12 2. TINJAUAN PUSTAKA
2.1 Habitat dan Penggunaannya
Menurut Odum (1993) habitat didefinisikan sebagai suatu tempat dimana organisme tinggal atau biasa ditemukan orang. Habitat terdiri dari komponen abiotik dan biotik yang bersama-sama menyusun kumpulan sumberdaya yang secara langsung maupun secara tak langsung mendukung kehidupan hewan untuk hidup di tempat tersebut. Tumbuh-tumbuhan merupakan bagian dominan dari habitat, dan juga berperan menyediakan berbagai macam makanan, tempat sarang serta tempat berlindung bagi hewan (Fleming 1992). Hutan primer, hutan sekunder dan semak merupakan habitat bagi burung, karena di semua tempat tersebut ditemukan berbagai jenis burung (Wiens 1992).
Tumbuhan yang terdapat di habitat tersebut merupakan faktor penting dalam kehidupan burung, karena beberapa bagian dari tumbuhan yaitu bagian generatif dan bagian vegetatif menjadi sumber makanan. Beberapa burung yang hidup di hutan memakan langsung material tumbuhan, seperti buah-buahan dan bunga (Fleming 1992). Buah yang dimakan disebar bijinya bersama feses, dan 50-80% tumbuhan hutan tropik dilakukan penyebaran bijinya oleh burung (Karr et al. (1992).
Penggunaan habitat oleh burung berubah-ubah tergantung ketersediaan sumberdaya yang dibutuhkannya. Perubahan penggunaan struktur vertikal tumbuhan untuk aktivitas makan burung sangat dipengaruhi oleh penyebaran makanan di pohon tersebut. Hasil penelitian Nurwatha (1994) menunjukkan Burung cabai jawa (Dicaeum trochileum), Cinenen kelabu (Orthotomus ruficeps) dan Burung-madu sriganti (Nectarinia jugularis) menggunakan lapisan tajuk yang berbeda pada habitat taman kota yang berbeda. Hal ini terkait dengan ketersediaan makanan pada ketinggian tumbuhan yang berbeda.
Komposisi komunitas dan kebiasaan hidup burung dapat dipengaruhi oleh perubahan komposisi spesies tumbuhan dalam suatu habitat (Lambert 1992). Sebagai contoh, perubahan habitat di hutan dataran rendah menjadi areal terbuka dan semak belukar, mengakibatkan beberapa spesies burung mengubah strata tempat mencari
13 makan dan memperluas daerah jelajahnya. Burung tidak memanfaatkan seluruh habitatnya, melainkan ada seleksi terhadap beberapa bagian dari habitat sesuai dengan yang dibutuhannya (Wiens 1992). Pengaruh keterbatasan sumberdaya di habitat untuk burung dapat menyebabkan persaingan baik intra-spesies atau inter-spesies (Karr et al. 1992).
2.2 Komunitas Burung
Komunitas burung berdasarkan terminologi adalah suatu kumpulan populasi dari spesies-spesies burung yang hidup di suatu habitat serta saling berinteraksi, membentuk sistem komposisi, struktur, perkembangan dan peranannya sendiri (Wiens 1992). Luasnya batasan yang melingkupi, menjadikan suatu komunitas sangat komplek, sehingga dalam memperlajarinya sering dilakukan pembagian-pembagian kajian. Morin (1999) menyatakan bahwa parameter penting dalam mempelajari suatu komunitas adalah taxocene dan guild.
Menurut Kaspari (2001) taxocene adalah pengelompokan secara ekologi berdasarkan kelompok taksa tertentu. Penentuan suatu komunitas berdasarkan taxocene terbatas pada organisme yang secara taksonomi relatif sama dan mendominasi komunitas tersebut, seperti komunitas burung. Taxocene merupakan unit dasar dalam penelitian makroekologi dan mempunyai parameter seperti kelimpahan dan keanekaan. Guild merupakan kumpulan spesies yang memanfaatkan suatu sumber daya dengan cara yang sama. Wiens (1992) menyatakan bahwa elemen kunci dari definisi guild adalah spesiesnya syntopic, kesamaan di antara spesies lebih ditentukan oleh kesamaan mereka dalam memanfaatkan suatu sumber daya dibandingkan secara taksonomi. Konsep guild diperkenalkan untuk mengklarifikasi beberapa kekeliruan sehubungan dengan konsep relung (niche), yang sebelumnya lebih banyak ditekankan pada seperangkat kondisi yang memungkinkan suatu spesies untuk tetap eksis dalam lingkungannya (Wiens 1992). Oleh karena itu, beberapa peneliti membatasi komunitas burung dengan batasan taksonomi dan guild yang berbeda, sehingga menjadi beberapa kelompok kecil dari burung seperti komunitas
14 burung air, paserin kecil, pemangsa, pemakan nektar, dan pemakan buah (Wiens 1992).
Komposisi spesies dari komunitas burung lokal ditentukan oleh penambahan spesies melalui pembentukan kolonisasi baru dan kehilangan spesies melalui kepunahan lokal. Perubahan tersebut terjadi dalam skala ruang dan waktu (Wiens 1992). Hal tersebut terkait dengan adanya perubahan habitat (Balen 1999). Habitat didominasi vegetasi semak, komposisi spesies burung yang menempatinya lebih banyak dari familia Sylviidae. Akan tetapi, habitat telah banyak ditumbuhi vegetasi pancang dan pohon komposisi spesies burung yang menempati bertambah dari familia Cuculidae, Picidae dan Capitonidae (Hadiprayitno 1999).
2.2.1 Keanekaan burung
Keanekaan spesies berhubungan dengan kekayaan (jumlah) spesies dalam suatu komunitas dan jumlah individu masing-masing spesies dalam komunitas tersebut (Krebs 1989; Wiens 1992). Keanekaan spesies adalah suatu karakteristik tingkatan komunitas berdasarkan organisasi biologisnya yang dapat digunakan untuk menyatakan struktur komunitas. Komponen utama keanekaan spesies adalah kekayaan jenis dan equitabilitas dalam pembagian individu yang merata diantara jenis (Odum 1993). Keanekaan spesies cenderung lebih rendah dalam ekosistem yang homogen dan lebih tinggi dalam ekosistem yang alami dan kompleks. Peningkatan jumlah spesies burung juga berkaitan dengan pertambahan luas habitat (Wiens 1992). Struktur komunitas dan kekayaan spesies burung berbeda antara suatu habitat dengan habitat yang lainnya (Johnsing & Joshua 1994). Keanekaan spesies di suatu habitat ditentukan oleh faktor seperti struktur vegetasi, komposisi spesies tumbuhan, sejarah habitat, tingkat gangguan dari predator dan manusia (Welty & Baptista 1988) serta ukuran luas habitat (Wiens 1992). Oleh karena itu, kondisi suksesi vegetasi berkaitan erat dengan perubahan komposisi spesies yang menempatinya (Alikodra 1990).
Penelitian mengenai hubungan keanekaan spesies burung dengan tahapan suksesi telah dilakukan di beberapa tempat. Welty & Baptista (1988) mendapatkan
15 spesies burung yang dominan berbeda di tiap tahapan suksesi proses reklamasi suatu lahan basah. Tiga tahun setelah reklamasi, spesies burung yang dominan adalah
Anthus pratensis. Emberiza schoeniculus menjadi burung yang dominan pada tempat
tersebut setelah lahan menjadi bentangan lumpur yang lembek. Pada bentangan lumpur yang keras (19 tahun setelah reklamasi), spesies burung yang dominan adalah
Montacilla flava. Selanjutnya ketika lahan tersebut telah berubah menjadi padang
rumput, spesies burung yang dominan adalah Alanda arvensis.
Hal serupa ditemukan Hadiprayitno (1999) di Gunung Tangkuban Parahu Jawa Barat, di habitat pinus yang berbeda usia. Di hutan pinus usia kurang dari 5 tahun ditemukan 6 spesies burung di dominasi oleh Cica-koreng jawa (Megalurus
palustris); hutan pinus usia 6-10 tahun ditemukan 7 spesies burung di dominasi
Kacamata biasa (Zosterops palpebrosus); hutan pinus berusia 11-15 tahun ditemukan 13 spesies burung di dominasi Kacamata biasa (Zosterops palpebrosus) dan Bentet kelabu (Lanius schach); dan hutan pinus berusia >15 tahun ditemukan 21 spesies burung didominasi Kacamata biasa (Zosterops palpebrosus) dan Gelatik-batu kelabu (Parus major).
2.2.2 Kelimpahan dan Distribusi Burung
Kelimpahan spesies burung dapat dinyatakan dengan jumlah individu suatu spesies di suatu habitat tertentu dalam waktu tertentu (Wiens 1992). Pada beberapa penelitian sering dinyatakan dengan kelimpahan relatif, yaitu jumlah total individu atau biomas suatu spesies dibandingkan jumlah total individu atau biomas seluruh spesies pada areal yang diamati (Morin 1999).
Menurut Wiens (1992) ada tiga model kelimpahan spesies yang bisa dijumpai dalam suatu komunitas yaitu: 1) jika secara numerik sebagian kecil spesies dalam komunitas mendominasi cukup besar, maka kelimpahan cenderung tidak seimbang. Model ini cenderung terjadi pada komunitas yang hanya memiliki sedikit spesies. Spesies dominan menguasai ruang tertentu dalam komunitas, sementara spesies lainnya menguasai bagian yang terpisah. Model ini dikenal juga dengan istilah
16 diantara mereka tanpa ada tumpang tindih. Jika kelimpahan sesuai dengan ukuran relung, distribusi dari kelimpahan spesies cenderung seimbang dengan hanya sedikit dominan secara numerik oleh sebagian kecil spesies. Model ini sesuai dengan yang dipopulerkan oleh MacArthur yaitu Broken Stick Model, 3) distribusi dan kelimpahan burung sesuai dengan distribusi log normal, terutama jika komunitas disusun oleh banyak spesies. Jika kelimpahan relatif dari spesies dibentuk oleh banyak faktor bebas yang saling berperan, faktor tersebut akan berlipat sehingga membentuk distribusi log normal
Menurut Karr et al. (1992) kelimpahan dan distribusi spesies burung di habitatnya dipengaruhi oleh kondisi struktur vegetasi. Ketersediaan stratifikasi vertikal vegetasi dapat memberikan pengaruh yang besar terhadap keberadaan dan kepadatan spesies burung. Oleh karena itu, kerusakan struktur maupun komposisi vegetasi hutan akibat kebakaran mempengaruhi distribusi dan kelimpahan burung karena terjadi perubahan struktur dan komposisi vegetasi (Ding et al. 1997; Hadiprayitno 1999). Selain itu, distribusi spesies burung juga dipengaruhi oleh fragmentasi habitat dan ketersediaan sumberdaya di habitat seperti makanan (Hobson & Bayne 2000, Haslem & Bennett 2008). Menurut Fleming (1992) kelimpahan buah yang tinggi berhubungan erat dengan kepadatan burung pemakan buah.
2.3 Karakteristik Burung Pemakan Buah 2.3.1 Morfologi Burung Pemakan Buah
Spesies-spesies burung berdasarkan jenis makanan yang dimakannya dapat dibagi 7 kategori (MacKinnon 1995), yaitu frugivora (pemakan buah), granivora (pemakan biji), insektivora (pemakan serangga), karnivora (pemakan daging dan bangkai), nektarivora (pemakan nektar), omnivora (pemakan segala misalnya buah dan serangga), dan piscivora (pemakan ikan). Kelompok spesies burung berdasarkan makanan tersebut mempunyai karakteristik yang berbeda satu dengan yang lainnya (Jordano 1992, 2000).
Berat tubuh burung pemakan buah merupakan faktor utama yang menentukan intensitas burung memakan buah. Kebutuhan jumlah makanan buah berhubungan erat
17 dengan besar tubuh burung pemakan buah (Herrera 1984a). Burung seperti
Acrocephalus spp. memakan buah yang ukuran sedang dengan komposisi volume
makanannya antara 30-70%. Ukuran tubuh yang kecil memakan buah berukuran kecil pula seperti Sylvia sp. dan Erithacus sp. (Jordano 1992, 2000).
Ukuran tubuh burung pemakan buah mempengaruhi intensitas memakan buah dengan cara membatasi jumlah maksimum dari buah-buahan yang ditelan dan daging buah yang dicerna (Herrera 1985). Sebagai contoh, rata-rata jumlah buah Prunus
mahaleb yang dimakan tiap kunjungan makan adalah 1,5 buah/kunjungan untuk Phoenicurus ochrusus (16,0 g); 9,0 buah/kunjungan untuk Turdus vircivorus (107,5
g), dan 21,0 buah/kunjungan untuk Columba palumbus (460,0 g) (Jordano & Schupp 2000). Oleh karena itu, berat tubuh sangat menentukan banyaknya jumlah maksimum biji yang dapat disebar oleh burung pemakan buah setelah makan (Jordano 1992, 2000).
Perbedaan cara mengambil buah oleh burung pemakan buah menunjukkan hubungan yang sangat erat dengan ekomorfologi dari burung tersebut, khususnya dengan morfologi sayap, karakteristik paruh dan morfologi alat gerak (Jordano 1986). Karakteristik bentuk dan ukuran paruh burung pemakan buah mempunyai peranan terhadap kerusakan biji dan keberhasilan penyebaran biji. Burung pemakan buah yang mempunyai ukuran paruh kecil dan kokoh seperti Emberiza spp. cenderung hanya dapat memakan daging buah, sedangkan bijinya dimuntahkan (Jordano 1992).
Besar bukaan paruh menunjukkan hubungan yang erat dengan ukuran buah yang dimakan; semakin besar bukaan paruh semakin besar pula ukuran buah yang dapat dimakan (Wiens 1992; Fukui 1995). Burung yang memiliki ukuran bukaan paruh kecil hanya memakan buah-buahan yang kecil, karena keterbatasan ukuran bukaan paruhnya (Wheelwright 1988; Herrera 1985).
2.3.2 Morfologi Sistem Pencernaan Burung Pemakan Buah
Burung mempunyai sistem pencernaan yang sangat berbeda dibanding mamalia, seperti tidak mempunyai gigi dan langit-langit mulut yang lunak, sehingga waktu ingesti di mulut lebih cepat. Secara umum sistem pencernaan burung terdiri
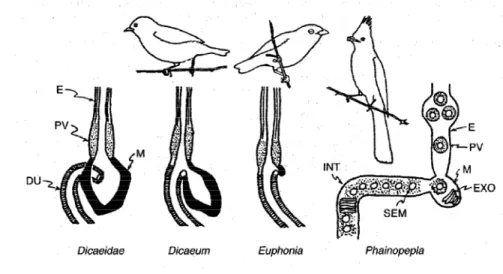
18 dari bagian rongga mulut, oesophagus, proventiculus, ventriculus, usus halus, usus besar dan kloaka (Pettingil 1970; Proctor & Lynch 1993). Pada beberapa burung yang mengalami spesialisasi berdasarkan makanan, beberapa bagian sistem pencernaannya mengalami modifikasi. Bagian sistem pencernaan yang termodifikasi khususnya pada burung pemakan buah adalah oesophagus. Oesophagus tidak dapat melebar pada kelompok burung Dicaeidae, sedangkan pada burung Ploceidae dan Pycnonotidae dapat melebar (Gambar 3). Modifikasi juga terjadi pada bagian proventiculus dan
ventriculus. Pada kedua bagian tersebut ototnya semakin tipis sehingga dapat
melewatkan biji secara utuh ke usus halus, usus besar dan kloaka (Jordano 1986).
Gambar 2. Skema sistem pencernaan burung (Proctor & Lynch 1993;hal 181)
Ciri umum modifikasi sistem pencernaan burung pemakan buah meliputi: (i)
oesopagus tereduksi dan proventrikulus yang sederhana, (ii) terdapat dinding
ventrikulus yang tipis, atau tidak mempunyai otot ventrikulus (rempela), (iii) posisi lateral dari ventrikulus dan hampir langsung dari oesophagus ke usus halus, serta (iv) panjang usus halus yang relatif pendek (Jordano 1992, 2000).
19 Gambar 3. Skema beberapa tipe proventrikulus dan ventrikulus pada burung pemakan buah. Sumber: Jordano (2000;hal 143).
E=oesophagus, PV=proventiculus, M=otot ventriculus, DU=duodenum, EXO=exocarp biji, SEM=biji, INT=usus halus
Burung-burung yang terspesialisasi sebagai pemakan serangga, memiliki otot ventrikulus lebih tebal dibandingkan pemakan buah (Gambar 3). Burung pemakan buah memiliki mekanisme proses pencernaan yaitu menghancurkan kulit buah dilakukan di ventrikulus yang sederhana dan bijinya dilewatkan melalui usus halus (Jordano 1992, 2000). Beberapa biji ditemukan hancur di feses burung, disebabkan pada saat penanganan buah tersebut di paruhnya.
2.4 Ketersediaan Buah
Buah merupakan bagian tumbuhan yang banyak mengandung nutrisi, sehingga burung lebih menyukai buah dibandingkan bagian lain dari tumbuhan. Selain itu, buah lebih mudah dipetik, ditangani dan ditelan oleh burung pemakan buah. Karakteristik buah yang diduga sebagai alasan dipilih oleh hewan pemakannya adalah penampilan buah (ukuran buah, berat daging buah terhadap berat buah, jumlah dan ukuran biji), kandungan nutrisi (karbohidrat, lipid, protein dan mineral), dan metabolit sekunder (Jordano 1992, 2000). Secara keseluruhan, ciri tadi memberi keuntungan kepada pemakannya yaitu total daging buah yang dapat dimakan dan kandungan nutrisi yang dapat diserap pada proses digesti (Herrera & Jordano 1981).
20 Ketersediaan buah di alam untuk burung dapat dilihat dari aspek kualitatif mencakup fenologi pembungaan dan buah, serta karakteristik buah berupa bentuk dan warna, aspek kuantitatif mencakup kelimpahannya (Radis 1997).
2.4.1 Fenologi Pembungaan dan Buah
Pola-pola fenologi tumbuhan buah di daerah tropik bervariasi dan kompleks. Komunitas tumbuhan buah mempunyai fase berbuah secara musiman. Spesies tumbuhan buah di daerah subtropik mengalami pembungaan dan menghasilkan buah pada musim semi, ketika suhu lingkungan meningkat, dan menghasilkan buah matang pada musim dingin. Oleh karena itu, ketersediaan buah maksimum di daerah subtropik cenderung terjadi di musim dingin (November-Januari), bertepatan dengan migrasi burung pemakan buah dari Palaearctic (Corlett 1998a; Noma & Yumoto 1997).
Fenologi pembungaan dan buah menunjukkan perbedaan di daerah tropik Asia dengan di subtropik. Fenologi pembungaan cenderung terjadi di musim kemarau dan buah matang pada musim hujan (Kimura et al. 2001). Kelimpahan buah tersedia secara maksimum tampak kurang mencolok di daerah tropik (Borges 1993; Corlett 1998b), tetapi beberapa spesies tumbuhan tertentu tampak sangat mencolok ketersediaannya antara musim kemarau dan musim hujan, seperti buah puspa dan kayu putih (Partasasmita 1998).
Fenologi tumbuhan tidak secara teratur menyesuaikan dengan musim panas dan musim hujan (Kimura et al. 2001). Walaupun seluruh studi mendeteksi siklus tahunan di tingkat komunitas, tetapi mempunyai hubungan yang lemah di tingkat populasi (Corlett & LaFrankie 1998; Corlett 1998b). Spesies tumbuhan buah tidak seluruhnya mempunyai fenologi berbuah supra-annual yaitu setiap individu tumbuhan buah mengalami periode berbuah yang terus-menerus sepanjang tahun. Akan tetapi, siklus tahunan atau dua tahunan tumbuhan berbuah juga tidak umum. Tumbuhan berbuah kadang-kadang terjadi beberapa kali dalam satu tahun, terutama spesies tumbuhan semak dengan periode berbuah yang hampir sama dari setiap individunya (Corlett & LaFrankie 1998; Corlett 1998b).
21 Pergantian musim berpengaruh terhadap penurunan jumlah buah masak di hutan subtropik dan hutan tropik pada beberapa tumbuhan. Sebagian besar pengaruh pergantian musim terjadi pada lamanya periode fase perkembangan buah, dan proses pematangan buah. Proses pematangan buah selalu lebih dari 1,5 bulan di hutan tropik, sedangkan di hutan subtropik lebih dari 4 bulan (Herrera 1984a).
2.4.2 Kelimpahan Buah
Kelimpahan buah sangat bervariasi pada ruang dan waktu. Distribusi horizontal dari tumbuhan buah berhubungan dengan kekayaan spesies tumbuhan dalam komunitas, sehingga menentukan pola distribusi sparsial buah di habitat. Jika tingkat suksesi dari vegetasi berbeda, maka kelimpahan buah untuk pemakan buah beda pula (Herrera 1985; Jordano 1992, 2000). Tumbuhan buah di hutan subtropik yang paling banyak adalah tumbuhan semak pada saat suksesi, tetapi tumbuhnya sangat sensitif terhadap naungan. Tumbuhan semak tersebut terkonsentrasi di daerah terbuka dan pinggiran hutan serta menjadi jarang di bagian dalam hutan (Herrera 1985).
Kelimpahan buah sangat dipengaruhi oleh musim dan keadaan lingkungan seperti curah hujan. Pembungaan cenderung menghasilkan buah lebih sedikit ketika menjelang musim hujan di daerah tropik. Hal ini karena curah hujan yang tinggi menghambat proses pembungaan, perkembangan buah dan pematangan buah (Kimura et al. 2001). Variasi pencahayaan dan kelembaban lingkungan berpengaruh secara langsung terhadap variasi fenologi di tingkat komunitas (Jordano 1992, 2000).
Kepadatan buah terjadi selalu di bawah 105 buah/ha (10 kg berat kering/ha) di hutan subropik. Tumbuhan semak di dataran rendah Mediterania memiliki jumlah kepadatan buah hampir sama dengan di beberapa hutan tropik, yaitu 80 kg berat kering/ha, dan kepadatan buah mencapai lebih dari 1,4 x 106 buah/ha/tahun (Herrera 1984a; Jordano 1995). Hutan hujan tropik menghasilkan banyaknya buah yang bervariasi. Umumnya kepadatan buah antara 180 - 1000 kg berat kering/ha, sedangkan semak di pegunungan berkisar antara 1- 8 kg berat kering/ha (Blake et al. 1990; Jordano 1992, 2000).
22 2.4.3 Karakteristik Buah
2.4.3.1 Warna Buah
Sebagian besar burung pemakan buah memakan buah yang hampir matang atau matang (Corlett 1998a, 1998b). Akan tetapi, beberapa burung paruh bengkok cenderung memakan buah yang masih muda seperti burung Nymphicus holandricus (Jones 1987) dan Psittacula alexandri (Partasasmita 1998). Buah berdaging di daerah subtropik umumnya mempunyai warna matang hitam atau merah (Corlett 1996). Pada umumnya ketersediaan buah matang berwarna coklat, kuning dan hijau lebih rendah di suatu habitat. Akan tetapi buah-buahan yang dimakan burung memiliki warna yang lebih cerah dibandingkan warna buah-buahan yang dimakan hewan mamalia (Leighton & Leighton 1983). Suryadi (1994) menemukan warna makanan burung rangkong lebih didominasi warna buah merah dan ungu.
Beberapa burung mempunyai mata yang bersel tetrachromatik dan dapat membedakan warna permukaan benda dalam kisaran ultraviolet (300-400 nm) dari spektrum (Corlett 1998b; Schmidt 2002). Sedangkan tipe sel trichromatik dimiliki mamalia terbatas pada primata, meliputi seluruh monyet dan kera (Osorio et al. 2004; Corlett 1998b). Seluruh mamalia herbivora lainnya mempunyai mata yang bersel
dichromat atau malam hari tidak bisa membedakan pola warna dengan jelas.
Perubahan pada primata dari memiliki tipe sel dichromatik menjadi trichromatik merupakan hasil evolusi sebagai bentuk adaptasi terutama bagi primata pemakan buah. Hasil perubahan tersebut mempermudah pemakan buah mendeteksi keberadaan buah-buahan yang berada diantara daun-daunan (Corlett 1998b; Schmidt 2002). 2.4.3.2 Ukuran Buah dan Biji
Berat buah dan biji lebih bervariasi di daerah garis katulistiwa seperti Singapura (Corlett 1998b) dan pinggiran daerah tropika seperti di Hongkong (Corlett 1996). Buah-buahan terkecil di daerah tersebut mempunyai berat segar ± 5 mg, terbesar ± 1 kg dengan berat biji ± 0,02 mg (Melastomataceae dan Rubiaceae), dan 5-10 g (Anacardiaceae, Burseraceae, Lauraceae, Myristicaceae dan Palmae) (Corlett
23 1998b). Berat buah beringin yang dimakan rangkong di pulau Sulawesi berkisar antara 0,08 – 15,3 g dengan diameter buah 5,43 –30 mm (Suryadi 1994).
Ukuran buah dan biji berinteraksi dengan karakteristik hewan penyebarnya yang potensial. Buah berukuran besar banyak tersedia di habitat, tetapi burung kesulitan untuk memakannya jika buah tersebut harus ditelan seluruhnya (Leighton & Leighton 1983). Buah yang berdiameter kecil (<8 mm) dapat dimakan oleh seluruh vertebrata pemakan buah. Akan tetapi hewan-hewan besar tidak menyukainya walaupun kadang-kadang memakannya, jika kepadatan buah tinggi atau satu pengambilan dapat diperoleh jumlah buah yang banyak (Corlett 1998b). Ukuran diameter buah (8-13 mm) berpotensi sebagai makanan untuk seluruh burung pemakan buah, tetapi hanya beberapa spesies burung yang memakannya seperti burung Zosteropidae dan Dicaeidae (Corlett 1998b). Ukuran diameter buah 22 mm dapat ditelan oleh beberapa spesies burung tertentu saja, seperti burung Enggang, Merpati buah, Kuau besar, Anis, Jalak, Bentet, dan Gagak (Leighton & Leighton 1983; Corlett 1998b; Ueda & Arima 2005). Diameter buah lebih dari 30 mm mungkin diluar kemampuan seluruh burung untuk menelannya. Namun ukuran buah seperti itu masih dapat dimakan oleh kebanyakan mamalia pemakan buah (Corlett 1998b).
Sebagai contoh, ukuran diameter buah Ficus drupacea adalah 20 mm dan hanya dimakan oleh mamalia pemakan buah yang lebih besar (Leighton & Leighton 1983). Akan tetapi, di Thailand burung yang sering memakan buah Ficus drupacea adalah Cabai (Dicaeum tangkas) dengan cara dipatuk sebagian-sebagian, dan hanya Rangkong yang menelan buah secara keseluruhan (Corlett 1998b).
Ukuran biji buah sangat berpengaruh terhadap kisaran buah yang dimakan oleh vertebrata. Ukuran buah di atas ambang, bijinya secara teratur dijatuhkan, diludahkan atau dimuntahkan tanpa melewati lambung. Ukuran biji buah yang dimuntahkan oleh kelompok monyet adalah ± 3-5 mm yaitu (Corlett & Lucas 1990; Corlett 1998b), walaupun hewan-hewan lebih kecil banyak menelan dan membuangnya melalui feses (Corlett & Lucas 1990). Burung dapat memakan buah dalam kisaran ukuran buah yang lebar, kecuali buah yang berukuran terlalu besar untuk ditelan dan terlalu keras untuk di patuk (Corlett 1998b).
24 2.4.3.3 Nutrisi Buah
Menurut Corlett (1996), komponen utama karakteristik 153 spesies buah (30% tumbuhan buah berdaging) didominasi buah-buahan berbiji tunggal di Hongkong. Buah-buahan tersebut mempunyai lapisan daging buah yang banyak mengandung air, lemak, karbohidrat dan berbiji banyak. Dari 58 spesies tumbuhan yang banyak mengandung karbohidrat menunjukkan bahwa burung pemakan buah memakan buah yang mengandung banyak hexosa, sementara mamalia memakan buah yang mengandung banyak hexosa dan sukrosa di Hongkong (Ko et al. 1998; Corlett 1998b). Menurut Cipollini (2000) metabolit sekunder membantu lebih dari satu fungsi adaptasi buah berdaging untuk dipilih sebagai pakan oleh burung.
Buah Ficus spp. termasuk jenis pakan yang sering dimakan oleh burung.
Walaupun buah tersebut memiliki kandungan serat yang tinggi dan nilai nutrisi yang rendah (Corlett 1998b). Di Hongkong, 8 spesies tumbuhan buah memiliki kisaran kandungan nutrisi dalam daging buahnya yang hampir sama, yaitu 45-71% total larutan karbohidrat, 9-25% serat, 2-11% protein dan 1-6% lemak (Corlett 1996). Beberapa burung pemakan buah sangat tergantung kebutuhan hidupnya pada ketersediaan buah Ficus spp. seperti Merpati hijau dan Rangkong. Kandungan nutrisi buah Ficus spp diduga telah mencukupi kebutuhan burung-burung tadi (Kinnaird 1992).
2.5 Perilaku Makan
Berdasarkan terminologi, perilaku makan terdiri dari serangkaian aktivitas makan yang dimulai dari mencari makanan, menangani sampai dengan memakannya (menelan). Hewan termasuk burung dalam menangani makanannya dilakukan dengan berbagai cara. Sebagai contoh, makanan diambil kemudian ditelan langsung atau diputar-putar bahkan dibanting-banting terlebih dahulu sebelum dimakan, jika makanan tersebut masih hidup seperti ikan, katak dan ular (Welty & Baptista 1988; Huntingford 1984).
Burung pemakan buah juga bervariasi dalam cara penanganan makanannya. Sebagai contoh, Rangkong akan mengapit buah di antara paruhnya dalam waktu 2-4
25 detik, kemudian dilemparkan ke atas, lalu ditangkap oleh paruhnya dan langsung ditelan (Suryadi 1994). Betet jawa menangani buah dengan cara dipegang oleh kakinya, kemudian buah digaruk dan disobek kulit buahnya serta dimakan sebagian-sebagian daging buahnya jika ukuran buah besar (Partasasmita 1998). Buah yang ditelan secara keseluruhan oleh burung berpotensi disebarkan bijinya, sedangkan buah yang dimakan sebagian-sebagian cenderung kurang berpotensi disebar bijinya (Wheelwright 1991).
2.5.1 Waktu Aktivitas Makan
Aktivitas makan hewan diurnal dimulai pada pagi hari sampai sore hari menjelang tidur. Burung melakukan aktivitas terbang pada umumnya bersifat bimodal, pagi hari meningkat kemudian pada siang hari menurun dan meningkat kembali pada sore hari. Fluktuasi tersebut ada hubungannya dengan kebutuhan makanan oleh burung (Marsden 1995), sedang tinggi rendahnya frekuensi aktivitas makan diduga dipengaruhi oleh suhu lingkungan dan perubahan intensitas cahaya matahari (Smith 1990). Pada pagi hari hewan berusaha untuk makan sebanyak-banyaknya dan energi yang diperoleh pada pagi hari dipergunakan untuk aktivitas siang dan sore hari. Aktivitas makan meningkat pada sore hari diduga sebagai strategi untuk tetap mempunyai energi pada malam hari (Marsden 1995). Pemilihan waktu aktivitas makan dipengaruhi juga oleh ketersediaan makanan, kesesuaian tempat, ada tidaknya pesaing dan predator (Krebs & Davis 1978).
Burung pemakan buah memiliki cara-cara tertentu dalam melakukan aktivitas makan, yaitu dengan mengambil buah sambil bertengger dekat dengan tandan buah, cara ini paling sering dipergunakan. Menurut Jordano (1992, 2000) burung pemakan buah mempunyai empat strategi manuver terbang yang berbeda untuk mengambil makanan dari tempat tenggerannya, yaitu pertama berhenti (hovering), metode ini digunakan oleh kelompok burung bondol (manakins), sikatan (flycatchers) dan Thraupidae; kedua memperlambat (stalling), dipakai oleh luntur (trogons); ketiga menyambar (swooping) dan memperlambat (stalling), memetik buah sambil terus bergerak dari satu tenggeran ke tenggeran yang lain, metoda ini digunakan oleh
26 kebanyakan familia Cotingidae; dan keempat mengambil buah dari tenggeran sambil berjalan memilih dan menggapai buah, kadang-kadang sambil menggantung (hanging) di dahan. Dua strategi pertama merupakan strategi yang paling umum digunakan burung pemakan buah.
2.5.2 Preferensi Makan
Setiap organisme untuk melangsungkan kehidupannya memerlukan makanan. Setiap makanan yang dimakan oleh hewan dapat ditinjau dari dua aspek yaitu aspek kualitatif dan kuantitatif. Aspek kuantitatif mencakup kelimpahannya di habitat dan aspek kualitatif meliputi ukuran, warna, palatabilitas, nilai gizi dan kemudahan dicerna (Krebs & Davis 1978).
Preferensi terhadap jenis pakan tertentu diduga dipengaruhi oleh warna, berat dan besar ukuran pakan, kelimpahan jenis pakan, dan kandungan nutrisinya. Pada burung yang berperan menentukan pola hidup dan jenis pakannya adalah ukuran tubuh, bentuk paruh dan sistem pencernaannya (Wiens 1992). Hubungan antara jenis makanan yang dikonsumsi oleh berbagai jenis burung dengan ketersediaannya di lingkungan dapat memperlihatkan fenomena pengalihan preferensi. Misalnya, apabila ketersediaan suatu jenis pakan di lingkungan rendah, maka penggunaan jenis pakan itu relatif rendah (tidak menampakkan preferensi), tetapi apabila ketersediaannya meningkat, maka hewan akan memperlihatkan preferensi yang tinggi terhadap jenis pakan tersebut (Smith 1990).
Seleksi pakan oleh hewan termasuk burung adalah merupakan strategi dalam mengoptimalkan perolehan energi untuk memenuhi kebutuhan hidupnya (Huntingford 1984; Krebs & Davis 1978). Semakin selektif pakan yang dimakan, maka waktu untuk mencari pakan tersebut akan semakin lama (Wheelwright 1991). Lamanya aktivitas makan burung akan menunjukkan kuantitas perolehan makanan. Semakin lama burung melakukan aktivitas makan di tumbuhan pakan, maka akan semakin banyak buah yang dimakan. Strategi tersebut sebagai bentuk mengoptimalkan perolehan sumberdaya pakan (Krebs & Davis 1978). Menurut Wheelwright (1991) burung pemakan buah harus tetap berada di tumbuhan pakan
27 hingga kenyang. Selain itu juga diduga bahwa lamanya kunjungan di tempat makan akan meningkatkan perolehan pakan oleh burung.
2.6 Interaksi Burung dan Tumbuhan Buah
Interaksi antara tumbuhan berbuah dan burung frugivora memainkan peranan penting dalam regenerasi tumbuhan dan ketersediaan makanan bagi hewan. Burung frugivora berkontribusi pada penyebaran biji tumbuhan dan memberi peluang pada biji tumbuhan untuk tumbuh di tempat yang jauh dari induknya (Herrera 2002; Kominami et al. 2003). Buah merupakan makanan esensial bagi burung frugivora dan hewan omnivora. Buah juga mendukung kehadiran populasi spesies burung pada musim panas di semak Mediterranean (Herrera 1984a). Kehilangan burung pemakan buah yang relatif lama akibat perburuan akan berdampak besar terhadap regenerasi tumbuhan yang sangat tergantung pada burung untuk penyebaran bijinya. Demikian pula sebaliknya jika habitat mengalami kerusakan vegetasinya akan berpengaruh terhadap ketersediaan buah, dan selanjutnya berdampak pada keanekaan dan kelimpahan burung yang sangat tergantung pada tumbuhan buah tersebut (Loiselle & Blake 2002).
2.6.1 Penyebaran biji
Banyak spesies tumbuhan dalam menyebarkan bijinya bergantung kepada burung pemakan buah di hutan tropis. Ketidakhadiran spesies burung karena pengaruh fragmentasi hutan, akan berpengaruh terhadap regenerasi spesies-spesies tumbuhan di tempat tersebut (Corlett 2002). Spesies burung yang membantu penyerbukan dan penyebaran biji beberapa jenis tumbuhan tertentu, antara lain Burung madu (Nectarinidae), burung-burung dari suku Anatidae, Columbidae, Turdidae, Corvidae dan Sittidae (Welty & Baptista 1988), Pycnonotidae (Fukui 1995). Delapan puluh lima persen dari hampir 200 spesies tumbuhan berbiji disebarkan bijinya oleh burung di hutan sekunder dan semak subtropik (Corlett, 1996), antara lain dari tumbuhan Lantana camara, Sapium discolor, Litsea sp., Ficus spp., dan Dendrophthoe spp. (Reid 1990).
28 Kepadatan burung frugivora dan pergerakannya sering dihubungkan secara langsung dengan penyebaran buah–buahan (Levey 1988). Pada saat ketersediaan buah menurun, burung pemakan buah Pipra mentalis berhenti berkembang biak, dan terjadi perubahan pola makan dengan lebih banyak memakan serangga sambil mencari tempat yang lebih banyak buahnya. Burung ini bergerak sepanjang hutan untuk mencari tempat dimana terdapat buah yang melimpah ( Levey et al. 1994).
Menurut hasil penelitian Herrera (1984b), burung passerine semak dapat dibagi menjadi tiga kategori sesuai dengan jenis makanan dan teknik penangan buah yaitu:
1. non-frugivores, spesies–spesies yang tidak pernah tercatat memakan buah-buah walaupun kelimpahan buah-buah tinggi.
2. fruit predators, spesies-spesies yang memakan daging buah, biji-bijian, atau keduanya tetapi tidak melakukan penyebaran biji.
3. seed dispersers, spesies-spesies yang memakan seluruh buah- buah berdaging dan memuntahkan bijinya kembali atau mengeluarkan bijinya bersama feses. Menurut Wheelwright (1991) buah yang memiliki biji besar ditemukan bijinya terjatuh dekat dengan tumbuhan induknya, karena tidak dicerna dan biji dimuntahkan lebih cepat oleh burung. Buah kecil dengan biji kecil jatuh jauh dari tumbuhan induk karena ditelan dan melalui proses digesti (pencernaan) burung sebelum jatuh bersama feses burung pemakan buah. Dua aspek penting yang mempengaruhi hasil penyebaran biji oleh burung adalah perilaku penanganan biji (baik ingesti maupun digesti) dan waktu retensi (Jordano 1992, 2000). Bentuk perilaku penanganan biji menentukan jumlah biji utuh yang dapat disebar.
Menurut Jordano (1995) burung penyebar biji sejati adalah burung yang menelan keseluruhan buah, memuntahkan atau mendefekasikan bijinya setelah meninggalkan tumbuhan induknya. Burung yang berperan sebagai penyebar biji adalah burung pemakan buah, tetapi tidak semua burung pemakan buah merupakan penyebar biji. Untuk membedakan jenis makanan yang dimakan oleh burung, dapat dilakukan dengan cara pengamatan langsung saat burung melakukan aktivitas makannya, analisis isi lambung, analisis muntahan, dan komposisi biji dalam feses
29 (Corlett 1998b). Analisis komposisi feses dimungkinkan untuk mengetahui jenis makanannya karena feses tersebut tersusun dari bagian-bagian makanan yang tidak tercerna dan bagian tersebut masih dapat diidentifikasi (Corlett 1996).
Kombinasi jenis biji dalam feses burung frugivora, bukan hasil proses acak dari buah yang tersedia di habitat, tetapi lebih mengindikasikan terdapat pemilihan yang konsisten oleh burung (Jordano 1992, 2000). Reid (1989) menemukan penyebaran tumbuhan parasit hanya terdapat di tumbuhan inang tertentu karena disebarkan oleh burung tertentu pula.
2.6.2 Daya Kecambah Biji
Berdasarkan terminology, biji secara struktural dapat dikatakan benih yaitu, bagian dari tanaman yang digunakan untuk memperbanyak atau mengembangbiakkan (Widajati et al. 2008). Mutu fisiologis dari biji dapat diukur dari viabilitasnya yaitu kemampuan hidup biji untuk tumbuh. Menurut Sadjad (1993) periode viabilitas biji yang sangat menentukan adalah pada proses pembentukan, perkembangan dan pemasakan fisiologis. Setelah biji masak secara fisiologis akan memasuki fase pengurangan kadar air dan biji menjadi dorman. Dormasi merupakan mekanisme yang mengatur perkecambahan biji, serta menjadi aspek penting dari spesies tumbuhan dan ekologinya. Perkecambahan terjadi ketika biji tumbuhan tumbuh, dan berkembang secara optimal. Dormansi adalah mencegah terjadinya perkecambahan ketika biji tersebar di tempat yang tidak cocok untuk tumbuh (Walck et al. 2002). Beberapa buah yang matang dari spesies tumbuhan memiliki biji yang terdapat tunas embrio di dalamnya, dengan ukuran lebih kecil dari bijinya. Embrio sangat kecil serta dapat dibedakan dari kotiledon dan bakal akar. Embrio tidak akan berkembang sebelum akar tumbuh memanjang. Embrio tidak berkembang karena dorman pada saat pematangan, ini berarti biji mempunyai dorman secara ekofisiologi, sehingga membutuhkan pemecah dorman seperti dengan air, panas atau dingin (Walck et al. 2002; Widajati et al. 2008). Menurut Widajati et al. (2008) dan Bewley & Black (1986) proses-proses yang terjadi di awal perkecambahan yaitu imbibisi. Kecepatan imbibisi sangat dipengaruhi oleh permeabilitas kulit biji, komposisi kimia biji, suhu
30 dan konsentrasi air. Setelah imbibisi, selanjutnya terjadi reaktivasi enzim, inisiasi pertumbuhan embrio dan retaknya kulit biji, serta kemudian munculnya akar menembus kulit biji (Bewley & Black 1986).
Spesies tumbuhan semak di hutan subtropik 87,5 % termasuk pada kelompok dorman pada saat pematangan. Kecepatan perkecambahan biji dari buah yang matang sebagian besar dihambat oleh keberadaan daging buah dan “exocarp” dari biji (Fukui 1995). Loiselle (1990) dan Jordano (2000) menunjukkan secara eksperimen bahwa kombinasi spesifik dari penyebaran biji dalam feses burung pemakan buah berpengaruh langsung terhadap perkecambahan biji dan kemampuan bertahan hidupnya. Demikian pula waktu retensi dari burung pemakan buah dapat meningkatkan daya kecambah biji (Barnea et al. 1990, 1991, 1992).