KANDUNGAN DAN HASIL ANTOSIANIN GALUR PADI BERAS MERAH PADA TIGA LINGKUNGAN TUMBUH BERBEDA
ANTHOCYANIN CONTENTS AND YIELDS OF RED RICE LINES AT THREE DIFFERENT GROWING ENVIRONMENTS
I Gusti Putu Muliarta Aryana
PS Pemuliaan Tanaman, Fakultas Pertanian Universitas Mataram ABSTRAK
Tujuan penelitian adalah untuk mengetahui kandungan dan hasil antosianin galur padi beras merah pada tiap genotipe dan responnya jika ditanam pada kondisi lingkungan tumbuh berbeda. Percobaan pada lingkungan gogo dilakukan pada lahan tegalan Desa Prian Kecamatan Montong Betok Kabupaten Lombok Timur, dengan jenis tanah Inseptisol. Percobaan pada lingkungan cekaman kekeringan dan sawah irigasi teknis dilakukan di Desa Grisak Kecamatan Ampenan Kota Mataram dengan jenis tanah Entisol. Rancangan percobaan yang digunakan di setiap lingkungan adalah Rancangan Acak Kelompok dengan 23 perlakuan yaitu 20 genotipe hasil seleksi back cross dan tiga tetua (Piong, Angka dan Kenya) yang diulang 3 kali. Kandungan antosianin dihitung dengan metode Shi at.al.(1984) dan hasil antosianin dihitung dengan mengalikan kandungan antosianin dengan hasil beras per ha. Analisis ragam gabungan berdasarkan Singh dan Chaudhary (1979). Hasil penelitian menunjukkan bahwa: a.) Setiap genotipe memiliki kandungan antosianin yang berbeda, namun memiliki respon yang sama terhadap lingkungan. Lingkungan kering memberikan kandungan antosianin tertinggi diikuti oleh lingkungan gogo dan lingkungan sawah irigasi. Kandungan antosianin tertinggi 42,19 ppm dicapai oleh galur G3(A2) diikuti oleh galur G4(A3) sebesar 39,23 ppm. b.) Hasil antosianin beras setiap genotipe berbeda di setiap lingkungan tumbuh, dengan nilai tertinggi di lingkungan tercekam kekeringan terdapat pada galur G4(A3) (59,43 gram/ha), di lingkungan gogo tertinggi pada galur G4(A3) (99,00 gram/ha) dan di lingkungan sawah irigasi teknis tertinggi pada galur G22(Angka) (96,88 gram/ha).
ABSTRACT
The aim of this research was to know anthocyanin contents and yields in each genotype of red rice lines, and its responses to various conditions of growing environments. For dryland experiment, the experiment was conducted at Prian village of Montong Betok district (East Lombok), on an Inseptisol soil. For cultivation in drought area and in an irrigated paddy field, the experiments were conducted at Grisak village, Ampenan (Mataram), on an Entisol soil type, during the dry season. Experimental Design used in each location was Randomized Complete Block Design with 23 treatments : 20 genotypes obtained from back cross selection and three parents (Piong, Angka and Kenya) with 3 replicates. Combined analysis of variance was based on Singh and Chaudhary (1979). Results indicated that: a). Every genotype had different level of anthocyanin content but responded similarly to the environments. Dry environment resulted in the highest anthocyanin content, followed by upland and technical irrigation environments. The highest anthocyanin content was on G3(A2) (42.19 ppm), followed by G4(A3) (39.23 ppm). b). Anthocyanin yields for every genotype varied with environments, with the highest value in drought environment was on G4(A3) (59.43 g ha-1), in upland area the highest value was on G4(A3) (99.00 g ha -1), and in technical irrigation area the highest value was on G22(Angka) (96.88 g ha-1).
_________________________
Kata kunci : antosianin, beras merah, kandungan, hasil. Key words: anthocyanin, red rice, content, yield.
PENDAHULUAN
Padi beras merah (Oryza sativa L.) merupakan bahan pangan pokok yang bernilai kesehatan tinggi. Selain mengandung karbohidrat, lemak, protein, serat dan mineral, beras merah juga mengandung antosianin. Antosianin merupakan pigmen merah yang
terkandung pada perikarp dan tegmen (lapisan kulit) beras, atau dijumpai pula pada setiap bagian gabah (Drake, Gebardt, dan Matthews, 1989; Chang dan Bardenas, 1965). Antosianin adalah senyawa fenolik yang masuk kelompok flavonoid dan berfungsi sebagai antioksidan, berperan penting baik bagi tanaman itu sendiri
maupun bagi kesehatan manusia. Peran antioksidan bagi kesehatan manusia untuk mencegah penyakit hati (hepatitis), kangker usus, stroke, diabetes, sangat esensial bagi fungsi otak dan mengurangi pengaruh penuaan otak (Nirmala, 2001). Oleh karena itu produksi olahan beras merah dipasarkan sebagai makanan bayi lanjutan dan konsumsi obat – obatan dengan harga 4 – 6 kali lebih mahal daripada harga beras merah pada umumnya. Saat ini kebutuhan akan beras merah terus meningkat sejalan dengan kesadaran masyarakat tentang manfaat kesehatan (Muliarta et al., 2003)
Potensi hasil dan kandungan antosianin pada tanaman selain dipengaruhi oleh faktor genetik juga dipengaruhi oleh faktor lingkungan. Pada lingkungan tercekam kekeringan dan tanpa tercekam kekeringan (basah) akan menyebabkan perbedaan potensi hasil yang diperoleh maupun biosintesa antosianin. Dari hasil penelitian yang dilakukan oleh Nabhushana dan Reddy (2004), dimana perlakuan kekeringan pada perkecam-bahan padi beras merah menghasilkan peningkatan kecepatan yang nyata terhadap akumulasi antosianin di daun, ini terdapat pada hampir semua genotipe padi yang diperlakukan yaitu R27, G962, Nagita 22, Prasanna, kecuali galur Hamsa (sedikit warna). Warna yang nampak pada daun sangat dipengaruhi oleh perbedaan genetik tanaman tersebut. Genotipe homozigot resesif pada beberapa lokus atau heterozigot untuk allel yang menghambat warna dominan, akan menghasilkan perubahan warna fenotipe baik pada yang berwarna maupun yang kurang berwarna. Diantara yang diberi perlakuan pengaruh kekeringan nyata lebih efektif dalam memacu lintasan flavonoid dibandingkan dengan perlakuan tanpa cekaman. Sementara semua komponen genotipe menunjukan peningkatan akumulasi pingmen antosianin akibat stres kekeringan, tetapi bagi genotipe yang tidak berwana tidak terjadi. Dari informasi ini dapat disimpulkan bahwa akumulasi antosianin pada komponen genotipe padi diakibatkan karena stres kekeringan .
Di Indonesia perbaikan varietas padi beras merah belum mendapatkan perhatian yang memadai baik sebagai padi gogo maupun padi sawah. Varietas unggul yang sudah dilepas oleh Balai Besar Penelitian Padi Sukamandi hingga kini ada sekitar 190 varietas. Dari 190 varietas yang dilepas baru satu varietas padi beras merah yang dilepas yaitu Aek Sibundong sebagai padi sawah (Suprihatno et al., 2007). Memperhatikan potensi genetik padi beras merah dan nilai ekonomi yang tinggi, Muliarta et.al., 2006 telah mendapatkan 20 galur harapan padi beras hasil seleksi silang balik antara kultivar Angka dan
Piong sebagai tetua berulang dengan tetua kultivar Kenya toleran kekeringan sebagai tetua donor. Bagaimana respon galur harapan padi beras merah di atas terhadap kandungan dan hasil antosianin pada lingkungan tumbuh berbeda perlu diteliti.
Tujuan penelitian ini adalah untuk mengetahui kandungan dan hasil antosianin galur padi beras merah pada tiap genotipe dan responnya jika ditanam pada kondisi lingkungan tumbuh berbeda.
METODE PENELITIAN
Penelitian dilaksanakan pada tiga lingkungan tumbuh berbeda. Penelitian pada lingkungan gogo dilakukan pada lahan tegalan di Desa Prian Kecamatan Montong Betok Kabupaten Lombok Timur, dengan jenis tanah inseptisol. Waktu kegiatan MH Desember 2006 - Maret 2007. Sedangkan pada lingkungan tercekam kekeringan dan sawah irigasi teknis dilakukan di Desa Grisak Kecamatan Ampenan Kota Mataram Lombok dengan jenis tanah entisol, pada MK April - Agustus 2007. Rancangan percobaan yang di gunakan di setiap lingkungan adalah Rancangan Acak Kelompok dengan 23 perlakuan yaitu 20 genotipe hasil dari seleksi back cross dan tiga tetua (Piong, Angka dan Kenya) yang di ulang 3 kali. Penanam setiap perlakuan genotipe dengan luasan 5,5 m x 1,25 m , jarak tanam 25 cm x 25 cm, dengan satu tanaman per rumpun. Pengairan di lingkungan gogo tergantung pada air hujan, pengairan di lingkungan sawah sesuai budidaya padi sawah pengairan teknis dan pengairan di lingkungan tercekam kekeringan pada kondisi tercekam 25 -30 % air tersedia. Karakter kandungan antosianin beras dihitung dengan menggunakan metode Shi et.al.,(1992) dan hasil antosianin dihitung dengan mengalikan kandungan antosianin dengan hasil beras per ha. Data hasil pengamatan dianalisis dengan analisis ragam pada taraf nyata 5 %. Beda antar galur, lingkungan dan interaksinya diuji dengan uji DMRT pada taraf nyata 1 %.
HASIL DAN PEMBAHASAN
Pada Tabel 1 nampak bahwa analisis ragam gabungan terhadap pengaruh genotipe dan lingkungan menunjukkan perbedaan nyata terhadap kandungan antosianin beras, tetapi tidak menunjukkan adanya pengaruh nyata pada interaksi genotipe x lingkungan.
Tabel 1. Analisis ragam gabungan pengaruh lingkungan, genotipe, dan interaksi lingkungan x genotipe karakter kandungan dan hasil antosianin padi beras merah pada tiga lingkungan tumbuh
Kuadrat Tengah Sumber
Keragaman db Kandungan antosiani Hasil antosianin Lingkungan (L) 2 1,453** 51,12** Ulangan(Lingk.) 6 3,304** 2,11* Genotipe(G) 22 7,882** 30,80** L x G 44 0,030ns 0,95* Galat 132 0,092 0,45 Keterangan : * = Nyata pada taraf uji 5%; ** =
Sangat nyata pada taraf uji 1 %; ns = Tidak nyata
Dari Tabel 2 nampak bahwa kandungan antosianin beras merah tertinggi pada genotipe G3(A2) yaitu sebesar 42,19 ppm. Genotipe ini berasal dari hasil persilangan silang balik tetua Kenya dengan Angka. Jika dibandingkan dengan asal tetuanya yaitu Angka dengan kandungan antosianin yang dimiliki sebesar 34,66 ppm. Maka genotipe G3(A2) mengalami peningkatan kandungan antosianin sebesar 21,73%. Peningkatan kandungan antosianin ini juga di jumpai pada genotipe G4(A3) yaitu sebesar 39,23 ppm sebesar 21,61%.
Dari hasil perkawinan silang balik tetua Kenya dengan Piong ada tiga genotipe yang menghasilkan peningkatan kandungan antosianin secara nyata jika dibandingkan dengan tetua Piong yang memiliki kandungan antosianin sebesar 32,26 ppm. Genotipe tersebut adalah G18(P16), G17(P15) dan G13(P4) dengan kandungan antosianinnya secara berturut-turut 37,64; 36,26 dan 35,07 ppm dengan kenaikan kandungan antosianinnya sebebesar 16,68 %, 12,40 % dan 8,71 %. Untuk genotipe lainnya mengalami penurunan.
Dari pengaruh lingkungan tumbuh, nampak bahwa kandungan antosianin tertinggi diperoleh pada lingkungan tercekam kekeringan yaitu sebesar 32,07 ppm. Kandungan antosianin menurun dengan berkurangnya cekaman kekeringan. Pada lingkungan gogo rerata kandungan antosianin beras genotipe tanaman sebesar 28,47 ppm. Pada lingkungan tampa cekaman kekeringan yaitu pada lingkungan sawah irigasi teknis, rerata kandungan antosianinnya 28,25 ppm.
Tabel 2. Rerata kandungan antosianin (ppm) genotipe padi beras merah pada tiga lingkungan tumbuh
No Perlakuan Kandungan antosianin beras (ppm)
A Genotipe 1 G3(A2) 42,19 a 2 G4(A3) 39,23 ab 3 G18(P16). 37,64 bc 4 G17(P15). 36,28 cd 5 G13(P4) 35,07 cd 6 G6(A5). 34,50 de 7 G22(ANGKA) 34,66 de 8 G12(P3). 33,61 def 9 G21(PIONG) 32,26 efg 10 G11(P2). 32,19 efg 11 G8(A7). 31,36 fg 12 G19(P18). 30,52 gh 13 G9(A8). 28,78 g 14 G10(P1). 26,56 i 15 G7(A6). 25,50 ijk 16 G14(P5) 25,33 ijk 17 G15(P13). 25,74 ij 18 G1(A0) 23,89 jk 19 G16(P14). 23,26 k 20 G20 (P19). 19,03 l 21 G2(A1) 17,62 l 22 G5(A4). 14,50 m 23 G23(KENYA) 0,00 n B Lingkungan 1 Kering 32,07 a 2 Gogo 28,47 b 3 Sawah 24,18 c Rerata 28,25 Keterangan: Data 0,00 (pada analisis keragaman
ditrasformasi ke √(x+1)). Angka-angka pada setiap kolom yang diikuti oleh huruf yang sama tidak berbeda nyata pada uji DMRT 1% Dari hasil percobaan tentang pewarnaan antosianin pada padi yang dilakukan Reddy (1996) nampak bahwa pembentukan pigmen atosianin di sebabkan oleh tiga gen dasar yaitu C (chromogen), A (activator) dan P (distributor). Dasar genetik biosintesa antosianin pada padi adalah dimana gen dapat dikelompokkan dalam gen struktural dan gen pengatur. Sejumlah gen struktural adalah C, A, Rc (brown pericarp), Rd (brown pericarp) menyandi enzim-enzim, sedangkan gen pengatur (regulator) adalah P (purple) dan Prp (purple pericarp) dengan
bermacam-macam allel yang menggambarkan lemahnya pengaturan pewarnaan. Dengan demikian pola pewarnaan antosianin pada padi utamanya ditentukan oleh status alel individu gen dan interaksi yang kompak antar allel.
Dari hasil penelitian yang dilakukan oleh Sbe et al. (1997), bahwa genotipe padi Nigita 22 bersifat homozigot dominan pada alle I-ib penghambat warna daun dan meniadakan pigmen antosianin. Terdapat hal yang menarik pada Nigita 22 yaitu kemampuannya untuk mengakumulasi antosianin bila diberikan perlakuan stres kekeringan yang ditandai dengan nilai A530, hal ini menunjukkan adanya expresi induksi stres terhadap gen yang relevan, selain itu pengaturan lintasan flavonoid dilak-sanakan secara pintas pada siklus regulasi yang meliputi kerja gen I-ib.
Sebagai perbandingan dengan tanaman kedelai menunjukkan toleransi yang luar biasa terhadap kekeringan, dengan mengakumulasi tiga sampai empat kali lebih banyak antosianin selama proses dehidrasi dibandingkan di dalam kondisi suka air (Chalker,1999.).
Penghindaran kekeringan umumnya nampak pada tanaman-tanaman yang mampu menu-runkan potensial osmotik daun-daunnya sampai ke titik evapotranspirasi minimum. Konsentrasi antosianin akan secara de facto menurunkan potensial osmotik daun (yaitu membuat semakin negatif), yang mengurangi potensial air daun dan barangkali mendukung penurunan konduktansi stomata yang tampak pada daun Brachystegia, Photinia X Fraseri ‘Red Top’ (Chalkerl, 1999.).
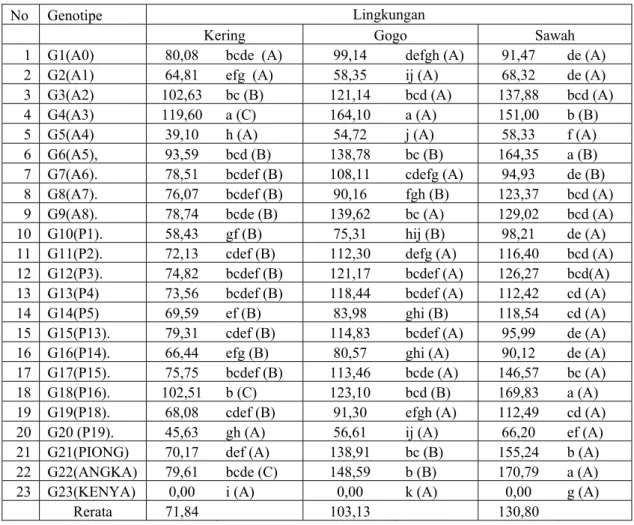
Tabel 3: Rerata hasil antosianin (gram/ha) pada genotipe padi di tiga lingkungan tumbuh
No Genotipe Lingkungan
Kering Gogo Sawah
1 G1(A0) 80,08 bcde (A) 99,14 defgh (A) 91,47 de (A) 2 G2(A1) 64,81 efg (A) 58,35 ij (A) 68,32 de (A) 3 G3(A2) 102,63 bc (B) 121,14 bcd (A) 137,88 bcd (A) 4 G4(A3) 119,60 a (C) 164,10 a (A) 151,00 b (B) 5 G5(A4) 39,10 h (A) 54,72 j (A) 58,33 f (A) 6 G6(A5), 93,59 bcd (B) 138,78 bc (B) 164,35 a (B) 7 G7(A6). 78,51 bcdef (B) 108,11 cdefg (A) 94,93 de (B) 8 G8(A7). 76,07 bcdef (B) 90,16 fgh (B) 123,37 bcd (A) 9 G9(A8). 78,74 bcde (B) 139,62 bc (A) 129,02 bcd (A) 10 G10(P1). 58,43 gf (B) 75,31 hij (B) 98,21 de (A) 11 G11(P2). 72,13 cdef (B) 112,30 defg (A) 116,40 bcd (A) 12 G12(P3). 74,82 bcdef (B) 121,17 bcdef (A) 126,27 bcd(A) 13 G13(P4) 73,56 bcdef (B) 118,44 bcdef (A) 112,42 cd (A) 14 G14(P5) 69,59 ef (B) 83,98 ghi (B) 118,54 cd (A) 15 G15(P13). 79,31 cdef (B) 114,83 bcdef (A) 95,99 de (A) 16 G16(P14). 66,44 efg (B) 80,57 ghi (A) 90,12 de (A) 17 G17(P15). 75,75 bcdef (B) 113,46 bcde (A) 146,57 bc (A) 18 G18(P16). 102,51 b (C) 123,10 bcd (B) 169,83 a (A) 19 G19(P18). 68,08 cdef (B) 91,30 efgh (A) 112,49 cd (A) 20 G20 (P19). 45,63 gh (A) 56,61 ij (A) 66,20 ef (A) 21 G21(PIONG) 70,17 def (A) 138,91 bc (B) 155,24 b (A) 22 G22(ANGKA) 79,61 bcde (C) 148,59 b (B) 170,79 a (A) 23 G23(KENYA) 0,00 i (A) 0,00 k (A) 0,00 g (A)
Rerata 71,84 103,13 130,80
Keterangan : Data 0,00 (pada analisis keragaman data ditransformasikan dalam akar x +1). Angka-angka dalam kolom yang di dampingi oleh huruf kecil yang sama tidak berbeda nyata; angka-angka dalam baris yang didampingi oleh huruf besar yang sama tidak berbeda nyata pada uji DMRT 1%
Hasil antosianin beras yang diperoleh per satuan luas merupakan nilai perkalian antara kandungan antosianin dengan hasil gabah. Hasil antosianin beras secara komulatif sangat dipengaruhi oleh tingkat produksi tanaman tersebut. Meskipun kandungan antosianin per satuan bobot sama, namun jika produktivitas yang dicapai berbeda, maka hasil antosianin kumulatif yang diperoleh juga berbeda. Semakin tinggi produktivitas tanaman, hasil antosianin komulatif juga semakin tinggi.
Hasil analisis ragam gabungan pengaruh interaksi genotipe x lingkungan menunjukkan perbedaan nyata terhadap hasil antosianin beras, artinya hasil antosianin suatu genotipe akan mengalami perubahan dengan berubahnya lingkungan tumbuh. Pada Tabel 3 tampak bahwa hasil antosianin tertinggi dilingkungan tercekam kekeringan di hasilkan oleh genotipe G4(A3) sebesar 119,60 gram/ha, kemudian diikuti oleh G3(A1) sebesar 102,63 gram/ha dan G18(P16) sebesar 102,51 gram/ha. Genotipe–genotipe tersebut bila ditanam di lingkungan gogo dan sawah irigasi teknis mengalami peningkatan hasil antosianin. Hasil antosianin tertinggi diperoleh bila ditanam di lingkungan sawah irigasi teknis, hal ini disebabkan karena hasil gabah tertinggi di peroleh pada lingkungan sawah irigasi teknis, kecuali G4(A3) dilingkungan irigasi teknis hasilnya lebih rendah dibandingkan dengan di lingkungan gogo. Genotipe G4(A1) merupakan genotipe yang menghasilkan antosianin tertinggi di lingkungan gogo yaitu sebesar 164,10 gram/ha. Pada lingkungan sawah irigasi teknis genotipe yang memberikan hasil antosianin tertinggi adalah G22(Angka) kemudian diikuti oleh G18(P16) dan G6(A5) dengan nilainya secara berurutan 170,79; 169,83 dan 164,35 gram/ha. Genotipe ini akan mengalami penurunan hasil antosianin jika ditanam di lingkungan gogo dan tercekam kekeringan. Hasil antosian terendah di hasilkan bila di tanam pada lingkungan tercekam kekeringan.
Bila dilihat hubungan antara hasil gabah, hasil antosiani dan kandungan antosianin dengan lingkungan tumbuh, nampak secara umum bahwa terjadi penurunan hasil antosianin dengan bertambahnya cekaman kekeringan, tetapi sebaliknya dengan kandungan antosianin secara relatif terjadi peningkatan, namun peningkatan kandungan antosianin karena bertambahnya cekaman kekeringan tidak sebanding dengan penurunan hasil antosianin.Terdapat indikasi bahwa genotipe hasil seleksi silang balik hingga empat kali memberikan hasil antosianin tertinggi bila dilakukan penanaman di lingkungan gogo.
KESIMPULAN Kesimpulan
a. Setiap genotipe memiliki kandungan antosianin yang berbeda, namun memiliki respon yang sama terhadap lingkungan. Lingkungan kering memberikan kandungan antosianin tertinggi diikuti oleh lingkungan gogo dan lingkungan sawah irigasi, kandungan antosianin tertinggi 42,19 ppm dicapai oleh G3(A2) diikuti oleh G4(A3) sebesar 39,23 ppm.
b. Hasil antosianin setiap genotipe berbeda di setiap lingkungan tumbuh, hasil antosianin beras tertinggi di lingkungan tercekam kekeringan terdapat pada G4(A3) yaitu 59,43 gram/ha. Di lingkungan gogo hasil antosianin tertinggi terdapat pada G4(A3) yaitu 99,00 gram/ha. Di lingkungan sawah irigasi teknis hasil antosianin terdapat pada G22(Angka), G18(P16) dan G6(A5) berturut-turut 96,88; 95,91 dan 90,06 gram/ha.
Saran
Genotipe-genotipe padi beras merah yang memiliki kandungan dan hasil antosianin beras tinggi perlu di lanjutkan ke uji multi lokasi .
UCAPATAN TERIMA KASIH
Terima kasih yang sedalam-dalamnya penulis sampaikan kepada Prof. Dr. Ir. Nur Basuki, Prof. Dr. Ir.Kuswanto, MS dan Prof. Dr. Ir. Astanto Kasno,MS. yang telah memberikan masukan dan saran sehingga tulisan ini terselesaikan.
DAFTAR PUSTAKA
Chalker S. 1999. Enviromental significant of anthosianins in plant stress responses,Photochem.Photobiol. 70: 1-9
Chang. T. T. and E. A. Bardenas. 1965. The morphology and varietals characteristics of the rice plant, Tech. Bull. IRRI 4 : 40 p. Drake, D. L., S. E. Gebardt, and R. H.
Matthews. 1989. Composition of foods; Cereal Grains and Pasta. United States Department of Agriculture.
Muliarta I G. P., N. Kantun, Sanisah dan N. Soemenaboedhy. 2006. Upaya mendapatkan padi beras merah tahan kekeringan melalui metode seleksi “Back Cross”. Penelitian Hibah Bersaing XI/4 (tidak dipublikasikan).
Fakultas Pertanian Universitas Mataram. 125 h.
Muliarta I G. P., N. Kantun, Sanisah, Kisman dan N. Soemenaboedhy. 2003. Upaya mendapatkan padi beras merah tahan kekeringan melalui metode seleksi “Back Cross”. Penelitian Hibah Bersaing XI/I (tidak dipublikasikan). Fakultas Pertanian Universitas Mataram. 75 h.
Nabhushana I, A. R. Reddy (2004).Rice flavonoid pathway genes, OsDfr and OsAns, are induced by dehydration, high salt and ABA, and contain stress responsive promoter elements that interact with the transcription activator, Osl-MYB. Plant Science 166 (2004) 1505-1513
Nirmala. 2001. Beras merah sumber vitamin B serat dan protein. PT Narya Gunatra. 96 h. Reddy, A. R. 1996. Genetic and molecular
analysis of anthocyanin pigmentation pathhway in rice. Proceedings of the third
international rice genetics symposium. IRRI. Manila.Phillipines. p 16-20
Sbe,K. Yamaguchi-Shinozaki, T. Urao, T.Iwasaki., D. Hosokawa, K.Shinozaki. 1997. Role of Arabidopsis MYC and MYB homolog in drought and Abscisic acid-regulated gen expression, Plant Cell 9 (1997) 1859-1868.
Shi Z.; M. Lin; F. J. Prancis. 1992. Stability of anthocyanin from Trandescantia pallida. J. Food Sci. 57: 758-780
Singh, R. K. and B. D. Chaudary. 1979. Biometrical methods in quantitative genetic analysis, Kalyani Publishers. New Delhi. 304 p.
Suprihatno B, A.A. Derajat, Satoto, S.E. Bahaki, N. Widiana,A.Setyono, S.D. Indrasti, O.S. Lesmana, dan H. Semiring. 2006. Deskripsi varietas padi. Balai Besar Penelelian Tanaman Padi.78 h.