I. PENDAHULUAN
Hormon estrogen merupakan salah satu hormon steroid kelamin, karena mempunyai struktur kimia berintikan steroid yang secara fisiologik sebagian besar diproduksi oleh kelenjar endokrin sistem produksi wanita. Pria juga memproduksi estrogen tetapi dalam jumlah jauh lebih sedikit, fungsi utamanya berhubungan erat dengan fungsi alat kelamin primer dan sekunder wanita.1,2
Hal yang spesifik bagi hormon ini pada wanita usia subur ialah sekresinya dari ovarium berlangsung secara siklik dan peranannya yang sangat penting dalam mempersiapkan kehamilan.1,3,4
Hormon ini juga berperan dalam proses perubahan habitus seorang anak perempuan menjadi wanita dewasa, kemudian menjelang akhir masa reproduksi produksinya mulai menurun dan sekresinya tidak lagi bersifat siklik.1,5
Hormon steroid termasuk ikatan hormon hidrogen, yang mempunyai bermacam-macam pengaruh yang khas, tergantung dari perbedaan dalam susunan gugus metal, ikatan rangkap, hidroksi atau kelompok keton. Hormon ini termasuk zat lipofil yang sedikit larut dalam air.2 Pada tahun 1926 Loewe dan Frank pertama kali melaporkan adanya aktifitas estrogen
dalam darah manusia, sedangkan Frank dan Goldberger pada tahun yang sama berhasil menemukan kondisi “double peak” selama siklus menstruasi normal dengan menggunakan teknik bioassay. Pada tahun 1935 Mac. Corquodale pertama kali mendapatkan kristal estradiol dari cairan folikuler ovarium dan juga estron ditemukan dalam cairan folikel tetapi dalam jumlah yang kecil.6
Estogen alamiah yang terpenting adalah estradiol (E2), estron (E1), dan estriol (E3).
Secara biologis, estradiol adalah yang paling aktif. Perbandingan khasiat biologis dari ketiga hormon tersebut E2 : E1 : E3 = 10 : 5 : 1.7
II. TATA NAMA
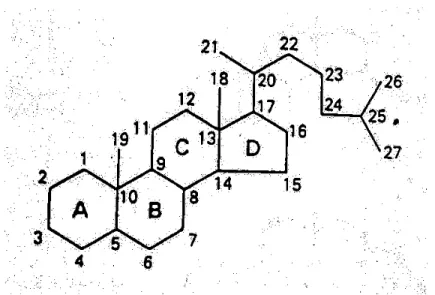
Semua hormon-hormon steroid pada dasarnya memiliki struktur yang sama, hanya saja mempunyai sedikit perbedaan kimiawi yang mengakibatkan terjadinya perbedaan aktivitas biokimiawi. Struktur dasarnya adalah molekul siklopentanolperhidrofenantren, molekul ini terdiri dari 3 buah cicin dari 6 atom karbon dan sebuah cicin dari 5 atom karbon. Cincin
dasar ini ditandai dengan huruf A, B, C, dan D, sedangkan atom karbon diberi angka (gambar 1).7,8
Gambar 1. Struktur dasar molekul siklopentanolperhidrofenantren (dikutip dari O’Malley B10 )
Hormon steroid seks dibagi menjadi 3 kelompok utama berdasarkan jumlah atom karbon yang dimiliki (gambar 2).
1. Seri karbon 21, struktur dasarnya adalah nucleus pregnane, termasuk disini kortikoid dan progestin
2. Seri karbon 19, struktur dasarnya adalah nukleus androstane termasuk disini hormon androgen
3. Seri karbon 18, struktur dasarnya adalah nukleus estrange termasuk disini hormon estrogen.6,7,8
Gambar 2. Pembagian hormon steroid seks (dikutip dari Wibowo7)
Derivat estrange memiliki 3 bentuk yaitu estron, estradiol dan estriol (gambar 3).
Gambar 3. Tiga bentuk derivat estrange (dikutip dari Wibowo7)
Penamaan dari hormon streroid ini menggunakan jumlah atom karbon yang ada, nama dasarnya didahului dengan jumlah yang menunjukkan posisi dari ikatan rangkap, nama-nama tersebut menunjukkan posisi dari ikatan rangkap, nama-nama-nama-nama tersebut menunjukkan apakah terdapat 1, 2 atau 3 ikatan yaitu : - ene, dan –diene, -triene. Setelah nama dasar
diikuti dengan nama kelompok hidroksi yang ditunjukkan dengan jumlah rantai karbon yang terikat, 1, 2 atau 3 kelompok hidroksi yaitu : - ol, - diol, - triol. Kemudian group keton menyusul dipaling akhir dengan nama sesuai jumlah karbon yang terikat 1, 2 atau 3 yaitu : - one, - dione dan – trione, sebagai contoh diperlihatkan pada gambar (gambar 4).8
Gambar 4. Penamaan hormon steroid ( dikutip dari Wibowo7 )
III. SINTESIS ESTROGEN
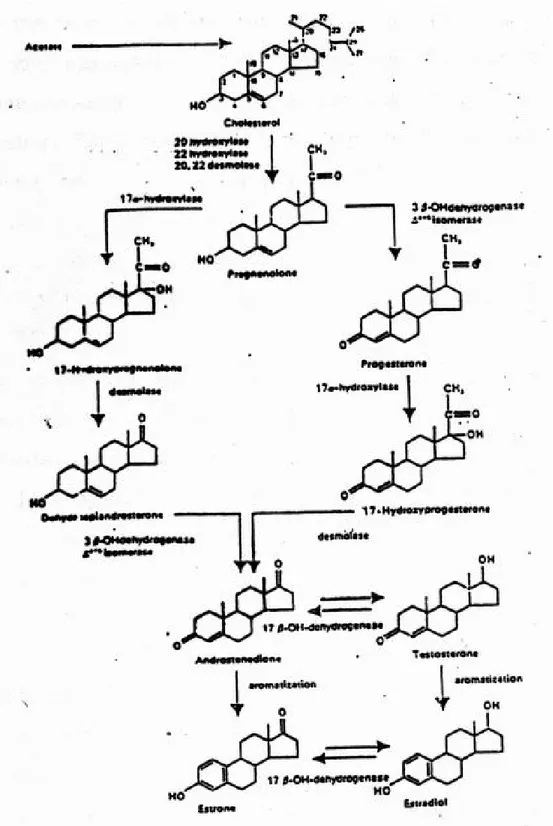
Sintesis hormon estrogen terjadi didalam sel-sel theka dan sel-sel granulose ovarium, dimana kolesterol merupakan zat pembakal dari hormon ini, yang pembentukannya melalui beberapa serangkaian reaksi enzimatik.3,7
Pada tahun 1959 Ryan dan Smith mengemukakan hipotesa 2 sel yakni mekanisme produksi hormon steroid dalam ovarium, hipotesa ini untuk menerangkan kerja sama antara sel theka dan sel granulose dalam pembentukan hormon.
LH diketahui berperan dalam sel theka untuk meningkatkan aktivitas enzim pembelah rantai sisi kolesterol melalui pengaktifan ATP menjadi cAMP, dan dengan melalui beberapa proses reaksi enzimatik terbentuklah androstenedion, kemudian androstenedion yang dibentuk dalam sel theka berfungsi kedalam sel granulose, selanjutnya melakukan aromatisasi membentuk estron dan estradiol 17 β.9,10,11
Kolesterol sebagai pembakal (prekursor) steroid disimpan dalam jumlah yang banyak di sel-sel theka. Pematangan folikel yang mengakibatkan meningkatnya biosintesa steroid dalam folikel diatur oleh hormon gonadotropin.3
Selama pembentukkan hormon steroid, jumlah atom karbon didalam kolesterol atau didalam molekul steroid lainnya dapat diproduksikan tapi tidak pernah ditingkatkan proses pembentukan hormon steroid dapat terjadi reaksi-reaksi sebagai berikut :
1. Reaksi desmolase : pemecahan / pembelahan rantai samping.
2. Konversi kelompok hidroksi menjadi keton atau kelompok keton menjadi kelompok hidroksil : reaksi dehidrogenase.
3. Reaksi hidroksilasi : perubahan kelompok OH. 4. Pemindahan hidrogen : terbentuknya ikatan ganda
5. Saturasi : penambahan hidrogen untuk mengurangi ikatan ganda.8
Kolesterol mengandung 27 atom karbon, setelah hidroksilasi dari kolesterol pada atom C20 dan atom C22 terjadi pemecahan rantai samping menjadi bentuk pregnenolon dan
asam isocaproat, pemecahan ini di samping adanya enzim 20β hidroksilasi dan 22 β hidroksilasi juga adanya peran LH dalam meningkatkan aktivitas enzim.3.6
Dari pregnenolan proses pembentukkan estrogen ada 2 cara yaitu : 1. Melalui ∆5 – 3 β hidroksi steroid Pathway / Pregnenolon pathway 2. Melalui ∆4 – 3 β ketone pathway / Progesteron pathway
Cara yang pertama melalui pembentukan dehidroepiandrosteron, sedangkan cara yang kedua melalui pembentukan progesterone, (gambar 5). Progesteron dibentuk dari pregnenolon melalui penghilangan atom hydrogen dari C3 dan pergeseran ikatan ganda
dari cincin B pada posisi 5-6 ke cincin A pada posisi 4-5, perubahan ini oleh adanya bantuan enzyme 3 β hidroksi dehidrogenase dan ∆4-5 isomerase, selanjutnya dengan bantuan enzyme 17α hidroksilase, progesteron akan diubah menjadi 17 hidroksi progesterone yang kemudian mengalami demolase menjadi bentuk testoteron, yang selanjutnya testosterone mengalami aromatisasi (pembentukan gugus hidroksi fenolik pada atom C3) menjadi estradiol (E2), sedangkan androstenedion juga dapat mengalami
aromatisasi membentuk eston (E1) Proses aromatisasi androstenedion dipengaruhi juga
oleh FSH. Sedangkan pembentukan estrogen melalui pembentukkan dehidroepiandrossteron yaitu dengan cara perubahan pregnenolon menjadi 17 hidroksi pregnenolon dengan bantuan enzim 17α hidroksilase, yang kemudian 17 hidroksi pregnenolon mengalami desmolase membentuk dehidroepiandrosteron. Dengan bantuan enzim 3β OH dehidrogenase serta ∆4-5 isomerase, dehidroepiandrosteron diubah menjadi androstenedion dengan cara penghilangan hydrogen dan atom C3 serta
selanjutnya sintesis hormon estrogen sama halnya seperti yang diperlihatkan melalui pembentukan progesteron.8,9,10
Pada wanita masa reproduksi, estradiol diproduksi sebanyak 0,09-0,25 mg/hari, estron 0,11-0,26 mg/hari. Kadar estradiol dalam darah berkisar antara 20-500 pg/ml dan estron 50-400 pg/ml, sedangkan pada wanita masa menopause kadar estrdiol dibawah 10 pg/ml, dan kadar estron dibawah 30 pg/ml, sebagai perbandingan diketahui kadar estradiol pada laki-laki berkisar antara 15-25 pg/ml dan kadar estron 40-75 pg/ml.6,10
Kadar estradiol mencapai puncaknya pada saat 2 hari sebelum ovulasi dengan kadar mencapai 150-400 pg/ml. Setelah ovulasi kadar estradiol menurun, untuk kemudian meningkat lagi sampai kira-kira hari ke 21, selanjutnya hormon ini menurun lagi sampai akhir siklus.7,11
Seperti diketahui zat awal untuk sintesis hormon steroid terdapat di semua kelenjar hormon steroid, hormon mana yang pasti dan dimana akan dihasilkan tergantung dari :
1. Reseptor yang tersedia untuk pengaturan hormon (ACTH, FSH, LH)
2. Enzim yang dominan untuk perubahan-perubahan susunan molekul steroid dalam setiap kelenjar hormon.3
IV. SUMBER-SUMBER ESTROGEN ENDOGEN
Sumber utama estradiol pada wanita adalah sel-sel teka dan granulosa ovarium dan turunan luteinisasi dari sel-sel ini. Berdasarkan teori sintesis estrogen kedua sel ini, sel-sel teka mensekresikan androgen yang menyebar ke sel-sel granulosa teraromarisasi menjadi estrogen. Kedua bentuk sel ini mungkin mampu untuk membentuk androgen dan estrogen. Estron dan estriol utamanya dibentuk di hati dari estradiol.2,10,13
Aktivitas aromatase juga telah terdeteksi pada otot, lemak, jaringan saraf, dan sel-sel Leydig dari testes. Selama kehamilan, estriol disintesis di sinsisiotrofoblas oleh aromatisasi dari 16αhidroksiandrostenedion. Ikatan selanjutnya berasal dari 16α-hidroksiepiandrosteron sulfat diubah menjadi dehidroepiandrosterone sulfat yang dihasilkan di kelenjar adrenal janin. Kombinasi kelenjar adrenal janin dan hati dan plasenta telah dirujuk sebagai “unit fetoplasenta dari biosintesis steroid”.2,10,13
Gambar 5.Dua cara pembentukan estrogen (dikutip daari Wibowo7 )
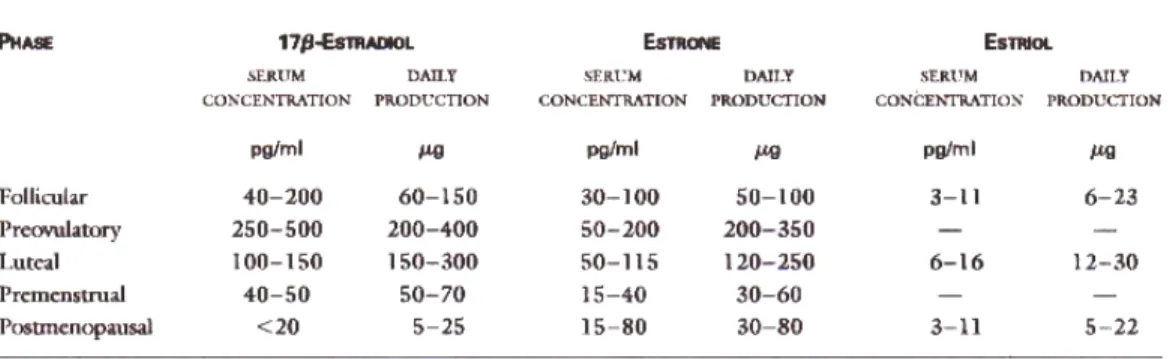
Pubertas pada gadis-gadis diinisiasi oleh gelombang-gelombang nokturnal amplitudo rendah gonadotropin yang meningkatkan konsentrasi estradiol serum menjadi 15-30 pg per milimeter (55-128 pmol per liter). Selama siklus menstruasi, produksi estradiol bervariasi secara siklis, dengan rata-rata paling tinggi dan konsentrasi pada fase preovulatoir (tabel 1). Produksi estradiol dan konsentrasi serum lebih rendah secara premenstrual. Pada periode perimenopause, deplesi folikel ovarium menyebabkan keadaan menurun pada produksi estradiol ovarium, walaupun konsentrasi estradiol serum memiliki pertimbangan bervariasi. Pada wanita-wanita postmenopause, konsentrasi estradiol serum sering lebih rendah dari 20 pg per mililiter (73 pmol per liter), dan kebanyakan estradiol dibentuk oleh konversi ekstragonad testosteron. Estron merupakan estrogen predominan pada wanita-wanita ini (tabel 1). Kadar sintesis estrogen pda jaringan-jaringan ekstragonad meningkat sesuai usia dan berat tubuh.2,10,13
Tabel 1. Rata-rata produksi dan konsentrasi serum estrogen dengan siklus menstruasi pada wanita-wanita normal.13
Sedikit diketahui tentang faktor-faktor yang meregulasi produksi estrogen pada wanita-wanita postmenopause, tetapi pada periode reproduktif kontrol dilakukan oleh gonadotropin. Gen-gen responsif terhadap follicle stimulating hormone, untuk mudahnya, mengatur ekspresi enzim-enzim steroidogenik. Kontrol trofik ini dimodifikasi oleh faktor-faktor parakrin. The insulin-like growth factors, contohnya, memfasilitasi kerja follicle stimulating hormone pada sel-sel granulosa. Pada mRNA reseptor-androgen di sel-sel granulosa diregulasi menurun oleh follicle stimulating hormone. Hubungan terbalik ini
merupakan bagian mekanisme yang menentukan folikel akan menjadi dominan sebagai folikel penseksresi hormon dalam siklus menstruasi. Pada jaringan-jaringan tepi, produksi estrogen yang berbeda dan interkonversinya tergantung pada ekspresi lokal dan aktivitas aromatase, 17β-hidroksisteroid dehidrogenase, dan estron sulfatase. Polimorfisme pada gen-gen pengkode untuk enzim-enzim steroidogenik mempengaruhi estrogen. Evaluasi selanjutnya dari polimorfisme ini dengan membandingkan risiko kanker atau kebutuhan untuk terapi estrogen bisa membuat kemungkinan suatu pendekatan terapi lebih individual pada wanita-wanita postmenopause13
V. TRANSPORT DAN METABOLISME ESTROGEN
Kolesterol adalah prekursor estrogen yang umum. Aktivitas 17β hidroksidehidrogenase akan mengkonversi androstenedion menjadi testoteron yang mana bukan merupakan produksi terbesar yang dihasilkan dari ovarium. Testoteron ini cepat mengalami demetilasi pada posisi C19 dan proses aromatisasi menjadi estradiol yang
merupakan estrogen terbanyak yang disekresi ovarium. Estradiol juga meningkat sampai mencapai jumlah yang cukup banyak dari androstenedion melalui estone. Estriol merupakan metabolit perifer dari estone dan estradiol dan bukan merupakan sekresi ovarium. Pembentukan estriol merupakan konversi dari material biologis yang aktif menjadi bentuk yang kurang aktif. Konversi steroid di jaringan perifer tidak selalu merupakan bentuk yang inaktif. Androgen bebas dikonversi di perifer untuk menjadi bebas, misalnya di kulit dan sel adiposa.3,6,8,12
Pada wanita menopause, kelenjar adrenal tetap merupakan sumber utama androgen bersikulasi. Pada laki-laki hampir semua estrogen bersikulasi dihasilkan dari konversi androgen di perifer.8
Dari keterangan diatas jelas bahwa estrogen yang bersikulasi pada wanita merupakan dari sekresi ovarium langsung yaitu estradiol dan estrone, ditambah konversi dari prekursor C19 Sejumlah estrogren telah ditemukan dalam cairan folikuler, urine, darah,
feses, plasenta dan kandung empedu wanita. Organ-organ tersebut juga ditemukan sejumlah estrogen.6,12
Didalam sirkulasi darah estrogen terdapat dalam bentuk terikat dan tidak terikat, sebagian besar estrogen terikat pada β globulin (69%), sebuah karier protein yang
diketahui sebagai seks hormon binding globulin (SHBG), 30% bagian lainnya terikat albumin dan sisanya sekitar 2-3% terlepas bebas.3,6,810
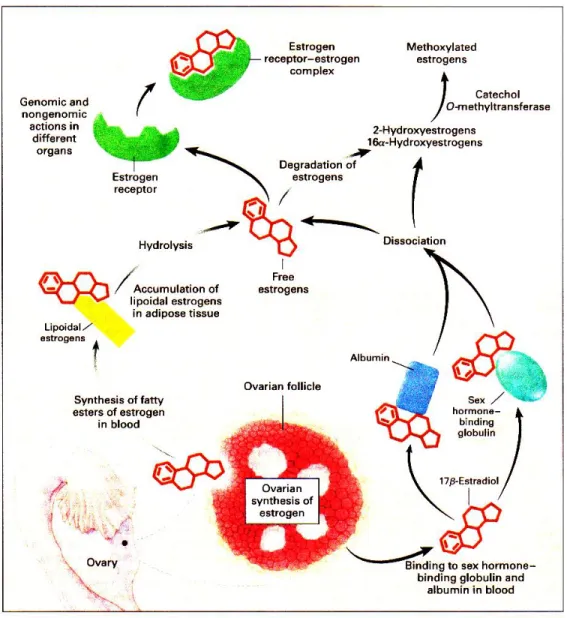
Estrogen lipoid adalah ester lemak dari estrogen yang terdiri dari beberapa kelas hormon-hormon steroid. Walaupun mereka diproduksi dalam berbagai jaringan in vitro, estrogen lipoid ditemukan predominannya dalam jaringan adiposa (gambar 6). Mereka disintesis dalam darah, ke sirkulasi dan berikatan dengan lipoprotein. Keseluruhan, kurang dari 10% estradiol serum berhubungan dengan lipoprotein, terutama lipoprotein densitas tinggi, tetapi estradiol serum bisa ditransfer ke lipoprotein densitas rendah oleh mekanisme karier yang tidak diketahui. Estrogen lipoid lebih resisten untuk katabolisme daripada estrogen bebas dan karena itu dijelaskan dengan lambat. Setelah pengambilan yang dimediasi-reseptor-lipoprotein oleh hidrolisis sel dan lokal, mereka mungkin memberikan secara potensial suatu simpanan steroid yang relevan. Estrogen lipoid juga menurunkan konsentrasi estradiol yang penting untuk menghambat aksidasi lipoprotein densitas rendah in vitro13
Estrogen dimetabolisme hepar menjadi bentuk terkonjugasi dengan sulfat atau glukuronat, metabolit ini bersifat inaktif dijaringan perifer.6
Telah ditemukan sejumlah estrogen metabolit dalam urine, metabolit ini terdapat dalam bentuk terkonjugasi dengan sulfat atau glukuronat . Gallagher dkk telah mengukur berbagai fraksi urine setelah pemberian estradiol berlabel karbon radiokatif, berapa banyaknya suatu fraksi dapat ditemukan. Estrogen dikonjugasikan di hepar dengan glucoronat dan sulfat sebelum diekskresikan ke urine. Sekitar 70% metabolit estrogen diekskresikan melalui urine sedangkan sisanya diekskresikan melalui feses.6,8
VI. AKSI MOLEKULER ESTROGEN
Kerja utama estrogen yang spesifik ditentukan oleh struktur hormon, subtipe atau isoform reseptor estrogen yang terlibat, karakteristik promotor gen target dan keseimbangan koaktivator dan koreseptor yang memodulasi respon transkripsional akhir dengan kompleks estrogen dan reseptor estrogen.2,13
Gambar 6. Sintesis ovarium, transport dan metabolisme estrogen13
VI.1. Aktivasi Reseptor Estrogen Tergantung Ligan – Jalur Klasik
Reseptor estrogen, tidak melekat dengan ligannya dan lepasnya ikatan pada sitoplasmanya atau lokasi utamanya, dilekatkan dengan protein yang berhubungan dengan reseptor. Protein-protein ini bertindak sebagai pengantar yang menstabilkan reseptor pada keadaan tidak aktif atau menutupi domain ikatan DNA dari reseptor. Protein yang berhubungan dengan reseptor lainnya bisa mengkontribusikan hubungan silang diantara jalur-jalur sinyal yang berbeda. Lokasi pasti reseptor estrogen dan reseptor hormon streoid lainnya tidak seluruhnya jelas. Mungkin dalam suatu distribusi seimbang di antara sitoplasma dan nukleus; keseimbangan ini selanjutnya digantikan setelah ikatan ligan.
Sebagai estrogen bebas yang berdifusi ke dalam sel, estrogen mengikat domain ikatan ligan dari reseptor, yang dipisahkan dari pengantar sitoplasmanya; kompleks estrogen dan reseptor estrogen selanjutnya berdifusi ke dalam nukleus sel. Kompleks estrogen-reseptor estrogen ini berikatan dengan bagian spesifik dari DNA yang disebut elemen-elemen respon estrogen sebagai homodimer atau heterodimer. Kompleks estrogen-reseptor estrogen tidak beikatan hanya pada elemen respon tetapi juga dengan koaktivator atau represor reseptor inti (gambar 7). Mekanisme pasti dari translokasi inti kompleks estrogen-reseptor estrogen tidak juga sepenuhnya diketahui, tetapi diketahui bahwa protein sitosolik Kaveolin-1 menstimulasi proses translokasi ini melalui interaksi langsung dengan molekul reseptor.2,10,1,14
Estrogen juga meregulasi transkripsi gen-gen yang elemen-elemen respon estrogen fungsionalnya lemah dengan memodulasi aktivitas faktor transkripsi lainnya. Pengikatan reseptor estrogen dengan subunit pengaktivasi protein 1, contohnya, menghasilkan pembentukan faktor transkripsi. Reseptor estrogen juga berinteraksi dengan faktor inti
KB.13
VI.2. Reseptor Estrogen α dan β
Reseptor estrogen merupakan anggota dari superfamili reseptor-hormon inti, yang memiliki kira-kira 150 anggota yang telah dikenal. Reseptor estrogen memiliki beberapa domain fungsional. Domain yang berikatan dengan DNA terdiri dari dua ikatan seng yang terlibat dalam pengikatan dan dimerisasi reseptor. Domain yang berikatan dengan ligan berisi perangkat asam amino berbeda yang mengikat ligan berbeda; domain ini juga berinteraksi dengan protein koregulator. Domain terminal-N memiliki derajat variabilitas yang tinggi dan normalnya terdiri dari domain transkripsi yang bisa berinteraksi secara langsung dengan faktor-faktor perlengkapan transkripsional. Domain terminal-C mengkontribusi kapasitas transaktivasi reseptor. 14
Ada dua subtipe reseptor estrogen dan beberapa isoform serta sambungan varian dari setiap subtipe. Subtipe pertama, reseptor estrogen α yang klasik, pertama kali diklon tahun 1986; subtipe kedua, reseptor estrogen β ditemukan paling terkini. Kedua subtipe reseptor ini bervariasi dalam struktur, dan gen-gen pengenkode mereka di dalam kromosom-kromosom yang berbeda. Gen reseptor estrogen α telah dipetakan pada lengan panjang
kromosom 6, sedangkan gen reseptor estrogen β berlokasi pada pita q22-24 dari kromosom 14.13,14
Walaupun domain ikatan DNA dari reseptor estrogen α dan β sangat mirip, derajat keseluruhan homologi dari reseptor adalah rendah. Khususnya sesungguhnya untuk domain berikatan ligan, yang hanya 55% dari bagian asam amino dibagikan. Hasilnya, beberapa ligan mengikat kedua reseptor dengan afinitas berbeda (tabel 2). Contohnya, 17α-estradiol kerja singkat dan secara biologis estron lemah memiliki afinitas lebih tinggi untuk reseptor estrogen α, tetapi akhirnya kedua fitoestrogen, genistein dan koumestrol, berikatan dengan afinitas lebih tinggi dengan reseptor estrogen β. Modulator reseptor estrogen selektif raloksifen berikatan dengan afinitas lebih tinggi dengan reseptor estrogen α, sedangkan beberapa polutan lingkungan, seperti alkilfenol, memiliki afinitas lebih tinggi untuk reseptor estrogen β.
Tabel 2. Afinitas-afinitas relatif ikatan ligan-ligan yang berbeda untuk reseptor estrogen α dan reseptor estrogen β.13
Distribusi jaringan reseptor estrogen α dan reseptor estrogen β berbeda, walaupun ada beberapa tumpang tindih. Sel-sel granulosa dan perkembangan spermatid berisi kebanyakan reseptor estrogen β dan subtipe ini ada pada beberapa jaringan-jaringan target nonklasik, termasuk ginjal, mukosa usus, parenkim paru, sumsum tulang, tulang, otak, sel-sel endotelial, dan kelenjar prostat. Kontrasnya, endometrium sel-sel-sel-sel kanker payudara, dan stroma ovarium isinya kebanyakan reseptor estrogen α.10,13,14
Seorang laki-laki dengan reseptor estrogen fungsional yang lemah ditemukan memiliki osteoporosis berat dan menurunkan fertilitas. Kedua tikus jantan dan betina dengan gen-gen untuk reseptor estrogen-gen α yang terganggu adalah infertil, densitas tulang-tulang femoral secara ringan menurun pada betina dan tandanya menurun pada jantan. Efek protektif estrogen setelah trauma pembuluh darah karotid tidak mengubah jatuhnya reseptor estrogen α tikus. Tikus-tikus betina yang gen untuk reseptor estrogen β rendah adalah infertil, dan tikus jantan dengan defek ini memiliki hiperplasia prostat dan hilangnya lemak abdominal tetapi subur. Pada tikus yang gen-gen untuk kedua reseptor estrogen α dan β rendah, betina memiliki ovarium yang berisi struktur seperti tubulus seminiferus yang berisi dengan sel-sel seperti Sertoli, dan jantannya infertil karena reduksi pada jumlah dan mortalitas sperma epididimis- suatu kemiripan fenotipe untuk reseptor estrogen α- membantu tikus.10,13,14
VI.3. Modulator Reseptor Estrogen Selektif
Istilah “modulator reseptor estrogen selektif” diperkenalkan untuk mendefinisikan ligan nonsteroid seperti tamoksifen yang antagonis dengan kerja estrogen dalam beberapa jaringan, seperti payudara dan mirip kerjanya pada tempat lainnya seperti uterus. Di antara wanita-wanita postmenopause, kerja agonis estrogen menyebabkan tulang untuk menjaga densitas dan dalam sistem kardiovaskuler dan otak untuk menjaga fungsinya, tetapi tidak pada payudara atau endometrium. Raloksifen memiliki efek seperti estrogen pada jaringan tulang dan pada konsentrasi lemak serum, tetapi tidak pada payudara dan jaringan endometrium. Pada otak, bagaimanapun, raloksifen lebih dari antagonis estrogen, karena meningkatnya gejala-gejala vasomotor dari defisiensi estrogen.10,13,14
Mekanisme selektivitas jaringan dari ikatan ini adalah kompleks. Pada kedua reseptor estrogen α47 dan reseptor estrogen β48, konformasi domain ikatan ligan mengubah jalur
yang berbeda ketika estradiol, raloksifen, atau genistein mengikat reseptor. Akibatnya, konformasi domain transaktivasi utama diubah sehingga permukaan berbeda terpapar dengan koaktivitator atau represor reseptor nukleus. Oleh karena itu, efek transkripsional bisa bervariasi. Contohnya, tamoksifen dan raloksifen bertindak sebagai aktivitor transkripsional pada tempat protein pengaktivasi-1 ketika membentuk kompleks dengan reseptor estrogen β tetapi menekan transkripsional ketika pembentukan kompleks dengan reseptor estrogen α. Estradiol mengaktivasi traskripsi ketika berikatan dengan dengan reseptor estrogen α tetapi menekankan efek transkripsional yang berlawanan ketika berikatan dengan reseptor estrogen β.10,13,14
VI.4. Koaktivator dan Korepresor Reseptor Estrogen
Reseptor estrogen berinteraksi dengan beberapa protein koregulator yang menghubungkan antara reseptor teraktivasi dan perlengkapan transkripsional, seperti yang dicatat sebelumnya (gambar 7). Untuk membentuk kompleks transkripsi-inisiasi, kumpulan berbagai faktor, seperti protein yang berikatan dengan kotak TATA dan faktor yang berhubungan lainnya di kotak TATA, dibutuhkan oleh RNA polimerase II. Kontrasnya, protein koregulator reseptor-nukleus berinteraksi dengan molekul reseptor untuk memodulasi kapasitas transkripsionalnya. Kompleks oligomerik ini, terdiri dari reseptor estrogen, faktor transkripsi dan protein koaktivator dan korepresor, dipercaya untuk menyajikan stabilitas dan spesifisitas transkripsional.10,13,14
Protein koaktivator reseptor-steroid-160kD dan protein pengikat elemen-respon siklik AMP-p300 tampak memiliki kapasitas paling besar untuk meningkatkan aktivitas transkripsional reseptor-reseptor estrogen. Kontrasnya, korepresor menekan transkripsi setelah ditambatkan ke suatu promotor oleh reseptor yang diikat DNA. Protein yang bekerja sebagai korepresor aktivitas reseptor-estrogen merupakan anggota dari kelas selanjutnya. Protein ini secara langsung berinteraksi dengan agonis- atau antagonis- dibuat reseptor tetapi juga bersaing dengan protein koaktivator reseptor-steroid. Hal ini merupakan suatu contoh bagaimana keseimbangan korepresor dan koaktivator mempengaruhi aktivitas transkripsional dari reseptor estrogen teraktivasi. Oleh karena itu, terlihat seperti bagian dari antagonis reseptor estrogen yang merupakan hasil rekrutmen korepresor.10,13,14
VI.5. Elemen-Elemen Respons-Estrogen dan Unit-Unit Respons-Estrogen
Reseptor estrogen merupakan faktor transkripsi yang, setelah teraktivasi, membuat interaksi nukleus langsung dengan mengikat elemen-elemen respons estrogen DNA, yang memberi indusibilitas estrogen pada gen. Elemen respon-estrogen tampak pada daerah regulator dari gen-gen target estrogen (gambar 7).10,13,14
Bagian 5’GGTCAnnnTGACC3’ dari Xenopus laevis gen vitelogenin (n menandakan nukleotid random) telah didefinisikan sebagai konsensus bagian elemen respons-estrogen. Hal ini merupakan suatu kebalikan bertahap 13-bp dengan jarak 3 vaiabel basa. Bagaimanapun, hanya sejumlah kecil dari kebanyakan gen-gen yang dapat menginduksi estrogen berisi konsensus elemen-elemen respon estrogen ini. Pada kebanyakan kasus, varian elemen respon-estrogen telah digambarkan. Pada promotor gen Bcl-x1 murin, untuk
mudahnya, bagian 5’GGTCAnnnTGGCC3’, yang berbeda dari bagian konsensus dengan 1 bp, memediasi indusibilitas estrogen. Bagian-bagian varian ini mengikat reseptor estrogen dengan afinitas kurang, tergantung pada sisi basa.
Lebih lanjut, varian elemen respons-estrogen atau bahkan elemen respons-estrogen parsial, sering terpisah oleh berbagai bagian basa, bisa bekerja dengan kombinasi untuk memberi keresposifan estrogen. Kombinasi ini dirujuk sebagai unit respons-estrogen. Pada gen untuk pengubahan faktor pertumbuhan α manusia, untuk mudahnya, bagian yang resposif estrogen adalah suatu elemen 5’GGTCAnnnTGCCC3’ yang dipisah oleh 20 bp dari suatu elemen 5’GGTGAnnnTAGCC3’.10,13,14
VII. AKSI ESTROGEN FISIOLOGIS
Estrogen menstimulasi pertumbuhan, aliran darah, dan retensi air pada organ seksual dan juga terlibat dalam penyebab kanker payudara dan kanker endometrium. Pada hati, estrogen meningkatkan reseptor lipoprotein, menghasilkan penurunan konsentrasi serum dari kolesterol low-density lipoprotein. Pada tulisan lainnya, estrogen meningkatkan potensial koagulasi. Pada saluran cerna, estrogen bisa sebagai pelindung melawan kanker kolon. Pada penuaan kulit, estrogen meningkatkan turgor dan produksi kolagen serta mengurangi kerutan yang dalam (gambar 8)2,9,10,13
VIII.1. Aksi pada Jaringan Payudara
Unit lobuler saluran terminal dari jaringan payudara wanita-wanita muda sangat responsif dengan estrogen. Pada jaringan payudara, estrogen menstimulasi pertumbuhan dan diferensiasi saluran epitelium, menginduksi aktivitas mitotik saluran sel-sel silindris, dan menstimulasi pertumbuhan jaringan penyambung. Estrogen juga menghasilkan efek seperti histamin pada mikrosirkulasi payudara. Densitas reseptor estrogen pada jaringan payudara sangat tinggi pada fase folikuler dari siklus menstruasi dan menurun setelah ovulasi. Estrogen menstimulasi pertumbuhan sel-sel kanker payudara. Pada wanita-wanita postmenopause dengan kanker payudara, konsentrasi estradiol tumor tinggi, karena aromatisasi in situ, meskipun adanya keonsentrasi estradiol serum yang rendah.2,9,10,13
VIII.2. Aksi pada Sistem Saraf Pusat
Hipotesis aromatisasi otak mengajukan bahwa diferensiasi seksual pada otak- yaitu kemampuan estrogen untuk menyebabkan pelepasan sekresi gonadotropin pada wanita-wanita- tergantung pada konversi lokal androgen menjadi estrogen. Rata-rata aromatisasi androgen menjadi estrogen pada otak rendah jika dibandingkan dengan jaringan-jaringan lainnya, tetapi setidaknya, produksi estrogen lokal dipercaya memiliki aksi penting. Salah satu contoh dari aksi sinergistis estrogen ini dengan neurotrofins yang direfleksikan pada regulasi reseptor resiprokal atau jalur-jalur sinyal berpasangan.2,9,10,13
Pada kehidupan selanjutnya, estrogen diduga memiliki aksi neuroprotektif. Pada jaringan otak dari tikus dewasa, estrogen menginduksi pembentukan ulang dendrit dan sinaptik dan menyebabkan aktivasi glial. Pada saraf-saraf hippokampus, suatu area yang melibatkan memori, estrogen meningkatkan densitas dari reseptor N-metil-D-aspartat dan meningkatkan sensitivitas saraf untuk masukan yang dimediasi oleh reseptor-reseptor ini.
Pada kultur sel-sel neuroblastoma manusia, estrogen memiliki efek neuroprotektif dan mereduksi generasi peptida beta-amiloid. Beberapa data epidemiologis mendukung bahwa pada wanita-wanita postmenopause, defisiensi estrogen berhubungan dengan penurunan fungsi kognitif dan meningkatkan risiko penyakit Alzheimer. Bagaimanapun, pada uji terandomisasi, pemberian estrogen tidak memiliki efek menguntungkan pada wanita-wanita yang menderita penyakit Alzheimer.2,9,10,13
VIII.3. Efek-Efek Vaskuler
Estrogen diduga menjadi agen-agen vasoprotektif alami. Reseptor estrogen telah terdeteksi pada sel-sel otot polos arteri koroner dan sel-sel endometrium pada berbagai tempat. Estrogen menyebabkan vasodilatasi jangka pendek dengan meningkatkan pembentukan dan pelepasan nitrat oksida dan prostasiklin pada sel-sel endotelial. Juga menurunkan tonus otot-otot polos vaskuler dengan pembukaan saluran kalsium spesifik melalui mekanisme yang tergantung pada siklik guanosin monofosfat. Peranan protektif estrogen melawan aterosklerosis didukung dengan penemuan bahwa pengobatan estrogen menurunkan progresi aterosklerosis arteri koroner pada monyet-monyet yang telah diooforektomi. Bagaimanapun juga, tidak ada efek pada keberadaan awal plak-plak. Pada tingkat seluler, estrogen menghambat apoptosis sel-sel endotelial dan mempromosikan aktivitas angiogenisnya in vitro.2,9,10,13
Walaupun penemuan ini ada, salah satu dari pertanyaan penting pada kesehatan wanita- apakah pengobatan estrogen pada periode postmenopause mencegah aterosklerosis- masih kontroversial. Penemuan yang dapat membantu dari studi-studi epidemiologis harus diseimbangkan dengan lemahnya keuntungan estrogen untuk proteksi sekunder melawan penyakit kardiovaskuler pada jantung dan Studi Sulih Progestin/Estrogen.2,9,10,13
VIII.4. Efek-Efek pada Tulang
Kedua osteoklas dan osteoblas mengekspresikan reseptor estrogen dan merupakan target langsung untuk estrogen, tetapi keseluruhan, estrogen diklasifikasikan sebagai agen-agen antiresoptif. Estrogen secara langsung menghambat fungsi osteoklas. Pada tikus yang diooforektomi, defisiensi estrogen meningkatkan produksi interleukin-6, interleukin-1, dan tumor nekrosis faktor pada osteoblas dan sel-sel stromal turunan tulang lainnya. Faktor-faktor ini secara tidak2,9,10,13
Langsung menstimulasi diferensiasi osteoklas. Pada ekstrak tulang dari wanita-wanita postmenopause dengan osteoporosis, konsentrasi interleukin-6 dan interleukin-1 mRNA juga tinggi. Defisiensi estrogen dikenal untuk mengakselerasikan pengeroposan tulang dan meningkatkan suseptibilitas untuk fraktur. Terapi estrogen mengurangi pengeroposan
tulang dan mereduksi risiko fraktur pada wanita-wanita dengan osteoporosis dan selanjutnya tanpa kondisi ini untuk lamanya terapi.2,9,10,13
Gambar 8. Efek-efek estrogen pada system organ yang berbeda(13)
IX. RINGKASAN
1. Estrogen merupakan salah satu hormon steroid kelamin. 2. Struktur estrogen yaitu siklopentanoperhidrofenantren.
3. Derivat estrange pada wanita masa reproduksi terdiri dari : estrone (E1), estradiol (E2)
dan estriol (E3).
4. Sintesis estrogen terutama terjadi di dalam sel teka dan sel granulose ovarium. 5. Metabolisme estrogen terjadi di hepar
6. Hasil metabolisme berupa konjugasi dengan asam sulfat atau dengan asam glucuronat diekskresikan sebagian besar molekul urine dan sisanya melalui feses.
7. Fungsi estrogen terutama mempengaruhi pertumbuhan dan perkembangan alat kelamin primer dan sekunder pada wanita.
X. RUJUKAN
1. Suherman KS. Farmakologi dan terapi. Edisi ke-4 Jakarta: Bagian Farmakologi FK-UI, 1995; 439-443
2. Guyton A. Fisiologi kedokteran. Edisi ke-7. Jakarta: EGC, 1994; 330-333
3. Jacob TZ, Baziad A. Endokrinologi reproduksi. Edisi ke-1. Jakarta: KSERI, 1994; 43-51
4. Supono. Ilmu kebidanan. Bagian kebidanan dan kandungan FK-Unsri. Palembang, 1985;28-29
5. Clayton SG, Lewis TLT, Pinker G. Gynaecology by Ten Teacher. 13th ed, London: Butler & Tanner ltd, 1981;43-47
6. Jones HW, Jones GS. Novaks textbook of gynaecology. 10th ed, Baltimore: The Williams & Wilkins Company, 1981; 29-39
7. Wibowo B. Ilmu kandungan. Edisi ke-2. Jakarta: Yayasan Bina Pustaka Sarwono Prawiroharjo, 1994; 70-74
8. Speroff L, Glass RH, Kase NG. Clinical gynaecology endocrinology and infertility. 6th ed, Baltimore: Williams and Wilkins, 1999; 279-289
9. Cunningham FG, Mac Donald PC, Gant NF. Williams obstetri. Edisi ke-18. Jakarta: EGC, 1995; 1097-1099
10. O’Malley B, Straat CA. Yen Jaffe. Reproductive endocrinology, 3th ed, Philadelpia: WB Saunders Company, 1991, 156-168
11. Badziad A, Endokrinologi ginekologi, edisi ke-2. Jakarta:2003; 113-122
12. Daryl K, Granner MD. Harper’s Biochemistry, 22th ed, London: Appleton & Lange, 1990; 521-523
13. Gruber CJ, Tschugguei W, Schneebeger C, Huber JC. Production and action of estrogens. N Engl J Med 2002; 346: 340-50
14. Wyeth-Ayerst Laboratories, The new sciense of estrogen receptors. US Pharmacist 2001