ISOLASI DAN KARAKTERISASI BAKTERI PENGHASIL
AHL-LAKTONASE ASAL LAHAN PERTANIAN LUAR
PULAU JAWA
AIDATUN FITRIYAH
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2011
ABSTRAK
AIDATUN FITRIYAH. Isolasi dan karakterisasi bakteri penghasil AHL-laktonase asal lahan
pertanian luar pulau Jawa. Dibimbing oleh IMAN RUSMANA dan ALINA AKHDIYA.
Quorum sensing (QS) adalah komunikasi antar sel bakteri yang dimediasi oleh autoinduser
(AI) untuk mengatur ekspresi gen-gen tertentu ketika populasi sel bakteri telah mencapai quorum.
AI bakteri Gram negatif adalah N-acyl homoserine lactone (AHL). Sistem QS bekerja pada
gen-gen untuk perilaku populasi bakteri seperti pembentukan biofilm, virulensi pada inang,
bioluminescence, dan pembentukan antibiotik. Proses QS dapat dirusak oleh senyawa anti-QS.
AHL-laktonase merupakan salah satu senyawa anti-QS yang disandikan oleh gen aiiA. Penelitian
ini bertujuan untuk mengisolasi dan mengkarakterisasi bakteri penghasil AHL-laktonase asal 16
lahan pertanian luar pulau Jawa. Bioesai aktivitas degradasi AHL dilakukan menggunakan
Chromobacterium violaceum sebagai bioindikator. Gen aiiA dideteksi dengan menggunakan
primer spesifik aiiAF dan aiiAR, sedangkan identifikasi molekuler dilakukan berdasarkan sekuen
gen 16S rRNA. Sebanyak 58 isolat bakteri berhasil diisolasi dan enam diantaranya memiliki
aktivitas degradasi AHL (PAD1a, BAL2a, NTT3a, NTT3e, NTT6B6, dan NTT6B7). Di antara
enam isolat tersebut, hanya isolat NTT3a dan NTT3e yang menunjukkan amplikon berukuran 800
bp ketika diamplifikasi menggunakan primer aiiAF dan aiiAR. Analisis terhadap sekuen gen aiiA
pada isolat NTT3a dan NTT3e menunjukkan tingkat homologi 99% dengan gen AHL-laktonase
Bacillus sp. 91.
Kata kunci: quorum sensing, autoinduser, AHL-laktonase, gen aiiA.
ABSTRACT
AIDATUN FITRIYAH. Isolation and characterization of AHL-lactonase producing bacteria from
agricultural land outside Java. Under supervision of IMAN RUSMANA and ALINA AKHDIYA.
Quorum sensing (QS) is communication between bacterial cells mediated by autoinducer
(AI) to regulate the expression of specific genes in certain bacterial population quorum. AI of
Gram negative bacteria is N-acyl homoserine lactone (AHL). QS system controls behaviors of
bacterial population on biofilm formation, virulence, bioluminescence, and formation of
antibiotics. QS process can be inhibited by anti-QS compound. AHL-lactonase is one of anti-QS
compound encoded by aiiA gene. The aims of this study were to isolate and characterize bacteria
producing AHL-lactonase from agricultural land outside Java. Soil samples were taken from 16
locations of agricultural lands. Bioassay was performed by AHL degradation of Chromobacterium
violaceum. Detection of aiiA gene was conducted using specific primer aiiAF and aiiAR, while
molecular identification was conducted based on 16S rRNA gene sequence. It was found that from
58 isolates, six isolates had AHL degradation activity i.e. PAD1a, BAL2a, NTT3a, NTT3e,
NTT6B6, dan NTT6B7 isolates. Two isolates (i.e. NTT3a and NTT3e isolates) had aiiA gene. And
indicated by 800 bp product. They were 99% closely related to AHL-lactonase gene from Bacillus
sp 91.
ISOLASI DAN KARAKTERISASI BAKTERI PENGHASIL
AHL-LAKTONASE ASAL LAHAN PERTANIAN LUAR
PULAU JAWA
AIDATUN FITRIYAH
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2011
Judul
: Isolasi dan Karakterisasi Bakteri Penghasil AHL-laktonase Asal
Lahan Pertanian Luar Pulau Jawa
Nama
: Aidatun Fitriyah
NIM
: G34070072
Menyetujui,
Mengetahui,
Ketua Departemen Biologi
Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
Dr. Ir. Ence Darmo Jaya Supena, M.S
NIP 196410021989031002
Tanggal Lulus:
Pembimbing I
Dr. Ir. Iman Rusmana, M.Si
NIP 196507201991031002
Pembimbing II
Alina Akhdiya, M.Si
NIP 196812082001122001
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga
karya ilmiah ini berhasil diselesaikan. Judul yang dipilih dalam penelitian yang dilaksanakan sejak
bulan November 2010 hingga Mei 2011 ini ialah Isolasi dan Karakterisasi Bakteri Penghasil
AHL-laktonase Asal Lahan Pertanian Luar Pulau Jawa.
Terima kasih penulis ucapkan kepada Bapak Dr. Ir. Iman Rusmana, M.Si dan Ibu Alina
Akhdiya, M.Si atas bimbingan, bantuan pendanaan, dan pengarahan yang diberikan selama
penelitian dan penyusunan skripsi ini. Terima kasih kepada Ibu Dr. Ir. Y. M. Diah Ratnadewi atas
saran dan masukan yang telah diberikan sehingga tulisan ini menjadi lebih baik. Ungkapan terima
kasih juga disampaikan kepada Ayah, Ibu, Ka Isma Haryanti, Ka Iyus Husniyati, Rizqy Rahman,
dan Dede Fina atas segala doa dan kasih sayangnya. Penulis juga mengucapkan terima kasih
kepada Fadhila Achmad S, Astri, Maulina, Yoshita, Syifa, Wika, Dina, Rizki, Ashna, Nisyful,
Komal, Henny, Fahmi, Rahmah, Rindi, Tira, Chintya, Ka Fina, Rani, teman-teman yang
melakukan penelitian di laboratorium Mikrobiologi, Yakub, Nia, Rina, Nita, Hokie, Debie, Susan,
Adian, Aya, Alma, Bang Jo, Ivan, Hana, Vita dan teman-teman yang telah membawakan sampel
tanah, Sepri, Made, Kokom, Adhi, serta seluruh teman-teman Biologi 44 atas segala doa,
dukungan, dan perhatiannya. Serta pihak-pihak yang secara tidak langsung telah membantu dalam
pengumpulan data karya ilmiah ini.
Semoga karya Ilmiah ini bermanfaat.
Bogor, Juli 2011
RIWAYAT HIDUP
Penulis dilahirkan di Indramayu pada tanggal 12 Juli 1988 dari ayah Bahrudin dan Ibu
Suprihatin. Penulis merupakan anak ketiga dari empat bersaudara.
Tahun 2007 penulis lulus dari SMA Negeri 4 Cirebon dan pada tahun yang sama lulus
seleksi masuk IPB melalui Undangan Seleksi Masuk IPB. Penulis memilih program studi Biologi,
Fakultas Matematika dan Ilmu Pengetahuan Alam.
Selama mengikuti perkuliahan, penulis aktif menjadi anggota Divisi Pengembangan
Sumber Daya Manusia (PSDM) Himpunan Mahasiswa Biologi (Himabio) dan anggota Divisi
Kekeluargaan Ikatan Kekeluargaan Cirebon (IKC) pada tahun 2008-2009, Sekretaris Divisi PSDM
Himabio tahun 2009-2010, Ketua Panitia Grand Biodiversity Biologi IPB tahun 2010, Ketua
Divisi Hubungan Masyarakat Workshop dan Diskusi Artikel Ilmiah Populer Biologi tahun 2010,
dan Ketua Divisi Konsumsi Seminar dan Pelatihan PKM tahun 2009. Penulis menjadi asisten
praktikum mata kuliah Biologi Tingkat Persiapan Bersama pada tahun ajaran
2009/2010-2010/2011, mata kuliah Mikrobiologi Dasar pada tahun ajaran 2009/2010-2010/2011, dan mata kuliah Ilmu
Lingkungan pada tahun ajaran 2010/2011.
Penulis melakukan studi lapangan di Kawasan Wana Wisata Cangkuang Sukabumi dengan
judul makalah “Eksplorasi Begonia Sebagai Tanaman Obat di Wana Wisata Cangkuang,
Sukabumi”. Penulis juga melakukan Praktik Lapangan di PT PERTAMINA (PERSERO)
Refinery Unit VI Balongan pada bulan Juli tahun 2010, dengan judul makalah “Evaluasi
Pengolahan Air Limbah dengan Sistem Activated Sludge di PT PERTAMINA (PERSERO)
Refinery Unit VI Balongan Indramayu”. Penulis juga menerima beasiswa PERTAMINA
Foundation tahun 2009-2011.
DAFTAR ISI
Halaman
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... viii
DAFTAR LAMPIRAN ... viii
PENDAHULUAN ... 1
Latar Belakang... 1
Tujuan ... 1
Waktu dan Tempat ... 1
BAHAN DAN METODE ... 1
Pengambilan Contoh Tanah... 1
Isolasi dan Penyimpanan Bakteri ... 2
Bioesai Aktivitas Degradasi AHL ... 2
Isolasi Genom dan Amplifikasi Gen Penyandi AHL-laktonase ... 2
Identifikasi Isolat Pendegradasi AHL ... 2
HASIL ... 3
Isolasi dan Penyimpanan Bakteri ... 3
Bioesai Aktivitas Degradasi AHL ... 3
Amplifikasi Gen Penyandi AHL-laktonase ... 3
Identifikasi Isolat Pendegradasi AHL ... 4
Analisis Filogenetik Gen Pendegradasi AHL ... 4
PEMBAHASAN... 5
SIMPULAN ... 6
SARAN ... 6
DAFTAR PUSTAKA ... 6
DAFTAR TABEL
Halaman
1 Sampel tanah bahan isolasi bakteri... 2
2 Isolat bakteri yang diperoleh dari setiap sampel tanah ... 3
3 Diameter zona degradasi AHL enam isolat terpilih... 3
4 Hasil analisis gen aiiA pada isolat NTT3a dan NTT3e ... 4
5 Hasil analisis gen 16S rRNA isolat-isolat pendegradasi AHL ... 4
DAFTAR GAMBAR
Halaman
1 Zona degradasi AHL oleh supernatan isolat PAD1a (A), BAL2a (B), NTT3e (C),
NTT3a (D), NTT6B6 (E), NTT6B7 (F), dan kontrol negatif (G) ... 3
2 Visualisasi pita DNA hasil amplifikasi gen aiiA pada isolat NTT3a dan NTT3e
(kiri-kanan; ladder 1 kb, NTT3e, dan NTT3a) ... 4
3 Visualisasi pita DNA hasil amplifikasi gen 16S rRNA pada isolat pendegradasi AHL
(kiri-kanan; PAD1a, BAL2a, NTT6B6, NTT6B7, NTT3e, NTT3a, dan ladder 1 kb) ... 4
4 Pohon filogenetik gen aiiA NTT3e dan NTT3a dibandingkan dengan beberapa spesies
penghasil AHL-laktonase dan AHL-acylase menggunakan metode Neighbor Joining
(NJ) dengan boostrap 1000x ... 5
DAFTAR LAMPIRAN
Halaman
1 Komposisi media pertumbuhan bakteri ... 9
2 Hasil pewarnaan Gram isolat-isolat pendegradasi AHL ... 10
3 Hasil pewarnaan endospora isolat NTT6B7 ... 11
4 Hasil sekuen gen aiiA, BLAST-N, dan BLAST-X isolat NTT3a dan NTT3e ... 12
5 Hasil sekuen gen 16S rRNA dan BLAST-N isolat PAD1a, BAL2a, NTT3a, NTT3e,
NTT6B6, dan NTT6B7 ... 15
PENDAHULUAN
Latar Belakang
Quorum sensing (QS) adalah komunikasi
antar sel bakteri yang dimediasi oleh
autoinduser (AI) pada quorum populasi
tertentu untuk mengatur ekspresi gen-gen
tertentu (Rukayadi & Hwang 2009). AI adalah
senyawa berbobot molekul rendah yang
disekresikan bakteri dan diakumulasikan
sehingga AI dikenali sel bakteri lainnya
kemudian diserap kembali. AI bakteri Gram
negatif berbeda dengan Gram positif. Gram
negatif menggunakan N-acyl homoserine
lactone (AHL), sedangkan Gram positif
menggunakan
senyawa
peptida
yang
dihasilkan melalui jalur ATP Binding Cassette
(ABC) (Rukayadi & Hwang 2009). Sistem QS
bekerja pada gen-gen yang berhubungan
dengan perilaku populasi bakteri seperti
pembentukan biofilm, virulensi pada inang,
bioluminescence, dan pembentukan antibiotik
(Gera & Srivastava 2006).
Anti-quorum sensing (anti-QS) merupakan
salah satu alternatif pengendalian penyakit
tanpa mematikan bakteri patogen. Senyawa
anti-QS dapat memblok atau mengacau proses
QS. Senyawa anti-QS dapat dikelompokkan
menjadi tiga jenis, yaitu: (1) senyawa
pendegradasi (degradator) yaitu senyawa yang
dapat mendegradasi AI atau komponen
pengatur QS lainnya dan senyawa ini biasanya
berupa enzim, (2) senyawa antagonis, dan (3)
senyawa kompetitor yaitu senyawa yang dapat
berkompetisi
dengan
AI
membentuk
kompleks dengan protein reseptor (Rukayadi
& Hwang 2009). Proses QS juga dapat
dicegah oleh reaksi laktonolisis (terbukanya
cincin lakton) akibat peningkatan pH >7 di
sekitar sel target. Namun ketika pH kembali
turun (asam) cincin lakton kembali terbentuk
(Rasmussen & Givskov 2006). Degradasi
enzimatik molekul AHL merupakan cara yang
efektif dalam mengcegah proses QS. Ada dua
jenis enzim pendegradasi AHL, yaitu AHL–
acylase dan AHL-laktonase.
AHL-acylase adalah enzim penghidrolisis
ikatan peptida pada rantai acyl molekul AHL
dan menghasilkan asam lemak bebas serta
homoserine lacton. Produk degradasi tersebut
digunakan kembali untuk proses metabolisme
bakteri yaitu sebagai sumber energi, nitrogen,
dan karbon. Beberapa gen yang menyandikan
AHL-acylase telah ditemukan yaitu aiiD dari
Ralstonia eutropha, PvdQ dan QuiP dari
Pseudomonas, AhlM dari Streptomyces sp.,
dan aiiC dari Anabaena sp. PCC7120
(Czajkowski & Jafra 2009).
AHL-laktonase
merusak
proses
QS
melalui hidrolisis enzimatis cincin lakton
molekul AHL. Salah satu gen penyandi
AHL-laktonase adalah gen aiiA. Gen aiiA pertama
kali ditemukan pada Bacillus sp. strain 240B1
(Dong et al. 2000). AHL-laktonase terdiri dari
dua klaster yaitu klaster aiiA pada genus
Bacillus dan Attm pada Agrobacterium
tumefaciens, Arthrobacter sp., dan Klebsiella
pneumonia (Dong & Zhang 2005). Homologi
aiiA kemudian ditemukan pada beberapa
spesies Bacillus lainnya yaitu B. subtilis, B.
cereus, B. mycoides dan beberapa strain dari
B. thuringiensis (Dong et al. 2002).
Kemampuan
suatu
bakteri
dalam
mendegradasi AHL dapat diuji dengan
menggunakan Chromobacterium violaceum
sebagai
bioindikator.
Chromobacterium
violaceum menggunakan proses QS untuk
mengatur ekspresi gen Vio yang menyandikan
pigmen berwarna ungu yang disebut violacein
(August et al. 2000).
Penelitian mengenai QS memiliki potensi
dan manfaat yang sangat besar karena dapat
dijadikan alternatif pengendalian bakteri
patogen yang ramah lingkungan dan tidak
memicu resistensi. Isolasi dan karakterisasi
bakteri penghasil AHL-laktonase dari lahan
pertanian dapat dijadikan sebagai langkah
awal memanfaatkan mekanisme anti-QS
untuk
mengendalikan
bakteri
patogen
terutama pada tanaman.
Tujuan
Penelitian ini bertujuan untuk mengisolasi
dan mengkarakterisasi bakteri penghasil
AHL-laktonase asal lahan pertanian di luar
Pulau Jawa.
Waktu dan Tempat
Penelitian ini dilaksanakan dari bulan
November 2010 sampai Mei 2011 di
Laboratorium
Mikrobiologi,
Departemen
Biologi, FMIPA, IPB.
BAHAN DAN METODE
Pengambilan Contoh Tanah
Contoh tanah diambil dari lahan pertanian
di Nusa Tenggara Timur, Lampung, Papua
Barat, Sumatera Barat, Bali, dan Bangka
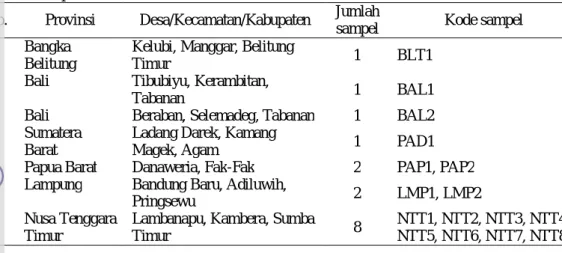
Belitung (Tabel 1).
2
Tabel 1 Sampel tanah bahan isolasi bakteri
No.
Provinsi
Desa/Kecamatan/Kabupaten
Jumlah
sampel
Kode sampel
1.
Bangka
Belitung
Kelubi, Manggar, Belitung
Timur
1
BLT1
2.
Bali
Tibubiyu, Kerambitan,
Tabanan
1
BAL1
3.
Bali
Beraban, Selemadeg, Tabanan
1
BAL2
4.
Sumatera
Barat
Ladang Darek, Kamang
Magek, Agam
1
PAD1
5.
Papua Barat
Danaweria, Fak-Fak
2
PAP1, PAP2
6.
Lampung
Bandung Baru, Adiluwih,
Pringsewu
2
LMP1, LMP2
7.
Nusa Tenggara
Timur
Lambanapu, Kambera, Sumba
Timur
8
NTT1, NTT2, NTT3, NTT4,
NTT5, NTT6, NTT7, NTT8
Isolasi dan Penyimpanan Bakteri
Sebanyak 1 g tanah disuspensikan dengan
garam fisiologis 0.85% kemudian diencerkan
secara serial sampai pengenceran 10
-6, 10
-7,
dan 10
-8. Masing-masing hasil pengenceran
diambil 100 µl dan disebarkan pada media
nutrient agar (NA) (Lampiran 1). Sisa hasil
pengenceran dipanaskan pada suhu 80º C
selama 10 menit lalu diambil sebanyak 100 µl
untuk disebarkan ke media NA yang baru.
Media NA yang telah diinokulasi kemudian
diinkubasi selama 24 jam pada suhu 37º C.
Koloni tunggal yang tumbuh dimurnikan
dengan metode cawan gores pada media
dengan bahan komposisi yang sama. Setelah
didapatkan koloni murni, isolat dipindahkan
pada media NA miring kemudian disimpan
dalam
lemari
pendingin
sampai
saat
digunakan.
Bioesai Aktivitas Degradasi AHL
Sebanyak
satu
lup
kultur
bakteri
diinokulasikan ke dalam 3 ml media luria
broth (LB) (Lampiran 1) lalu diinkubasi pada
suhu ruang. Setelah 16-18 jam, kultur
disentrifugasi (12000 rpm, 10 menit) untuk
diambil supernatannya. Sebanyak 100 µL
supernatan diteteskan pada paper disc di atas
media luria agar (LA) semi-padat (Lampiran
1) yang telah diinokulasikan dengan 1%
kultur
Chromobacterium
violaceum.
Selanjutnya cawan LA tersebut diinkubasi
pada suhu ruang selama 24 jam. Paper disc
yang dikelilingi oleh zona berwarna tidak
ungu menunjukkan adanya aktivitas degradasi
AHL oleh enzim yang dieksresikan oleh isolat
bakteri yang diuji.
Isolasi Genom dan Amplifikasi Gen
Penyandi AHL-laktonase
DNA bakteri diisolasi menggunakan
metode Lazzo (Sambrook & Russell 2001).
DNA total yang diperoleh digunakan sebagai
template untuk amplifikasi gen penyandi
AHL-laktonase menggunakan primer aiiAF
(5’- ATC GGA TCC ATG ACA GTA AAG
AAG CTT TAT TTC G-3’) dan aiiAR
(5’-GTC GAA TTC CTC AAC AAG ATA CTC
CTA ATG ATG T-3’) (Dong et al. 2000).
Campuran reaksi PCR yang digunakan terdiri
dari 32 µl ddH
2O, 5 µl 10x buffer, 3 µl 25
mM MgSO
4, 5 µl dNTPS, 1,5 µl
masing-masing primer, 1 µl template, dan 1 µl DNA
polymerase (KOD hot start). Amplifikasi
dilakukan selama 32 siklus dengan kondisi
pra-denaturasi (94ºC, 10 menit), denaturasi
(94ºC, 30 detik), annealing (52ºC, 30 detik),
elongasi (72ºC, 1 menit), dan post-elongasi
(72ºC, 5 menit) (Chan et al. 2007). Hasil PCR
kemudian dilarikan pada 1% gel elektroforesis
dan diwarnai dengan Ethidium Bromide
(EtBr).
Identifikasi Isolat Pendegradasi AHL
Isolat-isolat yang menunjukkan hasil
positif bioesei degradasi AHL dan amplifikasi
gen penyandi AHL-laktonase diidentifikasi
secara molekuler. Amplifikasi gen 16S rRNA
dilakukan menggunakan primer 63f (5’-CAG
GCC TAA CAC ATG CAA GTC-3’) dan
1387r (5’-GGG CGG WGT GTA CAA
GGC-3’) (Marchesi et al. 1998). Amplifikasi
dilakukan selama 30 siklus dengan kondisi
pra-denaturasi (95ºC, 5 menit), denaturasi
(95ºC, 30 detik), annealing (55ºC, 1 menit),
elongasi (72ºC, 5 menit), dan post-elongasi
(72ºC, 5 menit). Hasil amplifikasi gen 16s
rRNA kemudian dikirim ke perusahaan
penyedia jasa sekuensing.
3
HASIL
Isolasi dan Penyimpanan Bakteri
Sebanyak 58 isolat berhasil diisolasi dan
enam
di
antaranya
memiliki
aktivitas
degradasi AHL (Tabel 2). Keenam isolat
tersebut merupakan bakteri Gram positif
berbentuk
batang.
Lima
di
antaranya
membentuk endospora (Tabel 3) (Lampiran 2
& 3).
Tabel 2 Isolat bakteri yang diperoleh dari
setiap sampel tanah
No.
Kode
sampel
Isolat
yang
diperoleh
Jumlah
isolat positif
bioesei
1.
BLT1
4
-
2.
BAL1
3
-
3.
BAL2
2
1
4.
PAD1
1
1
5.
PAP1
4
-
6.
PAP2
2
-
7.
LMP1
6
-
8.
LMP2
3
-
9.
NTT1
2
-
10.
NTT2
-
-
11.
NTT3
6
2
12.
NTT4
2
-
13.
NTT5
9
-
14.
NTT6
11
2
15.
NTT7
1
-
16.
NTT8
2
-
Total
58
6
Bioesai Aktivitas Degradasi AHL
Isolat PAD1a menunjukan diameter zona
degradasi AHL terbesar yaitu 0.9 cm.
Sedangkan isolat NTT3a, NTT3e, dan
NTT6B7 memiliki diameter zona degradasi
AHL yang paling kecil yaitu sebesar 0.3 cm
(Tabel 3) (Gambar 1).
Amplifikasi Gen Penyandi AHL-laktonase
Hasil amplifikasi gen aiiA menunjukkan
terdapat dua isolat yang mengandung gen
aiiA. Dua isolat tersebut adalah NTT3a dan
NTT3e. Ukuran amplikon 800 bp (Gambar 2).
Hasil analisis BLAST-N dan BLAST-X
menunjukkan gen aiiA isolat NTT3a dan
NTT3e homolog dengan AHL-laktonase
Bacillus sp. 91 (Lampiran 4) (Tabel 4).
Tabel 3 Diameter zona degradasi AHL enam
isolat terpilih
No.
Kode
isolat
Diameter
zona
degradasi
(cm)
Morfologi sel
1.
PAD1a
0.9
Gram positif,
batang, tidak
berendospora
2.
BAL2a
0.4
Gram positif,
batang,
berendospora
3.
NTT3e
0.3
Gram positif,
batang,
berendospora
4.
NTT3a
0.3
Gram positif,
batang,
berendospora
5.
NTT6B6
0.5
Gram positif,
batang,
berendospora
6.
NTT6B7
0.3
Gram positif,
batang,
berendospora
Gambar 1 Zona degradasi AHL oleh
supernatan isolat PAD1a (A),
BAL2a
(B),
NTT3e
(C),
NTT3a (D), NTT6B6 (E),
NTT6B7 (F), dan kontrol
negatif (G).
A
B
D
E
G
F
C
4
Tabel 4 Hasil analisis gen aiiA pada isolat NTT3a dan NTT3e
Kode isolat
Homologi bakteri
(BLAST-N) (a)
Homologi protein (BLAST-X)
(b)
Nilai identitas
NTT3a
Bacillus sp. 91
AHL-lactonase
gene Length= 753
AHL-lactonase [Bacillus sp.
91] Length= 250
724/725 (99%) (a),
239/240 (99%) (b)
NTT3e
Bacillus sp. 91
AHL-lactonase
gene Length= 753
AHL-lactonase [Bacillus sp.
91] Length= 250
724/725 (99%) (a),
239/240 (99%) (b)
Tabel 5 Hasil analisis gen 16S rRNA isolat-isolat pendegradasi AHL
Kode isolat
Homologi bakteri
(BLAST-N)
Nilai identitas
PAD1a
Microbacterium sp. Bg-7 Length=1383
1025/1039 (99%)
BAL2a
Uncultured bacterium clone ncd1261e03c1
Length=1372
609/683 (89%)
NTT3a
Bacillus sp. AF-777 Length=1522
455/471 (99%)
NTT3e
Bacillus sp. NIOT-3 Length=1445
807/814 (97%)
NTT6B6
Bacillus subtilis strain HU48 Length=1427
1147/1149 (99%)
NTT6B7
Bacillus subtilis strain GD1 Length=1452
1025/1028 (99%)
Identifikasi Isolat Pendegradasi AHL
Berdasarkan hasil bioesai dan deteksi
gen aiiA dilakukan identifikasi gen 16S
rRNA terhadap enam isolat yang memiliki
aktivitas
degradasi
AHL.
Amplikon
berukuran 1300 bp (Gambar 3). Analisis gen
16S rRNA menunjukkan isolat NTT3a,
NTT3e, NTT6B6, dan NTT6B7 termasuk
kedalam genus Bacillus, isolat PAD1a mirip
dengan Microbacterium sp. Bg-7, dan isolat
BAL2a mirip dengan Uncultured bacterium
clone ncd1261e03c1 (Lampiran 5) (Tabel 5).
Gambar 3 Visualisasi pita DNA hasil
amplifikasi gen 16S rRNA
pada isolat pendegradasi AHL
(kiri-kanan; PAD1a, BAL2a,
NTT6B6, NTT6B7, NTT3a,
NTT3e, dan ladder 1 kb).
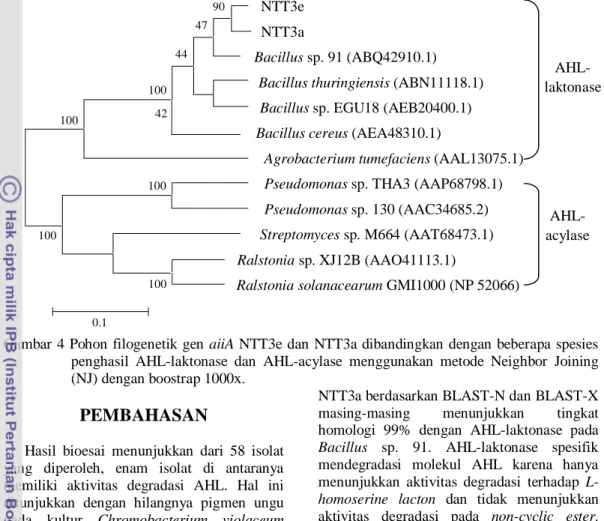
Analisis Filogenetik Gen Pendegradasi
AHL
Analisis filogenetik gen pendegradasi
AHL
menggunakan
metode
Neighbor
Joining (NJ) (Tamura et al. 2007) dengan
boostrap 1000x menunjukkan bahwa isolat
NTT3e dan isolat NTT3a terletak dalam satu
klaster dengan bakteri penghasil
AHL-laktonase Bacillus Sp. 91 dengan nilai
konsistensi boostrap hanya 47%. Pohon
filogenetik tersebut membentuk dua klaster
besar yang terpisah yaitu klaster bakteri
yang menghasilkan AHL-laktonase dan
AHL-acylase (Gambar 4).
1500
1000
750
500
250
Gambar 2 Visualisasi pita DNA hasil
amplifikasi gen aiiA pada isolat
NTT3a dan NTT3e (kiri-kanan;
ladder 1 kb, NTT3e, dan
NTT3a).
250
500
750
1000
5
Gambar 4 Pohon filogenetik gen aiiA NTT3e dan NTT3a dibandingkan dengan beberapa spesies
penghasil AHL-laktonase dan AHL-acylase menggunakan metode Neighbor Joining
(NJ) dengan boostrap 1000x.
PEMBAHASAN
Hasil bioesai menunjukkan dari 58 isolat
yang diperoleh, enam isolat di antaranya
memiliki aktivitas degradasi AHL. Hal ini
ditunjukkan dengan hilangnya pigmen ungu
pada kultur Chromobacterium violaceum
setelah diinkubasi selama 24 jam (Gambar 1).
Isolat
PAD1a
menunjukkan
aktivitas
degradasi terbaik dengan diameter zona
degradasi 0.9 cm. Dibandingkan dengan
diameter zona degradasi AHL empat isolat
lainnya, diameter zona degradasi AHL isolat
NTT3a dan NTT3e relatif lebih kecil.
Walaupun demikian, dari enam isolat yang
menunjukkan hasil positif pada bioesai
degradasi AHL hanya isolat NTT3a dan
NTT3e yang berhasil diamplifikasi gen
aiiAnya. Isolat BAL2a, NTT6B6, PAD1a, dan
NTT6B7
merupakan
isolat-isolat
yang
memiliki aktivitas degradasi AHL namun
tidak menunjukkan produk amplifikasi gen
aiiA. Hal ini dapat disebabkan oleh perbedaan
gen penyandi AHL-laktonase yang dimiliki
oleh isolat-isolat tersebut. Selain
AHL-laktonase
degradasi
AHL
juga
dapat
disebabkan oleh aktivitas AHL-acylase.
Amplifikasi gen aiiA pada isolat NTT3a
dan NTT3e menunjukkan amplikon yang
berukuran 800 bp (Gambar 2), hal ini sesuai
dengan penelitian Chan et al. (2007) yang
membuktikan bahwa amplikon hasil PCR gen
aiiA pada bakteri tanah sebesar 800 bp.
Analisis sekuen gen aiiA isolat NTT3e dan
NTT3a berdasarkan BLAST-N dan BLAST-X
masing-masing
menunjukkan
tingkat
homologi 99% dengan AHL-laktonase pada
Bacillus sp. 91. AHL-laktonase spesifik
mendegradasi molekul AHL karena hanya
menunjukkan aktivitas degradasi terhadap
L-homoserine lacton dan tidak menunjukkan
aktivitas degradasi pada non-cyclic ester.
Enzim ini aktivitasnya stabil pada suhu di
bawah 37ºC tetapi tidak stabil pada suhu yang
tinggi (Wang et al. 2004).
Amplifikasi gen 16S rRNA isolat PAD1a,
BAL2a, NTT3e, NTT3a, NTT6B6, dan
NTT6B7 menghasilkan amplikon berukuran
1300 bp (Gambar 2). Analisis gen 16S rRNA
isolat PAD1a menunjukkan kemiripan dengan
Microbacterium sp. Bg-7 (nilai identitas
99%). Wang et al. (2010) juga melaporkan
adanya
aktivitas
degradasi
AHL
pada
Microbacterium testaceum StLB037 yang
diisolasi
dari
daun
kentang.
Aktivitas
degradasi AHL pada bakteri ini mirip dengan
aktivitas degradasi AHL pada beberapa
bakteri yang memiliki AHL-laktonase seperti
Bacillus sp., Arthrobacter sp., Agrobacterium
tumefaciens, dan Rhodococcus erythropolis.
AHL-laktonase
pada
Microbacterium
testaceum StLB037 disandikan oleh gen aiiM.
Hasil analisis 16S rRNA menunjukkan isolat
BAL2a mirip dengan Uncultured bacterium
clone ncd1261e03c1 (nilai identitas 89%),
isolat NTT6B6 mirip dengan Bacillus subtilis
strain HU48 (nilai identitas 99%). Hasil
analisis 16S rRNA isolat NTT6B7 mirip
dengan Bacillus subtilis strain GD1 (nilai
NTT3e
NTT3a
Bacillus sp. 91 (ABQ42910.1)
Bacillus thuringiensis (ABN11118.1)
Bacillus sp. EGU18 (AEB20400.1)
Bacillus cereus (AEA48310.1)
Agrobacterium tumefaciens (AAL13075.1)
Pseudomonas sp. THA3 (AAP68798.1)
Pseudomonas sp. 130 (AAC34685.2)
Streptomyces sp. M664 (AAT68473.1)
Ralstonia sp. XJ12B (AAO41113.1)
Ralstonia solanacearum GMI1000 (NP 52066)
100 100 100 100 100 47 44 42 0.1 90
AHL-laktonase
AHL-acylase
6
identitas 99%), isolat NTT3e mirip dengan
Bacillus sp. NIOT-3 (nilai identitas 99%), dan
isolat NTT3a mirip dengan Bacillus sp.
AF-777 (nilai identitas 97%). Menurut Dong et al.
(2002), genus Bacillus sebagian besar
memiliki gen aiiA yang menyandikan
AHL-laktonase. Chan et al. (2007) juga melaporkan
bahwa sebanyak 12 isolat dari 50 isolat
Bacillus yang diisolasi dari tanah tropis
memiliki gen yang homolog dengan gen aiiA.
Analisis filogenetik gen pendegradasi AHL
menggunakan metode Neighbor Joining (NJ)
dengan boostrap 1000x menunjukkan bahwa
isolat NTT3a dan isolat NTT3e terletak dalam
satu klaster dengan bakteri penghasil
AHL-laktonase Bacillus Sp. 91 (Gambar 4). Nilai
konsistensi boostrap antara NTT3a dan
NTT3e mencapai 90% yang menunjukkan
bahwa gen aiiA pada kedua isolat ini sangat
mirip. Pohon filogenetik tersebut membentuk
dua klaster besar yang terpisah yaitu klaster
bakteri yang menghasilkan AHL-laktonase
dan acylase. Sekuen enzim
AHL-laktonase terdistribusi luas pada beberapa
organisme meliputi Actinobacteria,
Acido-bacteria, Bacteroidetes, Chloroflexi,
Deino-coccus-Thermus, Firmicutes,
α-Proteobac-teria, β-Proteobacα-Proteobac-teria, γ-Proteobacα-Proteobac-teria,
δ-Proteobacteria, Euryarchaeota, Crenarchaeota,
Sphingobacteria, Spirochaetales,
Nitrospira-les, and Planctomycetes. Namun hanya dua
kelompok yaitu Firmicutes (12 genus dan 22
spp.) dan α-Proteobacteria (18 genus dan 22
spp.) yang memiliki aktivitas AHL-laktonase.
Distribusi sekuen AHL-acylase lebih sempit
dibandingkan
sekuen
AHL-laktonase.
Distribusi
sekuen
enzim
AHL-acylase
meliputi Actinobacteria, Cyanobacteria,
Bac-teroidetes, Deinococcus-Thermus, Firmicutes,
α-Proteobacteria, β-Proteobacteria,
γ-Proteo-bacteria, δ-Proteoγ-Proteo-bacteria, Euryarchaeota dan
Crenarchaeota.
Distribusi
sekuen
acylase sangat berbeda dengan sekuen
AHL-laktonase.
γ-Proteobacteria
merupakan
kelompok terbesar yang mengandung aktivitas
AHL-acylase (20 genus dan 31 spp.) (Kalia et
al. 2011).
SIMPULAN
Isolat NTT3e dan NTT3a memiliki gen
aiiA yang homolog dengan AHL-laktonase
pada Bacillus sp. 91. Gen 16S rRNA isolat
NTT3e mirip dengan Bacillus sp. NIOT-3
(nilai identitas: 99%). Sedangkan isolat
NTT3a mirip dengan Bacillus sp. AF-777
(nilai identitas: 97%). Isolat PAD1a, BAL2a,
NTT6B6, dan NTT6B7 memiliki aktivitas
degradasi
AHL
namun
tidak
berhasil
diamplifikasi
gen
aiiAnya.
Berdasarkan
analisis gen 16S rRNA, isolat PAD1a mirip
dengan Microbacterium sp. Bg-7, BAL2a
mirip dengan Uncultured bacterium clone
ncd1261e03c1,
NTT6B6
mirip
dengan
Bacillus subtilis strain HU48, dan NTT6B7
mirip dengan Bacillus subtilis strain GD1.
SARAN
Perlu dilakukan karakterisasi molekuler
gen pendegradasi AHL pada isolat yang
memiliki aktivitas degaradasi AHL namun
tidak berhasil diamplifikasi gen aiiAnya
menggunakan primer aiiAF dan aiiAR.
DAFTAR PUSTAKA
August PR, Grossman TH, Minor C, Draper
MP, MacNeil IA, Pemberton JM, Call
KM, Holt D, Osburne S. 2000. Sequence
analysis and functional characterization of
the violacein biosynthetic pathway from
Chromobacterium
violaceum.
J
Mol
Microbiol Biotechnol 2: 513-519.
Chan KG, Tiew SZ. Ng CC. 2007. Rapid
isolation method of soil bacilli and
screening of their quorum quenching
activity. Asia Pacific J Mol Biol and
Biotechnol 15: 153-156.
Czajkowski R, Jafra S. 2009. Quenching of
acyl-homoserine
lactone-dependent
quorum sensing by enzymatic disruption
of signal molecules. Acta Biochim
Polonica 56: 1-16.
Dong YH, Xu JL, Li XC, Zang LH. 2000.
aiiA, a novel enzyme inactivates acyl
homoserine-lactone quorum-sensing signal
and attenuates the virulence of Erwinia
carotovora. Proc Natl Acad Sci 97:3526–
3531.
Dong YH, Gusti AR, Zhang Q, Xu JL, Zhang
LH. 2002. Identification of
quorum-quenching N-acyl homoserine lactonases
from Bacillus species. Appl Environ
Microbiol 68:1754–1759.
Dong YH, Zhang LH. 2005. Quorum sensing
and
quorum-quenching
enzymes.
J
Microbiol 43: 101-109.
Gera C, Srivastava S. 2006. Quorum sensing:
the
phenomenon
of
microbial
communication. Current Sci 90: 666-675.
Kalia VC, Raju SC, Purohit HJ. 2011.
Genomic
analysis
reveals
versatile
organisms
for
quorum
quenching
7
enzymes: acyl-homoserine lactone-acylase
and –lactonase. Open Microbiol J 5: 1-13.
Marchesi JR, Sato T, Weightman AJ, Martin
TA, Fry JC, Hiom SJ, Wade WG. 1998.
Design
and
evaluation
of
useful
bacterium-specific
PCR
primer
that
amplify genes coding for bacterial 16S
rRNA. Appl Environ Microbiol 64:
795-799.
Rasmussen TB, Givskov M. 2006. Quorum
sensing inhibitors: a bargain of effects.
Microbiology 152: 895-904.
Rukayadi Y, Hwang JK. 2009. Pencegahan
quorum sensing: suatu pendekatan baru
dalam
mengatasi
infeksi
bakteri.
Medicinus 22: 22-27.
Sambrook J, Russell DW. 2001. Molecular
Cloning: a Laboratory Manual Third
Edition. New York: Cold Spring Harbor
Laboratory Pr.
Tamura K, Dudley J, Nei M, Kumar S. 2007.
MEGA4: molecular evolutionary genetics
analysis (MEGA) software version 4.0.
Mol Biol Evol 24: 1596-1599.
Wang LH, Weng LX, Dong YH, Zhang LH.
2004. Specifity and enzyme kinetics of the
quorum-quenching N-acyl homoserine
lactone lactonase (AHL-lactonase). J Biol
Chem 279: 13645-13651.
Wang WZ, Morohoshi T, Ikenoya M, Someya
N, Ikeda T. 2010. AiiM, a novel class of
N-acylhomoserine lactonase from the
leaf-associated
bacterium
Microbacterium
testaceum. Appl and Environ Microbiol
76: 2524-2530.
9
Lampiran 1 Komposisi media pertumbuhan bakteri
1. Nutrient Agar (NA)
0.8 g nutrient broth (NB)
1.5 g agar
100 ml akuades
2. Luria Broth (LB)
1 g Trypton
1 g NaCl
0.5 g yeast extract
100 ml akuades
3. Luria Agar (LA) untuk pertumbuhan Chromobacterium violaceum
1 g Trypton
1 g NaCl
0.5 g yeast extract
0.1 g CaCO
3 1.5 g agar
100 ml akuades
4. Luria Agar (LA) semi padat
1 g Trypton
1 g NaCl
0.5 g yeast extract
0.1 g CaCO
3 1 g agar
100 ml akuades
10
Lampiran 2 Hasil pewarnaan Gram isolat-isolat pendegradasi AHL
PAD1a
(perbesaran 100 x 10)
BAL2a
(perbesaran 100 x 10)
NTT6B6
(perbesaran 100 x 10)
NTT6B7
(perbesaran 100 x 10)
NTT3e
(perbesaran 100 x 10)
NTT3a
(perbesaran 100 x 10)
11
Lampiran 3 Hasil pewarnaan endospora isolat NTT6B7
Keterangan: bagian yang ditunjuk panah adalah endospora (gelap dan berbentuk bulat di ujung sel)
sedangkan sel vegetatifnya berwarna merah muda (perbesaran 100 x 10).
12
Lampiran 4 Hasil sekuen gen aiiA, BLAST-N, dan BLAST-X isolat NTT3a dan NTT3e
A) Urutan nukleotida gen aiiA isolat NTT3a
TTTTCAGCAGGTCGTTGTATGTTAGATCATTCTTCTGTTAATAGTACACTCGCGCCGGGGAATTTATTGAA CTTACCTGTATGGTGTTATCTTTTGGAGACAGAAGAGGGGCCTATTTTAGTAGATACAGGTATGCCAGAA AGTGCAGTTAATAATGAAGGGCTTTTTAACGGTACATTTGTTGAAGGACAGATTTTACCGAAAATGACTG AAGAAGATAGAATCGTGAATATATTAAAACGTGTAGGGTATGAGCCGGACGACCTTTTATATATTATTAG TTCTCACTTACATTTTGATCATGCAGGAGGAAACGGTGCTTTTACAAATACACCGATTATTGTGCAACGA ACGGAATATGAGGCAGCACTTCATAGAGAAGAATATATGAAAGAATGTATATTACCGCATTTGAACTAC AAAATTATTGAAGGGGATTATGAAGTGGTACTAGGTGTGCAATTATTGTATACGCCAGGCCATTCTCCAG GCCATCAGTCGCTATTAATTGAGACAGAAAAATCCGGTCCTGTATTATTAACGATTGATGCATCTTATAC GAAAGAAAATTTTGAAGATGAAGTGCCGTTCGCGGGATTTGATTCGGAATTAGCTTTATCTTCCATTAAA CGTTTAAAAGAAGTTGTGGCGAAAGAGAAACCAATTATTTTCTTTGGTCATGATATAGAGCAGGAAAAG GGTTGTAAAGTGTTCCCGGAATATATATAGTGCAAAAAGTCATGAGCTTACGTGCTCATGACTTTTTCGT TTAAATAATTTTTTTGAATAAGTTATAAACCTTTTTTGAGCTATCTTCATTTAATTGATAGTACTTGAGGTT TACATCATTAGGAGTATCTTGTTGAGAAAATTCGACAAAANNNAAAAAAGAA
B) Hasil BLAST-N sekuen gen aiiA isolat NTT3a dengan data Genbank
|EF537015.1| Bacillus sp. 91 AHL-lactonase gene, complete cds
Length = 753; Score = 1334 bits (722); Expect = 0.0; Identities = 724/725 (99%); Gaps = 0/725
(0%); Strand = Plus/Plus
Query 6 CAGCAGGTCGTTGTATGTTAGATCATTCTTCTGTTAATAGTACACTCGCGCCGGGGAATT 65 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 29 CAGCAGGTCGTTGTATGTTAGATCATTCTTCTGTTAATAGTACACTCGCGCCGGGGAATT 88 Query 66 TATTGAACTTACCTGTATGGTGTTATCTTTTGGAGACAGAAGAGGGGCCTATTTTAGTAG 125 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 89 TATTGAACTTACCTGTATGGTGTTATCTTTTGGAGACAGAAGAGGGGCCTATTTTAGTAG 148 Query 126 ATACAGGTATGCCAGAAAGTGCAGTTAATAATGAAGGGCTTTTTAACGGTACATTTGTTG 185 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 149 ATACAGGTATGCCAGAAAGTGCAGTTAATAATGAAGGGCTTTTTAACGGTACATTTGTTG 208 Query 186 AAGGACAGATTTTACCGAAAATGACTGAAGAAGATAGAATCGTGAATATATTAAAACGTG 245 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 209 AAGGACAGATTTTACCGAAAATGACTGAAGAAGATAGAATCGTGAATATATTAAAACGTG 268 Query 246 TAGGGTATGAGCCGGACGACCTTTTATATATTATTAGTTCTCACTTACATTTTGATCATG 305 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 269 TAGGGTATGAGCCGGACGACCTTTTATATATTATTAGTTCTCACTTACATTTTGATCATG 328 Query 306 CAGGAGGAAACGGTGCTTTTACAAATACACCGATTATTGTGCAACGAACGGAATATGAGG 365 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 329 CAGGAGGAAACGGTGCTTTTACAAATACACCGATTATTGTGCAACGAACGGAATATGAGG 388 Query 366 CAGCACTTCATAGAGAAGAATATATGAAAGAATGTATATTACCGCATTTGAACTACAAAA 425 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 389 CAGCACTTCATAGAGAAGAATATATGAAAGAATGTATATTACCGCATTTGAACTACAAAA 448 Query 426 TTATTGAAGGGGATTATGAAGTGGTACTAGGTGTGCAATTATTGTATACGCCAGGCCATT 485 ||||||||||||||||||||||||||| |||||||||||||||||||||||||||||||| Sbjct 449 TTATTGAAGGGGATTATGAAGTGGTACCAGGTGTGCAATTATTGTATACGCCAGGCCATT 508 Query 486 CTCCAGGCCATCAGTCGCTATTAATTGAGACAGAAAAATCCGGTCCTGTATTATTAACGA 545 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 509 CTCCAGGCCATCAGTCGCTATTAATTGAGACAGAAAAATCCGGTCCTGTATTATTAACGA 568 Query 546 TTGATGCATCTTATACGAAAGAAAATTTTGAAGATGAAGTGCCGTTCGCGGGATTTGATT 605 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 569 TTGATGCATCTTATACGAAAGAAAATTTTGAAGATGAAGTGCCGTTCGCGGGATTTGATT 628 Query 606 CGGAATTAGCTTTATCTTCCATTAAACGTTTAAAAGAAGTTGTGGCGAAAGAGAAACCAA 665 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 629 CGGAATTAGCTTTATCTTCCATTAAACGTTTAAAAGAAGTTGTGGCGAAAGAGAAACCAA 688 Query 666 TTATTTTCTTTGGTCATGATATAGAGCAGGAAAAGGGTTGTAAAGTGTTCCCGGAATATA 725 ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||13
Sbjct 689 TTATTTTCTTTGGTCATGATATAGAGCAGGAAAAGGGTTGTAAAGTGTTCCCGGAATATA 748 Query 726 TATAG 730
|||||
Sbjct 749 TATAG 753
C) Hasil BLAST-X sekuen gen aiiA isolat NTT3a dengan data Genbank
|ABQ42910.1| AHL-lactonase [Bacillus sp. 91]
Length = 250; Score = 488 bits (1256); Expect = 5e-136; Identities = 239/240 (99%); Positives
= 239/240 (99%); Gaps = 0/240 (0%); Frame = +1
Query 8 AGRCMLDHSSVNSTLAPGNLLNLPVWCYLLETEEGPILVDTGMPESAVNNEGLFNGTFVE 187 AGRCMLDHSSVNSTLAPGNLLNLPVWCYLLETEEGPILVDTGMPESAVNNEGLFNGTFVE Sbjct 11 AGRCMLDHSSVNSTLAPGNLLNLPVWCYLLETEEGPILVDTGMPESAVNNEGLFNGTFVE 70 Query 188 GQILPKMTEEDRIVNILKRVGYEPDDLLYIISSHLHFDHAGGNGAFTNTPIIVQRTEYEA 367 GQILPKMTEEDRIVNILKRVGYEPDDLLYIISSHLHFDHAGGNGAFTNTPIIVQRTEYEA Sbjct 71 GQILPKMTEEDRIVNILKRVGYEPDDLLYIISSHLHFDHAGGNGAFTNTPIIVQRTEYEA 130 Query 368 ALHREEYMKECILPHLNYKIIEGDYEVVLGVQLLYTPGHSPGHQSLLIETEKSGPVLLTI 547 ALHREEYMKECILPHLNYKIIEGDYEVV GVQLLYTPGHSPGHQSLLIETEKSGPVLLTI Sbjct 131 ALHREEYMKECILPHLNYKIIEGDYEVVPGVQLLYTPGHSPGHQSLLIETEKSGPVLLTI 190 Query 548 DASYTKENFEDEVPFAGFDSELALSSIKRLKEVVAKEKPIIFFGHDIEQEKGCKVFPEYI 727 DASYTKENFEDEVPFAGFDSELALSSIKRLKEVVAKEKPIIFFGHDIEQEKGCKVFPEYI Sbjct 191 DASYTKENFEDEVPFAGFDSELALSSIKRLKEVVAKEKPIIFFGHDIEQEKGCKVFPEYI 250D) Urutan nukleotida gen aiiA isolat NTT3e
TTTCAGCAGGTCGTTGTATGTTAGATCATTCTTCTGTTAATAGTACACTCGCGCCGGGGAATTTATTGAAC TTACCTGTATGGTGTTATCTTTTGGAGACAGAAGAGGGGCCTATTTTAGTAGATACAGGTATGCCAGAAA GTGCAGTTAATAATGAAGGGCTTTTTAACGGTACATTTGTTGAAGGACAGATTTTACCGAAAATGACTGA AGAAGATAGAATCGTGAATATATTAAAACGTGTAGGGTATGAGCCGGACGACCTTTTATATATTATTAGT TCTCACTTACATTTTGATCATGCAGGAGGAAACGGTGCTTTTACAAATACACCGATTATTGTGCAACGAA CGGAATATGAGGCAGCACTTCATAGAGAAGAATATATGAAAGAATGTATATTACCGCATTTGAACTACA AAATTATTGAAGGGGATTATGAAGTGGTACTAGGTGTGCAATTATTGTATACGCCAGGCCATTCTCCAGG CCATCAGTCGCTATTAATTGAGACAGAAAAATCCGGTCCTGTATTATTAACGATTGATGCATCTTATACG AAAGAAAATTTTGAAGATGAAGTGCCGTTCGCGGGATTTGATTCGGAATTAGCTTTATCTTCCATTAAAC GTTTAAAAGAAGTTGTGGCGAAAGAGAAACCAATTATTTTCTTTGGTCATGATATAGAGCAGGAAAAGG GTTGTAAAGTGTTCCCGGAATATATATAGTGCAAAAAGTCATGAGCTTACGTGCTCATGACTTTTTCGTTT AAATAATTTTTTTGAATAAGTTATAAACCTTTTTTGAGCTATCTTCATTTAATTGATAGTACTTGAGGTTT ACATCATTAGGAGTATCTTGTTGAGGAATTCAACAAAAAAAATATGCGNTAGATGTTAGCGTTCTGGTTG GAAATCCGCATCAATAGTCGGTACATCTTCACCGACTAGNACGTGTAGC
E) Hasil BLAST-N sekuen gen aiiA isolat NTT3e dengan data Genbank
|EF537015.1| Bacillus sp. 91 AHL-lactonase gene, complete cds
Length = 753; Score = 1334 bits (722); Expect = 0.0; Identities = 724/725 (99%); Gaps = 0/725
(0%); Strand=Plus/Plus
Query 4 CAGCAGGTCGTTGTATGTTAGATCATTCTTCTGTTAATAGTACACTCGCGCCGGGGAATT 63 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 29 CAGCAGGTCGTTGTATGTTAGATCATTCTTCTGTTAATAGTACACTCGCGCCGGGGAATT 88 Query 64 TATTGAACTTACCTGTATGGTGTTATCTTTTGGAGACAGAAGAGGGGCCTATTTTAGTAG 123 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 89 TATTGAACTTACCTGTATGGTGTTATCTTTTGGAGACAGAAGAGGGGCCTATTTTAGTAG 148 Query 124 ATACAGGTATGCCAGAAAGTGCAGTTAATAATGAAGGGCTTTTTAACGGTACATTTGTTG 183 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 149 ATACAGGTATGCCAGAAAGTGCAGTTAATAATGAAGGGCTTTTTAACGGTACATTTGTTG 208 Query 184 AAGGACAGATTTTACCGAAAATGACTGAAGAAGATAGAATCGTGAATATATTAAAACGTG 243 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 209 AAGGACAGATTTTACCGAAAATGACTGAAGAAGATAGAATCGTGAATATATTAAAACGTG 268 Query 244 TAGGGTATGAGCCGGACGACCTTTTATATATTATTAGTTCTCACTTACATTTTGATCATG 303 ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||14
Sbjct 269 TAGGGTATGAGCCGGACGACCTTTTATATATTATTAGTTCTCACTTACATTTTGATCATG 328 Query 304 CAGGAGGAAACGGTGCTTTTACAAATACACCGATTATTGTGCAACGAACGGAATATGAGG 363 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 329 CAGGAGGAAACGGTGCTTTTACAAATACACCGATTATTGTGCAACGAACGGAATATGAGG 388 Query 364 CAGCACTTCATAGAGAAGAATATATGAAAGAATGTATATTACCGCATTTGAACTACAAAA 423 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 389 CAGCACTTCATAGAGAAGAATATATGAAAGAATGTATATTACCGCATTTGAACTACAAAA 448 Query 424 TTATTGAAGGGGATTATGAAGTGGTACTAGGTGTGCAATTATTGTATACGCCAGGCCATT 483 ||||||||||||||||||||||||||| |||||||||||||||||||||||||||||||| Sbjct 449 TTATTGAAGGGGATTATGAAGTGGTACCAGGTGTGCAATTATTGTATACGCCAGGCCATT 508 Query 484 CTCCAGGCCATCAGTCGCTATTAATTGAGACAGAAAAATCCGGTCCTGTATTATTAACGA 543 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 509 CTCCAGGCCATCAGTCGCTATTAATTGAGACAGAAAAATCCGGTCCTGTATTATTAACGA 568 Query 544 TTGATGCATCTTATACGAAAGAAAATTTTGAAGATGAAGTGCCGTTCGCGGGATTTGATT 603 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 569 TTGATGCATCTTATACGAAAGAAAATTTTGAAGATGAAGTGCCGTTCGCGGGATTTGATT 628 Query 604 CGGAATTAGCTTTATCTTCCATTAAACGTTTAAAAGAAGTTGTGGCGAAAGAGAAACCAA 663 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 629 CGGAATTAGCTTTATCTTCCATTAAACGTTTAAAAGAAGTTGTGGCGAAAGAGAAACCAA 688 Query 664 TTATTTTCTTTGGTCATGATATAGAGCAGGAAAAGGGTTGTAAAGTGTTCCCGGAATATA 723 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 689 TTATTTTCTTTGGTCATGATATAGAGCAGGAAAAGGGTTGTAAAGTGTTCCCGGAATATA 748 Query 724 TATAG 728 ||||| Sbjct 749 TATAG 753F) Hasil BLAST-X sekuen gen aiiA isolat NTT3e dengan data Genbank
|ABQ42910.1| AHL-lactonase [Bacillus sp. 91]
Length = 250; Score = 488 bits (1256); Expect = 6e-136; Identities = 239/240 (99%); Positives
= 239/240 (99%); Gaps = 0/240 (0%); Frame = +3
Query 6 AGRCMLDHSSVNSTLAPGNLLNLPVWCYLLETEEGPILVDTGMPESAVNNEGLFNGTFVE 185 AGRCMLDHSSVNSTLAPGNLLNLPVWCYLLETEEGPILVDTGMPESAVNNEGLFNGTFVE Sbjct 11 AGRCMLDHSSVNSTLAPGNLLNLPVWCYLLETEEGPILVDTGMPESAVNNEGLFNGTFVE 70 Query 186 GQILPKMTEEDRIVNILKRVGYEPDDLLYIISSHLHFDHAGGNGAFTNTPIIVQRTEYEA 365 GQILPKMTEEDRIVNILKRVGYEPDDLLYIISSHLHFDHAGGNGAFTNTPIIVQRTEYEA Sbjct 71 GQILPKMTEEDRIVNILKRVGYEPDDLLYIISSHLHFDHAGGNGAFTNTPIIVQRTEYEA 130 Query 366 ALHREEYMKECILPHLNYKIIEGDYEVVLGVQLLYTPGHSPGHQSLLIETEKSGPVLLTI 545 ALHREEYMKECILPHLNYKIIEGDYEVV GVQLLYTPGHSPGHQSLLIETEKSGPVLLTI Sbjct 131 ALHREEYMKECILPHLNYKIIEGDYEVVPGVQLLYTPGHSPGHQSLLIETEKSGPVLLTI 190 Query 546 DASYTKENFEDEVPFAGFDSELALSSIKRLKEVVAKEKPIIFFGHDIEQEKGCKVFPEYI 725 DASYTKENFEDEVPFAGFDSELALSSIKRLKEVVAKEKPIIFFGHDIEQEKGCKVFPEYI Sbjct 191 DASYTKENFEDEVPFAGFDSELALSSIKRLKEVVAKEKPIIFFGHDIEQEKGCKVFPEYI 25015
Lampiran 5 Hasil sekuen gen 16S rRNA dan BLAST-N isolat PAD1a, BAL2a, NTT3a, NTT3e,
NTT6B6, dan NTT6B7
A) Urutan nukleotida gen 16S rRNA isolat PAD1a
TTTGGAAAGATTTTTCGGTTGGGGATGGGCTCGCGGCCTATCAGCTTGTTGGTGAGGTAATGGCTCACCA AGGCGTCGACGGGTAGCCGGCCTGAGAGGGTGACCGGCCACACTGGGACTGAGACACGGCCCAGACTCC TACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGAAAGCCTGATGCAGCAACGCCGCGTGAGGG ATGACGGCCTTCGGGTTGTAAACCTCTTTTAGCAGGGAAGAAGCGAAAGTGACGGTACCTGCAGAAAAA GCGCCGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGGCGCAAGCGTTATCCGGAATTATTGGG CGTAAAGAGCTCGTACGCGGTTTGTCGCGTCTGCTGTGAAATCCCGAGGCTCAACCTCAGGCCTGCAGTG AGTACGGGCAGACTAGAGTGCGGTAGGGGAGATTGGAATTCCTGGTGTAGCGGTGGAATGCGCAGATAT CAGGAGGAACACCGATGGCGAAGGCAGATCTCTGGGCCGTAACTGACGCTGAGGAGCGAAAGGGTGGG GAGCAAACAGGCTTAGATACCCTGGTAGTCCACCCCGTAAACGTTGGGAACTAGTTGTGGGGTCCATTCC ACGGATTCCGTGACGCAGCTAACGCATTAAGTTCCCCGCCTGGGGAGTACGGCCGCAAGGCTAAAACTC AAAGGAATTGACGGGGACCCGCACAAGCGGCGGAGCATGCGGATTAATTCGATGCAACGCGAAGAACC TTACCAAGGCTTGACATATACGAGAACGGGCCAGAAATGGTCAACTCTTTGGACACTCGTAAACAGGTG GTGCATGGTTGTCATCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGAGCAACCCTCGTTC TATGTTGCCAGCACGTAATGGTGGGGAACTCATGGGATACTGCCCGGGTCCACTTCGAAGGAAGGTGGG GATGACGTCAAATCATCATGCCCCTTAAGGTCTTGGGCTTCAAGCATGCTACAATGGCCAGGTACAAAGG GC
B) Hasil BLAST-N sekuen gen 16S rRNA isolat PAD1a dengan data Genbank
|HQ916747.1| Microbacterium sp. Bg-7 16S ribosomal RNA gene, partial sequence
Length = 1383; Score = 1838 bits (995); Expect = 0.0; Identities = 1025/1039 (99%); Gaps =
4/1039 (0%); Strand = Plus/Plus
Query 1 TTTGGAAAGATTTTTCGGTTGGGGATGGGCTCGCGGCCTATCAGCTTGTTGGTGAGGTAA 60 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 145 TTTGGAAAGATTTTTCGGTTGGGGATGGGCTCGCGGCCTATCAGCTTGTTGGTGAGGTAA 204 Query 61 TGGCTCACCAAGGCGTCGACGGGTAGCCGGCCTGAGAGGGTGACCGGCCACACTGGGACT 120 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 205 TGGCTCACCAAGGCGTCGACGGGTAGCCGGCCTGAGAGGGTGACCGGCCACACTGGGACT 264 Query 121 GAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGAAA 180 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 265 GAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGAAA 324 Query 181 GCCTGATGCAGCAACGCCGCGTGAGGGATGACGGCCTTCGGGTTGTAAACCTCTTTTAGC 240 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 325 GCCTGATGCAGCAACGCCGCGTGAGGGATGACGGCCTTCGGGTTGTAAACCTCTTTTAGC 384 Query 241 AGGGAAGAAGCGAAAGTGACGGTACCTGCAGAAAAAGCGCCGGCTAACTACGTGCCAGCA 300 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 385 AGGGAAGAAGCGAAAGTGACGGTACCTGCAGAAAAAGCGCCGGCTAACTACGTGCCAGCA 444 Query 301 GCCGCGGTAATACGTAGGGCGCAAGCGTTATCCGGAATTATTGGGCGTAAAGAGCTCGTA 360 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 445 GCCGCGGTAATACGTAGGGCGCAAGCGTTATCCGGAATTATTGGGCGTAAAGAGCTCGTA 504 Query 361 CGCGGTTTGTCGCGTCTGCTGTGAAATCCCGAGGCTCAACCTCAGGCCTGCAGTGAGTAC 420 |||||||||||||||||||||||||||||||||||||||||| ||||||||||| |||| Sbjct 505 GGCGGTTTGTCGCGTCTGCTGTGAAATCCCGAGGCTCAACCTCGGGCCTGCAGTGGGTAC 564 Query 421 GGGCAGACTAGAGTGCGGTAGGGGAGATTGGAATTCCTGGTGTAGCGGTGGAATGCGCAG 480 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 565 GGGCAGACTAGAGTGCGGTAGGGGAGATTGGAATTCCTGGTGTAGCGGTGGAATGCGCAG 624 Query 481 ATATCAGGAGGAACACCGATGGCGAAGGCAGATCTCTGGGCCGTAACTGACGCTGAGGAG 540 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 625 ATATCAGGAGGAACACCGATGGCGAAGGCAGATCTCTGGGCCGTAACTGACGCTGAGGAG 684 Query 541 CGAAAGGGTGGGGAGCAAACAGGCTTAGATACCCTGGTAGTCCACCCCGTAAACGTTGGG 600 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 685 CGAAAGGGTGGGGAGCAAACAGGCTTAGATACCCTGGTAGTCCACCCCGTAAACGTTGGG 74416
Query 601 AACTAGTTGTGGGGTCCATTCCACGGATTCCGTGACGCAGCTAACGCATTAAGTTCCCCG 660 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 745 AACTAGTTGTGGGGTCCATTCCACGGATTCCGTGACGCAGCTAACGCATTAAGTTCCCCG 804 Query 661 CCTGGGGAGTACGGCCGCAAGGCTAAAACTCAAAGGAATTGACGGGGACCCGCACAAGCG 720 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 805 CCTGGGGAGTACGGCCGCAAGGCTAAAACTCAAAGGAATTGACGGGGACCCGCACAAGCG 864 Query 721 GCGGAGCATGCGGATTAATTCGATGCAACGCGAAGAACCTTACCAAGGCTTGACATATAC 780 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 865 GCGGAGCATGCGGATTAATTCGATGCAACGCGAAGAACCTTACCAAGGCTTGACATATAC 924 Query 781 GAGAACGGGCCAGAAATGGTCAACTCTTTGGACACTCGTAAACAGGTGGTGCATGGTTGT 840 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 925 GAGAACGGGCCAGAAATGGTCAACTCTTTGGACACTCGTAAACAGGTGGTGCATGGTTGT 984 Query 841 CATCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGAGCAACCCTCGTTCTA 900 | |||||||||||||||||||||||||||||||||||||||||| ||||||||||||||| Sbjct 985 CGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTCGTTCTA 1044 Query 901 TGTTGCCAGCACGTAATGGTGGGGAACTCATGGGATACTGCCCGGGTCCACTTCGAAGGA 960 ||||||||||||||||||||||| |||||||||||||||||| ||||| ||| || |||| Sbjct 1045 TGTTGCCAGCACGTAATGGTGGG-AACTCATGGGATACTGCCGGGGTCAACT-CGGAGGA 1102 Query 961 AGGTGGGGATGACGTCAAATCATCATGCCCCTTAAGGTCTTGGGCTTCAAGCATGCTACA 1020 |||||||||||||||||||||||||||||||||| | |||||||||||| |||||||||| Sbjct 1103 AGGTGGGGATGACGTCAAATCATCATGCCCCTTATG-TCTTGGGCTTCACGCATGCTACA 1161 Query 1021 ATGGCCAGGTACAAAGGGC 1039 |||||| |||||||||||| Sbjct 1162 ATGGCC-GGTACAAAGGGC 1179C) Urutan nukleotida gen 16S rRNA isolat BAL2a
GCNCNGTGANTTAGCGGCGGACGGGAGAGTCACACGTGGGTCACCTACCTATNAGACTGGTATCACTCC GGGAAACCGGGGCTAATGCCGGATAACATTTAGAACCGCGTGGTTCTCAAGTGAAGGATGGTTTTGCTAT CACTTATAGATGGACCCGCGCCGTATTAGCTAGTGGGGAGGGTAACGGCTTACCAAGGCAACGATACGT ACCCGACCTGAAAGGGGGATCGGCCCCCCGGGAACGGAAACACGGCCCAAACTCCTACGGGAGGCAGC ATTAGGGAATCTTCCGCAAGGGGCAAAAGCCTGACGGANCACCGCCCCGGGGGTGATGAAGGTCTTCGA ATCGAAAAATTTTGTTATTAGGGAAAAACAAATGTGTAAGTAACTGGGCGCGTCTTGACGGTACCTAATC AGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGTGGCAAGCGTTGTCCGGAATTA TTGGGCGTAAGGGCGCGTAGGCGGTTTCTTAAGTCTGATGTGAAGCCCCGGCTCANNGGNGAGGTCATT GGAACTGGAACTTGAGTGCAGANAGAAGTGNATTCNTGTGTAGCGTGAATGCGCAGAGATATGAGNNCA CNGTGCGAAGCGANTTCTGTCTGTACTGACGCTGATGNGCGAAGCGTNNNNCNNCAGNTAGATACNTGT AGTCACGCNTAANGATGAGTGCTAGTGTAGGNNNNCNNNNTAGTGCTGCAGCTANGCATTAGCA
D)
Hasil BLAST-N sekuen gen 16S rRNA isolat BAL2a dengan data Genbank
|JF072163.1| Uncultured bacterium clone ncd1261e03c1 16S ribosomal RNA gene, partial
sequence
Length = 1372; Score = 854 bits (462); Expect = 0.0; Identities = 609/683 (89%); Gaps =
24/683 (4%); Strand = Plus/Plus
Query 11 TTAGCGGCGGACGGGAGAGTCACACGTGGGTCACCTACCTATNAGACTGGTATCACTCCG 70 ||||||||||||||| |||| |||||||||| |||||||||| ||||||| || |||||| Sbjct 70 TTAGCGGCGGACGGGTGAGTAACACGTGGGTAACCTACCTATAAGACTGGAATAACTCCG 129 Query 71 GGAAACCGGGGCTAATGCCGGATAACATTTAGAACCGCGTGGTTCTCAAGTGAAGGATGG 130 |||||||||||||||||||||||||||||||||||||| ||||||| ||||||| ||||| Sbjct 130 GGAAACCGGGGCTAATGCCGGATAACATTTAGAACCGCATGGTTCTAAAGTGAAAGATGG 189 Query 131 TTTTGCTATCACTTATAGATGGACCCGCGCCGTATTAGCTAGTGGGGAGGGTAACGGCTT 190 ||||||||||||||||||||||||||||||||||||||||||| || | ||||||||||| Sbjct 190 TTTTGCTATCACTTATAGATGGACCCGCGCCGTATTAGCTAGTTGGTAAGGTAACGGCTT 249 Query 191 ACCAAGGCAACGATACGTACCCGACCTGAAAGGGGGATCGGCCCCCCGGGAACGGAAACA 250 ||||||||||||||||||| ||||||||| |||| |||||||| | | ||||| || ||| Sbjct 250 ACCAAGGCAACGATACGTAGCCGACCTGAGAGGGTGATCGGCCACACTGGAACTGAGACA 309 Query 251 CGGCCCAAACTCCTACGGGAGGCAGCATTAGGGAATCTTCCGCAAGGGGCAAAAGCCTGA 31017
||| ||| ||||||||||||||||||| ||||||||||||||||| |||| ||||||||| Sbjct 310 CGGTCCAGACTCCTACGGGAGGCAGCAGTAGGGAATCTTCCGCAATGGGCGAAAGCCTGA 369 Query 311 CGGANCACCGCCCCGGGGGTGATGAAGGTCTTCGAATCGAAAAATTTTGTTATTAGGGAA 370 |||| || |||| || |||||||||||||||||| |||| |||| | ||||||||||||| Sbjct 370 CGGAGCAACGCCGCGTGGGTGATGAAGGTCTTCGGATCGTAAAACTCTGTTATTAGGGAA 429 Query 371 AAACAAATGTGTAAGTAACTGGGCGCGTCTTGACGGTACCTAATCAGAAAGCCACGGCTA 430 |||||||||||||||||||| || ||||||||||||||||||||||||||||||||||| Sbjct 430 GAACAAATGTGTAAGTAACTGTGCACGTCTTGACGGTACCTAATCAGAAAGCCACGGCTA 489 Query 431 ACTACGTGCCAGCAGCCGCGGTAATACGTAGGTGGCAAGCGTTGTCCGGAATTATTGGGC 490 ||||||||||||||||||||||||||||||||||||||||||| |||||||||||||||| Sbjct 490 ACTACGTGCCAGCAGCCGCGGTAATACGTAGGTGGCAAGCGTTATCCGGAATTATTGGGC 549 Query 491 GTAAGG-GCGCGTAGGCGGTTTCTTAAGTCTGATGTGAA-GCCC-CGGCTCANNGGNG-A 546 |||| | |||||||||||||||||||||||||||||||| |||| ||||||| | | | Sbjct 550 GTAAAGCGCGCGTAGGCGGTTTCTTAAGTCTGATGTGAAAGCCCACGGCTCAACCGTGGA 609 Query 547 GG-TCATTGGAA-CTGG-AA-CTTGAGTGCAGAN-AG-AA-GTGNA-TTCN-TGTGTAGC 597 || ||||||||| |||| || |||||||||||| || || ||| | ||| |||||||| Sbjct 610 GGGTCATTGGAAACTGGGAAACTTGAGTGCAGAAGAGGAAAGTGGAATTCCATGTGTAGC 669 Query 598 G-TGAA-TGCGCAGAGATATG-AGNN-CACN-GTG-CGAAG-CGANTT-CTG-TCTGTA- 647 | |||| |||||||||||||| || ||| ||| ||||| ||| || ||| |||||| Sbjct 670 GGTGAAATGCGCAGAGATATGGAGGAACACCAGTGGCGAAGGCGACTTTCTGGTCTGTAA 729 Query 648 CTGACGCTGATGNGCGAA-GCGT 669 |||||||||||| ||||| |||| Sbjct 730 CTGACGCTGATGTGCGAAAGCGT 752E) Urutan nukleotida gen 16S rRNA isolat NTT3a
GGGAGAAGTATAGAGCTGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACGTGGTGTAACCTGCC CATAAGACTGGTATAACTCCGGGAAACCGGGGCTAATACCGGAATAATATTTTGAACCGCATGGTTCGC AATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACCCGCGTCGCTTTAGCTATGTGGTGAGGTAACG GCTCACCAAGGCGACGATGCGTAGCCGACCTGAGAGGGTGATCGGGCACACTGGGACTGACACGCGGCC CAGACTCCTACGGGAGGCAGCAGTAGGGAATCTTCCACAATGGACGAAAGTCTGACGGAGCAACGCCGC GTGAGTGATGAAGGCTTTCGGGTCGTAAAACTCTGTTGTTAGGGAAGAACAAGTGCTAGTTGAATAAGC TGGCACCTTGACGGTACCTAACCATAAAGCCACGGCTAACTACGTGCCAGCACCCGCGATTATACT
F)
Hasil BLAST-N sekuen gen 16S rRNA isolat NTT3a dengan data Genbank
|FJ611939.1| Bacillus sp. AF-777 16S ribosomal RNA gene, partial sequence
Length = 1522; Score = 776 bits (420); Expect = 0.0; Identities = 455/471 (97%); Gaps = 5/471
(1%); Strand = Plus/Plus
Query 12 AGAGC-TGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACGTGGTGTAACCTGCCC 70 ||||| |||||||||||||||||||||||||||||||||||||||||| ||||||||||| Sbjct 71 AGAGCTTGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACGTGG-GTAACCTGCCC 129 Query 71 ATAAGACTGGTATAACTCCGGGAAACCGGGGCTAATACCGGAATAATATTTTGAACCGCA 130 |||||||||| |||||||||||||||||||||||||||||| |||||||||||||||||| Sbjct 130 ATAAGACTGGGATAACTCCGGGAAACCGGGGCTAATACCGG-ATAATATTTTGAACCGCA 188 Query 131 TGGTTCGCAATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACCCGCGTCGCTTTAG 190 ||||||| ||||||||||||||||||||||||||||||||||||||||||||||| |||| Sbjct 189 TGGTTCGAAATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACCCGCGTCGCATTAG 248 Query 191 CTATG-TGGTGAGGTAACGGCTCACCAAGGCGACGATGCGTAGCCGACCTGAGAGGGTGA 249 ||| | ||||||||||||||||||||||||| |||||||||||||||||||||||||||| Sbjct 249 CTA-GTTGGTGAGGTAACGGCTCACCAAGGCAACGATGCGTAGCCGACCTGAGAGGGTGA 307 Query 250 TCGGGCACACTGGGACTGACACGCGGCCCAGACTCCTACGGGAGGCAGCAGTAGGGAATC 309 |||| |||||||||||||| || ||||||||||||||||||||||||||||||||||||| Sbjct 308 TCGGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTAGGGAATC 367 Query 310 TTCCACAATGGACGAAAGTCTGACGGAGCAACGCCGCGTGAGTGATGAAGGCTTTCGGGT 369 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 368 TTCCACAATGGACGAAAGTCTGACGGAGCAACGCCGCGTGAGTGATGAAGGCTTTCGGGT 42718
Query 370 CGTAAAACTCTGTTGTTAGGGAAGAACAAGTGCTAGTTGAATAAGCTGGCACCTTGACGG 429 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 428 CGTAAAACTCTGTTGTTAGGGAAGAACAAGTGCTAGTTGAATAAGCTGGCACCTTGACGG 487 Query 430 TACCTAACCATAAAGCCACGGCTAACTACGTGCCAGCACCCGCGATTATAC 480 |||||||||| ||||||||||||||||||||||||||| ||||| | |||| Sbjct 488 TACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATAC 538G) Urutan nukleotida gen 16S rRNA isolat NTT3e
GGGGAAGGGTTAGAGCTTGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACGTGGGTAACCTGCC CATAAGACTGGGATAACTCCGGGAAACCGGGGCTAATACCGGATAACATTTTGAACCGCATGGTTCGAA ATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACCCGCGTCGCATTAGCTAGTTGGTGAGGTAACGG CTCACAAGGCAACGATGCGTAGCCGACCTGAGAGGGTGATCGGCCACACTGGGACTGAGACACGGCCCA GACTCCTACGGGAGGCAGCAGTAGGGAATCTTCCGCAATGGACGAAAGTCTGACGGAGCAACGCCGCGT GAGTGATGAAGGCTTTCGGGTCGTAAAACTCTGTTGTTAGGGAAGAACAAGTGCTAGTTGAATAAGCTG GCACCTTGACGGTACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGT GGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGCGCGCAGGTGGTTTCTTAAGTCTGATGTGAAAGCC CACGGCTCAACCGTGGAGGGTCATTGGAAACTGGGAGACTTGAGTGCAGAAGAGGAAAGTGGAATTCCA TGTGTAGCGGTGAAATGCGTAGAGATATGGAGGAACACCAGTGGCGAAGGCGACTTTCTGGTCTGTAAC TGACACTGAGGCGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCCTTAAACG ATGAGTGCTAAGTGTTAGAGGGTTTCCGCCCTTTACTGCTGCAGTTTAACGCATTTAATAATTTC
H) Hasil BLAST-N sekuen gen 16S rRNA isolat NTT3e dengan data Genbank
|AM981260.1| Bacillus sp. NIOT-3 partial 16S rRNA gene, isolate NIOT-3
Length = 1445; Score = 1461 bits (791); Expect = 0.0; Identities = 807/814 (99%); Gaps =
3/814 (0%); Strand = Plus/Plus
Query 4 GAAGGGTTAGAGCTTGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACGTGGGTAA 63 ||||| |||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 28 GAAGGATTAGAGCTTGCTCTTATGAAGTTAGCGGCGGACGGGTGAGTAACACGTGGGTAA 87 Query 64 CCTGCCCATAAGACTGGGATAACTCCGGGAAACCGGGGCTAATACCGGATAACATTTTGA 123 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 88 CCTGCCCATAAGACTGGGATAACTCCGGGAAACCGGGGCTAATACCGGATAACATTTTGA 147 Query 124 ACCGCATGGTTCGAAATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACCCGCGTCG 183 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 148 ACCGCATGGTTCGAAATTGAAAGGCGGCTTCGGCTGTCACTTATGGATGGACCCGCGTCG 207 Query 184 CATTAGCTAGTTGGTGAGGTAACGGCTCA-CAAGGCAACGATGCGTAGCCGACCTGAGAG 242 ||||||||||||||||||||||||||||| |||||||||||||||||||||||||||||| Sbjct 208 CATTAGCTAGTTGGTGAGGTAACGGCTCACCAAGGCAACGATGCGTAGCCGACCTGAGAG 267 Query 243 GGTGATCGGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTAGG 302 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 268 GGTGATCGGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTAGG 327 Query 303 GAATCTTCCGCAATGGACGAAAGTCTGACGGAGCAACGCCGCGTGAGTGATGAAGGCTTT 362 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 328 GAATCTTCCGCAATGGACGAAAGTCTGACGGAGCAACGCCGCGTGAGTGATGAAGGCTTT 387 Query 363 CGGGTCGTAAAACTCTGTTGTTAGGGAAGAACAAGTGCTAGTTGAATAAGCTGGCACCTT 422 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 388 CGGGTCGTAAAACTCTGTTGTTAGGGAAGAACAAGTGCTAGTTGAATAAGCTGGCACCTT 447 Query 423 GACGGTACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAG 482 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 448 GACGGTACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAG 507 Query 483 GTGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGCGCGCAGGTGGTTTCTTAAGTCT 542 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 508 GTGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGCGCGCAGGTGGTTTCTTAAGTCT 567 Query 543 GATGTGAAAGCCCACGGCTCAACCGTGGAGGGTCATTGGAAACTGGGAGACTTGAGTGCA 602 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 568 GATGTGAAAGCCCACGGCTCAACCGTGGAGGGTCATTGGAAACTGGGAGACTTGAGTGCA 62719
Query 603 GAAGAGGAAAGTGGAATTCCATGTGTAGCGGTGAAATGCGTAGAGATATGGAGGAACACC 662 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 628 GAAGAGGAAAGTGGAATTCCATGTGTAGCGGTGAAATGCGTAGAGATATGGAGGAACACC 687 Query 663 AGTGGCGAAGGCGACTTTCTGGTCTGTAACTGACACTGAGGCGCGAAAGCGTGGGGAGCA 722 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 688 AGTGGCGAAGGCGACTTTCTGGTCTGTAACTGACACTGAGGCGCGAAAGCGTGGGGAGCA 747 Query 723 AACAGGATTAGATACCCTGGTAGTCCACGCCCTTAAACGATGAGTGCTAAGTGTTAGAGG 782 ||||||||||||||||||||||||||||||| | |||||||||||||||||||||||||| Sbjct 748 AACAGGATTAGATACCCTGGTAGTCCACGCCGT-AAACGATGAGTGCTAAGTGTTAGAGG 806 Query 783 GTTTCCGCCCTTTACTGCTGCAGTTTAACGCATT 816 |||||||||||||| ||||| |||| |||||||| Sbjct 807 GTTTCCGCCCTTTAGTGCTGAAGTT-AACGCATT 839I) Urutan nukleotida gen 16S rRNA isolat NTT6B6
GGGGGGGAAAAGGGACTTGCTCCCTGATGTTAGCGGCGGACGGGTGAGTAACACGTGGGTAACCTGCCT GTAAGACTGGGATAACTCCGGGAAACCGGGGCTAATACCGGATGGTTGTTTGAACCGCATGGTTCAAAC ATAAAAGGTGGCTTCGGCTACCACTTACAGATGGACCCGCGGCGCATTAGCTAGTTGGTGAGGTAACGG CTCACCAAGGCAACGATGCGTAGCCGACCTGAGAGGGTGATCGGCCACACTGGGACTGAGACACGGCCC AGACTCCTACGGGAGGCAGCAGTAGGGAATCTTCCGCAATGGACGAAAGTCTGACGGAGCAACGCCGCG TGAGTGATGAAGGTTTTCGGATCGTAAAGCTCTGTTGTTAGGGAAGAACAAGTACCGTTCGAATAGGGC GGTACCTTGACGGTACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGG TGGCAAGCGTTGTCCGGAATTATTGGGCGTAAAGGGCTCGCAGGCGGTTTCTTAAGTCTGATGTGAAAGC CCCCGGCTCAACCGGGGAGGGTCATTGGAAACTGGGGAACTTGAGTGCAGAAGAGGAGAGTGGAATTCC ACGTGTAGCGGTGAAATGCGTAGAGATGTGGAGGAACACCAGTGGCGAAGGCGACTCTCTGGTCTGTAA CTGACGCTGAGGAGCGAAAGCGTGGGGAGCGAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACG ATGAGTGCTAAGTGTTAGGGGGTTTCCGCCCCTTAGTGCTGCAGCTAACGCATTAAGCACTCCGCCTGGG GAGTACGGTCGCAAGACTGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTT TAATTCGAAGCAACGCGAAGAACCTTACCAGGTCTTGACATCCTCTGACAATCCTAGAGATAGGACGTCC CCTTCGGGGGCAGAGTGACAGGTGGTGCATGGTTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTC CCGCAACGAGCGCAACCCTTGATCTTAGTTGCCAGCATTCAGTTGGGCACTCTAAGGTGACTGCCGGTGA CAAACCGGAGGAAGGTGGGGATGACGTCAAATCATCATGCCCCT