4 HASIL DAN PEMBAHASAN
4. 1 . HasilHasil analisa proksimat tubuh ikan menunjukkan bahwa secara umum terjadi peningkatan kandungan protein dan lemak tubuh ikan uji pada akhir percobaan seiring dengan peningkatan kadar protein dan rasio energi protein pakan. Sebaliknya kadar abu tubuh ikan cenderung menurun dengan bertambahnya kadar protein dan rasio energi protein pakan, sedangkan kadar BETN tubuh ikan pada perlakuan A1B1, A1B2, A2B1, A2B2 dan A3B1 terlihat menurun dan kemudian meningkat pada perlakuan A3B2, A4B1 dan A4B2. Pengaruh pakan uji terhadap komposisi proksimat tubuh ikan pada setiap perlakuan disajikan pada Tabel 4 dan hasil proksimat lengkapnya dapat dilihat pada Lampiran 2.
Tabel 4. Komposisi proksimat tubuh ikan kuwe (% bobot kering)
Parameter Awal
Perlakuan (Kadar Protein dan Rasio Energi Protein C/P)
A1B1 (33;9) A1B2 (33;11) A2B1 (37;9) A2B2 (37;11) A3B1 (41;9) A3B2 (41;11) A4B1 (45;9) A4B2 (45;11) Protein 62.88 61.76±1.22 64.13±0.93 64.90±1.19 63.20±1.38 66.58±1.30 63.07±1.00 64.44±0.79 64.09±0.50 Lemak 11.72 15.26±0.51 15.97±0.86 14.25±0.43 14.56±0.97 14.20±0.34 14.97±0.77 15.18±0.57 14.67±1.21 BETN 3.88 2.91±0.94 2.75±1.05 2.82±0.74 3.02±0.61 3.38±0.83 5.55±0.90 5.28±0.0.29 7.09±1.02 Kadar Abu 21.52 20.08±0.64 17.15±0.26 18.04±0.76 19.22±0.52 15.84±0.48 16.41±1.71 15.10±0.57 14.14±0.58
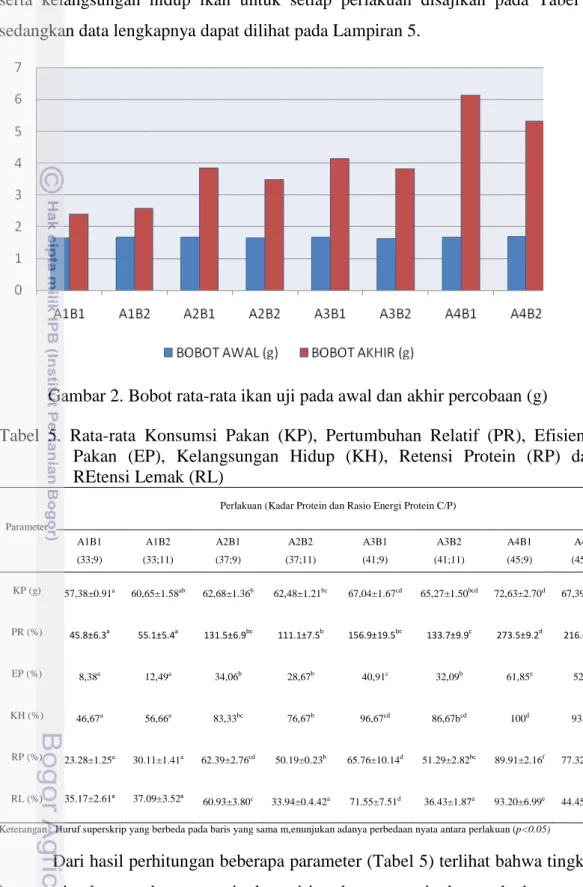
Hasil percobaan pemberian pakan dengan tingkat protein dan rasio energi protein yang berbeda mempengaruhi pertumbuhan juvenile ikan kuwe. Data perubahan bobot rata-rata ikan per ekor dapat dilihat pada gambar 2, sedangkan data lengkapnya terdapat pada Lampiran 5.
Perbedaan tingkat protein dan rasio energi protein dalam pakan juga berpengaruh pada tingkat konsumsi pakan, pertumbuhan relatif, efisiensi pakan, retensi protein dan lemak serta kelangsungan hidup ikan. Data mengenai tingkat konsumsi pakan, pertumbuhan relatif, efisiensi pakan, retensi protein dan lemak
serta kelangsungan hidup ikan untuk setiap perlakuan disajikan pada Tabel 5 sedangkan data lengkapnya dapat dilihat pada Lampiran 5.
Gambar 2. Bobot rata-rata ikan uji pada awal dan akhir percobaan (g)
Tabel 5. Rata-rata Konsumsi Pakan (KP), Pertumbuhan Relatif (PR), Efisiensi Pakan (EP), Kelangsungan Hidup (KH), Retensi Protein (RP) dan REtensi Lemak (RL)
Parameter
Perlakuan (Kadar Protein dan Rasio Energi Protein C/P)
A1B1 (33;9) A1B2 (33;11) A2B1 (37;9) A2B2 (37;11) A3B1 (41;9) A3B2 (41;11) A4B1 (45;9) A4B2 (45;11) KP (g) 57,38±0.91a 60,65±1.58ab 62,68±1.36b 62,48±1.21bc 67,04±1.67cd 65,27±1.50bcd 72,63±2.70d 67,39±1.59e PR (%) 45.8±6.3a 55.1±5.4a 131.5±6.9bc 111.1±7.5b 156.9±19.5bc 133.7±9.9c 273.5±9.2d 216.6±9.9 EP (%) e 8,38a 12,49a 34,06b 28,67b 40,91c 32,09b 61,85e 52,48 KH (%) d 46,67a 56,66a 83,33bc 76,67b 96,67cd 86,67bcd 100d 93,33 RP (%) cd 23.28±1.25a 30.11±1.41a 62.39±2.76cd 50.19±0.23b 65.76±10.14d 51.29±2.82bc 89.91±2.16f 77.32±2.25 RL (%) e 35.17±2.61ª 37.09±3.52ª 60.93±3.80c 33.94±0.4.42a 71.55±7.51d 36.43±1.87a 93.20±6.99e 44.45±6.16a
Keterangan : Huruf superskrip yang berbeda pada baris yang sama m,enunjukan adanya perbedaan nyata antara perlakuan (p<0.05)
Dari hasil perhitungan beberapa parameter (Tabel 5) terlihat bahwa tingkat konsumsi pakan cenderung meningkat seiring dengan meningkatnya kadar protein pakan namun cenderung menurun pada perlakuan rasio energi yang lebih tinggi pada kadar protein yang sama. Pada kadar Protein A4 (45%) tingkat konsumsi pakan lebih tinggi pada rasio energi protein B1 (9 kkal GE/g) dan berbeda nyata
(p<0,05) dibandingkan dengan rasio energi protein B2 (11 kkal GE/g). Demikian pula dengan perlakuan kadar protein A3 (41%) dan A2 (37%) dimana konsumsi pakan lebih tinggi pada rasio energi B1 namun tidak berbeda nyata dengan rasio energi protein B2 (p>0,05). Pada perlakuan A1B1 dan A1B2 (kadar protein terendah) konsumsi pakan meningkat seiring dengan peningkatan rasio energi protein pada kadar protein yang sama namun tidak menunjukkan perbedaan yang nyata (p>0.05). Tingkat konsumsi pakan tertinggi terdapat pada perlakuan A4B1 (45;9) dan berbeda nyata dengan semua perlakuan lainnya (p<0.05). Data lengkap tingkat konsumsi pakan terdapat pada Lampiran 5.
Gambar 3. Tingkat konsumsi pakan (g)
Demikian pula dengan laju pertumbuhan relatif, dimana terlihat bahwa meningkatnya kadar protein akan meningkatkan laju pertumbuhan relatif namun cenderung menurun pada perlakuan dengan rasio energi yang lebih tinggi pada level protein yang sama namun tidak menunjukkan perbedaan yang nyata (p>0,05), kecuali pada level protein A4 (45%) rasio energi protein B1 menghasilkan pertumbuhan relatif yang lebih tinggi dan berbeda nyata dengan rasio energi protein B2 (p<0,05). Laju pertumbuhan relatif tertinggi diperoleh pada perlakuan A4B1 (45;9) yang berbeda nyata dengan tujuh perlakuan lainnya (p<0,05). Data lengkapnya dapat dilihat pada Lampiran 5.
Gambar 4. Laju pertumbuhan relatif (%)
Efisiensi pakan ikan uji dalam percobaan ini memiliki kecenderungan yang sama dengan tingkat konsumsi pakan dan laju pertumbuhan relatif dimana peningkatan kadar protein dalam pakan meningkatkan efisiensi dan peningkatan kadar rasio energi protein pada level protein yang sama akan menurunkan efisiensi, kecuali pada perlakuan dengan level protein terendah dalam percobaan ini yaitu A1B1 (33;9) dan A1B2 (33;11) namun tidak berbeda nyata (p>0,05). Efisiensi pakan tertinggi dalam percobaan ini terjadi pada perlakuan A4B1 (45;9) dan terendah pada perlakuan A1B1 (33;9). Data lengkap pada Lampiran 5.
Gambar 5. Efisiensi pakan (%)
d a a bc b bc c d e
Pemberian pakan dengan kadar protein dan rasio energi protein yang berbeda memberikan pengaruh tingkat kelangsungan hidup yang berbeda pada masing-masing perlakuan. Semakin tinggi kadar protein dalam pakan maka semakin tinggi pula tingkat kelangsungan hidup ikan uji. Tingkat kelangsungan hidup bervariasi dan berada pada kisaran 46.67 – 100 %. Pada level protein yang sama tingkat kelangsungan hidup lebih tinggi pada rasio energi protein B1, kecuali pada level protein A1, dimana tingkat kelangsungan hidup lebih tinggi pada rasio energi protein B2. Hasil analisis statistik menunjukkan perbedaan rasio energi protein pada level protein yang sama menghasilkan tingkat kelangsungan hidup yang tidak berbeda nyata (p>0,05), Sedangkan perbedaan tingkat protein dalam pakan memberikan pengaruh yang berbeda nyata terhadap kelangsungan hidup ikan uji (p<0,05).
Dari hasil perhitungan retensi protein (Tabel 5) menunjukkan bahwa pemberian pakan dengan kadar protein dan rasio energi protein yang berbeda berpengaruh pada retensi protein dimana nilainya akan semakin meningkat seiring dengan peningkatan kadar protein dan rasio energi protein. Retensi protein tertinggi terdapat pada perlakuan A4B1 (p<0,05) yang diikuti oleh perlakuan A4B2 (45;11) dan semakin menurun pada perlakuan A3B1 (41;9), A2B1 (37;9), A3B2 (41;11), A2B2 (37;11), A1B2 (33;11) dan A1B1(33;9). Data lengkap pada Lampiran 6.
Pemberian pakan dengan kadar protein dan rasio energi protein yang berbeda juga berpengaruh pada nilai retensi lemak. Peningkatan kadar protein akan semakin meningkatkan retensi lemak kecuali pada perlakuan A2B2 (37;11) namun tidak menunjukkan perbedaan yang nyata dengan perlakuan A1B1, A3B2, A1B2 dan A4B2. Peningkatan rasio energi protein pada level protein yang sama akan menurunkan nilai retensi lemak. Pada level protein A4, perlakuan dengan rasio energi protein B1 menghasilkan nilai retensi lemak yang lebih tinggi dan berbeda nyata dengan rasio energi protein B2. Demikian pula halnya dengan perlakuan pada level protein A3 dan A2. Retensi lemak tertinggi dalam percobaan ini terdapat pada perlakuan A4B1 (45;9) dan berbeda nyata dengan tujuh perlakuan lainnya (p<0,05). Data lengkap retensi lemak masing-masing perlakuan terdapat pada Lampiran 7.
Hasil analisa kandungan glikogen hati, glikogen otot, lemak hati dan lemak otot dapat dilihat pada Tabel 6.
Tabel 6. Kandungan glikogen hati (GH), glikogen otot (GO), lemak hati (LH) dan lemak otot (LO)
Parameter Awal
Perlakuan (Kadar Protein dan Rasio Energi Protein C/P)
A1B1 (33;9) A1B2 (33;11) A2B1 (37;9) A2B2 (37;11) A3B1 (41;9) A3B2 (41;11) A4B1 (45;9) A4B2 (45;11) GH (%) 0.014 0.004 0.012 0.002 0.016 0.008 0.177 0.339 0.161 GO (%) 3.139 0.383 1.196 0.707 0.130 0.756 0.238 3.008 1.187 LH (%) 37.88 46.22 54.08 48.65 68.24 49.24 61.64 42.82 52.07 LO (%) 15.55 12.03 12.13 14.24 13.72 13.95 14.95 12.39 12.68
Berdasarkan hasil analisa glikogen hati dan otot serta lemak hati dan otot pada Tabel 6, terlihat bahwa kadar glikogen hati cenderung naik pada semua perlakuan dengan rasio energi protein yang tinggi pada level protein A2 (37%) dan A3 (41%). Pada level protein A4 (45%) kenaikan kadar glikogen hati terjadi pada kedua level rasio energi protein dan kenaikan terbesar ada pada rasio energi protein 9 kkal GE/g (B1). Kadar glikogen hati tertinggi terdapat pada perlakuan A4B1 (45;9) yaitu sebesar 0.339% dan yang terendah pada perlakuan A2B1 (37;9) sebesar 0.002 %. Pemberian pakan dengan kadar protein dan rasio energi protein yang berbeda akan menurunkan kadar glikogen otot ikan uji pada semua perlakuan, terutama pada perlakuan dengan rasio energi yang lebih tinggi. Kadar glikogen otot tertinggi dalam percobaan ini terdapat pada perlakuan A4B1 (45;9) sebesar 3,008 % dan yang terendah pada perlakuan A2B2 (37;11) sebesar 0,130 %. Lemak hati ikan uji cenderung meningkat seiring dengan peningkatan kadar protein pakan sampai pada level protein A2 (37%) dan peningkatan kadar lemak hati akan semakin besar pada level rasio energi protein 11 kkal GE/g pada semua perlakuan. Kadar lemak hati tertinggi terdapat pada perlakuan A3B2 (41;11) dan terendah pada perlakuan A4B1 (45;9). Kadar lemak otot ikan uji dalam percobaan ini mengalami penurunan pada semua perlakuan. Lemak otot
tertinggi ada pada pelakuan A3B2 (41;11) sebesar 14,95 % dan yang terendah pada perlakuan A1B1 (33;9) sebesar 12,03 %.
Hasil pengukuran eskresi total ammonia nitrogen (NH3-N) dan hepatosomatik indeks disajikan pada Tabel 7 dan data lengkap setiap ulangan dapat dilihat pada Lampiran 8 dan 9.
Tabel 7. Hepatosomatik Indeks (HSI) dan Eskresi Total Amonia Nitrogen (TAN)
Parameter
Perlakuan (Kadar Protein dan Rasio Energi Protein C/P)
A1B1 A1B2 A2B1 A2B2 A3B1 A3B2 A4B1 A4B2
HSI (%) 1.38±0.03a 1.45±0.03a 1.57±0.02b 1.56±0.04b 1.61±0.02bc 1.58±0.05b 1.67±0.02c 1.62±0.02bc
TAN (mg/g tubuh/jam)
0.016 0.014 0.022 0.021 0.027 0.016 0.028 0.024
Secara umum terlihat bahwa nilai hepatosomatik indeks ikan uji (Tabel 7) cenderung meningkat seiring dengan meningkatnya kadar protein dan rasio energi protein. Di level protein yang sama nilai hepatosomatik indeks cenderung lebih tinggi pada rasio protein energi yang lebih rendah (B1), kecuali pada level protein A1 (33) nilai hepatosomatik indeks lebih tinggi pada rasio energi protein B2, namun secara statatistik hasilnya tidak berbeda nyata Nilai hepatosomatik indeks tertinggi dalam percobaan ini adalah pada perlakuan A4B1 (45;9) yang tidak berbeda nyata dengan perlakuan A4B2 (45;11) dan A3B1 (41;9) namun ketiganya menunjukkan perbedaan yang nyata dengan 4 perlakuan lainnya (p<0,05).
Eskresi total amonia nitrogen (NH3-N) yang diukur selama tiga jam menunjukkan adanya peningkatan seiring dengan meningkatnya kadar protein pakan pada setiap perlakuan. Berdasarkan data pada Tabel 7 terlihat bahwa eskresi ammonia akan semakin tinggi di level protein 41% (A3) dan 45% (A4) pada rasio energi protein 9 kkal GE/g (B1). Nilai eskresi total amonia tertinggi dalam percobaan ini terdapat pada perlakuan A4B1 (45;9) dan yang terendah pada perlakuan A1B2 (33;11).
4.2. Pembahasan
Pakan uji dengan kadar protein dan rasio energi protein yang berbeda memberikan pengaruh terhadap komposisi proksimat tubuh ikan (Tabel 4). Pada setiap perlakuan terlihat bahwa kadar protein dan lemak tubuh ikan semakin tinggi seiring dengan peningkatan kadar protein dan rasio energi protein pakan. Sebaliknya kadar abu tubuh ikan cenderung menurun dengan bertambahnya kadar protein dan rasio energi protein pakan, sedangkan kadar BETN tubuh ikan pada perlakuan A1B1, A1B2, A2B1, A2B2 dan A3B1 terlihat menurun dan kemudian meningkat pada perlakuan A3B2, A4B1 dan A4B2. Berdasarkan hasil analisis terhadap komposisi proksimat tubuh ikan terlihat bahwa kadar protein tubuh ikan semakin meningkat dengan meningkatnya kadar protein pakan. Meningkatnya kadar protein tubuh ikan disebabkan oleh adanya peningkatan kadar protein yang dikonsumsi sebagai akibat dari meningkatnya kadar protein dan dalam pakan.
Tabel 4 menunjukkan bahwa komposisi kandungan protein tubuh ikan diimbangi oleh kandungan lemak. Adelina et al. (2000) menyatakan bahwa adanya penyimpanan lemak tubuh yang tinggi dan penyimpanan protein pada batas tertentu sesuai kemampuan ikan untuk mensintesis protein tubuh maka akan menyebabkan kandungan protein tubuh ikan cenderung menurun. Itulah sebabnya pada penelitian ini ikan uji yang memiliki kandungan lemak lebih tinggi cenderung akan memiliki kandungan protein tubuh yang lebih rendah (A1B1, A1B2, A4B1, A3B2).
Pemberian pakan dengan kadar protein yang sama dengan rasio energi protein berbeda menghasilkan laju pertumbuhan relatif, konsumsi pakan, efisiensi pakan, retensi protein, retensi lemak dan kelangsungan hidup yang berbeda. Pakan dengan rasio energi protein lebih tinggi pada kadar protein yang sama menghasilkan laju pertumbuhan relatif, konsumsi pakan, efisiensi pakan, retensi protein, retensi lemak dan kelangsungan hidup yang lebih rendah. Hasil penelitian ini menunjukkan bahwa kadar protein pakan A4B2 (45%) yang sama dengan pakan A4B1 tetapi total energi pakan A4B2 lebih tinggi (Tabel 2) menghasilkan laju pertumbuhan relatif, konsumsi pakan, efisiensi pakan, retensi protein, retensi lemak dan kelangsungan hidup yang lebih rendah. Hal ini menunjukkan bahwa penambahan total energi pakan pada kadar protein sama mempengaruhi jumlah
simpanan protein tubuh yang pada akhirnya mempengaruhi laju pertumbuhan relatif, konsumsi pakan, efisiensi pakan, retensi protein, retensi lemak dan kelangsungan hidup. Hal ini terjadi mungkin karena pakan A4B2 memiliki total energi yang terlalu tinggi yang disebabkan kandungan lemak yang tinggi (19,45%). Menurut Cho dan Watanabe (1985) lemak yang dibutuhkan ikan berkisar antara 4 - 18%, sedangkan hasil penelitian Chu et al. (1996) ikan kerapu
E. aerolatus yang diberi pakan dengan kandungan lemak 10 % pada pakan yang
mengandung 60% protein menghasilkan pertumbuhan yang baik. Tingginya total energi pakan karena kadar lemak yang tinggi menyebabkan ikan merasa cepat kenyang yang berakibat pada konsumsi pakan menjadi rendah. Keadaan ini akan membatasi banyaknya protein yang masuk ke dalam tubuh ikan sehingga protein yang disimpan menjadi protein tubuh lebih sedikit (Seenappa dan Devaraj 1995).
Perbedaan tingkat konsumsi pakan dalam penelitian ini selain disebabkan oleh pengaruh pakan dengan kadar protein dan rasio energi yang berbeda juga dapat disebabkan oleh perbedaan tingkat pertumbuhan ikan (ukuran), kandungan energi pakan dan kadar lemak pakan. Bobot ikan yang semakin besar akan menyebabkan tingkat konsumsi pakan yang tinggi seperti pada perlakuan A4B1 (45;9) dan A4B2 (45;11), sebaliknya untuk ikan yang memiliki bobot lebih kecil memiliki tingkat konsumsi pakan yang rendah seperti pada perlakuan A1B1 (33;9) dan A1B2 (33;11). Pada perlakuan pakan yang memiliki kandungan energi dan lemak yang tinggi tingkat konsumsi pakan juga cenderung menurun dibandingkan dengan konsumsi pakan pada perlakuan dengan level protein yang sama namun kandungan energi dan lemak yang lebih rendah. Menurut Robinson et al. (2001) energi dalam pakan akan mempengaruhi asupan pakan pada ikan yang diberi makan secara ad libitum. Jika energi dalam pakan terlalu tinggi, ikan akan cepat kenyang sehingga menghentikan konsumsi pakan. Nematipour et al. (1992) menambahkan bahwa tingginya energi dalam pakan ikan menyebabkan terjadinya akumulasi lemak yang tinggi pada tubuh ikan sehingga akan membatasi jumlah pakan yang dikonsumsi.
Peningkatan kadar protein pada perlakuan rasio energi protein B1 (9 kkal GE/g) akan meningkatkan efisiensi pakan dan retensi protein. Hal ini menunjukkan bahwa protein efektif diserap untuk meningkatkan pertambahan
bobot atau persentasi pakan yang diubah menjadi daging meningkat. Meningkatnya efisiensi pakan dan retensi protein tersebut kemungkinan juga disebabkan oleh penggunaan protein hewani yang meningkat pada perlakuan rasio energi protein B1(9 kkal GE/g) dibandingkan dengan rasio energi protein B2 (11 kkal GE/g). Menurut Zonneveld et al. (1991) ikan lebih mudah mencerna protein hewani dibanding protein nabati.
Dalam penelitian ini nilai retensi lemak akan meningkat seiring dengan peningkatan kadar protein pakan pada rasio energi protein B1 (9 kkal GE/g). Hal ini kemungkinan disebabkan oleh kandungan lemak pakan pada perlakuan ini cukup seimbang dan memenuhi kebutuhan lemak ikan sehingga meningkatkan nilai retensinya. Berdasarkan analisis proksimat pakan uji (Tabel 2) kandungan lemak pakan pada perlakuan rasio energi protein B1 berkisar antara 8 – 11 g/100 g pakan (8 -11 %). Ikan kerapu E. aerolatus yang diberi pakan dengan kandungan lemak 10 % pada pakan yang mengandung 60% protein menghasilkan pertumbuhan yang baik (Chu et al. 1996). Menurut Cho dan Watanabe (1985) lemak yang dibutuhkan ikan berkisar antara 4 - 18 %. Kadar lemak yang tepat dalam pakan yang sesuai dengan kebutuhan energi ikan akan menyebabkan protein pakan dapat lebih efisien digunakan untuk pertumbuhan. Berkurangnya penggunaan protein sebagai sumber energi berkaitan dengan protein sparing effect dari lemak untuk ikan-ikan karnivora. Hal ini terbukti dalam penelitian ini dimana peningkatan kadar protein pada semua perlakuan dengan rasio energi B1 (9 kkal GE/g) yang memiliki kadar lemak 8 – 11 % akan meningkatkan kinerja pertumbuhan ikan uji (Tabel 5).
Pertumbuhan ikan sangat terkait dengan faktor luar dan faktor dari dalam tubuh. Faktor luar yang sangat berpengaruh selain lingkungan perairan adalah makanan. Unsur makanan yang sangat terkait dengan pertumbuhan adalah protein, dimana fungsi utama protein adalah untuk pertumbuhan. Tinggi rendahnya kadar protein dan rasio energi protein (kandungan energi total) pakan dapat membatasi pertumbuhan dan pertambahan bobot tubuh.
Hasil penelitian ini menunjukkan bahwa setelah 40 hari pemeliharaan terlihat adanya peningkatan bobot rata-rata individu pada setiap perlakuan (Lampiran 5). Semakin tinggi perubahan bobot tubuh dalam kurun waktu tertentu,
maka laju pertumbuhan akan semakin tinggi (Gambar 2 dan 4). Laju pertumbuhan erat kaitannya dengan bobot tubuh dan bobot tubuh erat kaitannya dengan protein. Hal tersebut dapat dimengerti karena sekitarr 45 – 75% berat kering tubuh ikan terdiri dari protein (Watanabe 1988).
Pakan uji dengan berbagai kadar protein dan rasio energi protein memberikan pengaruh terhadap laju pertumbuhan relatif ikan. Peningkatan kadar protein dalam pakan akan meningkatkan pertumbuhan relatif dan pada kadar protein yang sama pertumbuhan relatif ikan lebih tinggi pada rasio energi protein yang lebih rendah. Hasil penelitian ini menunjukkan bahwa pakan pada perlakuan A4B1 (45;9) menghasilkan laju pertumbuhan relatif tertinggi. Berdasarkan Hasil penelitian Suwirya at al. (2007) pakan dengan kandungan protein 42–46 % memberikan respon pertumbuhan terbaik bagi ikan kuwe Gnathanodon speciosus.
Tingginya laju pertumbuhan relatif pada perlakuan A4B1 diduga karena pakan pada perlakuan ini memiliki keseimbangan protein dan energi yang memenuhi kebutuhan ikan sehingga lemak dan karbohidrat yang dikonsumsi dapat dimanfaatkan dengan efisien sebagai sumber energi, sementara protein dimanfaatkan untuk sintesis protein dalam tubuh ikan. Hal ini terbukti dengan rendahnya kadar lemak ikan pada perlakuan ini yang menandakan bahwa lemak yang dideposit lebih sedikit dibandingkan dengan lemak yang dimanfaatkan sebagai sumber energi. Selain itu tersedianya sumber energi dari non-protein untuk kebutuhan hidup menyebabkan protein yang dikonsumsi ikan sebagian besar dimanfaatkan untuk membangun protein tubuh. Keseimbangan protein dan energi dalam pakan sangat berperan dalam menunjang pertumbuhan ikan dan pemanfaatan energi dan protein tergantung pada tingkat konsumsi pakan. Ketika konsumsi meningkat, maka metabolisme basal dan kebutuhan energi untuk pemeliharaan secara relatif akan berkurang terhadap budget energi, sehingga secara umum akan meningkatkan efisiensi pakan (Saether et al. 1996). Hasil penelitian ini sesuai dengan pendapat Lovell (1988) yang menyatakan bahwa pertumbuhan atau pembentukan jaringan tubuh paling besar dipengaruhi oleh keseimbangan protein dan energi di dalam pakan. Watanabe (1988) menyatakan bahwa pertumbuhan dipengaruhi oleh kualitas makanan dalam hal keseimbangan nutrien-nutriennya, kandungan energi, ketersediaan setiap nutriennya dan kondisi
lingkungan. Ketepatan energi pakan pada perlakuan ini (A4B1) mengakibatkan protein yang ada disimpan cukup banyak dan digunakan untuk pembentukan jaringan tubuh. Ini ditunjukkan oleh nilai retensi protein yang tinggi dan kadar protein tubuh yang juga tinggi.
Kekurangan protein dan energi dalam pakan menyebabkan pertumbuhan ikan menjadi rendah. Hal ini terlihat pada perlakuan A1B1 (33;9) dan A1B2 (33;11), kadar protein pada kedua perlakuan ini kemungkinan masih rendah sehingga banyaknya protein yang masuk untuk disimpan menjadi protein tubuh juga rendah. Walaupun protein tidak dipakai sebagai sumber energi yang utama, namun karena kadar protein pakan masih rendah dan belum mencukupi kebutuhan optimal ikan maka kedua pakan ini menghasilkan laju pertumbuhan relatif ikan yang rendah. Hal ini juga sangat berkaitan dengan efisiensi pakan, dimana berdasarkan data (Tabel 5) ikan uji pada perlakuan ini memiliki nilai efisiensi yang paling rendah. Reis et al. (1988) yang meneliti ikan chanel catfish juga menemukan bahwa ikan yang diberi pakan dengan kadar protein yang rendah mempunyai pertumbuhan yang rendah yang disebabkan karena rendahnya protein yang masuk ke dalam tubuh ikan.
Perlakuan pakan dengan level rasio energi protein yang tinggi ternyata tidak memberikan pengaruh pertumbuhan yang terbaik bagi ikan. Perlakuan A4B2 dengan kadar protein 45% dan rasio energi protein 11 kkal GE/g menghasilkan pertumbuhan ikan kembali menjadi rendah dibandingkan dengan perlakuan dengan level protein yang sama namun dengan rasio energi protein yang lebih rendah (9 kkal GE/g). Demikian pula halnya yang terjadi dengan perlakuan lainnya kecuali pada perlakuan A1B1(33;9) dan A1B2 (33;11) dimana rasio energi yang lebih tinggi menghasilkan pertumbuhan yang lebih baik pada level protein yang sama (level protein paling rendah dalam penelitian ini). Rendahnya pertumbuhan ikan pada perlakuan rasio energi yang lebih tinggi pada level protein yang sama kemungkinan disebabkan tingginya energi total pakan terutama yang berasal dari lemak (Tabel 2). Hal ini karena kandungan lemak dalam pakan juga menentukan pemanfaatan protein sebagai sumber energi untuk pembentukan jaringan dan aktivitas ikan. Peres dan Teles (1999) mengemukakan bahwa lemak yang tinggi dalam pakan tidak meningkatkan pertumbuhan dan efisiensi pakan
pada ikan kakap Eropa Dicentrarchus labrax yang diberi pakan dengan kandungan lemak 12, 18, 24, dan 30%. Hal yang sama dikemukakan oleh Webster et al. (1995) bahwa pertumbuhan dan konversi pakan ikan sunshine bass yang diberi lemak rendah (6%), ternyata lebih tinggi dibandingkan dengan ikan yang diberi pakan dengan kandungan lemak tinggi (9, 16, dan 17%). Lovell (1988) dan Alanara (1994) menyatakan bahwa pakan berenergi tinggi karena keberadaan lemak yang tinggi tidak memberikan pertumbuhan terbaik, karena kadar lemak yang tinggi menyebabkan konsumsi pakan ikan menjadi rendah. Selanjutnya hasil penelitian Seenappa dan Devaraj (1995) pertumbuhan ikan Indian major carps yang diberi lemak rendah (4%) lebih tinggi dibandingkan dengan ikan yang diberi lemak tinggi (8% dan 12%) yang disebabkan oleh konsumsi pakan yang rendah. Hasil penelitian Hung et al. 1997 pada ikan white sturgeon Acipenser
transmontanus yang diberi pakan dengan kandungan lipid tinggi akan
menyebabkan laju pertumbuhan spesifiknya juga paling rendah, sedangkan hasil penelitian Suwirya et al. (2007) ikan kuwe Gnathanodon speciosus tidak dapat memanfaatkan lemak secara efektif sebagai sumber energi dalam pertumbuhan. Hal ini dibuktikan dengan data tingkat konsumsi pakan dan efisiensi pakan pada penelitian ini (Tabel 5) dimana pada perlakuan dengan rasio energi yang lebih tinggi pada level protein yang sama tingkat konsumsi dan efisiensi pakan ikan uji cenderung menurun. Keadaan ini akan membatasi banyaknya protein yang masuk ke dalam tubuh ikan sehingga protein yang disimpan menjadi protein tubuh lebih sedikit, akibatnya pertumbuhan ikan menjadi rendah. Rosenlund et al. (2004) melaporkan adanya pemanfaatan protein yang meningkat apabila kadar protein diturunkan dan kadar lemak ditingkatkan pada kadar tertentu. Namun pada penelitian ini ini, efek tersebut tidak tampak pada ikan yang mendapat lemak tinggi. Ini mungkin karena ikan yang mengkonsumsi pakan dengan lemak lebih rendah akan mendapatkan protein lebih banyak, karena konsumsi pakan yang lebih banyak.
Perbedaan kadar protein dan rasio energi protein dalam pakan juga berpengaruh pada kelangsungan hidup ikan uji. Pada kadar protein A4 dan A3, tingkat kelangsungan hidup ikan berada pada kisaran 86 - 100% yang berdasarkan hasil analisis statistika ketiga perlakuan ini tidak menunjukkan hasil yang berbeda
nyata. Pada perlakuan dengan level protein rendah (A1 dan A2) tingkat kelangsungan hidup ikan uji hanya berada pada kisaran 46 – 76 % dan berbeda nyata dengan perlakuan dengan level protein yang lebih tinggi (A3 dan A4). Hal ini menunjukkan bahwa kadar protein 33% dan 37% tidak cukup bagi ikan kuwe untuk menjamin berlangsungnya seluruh proses metabolisme sel dalam tubuh, walaupun pada perlakuan dengan level protein ini masih terlihat adanya pertumbuhan. Beberapa peneliti melaporkan kebutuhan protein beberapa jenis ikan karnivora laut berkisar antara 40 – 55 % dan bervariasi menurut spesiesnya. Suwirya et al. (2007) menyatakan bahwa ikan kuwe membutuhkan protein 42% - 46%, sementara ikan kakap merah membutuhkan pakan dengan kadar protein 42.5% (SEAFDEC, 1998). Dalam keadaan kekurangan protein maka fungsi lain protein sebagai penyusun utama enzim dan antibodi untuk pertahanan terhadap penyakit menjadi berkurang. Lovell (1989) menambahkan bahwa protein dapat digunakan sebagai sumber energi jika kebutuhan energi dari lemak dan karbohidrat tidak mencukupi dan juga sebagai penyusun utama enzim, hormon dan antibodi. Selanjutnya Anonim (2007) mengemukakan bahwa keseimbangan protein dalam formulasi pakan sangat penting karena berperan besar dalam kesintasan, pertumbuhan, serta ketahanan tubuh ikan, terutama pada stadia larva. Berdasarkan hal ini maka rendahnya tingkat kelangsungan hidup pada perlakuan dengan kadar protein A1 dan B2 diduga disebabkan karena berkurangnya ketahanan tubuh ikan sebagai akibat dari kekurangan protein.
Hasil analisa kandungan glikogen hati dan otot ikan uji (Tabel 6) menunjukkan bahwa pemberian pakan dengan kadar protein dan rasio energi protein yang berbeda berpengaruh terhadap kadar glikogen hati pada setiap perlakuan. Kadar glikogen hati ikan uji cenderung naik pada semua perlakuan dengan rasio energi protein yang tinggi pada level protein A2 (37%) dan A3 (41%). Pada level protein A4 (45%) kenaikan kadar glikogen hati terjadi pada kedua level rasio energi protein. Naiknya kadar glikogen hati pada perlakuan dengan rasio energi protein tinggi kemungkinan disebabkan oleh besarnya total energi dalam pakan sehingga cadangan energi dalam bentuk glikogen dalam hati tidak dimanfaatkan oleh ikan. Selain itu tingginya kadar BETN (Tabel 2) dalam pakan pada perlakuan A1B2, A2B2, A3B2 dan A4B1 berakibat pada tingginya
nilai kadar glikogen hati pada ke-tiga perlakuan tersebut. Subandiyono (2009) mengemukakan bahwa pemberian pakan dengan tingkat karbohidrat yang tinggi dalam waktu yang lama pada ikan karnivora akan menyebabkan pertumbuhan ikan terhambat dan meningkatkan kadar glikogen hati. Berbeda dengan mamalia yang dengan segera dapat memanfaatkan cadangan energi dalam bentuk glikogen, Ikan tidak dapat dengan cepat memobilisasi cadangan glikogen yang tersimpan dalam hati saat ikan tersebut kelaparan. Ikan cenderung merombak protein dan lemak menjadi energi terlebih dahulu lewat proses glukoneogenesis. Hal ini diperkuat dengan pernyatan Tacon & Cowey (1985) serta Halver (1989) yang mengemukakan bahwa ikan cenderung menggunakan protein sebagai sumber energi dibandingkan lemak dan karbohidrat.
Lemak hati ikan uji cenderung meningkat seiring dengan peningkatan kadar protein pakan sampai pada level protein 37% (A2) dan kemudian sedikit menurun pada level protein 41% (A3) dan 45% (A4). Peningkatan kadar lemak hati akan semakin tinggi pada level rasio energi protein 11 kkal GE/g (B2) pada semua perlakuan. Peningkatan kadar lemak hati yang cukup tinggi pada level protein yang lebih rendah dan rasio energi yang lebih tinggi diduga akibat ikan yang mengkonsumsi pakan dengan protein dan rasio energi tinggi cenderung akan menyimpan energi lebih banyak pada jaringan tubuhnya. Hatlen et al. (2007) mengemukakan bahwa ikan yang mengkonsumsi pakan rendah protein akan menyimpan energi lebih banyak pada hati (berupa lemak) dan lebih sedikit pada jaringannya (protein) daripada yang mengkonsumsi pakan tinggi protein. Hal ini sesuai dengan data hasil analisis (Tabel 6), dimana ikan uji yang diberi perlakuan pakan dengan kadar protein yang lebih tinggi (A3B1, A3B2, A4B1 dan A4B2) memiliki kadar glikogen otot dan lemak otot yang cenderung lebih tinggi dibandingkan dengan ikan uji yang diberi pakan dengan kadar protein yang lebih rendah (A1B1, A1B2, A2B1, A2B2).
Dari hasil pengukuran hepatosomatik indeks (HSI) terlihat bahwa nilainya berada pada kisaran 1,38-1,67. Nilai ini didapat dari perbandingan antara berat hati dan berat tubuh ikan dalam bobot basah. Nilai HSI cenderung meningkat seiring dengan peningkatan kadar protein dan rasio energi protein serta peningkatan bobot ikan pada masing-masing perlakuan yang secara langsung mempengaruhi tingkat
konsumsi pakan. Peningkatan jumlah konsumsi pakan menyebabkan jumlah nutrien yang diserap meningkat sehingga nilai HSI meningkat. Peningkatan HSI yang seiring dengan peningkatan kadar protein dan rasio energi protein menunjukkan bahwa dengan meningkatnya jumlah nutrien yang diserap menyebabkan jumlah nutrient yang terakumulasi pada hati meningkat. Dalam penelitian ini tidak terlihat adanya penumpukan lemak pada hati yang dapat menyebabkan peningkatan nilai HSI secara tidak normal. Pembengkakan hati biasanya disebabkan oleh peningkatan kadar lemak di hati serta defisiensi lemak. Tacon (1992) menyebutkan bahwa pembengkakan dan perubahan warna organ hati menjadi lebih pucat disebabkan oleh defisiensi asam lemak akibat kurang baiknya komposisi pakan yang digunakan.
Data Total eskresi ammonia ikan uji pada Tabel 7 menunjukkan adanya perbedaan nilai pada masing-masing perlakuan. Eskresi ammonia sangat berhubungan dengan katabolisme asam amino yang digunakan sebagai energi. Ikan akan mengeluarkan nitrogen yang dihasilkan dari katabolisme asam amino ke dalam air sebagai amonia dari insang (Halver dan Hardy, 2002). Semakin tinggi nilai TAN menunjukkan semakin banyaknya protein yang dideaminasi. Dalam penelitian ini, eskresi total amonia nitrogen yang diukur selama tiga jam menunjukkan adanya peningkatan seiring dengan meningkatnya kadar protein pakan pada rasio energi pakan yang lebih rendah (9 Kkal GE/g). Nilai eskresi total amonia nitrogen tertinggi diperoleh pada perlakuan A4B1 (45:9) yang merupakan perlakuan pakan dengan kadar protein tertinggi dan rasio energi protein yang lebih rendah. Pada level protein yang sama (45%) nilai eskresi total ammonia nitrogen cenderung menurun pada rasio energi yang lebih tinggi. Hasil penelitian ini sesuai dengan pendapat Adelina at al. (2000) bahwa peningkatan protein pakan akan meningkatkan eskresi amonia, akan tetapi eskresi ammonia akan semakin kecil dengan bertambahnya rasio energi protein. Ming (1985) menyatakan bahwa eskresi amonia meningkat dengan cepat sebagai respon terhadap penambahan protein pakan. Selanjutnya Degani et al. (1985) mengemukakan bahwa produksi amonia berkolerasi secara linier dengan kadar protein pakan.