BAB IV
HUBUNGAN ANTARA JAR AK GENETIK AN TAR TETUA DAN
HIBRIDA F1 KAKAO DENGAN RESISTENSI TERHADAP IN FEKSI
Phytophthora palmivora Butl.
Abstrak
Peningkatan permintaan biji kakao mendorong tersedianya bibit kakao yang unggul da n be rmutu, yaitu bibit yang berda ya hasil tinggi da n tahan terhadap penyakit khusus nya pe nyakit busuk buah yang disebabka n oleh Phytophthora palmivora. Salah satu upa ya untuk mendapatka n bibit tersebut adalah dengan melakukan penyilangan antara tetua yang memiliki daya hasil tinggi dengan yang tahan terhadap P.palmivora. Salah satu keberhasilan penyilangan ditentukan oleh keragaman sifat dari tetua yang disilangkan. Pada penelitian sebelumnya telah dipilih lima tetua de ngan ke tahanan terhadap infeks i busuk buah da n da ya hasil yang tinggi dan digunakan untuk memproduksi satu rangkaian populasi hibrida F1 dengan metode semi-dialel. Tetua terpilih yang digunakan dalam persilangan tersebut adalah klon kakao ICCRI 3, TSH 858, ICS 13, DR1 dan Sca 6. Penelitian ini bertuj uan untuk menganalisis jarak genetik antar tetua da n antar individu pada populasi hibrida kakao berdasarkan marka SSR, juga mempelajari hubungan antara jarak genetik antar tetua dan jarak genetik antar individu hibrida F1 dengan sifat resistensi terhadap penyakit busuk buah akibat infeksi P. palmivora. Hasil analisis klaster menunjukkan ba hwa tingkat kesamaan di antara lima klon kakao yang dijadikan sebagai tetua bervariasi antara 0.15 – 0.48. Hal ini berarti jarak genetik antar tetua yang digunakan untuk menghasilkan galur kakao hibrida F1 memiliki nilai yang relatif tinggi, yaitu berkisar antara 0.52 – 0.85. Semakin jauh jarak genetic diharapkan keragaman individu hibrida semakin tinggi. Hasil analisis SSR pada sepuluh silangan individu hibrida, menunjukkan bahwa karagaman genetik individu hibrida relatif tinggi. Tetua dengan jarak genetic yang jauh ternyata juga menghasilkan hibrida dengan keragaman yang relatif tinggi.
GENETIC DISTANCE AND THE RELATIONS WITH CACAO RESISTANCE AGAINST Phytophthora palmivora Butl.
Abstrac t
Demands for cocoa beans have encourage the availability of cocoa seedlings that have superior quality, high yield and resistant to diseases, especially pod rot caused by Phytophthora palmivora. One of the efforts to get these kinds of seeds is by crossing the parents which have a high yield with the parents which have a resistance to P.palmivora. The success of the crossing is determined by the diversity of parents. Before, it has selected five parents which have high yield and resistance to black pod infection to be used to produce a series of F1 hybrid population using semi-dialel method. Selected parents used in these crosses are clones of cocoa ICCRI 3, TSH 858, ICS 13, DR1 and SCA 6. The aims of this study is to analyze the genetic distance between parents and among individuals in a population of hybrid cocoa based on SSR markers, and also to study the relationship of genetic distance among the parents and genetic distance between the F1 hybrid individuals with the nature of resistance to black pod disease caused by infection with P. palmivora. The results of cluster analysis showed that the degree of similarity among the five parents varies between 0.15 to 0.48. This means that the genetic distance between the parents used to produce F1 hybrid strains of cocoa has a relatively high value, which ranges from 0.52 0.85. The farther the distance of genetic between the parents the higher genetic diversity of hybrid individuals is expected. SSR analysis results of ten individual crossed hybrids, suggests that individual genetic hybrid has a relatively high genetic diversity. Parents which have a further genetic distance were also produced hybrids with a relatively high diversity.
Pendahuluan
Tanaman kakao telah lama dibudidayakan di Indonesia, baik oleh masyarakat maupun perkebunan yang dikelola perusahaan swasta ataupun pemerintah. Indonesia merupakan salah satu negara yang memiliki areal pertanaman kakao paling luas di dunia dengan luas areal penenaman mencapai 917 ribu hektar dan teersebar di seluruh provinsi kecuali DKI Jakarta. Sealain itu Indonesia juga termasuk negara penghasil kakao terbesar ke tiga setelah Pantai Gading dan Ghana, dengan nilai produksi tahunan mencapai 572 ribu ton (Wahyudi dan Raharjo 2008). Akan tetapi di pasaran internasional harga kakao asal Indonesia masih rendah karena kualitas bijinya yang kurang baik, yaitu
didominasi oleh biji-biji yang tanpa fermentasi dan kadar kotoran tinggi serta terko ntaminasi serangga, jamur, atau mikotok sin.
Hal tersebut tentunya menjadi keprihatinan kita bersama untuk mengatasinya, karena dengan potensi lahan yang luas sangat memungkinkaan untuk meningkatkan kualitas biji yang dihasilkan tanaman kakao produksi negara kita. Dengan demikian maka peluang Indonesia sebagai negara pengekspor biji kakao terbuka dengan lebar, sehingga dapat meningkatkan devisa negara.
Hingga saat ini pengembangan budi daya kakao masih menemui berbagai kendala, antara lain serangan hama dan penyakit yang dapat menurunkan kualitas serta kuantitas produksi biji kakao. Berbagai penyakit yang disebabkan oleh patogen dapat menyerang tanaman kakao, salah satunya adalah penyakit busuk buah (Black pod) yang disebabkan oleh cendawan Phytophthora palmivora. Serangan penyakit busuk buah ini menyebabkan kerugian yang bervariasi antara daerah satu dengan yang lainnya bahkan antar negara. Secara umum, kerugian antara 20-30% pertahun terjadi akibat serangan penyekit busuk buah pada pertanaman kakao di lapangan (Wood dan Lass 1985).
Pengadaan bibit yang unggul dan bermutu merupakan salah satu jalan ke luar yang dapat ditempuh untuk mengatasi masalah tersebut. Usaha-usaha yang harus dilakukan untuk memperoleh bahan tanam yang unggul antara lain dengan melakukan pemuliaan tanaman kakao meliputi rangkaian kegiatan seperti memperisapkan koleksi plasma nutfah, melakukan pengujian klon, menyilangkan antar klon, dan melakukan pengujian hasil silangan klon (Winarno 2008).
Salah satu upa ya yang telah dilakuka n Pusat Penelitian Kop i da n Kakao Indonesia adalah dengan melakukan penyilangan antara klon-klon kakao yang resisten terhadap penyakit sebagai tetua donor dengan kon-klon yang berda ya hasil tinggi sebagai tetua resipien. Dengan demikian diharapkan akan diperoleh klon-klon baru yang memiliki perpaduan karakter tersebut. Untuk mendapatkan hibrida yang sesuai de ngan yang diharapka n maka salah satu faktor penentunya adalah pemilihan tetua yang tepat. Untuk itu dipe rluka n infor masi yang akurat tentang keragaman genetik plasma nutfah kakao yang akan dipilih sebagai tetua. Pengetahuan tentang keragaman ge netik di antara klon-klon kakao yang biasa ditanaman, juga jenis-jenis gen yang terlibat dalam ketahanan terhadap penyakit,
daya hasil, kualitas, dan karakter agronomis lainnya sangat diperluka n oleh para pemulia kakao untuk menyeleksi klon-klon tetua secara efektif dan efisien (Martinez et al. 2006).
Dalam kegiatan yang dilakukan pada penelitian sebelumnya mengenai persilangan untuk perakitan varietas baru dari tetua yang memiliki deversitas genetik tinggi resistensi terhadap hama PBK, Penyakit Busuk buah, VSD dan produksi tinggi, telah dipilih lima tetua dengan ke tahanan terhadap infeks i busuk buah dan daya hasil yang tinggi dan digunakan untuk memproduksi satu rangkaian populasi hibrida F1 dengan metode semi-dialel (Rubiyo 2009). Tetua terpilih yang digunakan dalam persilangan tersebut adalah klon kakao ICCRI 3, TSH 858, ICS 13, DR1 dan Sca 6. Untuk menguji apakah pilihan ke-lima tetua tersebut telah dilakukan dengan benar atau belum, DNA dari lima klon kakao yang digunakan untuk menghasilkan hibrida F1 telah diisolasi dan dianalisis menggunakan marker SSR pada lokus- lokus yang telah digunakan sebelumnya. Data yang dihasilkan selanjutnya dianalisis untuk mendapatkan informasi tingkat kesamaan antar klon dan digunakan untuk mengelompokkan masing- masing klon dengan analisis klaster.
Kegiatan penelitian bertujuan unt uk mempelajari:` (1) menganalisis jarak genetik antar tetua yang digunakan dalam persilangan untuk menghasilkan populasi hibrida F1 berdasarkan marka SSR; (2) Jarak genetik antar individu hibrida F1 hasil persilangan antara dua klon kakao terpilih berdasarkan marka SSR; (3) Hubungan antara jarak genetik antar tetua dan jarak genetik antar individu hibrida F1 dengan sifat resistensi terhadap penyakit busuk buah akibat infeksi P. palmivora.
Bahan dan Metode
Waktu dan Tempat Penelitian
Penelitian dilaksanakan di laboratorium Biologi Molekuler Tanaman, Departemen Agronomi da n Hortikultura, Fakultas Pertanian Intitut Pertanian Bogor, mulai bulan Juli 2009 sampai dengan Juli 2010.
DNA diisolasi dari tanaman F1 hasil silangan antar lima tetua yang menghasilkan sepuluh hibrida (Tabe l 6), diperoleh dari hasil penelitian Rubiyo (2009), masing- masing diambil 10 individu tanaman unt uk dianalisis. Sebanyak 2 gram daun segar dipotong-potong halus dan dimasukkan ke dalam tabung ependorf 2 ml yang telah berisi buffer ekstraksi.
Tabe l 6. Tetua yang digunakan dalam persilangan hibrida kakao
TETUA ICCRI 3 TSH 858 DR 1 ICS 13 Sca 6
ICCRI 3 1 2 3 4
TSH 858 5 6 7
DR 1 8 9
ICS 13 10
Sca 6
Total genomic DNA diekstraksi berdasarkan prosedur Cetye Trietye Ammonium Bromide (CTAB) dari wilkie (1997 ) yang dimod ifikasi. Daun yang telah direndam dalam bufer ekstraksi digerus di dalam mortar bersama dengan 1,5 ml buffer ekstraksi (2% CTAB; 1,4 M NaCl; 40 mM EDTA; 100 mM Tris-HCl pH 8), 1% B- mercaptoethanol dan 1% pvp, setelah halus disimpan pada tabung miccrotube 2 ml. Selanjutnya diinkubasi pada suhu 65oC selama 30 menit dalam kloroform isoamil alcohol/ CIA (24:1) sebanyak 2 x volume ke dalam masing-masing tabung kemudian divortek selama 3 menit, dan disentrifugasi 12.000 rpm selama 10 menit. Supernatan dipindahkan ke tabung baru, ditambahkan 1 ml kloroform; Isoamil alcohol dan disentrifugasi lagi seperti sebelumnya. Supernatan ke mud ian dipinda hkan ke tabung baru dan selanjutnya ditambahkan 2-propanol dingin sambil dibo lak-balik tabungnya hingga terbentuk gumpalan putih, kemudian diinkubasi dalam freezer pada suhu selama 30 menit. Presipitasi DNA dengan 3 M sodium asetat (pH 8.0) dan ditambahkan 1 ml etanol absolute dingin, pellet dicuci dengan etanol 70 % kemudian dikeringkan dengan membalik tabung di atas kertas tissue. DNA dillarutka n dalam 300 ul TE (10 mM Tris-HCl; 0,1 mM EDTA (pH 8.0). DNA yang sudah diisolasi siap untuk PCR.
PCR dan Analisis SSR
Template DNA dari masing- masing individu kakao diuji dengan 22 primer SSR yang telah dikembangkan oleh Pugh et al (2004) dan dipilih dari hasil penelitian sebelumnya (Tabel 7). PCR dilakukan dengan total volume 15 µl, terdiri atas 2 µl DN A template, 1 µl primer, 1.5 µl 10x buffer, 0.15 µl MgCl2,
0.075 Taq DN A polymerase, dan 2.7 µl dNTP
Tabe l 7. Microsatellite (SSR) yang digunakan unt uk mengamplifikasi genomic DNA F1 cacao
No LG Nama M arker
Primer Jml Basa PCR size
(bp)
Repeat Sekuens
Tm (oC)
1 1 mT cCIR144 F: AACCACT GACACGCAAT GAA R: T GTTT GCAAAT AAAGAAGAGAGGA
20 24
242 (CT )2TTT(CT)9 60.16 59.45 2 1 mT cCIR184 F: ACT GCT GCAGCCT CT CTTTC
R: ACAT GGAGGGAGGGAGA GAT
20 20
204 (CA)8(CT)13 59.90 59.89 3 2 mT cCIR162 F: GACCTTTTTCCCCCT GATTC
R: T GGCAAAAATTCACCAGT CA
20 20
250 (GA)19 59.74 60.09 4 3 mT cCIR167 F: AAT CGGT GCAT GGTAGAACC
R: AGCATAGT GT CGTTTCT GTT GC
20 22
244 (GA)18 59.82 59.45 5 3 mT cCIR198 F: GGGACCAT AAGGAAAT CAT GC
F: GCTT GCCCAGGT GAAGT AAG
21 20
192 (CA)3TA(CA)6 60.53 59.88 6 4 mT cCIR95 R: GTT CTCGACAT GGGCT CCTA
F: T GCAT GGAT GCT GAAACAAG
20 20
237 (T C)4CC(T C)21 60.22 60.81 7 4 mT cCIR213 F: T CCAAT GTT GAT CT CGCAAA
R: TTTTCATTCCT GCTT GCGT A
20 20
186 (CT )26 60.20 59.44 8 4 mT cCIR67 F: GGTT CT CGCTT GAAAAT CCA
R: CCT CTTTTCCAAGCCT CCAT
20 20
176 (CT )7(CA)12 60.19 60.57 9 5 mT cCIR69 F: GGACAT CGGT GTT CCATCAG
R: T GCTAT GAGATT GAAAGAGAATT GA 20 25
208 (CT )20 61.36 59.43 10 5 mT cCIR109 F: CCCGT AAGCTT CCATTTT CC
R: CAAAGGGACCAAAAAGAGCA
20 20
221 (CT)12 60.79 60.22 11 5 mT cCIR10 F: CGAATT GACAGAT GGCCT ACA
R: CCCAAGCAAGCCT CATACT C
21 20
122 (T G)13 61.04 59.84 12 6 mT cCIR16, F: CTT CACCAGCT CACCGAT CT
R: AT CAAT GGGTTCGGGT AGT G
20 20
207 (CT )13 60.41 59.67 13 6 mT cCIR255 F: GCCTTACAGCATT CCCAT GA
R: AT CT GCAGGACTT GGACCAC 20 20 193 (AC)11 61.00 60.12 14 6 mT cCIR276 F: T GT GT GTTTAATT GCT CCT GCT R: T GT CT GCCCTTTGACCTTTC 22 20 199 (GA)14 59.81 60.23 15 6 mT cCIR291 F: TT GCAATT GTCCCAAGCAT A
R: AT GT CAAGCAT GGCAGT GTT
20 20
212 (CT )12 60.07 59.17 16 7 mT cCIR190 F: CT GAAGCACAATTATTCCAT CAA
R: CCAATT GCT CCACAAAGAGC
23 20
172 (T G)12 59.13 60.78
17 8 mT cCIR211 F: GGGATT GCACTTCACAAGGT R: T CCAAGTT CCGT AT GT GCT G 20 20 179 (CT )9 59.97 59.72 18 8 mT cCIR218 F: CAT GCGTT GACCAAGGAAG
R: AT CAAT GCAT GGGAACACCT
19 20
181 (CT )11 60.25 60.20 19 9 mT cCIR251 F: T CAT GCCCAGT GACACAAAT
R: AAT GGACT GGAGCAT GGAAG
20 20
228 (CT )7(CA)12 59.97 60.07 20 10 mT cCIR91 F: GCCCAT GCTT CTCTT CAT GT
R; GGGAAAT GAGAAGGGT GT GA 20 20 191 (CT )10 60.23 59.90 21 10 mT cCIR155 F: CTTAGAGGCTT GT GCCGT GA R: GCCAT GCCAATTTCCAAT AA 20 20 197 (T C)12 61.50 60.65 22 10 mT cCIR209 F: T GT CCTT CACATAAGCCAT GA R: T GTT GCCCTTCCTT GTTAGG 21 20 243 (GT)6AT(GA)9 59.14 60.10
Proses PCR dilakukan dengan 39 siklus, diawali denaturasi pada 94 oC selama 4 menit, kemudian 39 siklus berikutnya yang terdiri atas denaturasi pada 94 oC selama 45 detik, penempelan (annealing) pada suhu 55 oC selama 45 detik, dan perpanjangan (extension) pada suhu 72 oC selama 30 detik. Tahap terakhir dilanjutkan dengan perpanjangan akhir (final extension) pada suhu 72 oC selama 5 menit dan pendinginan (cooling) sampai suhu 16 o
Hasil amplifikasi PCR dievaluasi de ngan melakukan runni ng pada gel agarose 1% selama 20 menit pada mesin elektroforesis dengan arus 300 Am dan 200 Volt untuk konfirmasi ada tidaknya produk amplifikasi.
C selama 5 menit.
Running PAGE DNA hasil amplifikasi dengan SSR
Proses elektroforesis dimulai dengan penyiapan kaca. Pertama disiapkan dua kaca panjang dan pendek. Kaca panjang dibersihkan dengan etanol 95%, dan dilanjutkan dengan clear view. Untuk kaca pendek dibersihkan dengan etanol 95%, dilanjutkan dengan campuran antara Bind Silan e dan asam asetat. Kemudian, dua kaca disusun dengan diberi plat plastik sebagai pembatas dan dijepit dengan empat pasang penjepit. Sementara itu, disiapka n “Microsatelite gel polyacrilamide” yang terdiri atas: 60 ml 4.5% acrilamide ditambah 10% APS µl dan 90 µl TEMED, kemudian diaduk perlahan sampai tercampur. Cairan acrilamide selanjutnya dituangkan secara perlahan ke celah diantara dua kaca, sisipka n sisirnya dan dibiarkan sampai gel terpolimerisasi.
Sampel berupa produk PCR sebanyak 2 µl ditambah 3-4 µl pewarna kemudian diaduk dan didenaturasi pada suhu 94° C selama 10 menit. Plat kaca
kemudian dipasang pada mesin elektroforesis, dan dilakukan pre running elektroforesis dengan buffer 1X TBE hingga mencapai suhu 60o
Pewarnaan perak nitrat dimulai dengan perendaman kaca dalam larutan acetic acid 10% selama 30 menit. Selanjutnya dicuci dengan akuades sebanyak 3 x masing- masing selama 10 menit, kemudian dilakukan perendaman pada larutan perak nitrat selama 30 menit, selanjutnya dicuci dengan akuades. Selanjutnya kaca direndam pada larutan developer sambil digoyang hingga nampak pita. Setelah nampak pita, larutan developer dibuang dan terakhir dicuci dengan acetic acid selama 5 detik dan dilanjutkan pencucian terakhir dengan air.
C. 2 µl sampel hasil amplifikasi dimasukkan pada celah sisir sampai semua sampel masuk, dan sampel siap di running pada mesin elektroforesis dengan kondisi 3000 volt, 300 mA, 65 Watt selama 2,5 jam. Setelah selesai, kaca dikeluarkan dan lepaskan kaca penutupnya dengan hati- hati. Hasil elektroforesis siap untuk d iwarnai.
Skoring dan analisis data
Skoring dilakukan terhadap posisi alel hasil amplifikasi PCR untuk masing- masing genotipe pada tiap lokus SSR. Hasil skor ing dilakukan analisis untuk melihat keragaman genetik dengan menggunakan software NTSys, sedangkan tingkat heterosigositas ditentukan dengan menggunakan software CERVUS.
Data hasil analisis klastering digabungkan denga n data respo n ketahanan terhadap P. palmivora yang diperoleh dari hasil penelitian sebelumnya (Rubiyo, 2009). Respon ketahanan ditentukan berdasarkan luas bercak daun yang muncul pada hari ke-6 setelah diinokulasi dengan P. palmivora. TH: tahan; AT: agak tahan; AR: agak rentan; RT: rentan; SR: sangat rentan terhadap infeksi P. palmivora menguunakan metode uji yang dilakukan.
Has il dan pe mbahas an
Analisis SSR
Amplifikasi DNA dilakukan dengan menggunakan 22 primer yang sebelumnya telah digunakan dalam mengamplifikasi DNA tetua. Pada amplifikasi DNA tetua digunakan 39 primer SSR, akan tetapi hanya 22 primer yang
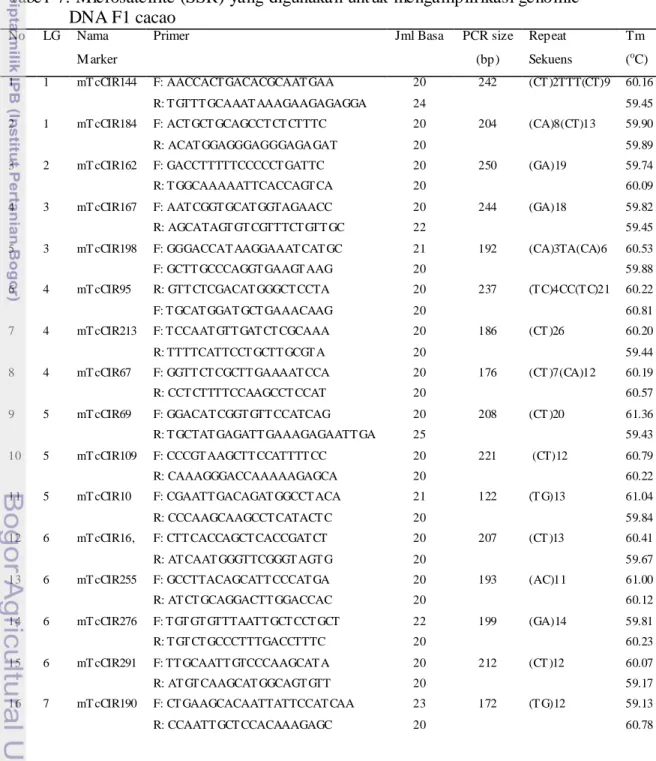
menunjukkan pita polimorfik untuk lima tetua yang digunakan. Hasil PCR menunjukkan bahwa seluruh primer dapat mengamplifikasi sepuluh hibrida (H1 sampai H10), meskipun tidak untuk semua individu. Hal ini dapat dilihat dari hasil running pada gel agarose (Gambar 12).
Gambar 12. Contoh Hasil PCR 10 hibrida F1 dengan Primer MTcCIR 67 pada gel agarose, M = marka
Hasil analisis tingkat kesamaan genetik antar lima klon tetua yang digunakan dapat dilihat pada Tabel 8, sedangkan hasil analisis klaster untuk pengelompokkan lima klon kakao yang digunakan sebagai tetua disajikan pada Gambar 13.
Tabe l 8. Tingkat kesamaan 5 klon kakao terpilih yang digunakan untuk menghasilkan populasi F1.
Klon kakao ICCRI3 TSH858 DR1 ICS13 Sca6 ICCRI3 -
TSH858 0.33 -
DR1 0.33 0.43 -
ICS13 0.40 0.48 0.33 -
Sca6 0.22 0.32 0.17 0.15 -
Keterangan: Tingkat kesamaan dianalisis berdasarkan keragaman alel dengan menggunakan sejumlah lokus marker SSR yang telah digunakan sebelumnya.
Dari Tabel 8 dapat diketahui bahwa tingkat kesamaan di antara lima klon kakao yang dijadikan sebagai tetua bervariasi antara 0.15 – 0.48. Hal ini berarti jarak genetik antar tetua yang digunakan untuk menghasilkan galur kakao hibrida F1 berkisar antara 0.52 – 0.85 yang menunjukkan bahwa nilainya relatif tinggi. Dari informasi jarak genetik antar tetua tersebut mengindikasikan bahwa populasi F1 yang dihasilkan akan mempunyai tingkat keragaman genetik yang tinggi. Keragaman genetik antar individu dalam populasi F1 yang dihasilkan diduga akan
H1 H2 H3 H4 H5 M H6 H7 H8 H9 H10
mempunyai nilai tinggi, terutama dari hasil kombinasi persilangan antara: klon kakao DR 1 x Sca 6, ICS 13 x Sca 6, dan ICCRI 3 x Sca 6.
Gambar 13. Pengelompokkan lima klon tetua untuk menghasilkan populasi F1 berdasarkan marka molekuler SSR dan berdasarkan responsnya terhadap infeksi Phytophthora palmivora.
Ketiga famili hibrida F1 hasil persilangan antara ketiga pasangan tetua tersebut diduga akan mengasilkan populasi F1 dengan keragaman genetik yang tinggi. Selain itu, karena dalam persilangan tersebut digunakan klon kakao Sca 6 (yang diketahui seba gai klon resisten) sebagai do nor sifat-sifat resistensi terhadap hama dan penyakitkya, maka besar kemungkinan populasi hibrida F1 hasil persilangan antara ketiga pasangan tetua tersebut diduga akan menghasilkan populasi F1 dengan ketahanan terhadap infeksi penyakit dan hama PBK.
Hasil analisis klaster yang dilakukan juga menjelaskan bahwa Sca 6 mempunyai tingkat kesamaan yang paling rendah dengan empat klon kakao yang lain. Sebaliknya, klon kakao TSH 858 dan ICS 13 mempunyai tingkat kesamaan yang tertinggi diantara lima tetua yang dievaluasi (Gambar 13). Berdasarkan hal tersebut, akan menjadi menarik untuk melihat keragaman yang ada pada individu-individu dalam populasi hibrida F1 turunan pasangan kombinasi tetua tertentu dan hubungannya dengan jarak genetik pasangan tetuanya .
Keragaman hibrida F1
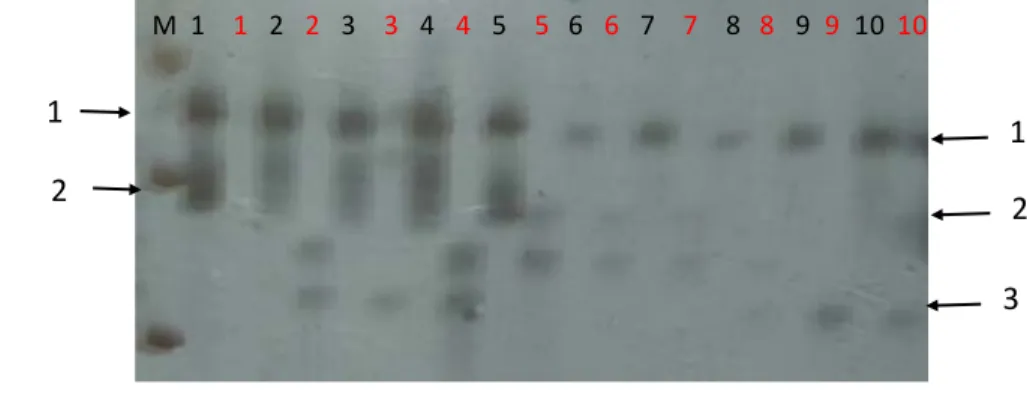
Hasil running pada gel akrilamid menunjukkan adanya keragaman pada individu- individu dalam populasi hibrida F1 (gambar 14).
Gambar 14. Contoh elektroferogram hasil PCR menggunakan primer SSR mTcCIR 167 (P1) dan mTcCIR 10 (P2) dengan 10 contoh DNA kakao F1. Amplifikasi dengan primer P1 (lokus P1) menghasilkan dua macam pita yang berbeda ukuran (masing- masing menjadi alel 1 dan alel 2 pada lokus P1). Primer P2 (lokus P2) menghasilkan tiga macam pita yang berbeda (masing- masing menjadi alel 1. Alel 2, dan alel 3 pada lokus P2).
Adanya keragaman di antara individu pada hasil silangan dapat terjadi karena berbagai faktor, salah satunya adalah adanya rekombinasi alel terutama pada genotipe yang heterozigot. Apabila dua tetua yang masing- masing memiliki dua alel heterozigot disilangkan, maka akan menghasilkan individu dengan empat ke mungkinan variasi genotipe (misalnya P1 memiliki alel “1” dan “2” disilangkan dengan P2 yang memiliki “3” dan “4”, maka hibrida F1 yang dihasilkan memiliki empat kemungkinan kombinasi alel yaitu A:“1” da n “3”; B: “1” da n “4”; C: “2” dan “3”; D:”2” da n “4”.
Seperti telah dijelaskan pada bagian sebelumnya bahwa selain tingkat heteros igos itas, hal lain yang perlu dipe rhatika n da lam menent uka n tetua untuk persilangan adalah ada tidaknya kesamaan alel dalam masing- masing primer yang digunakan untuk menghasilkan marka molekuler SSR. Berkaitan de ngan hal tersebut, klon yang diharapkan menghasilkan hibrida yang baik adalah klon-klon yang tingkat heterosigositasnya tinggi dan kombinasi alel dalam setiap primernya berbeda. Sebagai contoh: klon ICCRI 3 mempunyai kombinasi alel 2,5 untuk primer mTcCIR 213 dan 1,3 untuk primer mTcCIR 16, sedangkan klon TSH 858 mempunyai kombinasi alel masing- masing 2,4 dan 1,3 untuk primer yang sama. Berdasarkan data tersebut dapat dilihat bahwa kedua klon sama-sama heterosigot dan memiliki kompos isi alel yang berbeda. Semakin besar tingkat
1 2 1 2 3 M 1 1 2 2 3 3 4 4 5 5 6 6 7 7 8 8 9 9 10 10
heterosigos itas dan semakin beragam ko mbinasi alel pada semua lokus yang diamplifikasi untuk sepasang klon kakao yang disilangkan, maka semakin besar keragaman genetik yang aka n dihasilka n diantara tur unan hasil persilangan dari kedua tetua tersebut (Sudarsono et al.2009).
Analisis keragaman hibrida kakao dilakukan berdasarkan marka molekuler dan respon terhadap P.palmivora. Hasil analisis clustering dengan menggunakan program NTSys pada masing- masing silangan menunjukkan adanya keragaman dan pengelompokan di antara individu. Analisis yang dilakukan terhadap sepuluh individu hasil persilangan antara ICCRI3 dengan TSH 858 menunjukkan adanya pengelompokan yang terdiri atas tiga kelompok pada koefisien keragaman 0.5 (Gambar 15). Individu nomor 1, 2, 6, 9 dan 10 berada pada satu kelompok yang sama, individu nomor 1,4, dan 7 terpisah dari kelompok pertama, sedangkan nomor 1 dan 8 berada pada kelompok lain.
Gambar 15. Pengelompokkan aksesi hasil persilangan antara klon ICCRI-3 x TSH858 berdasarkan marker molekuler SSR dan berdasarkan responsnya terhadap infeksi Phytophthora palmivora.
Berdasarkan responnya terhadap penyakit busuk buah yang disebabkan P.palmivora, persilangan dua tetua ini menghasilkan delapan individu yang tahan, satu agak tahan dan satu agak rentan (Gambar 15). individu- individu yang tahan berada pada kelompok dua dan tiga, sedangkan kelompok pertama sebagian individu tahan tetapi ada yang agak tahan da n agak r entan.
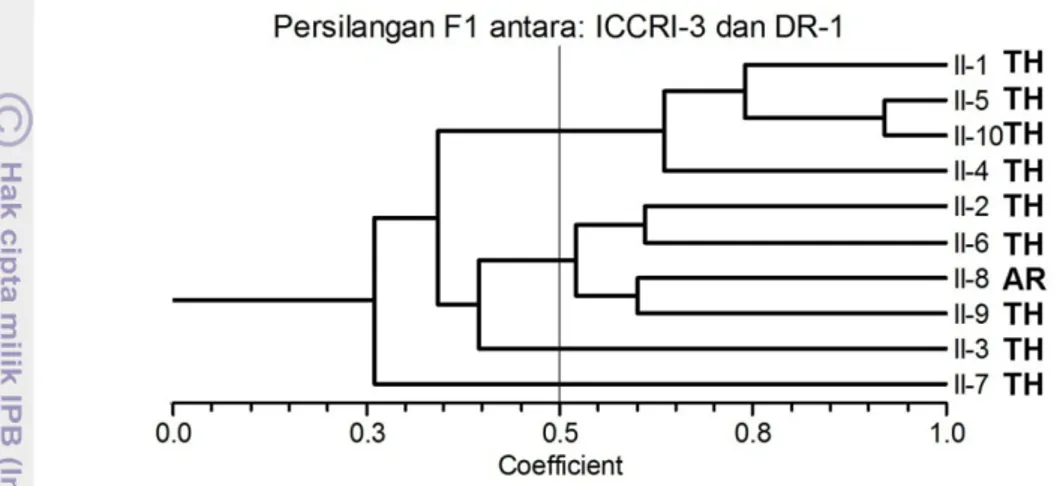
Analisis yang dilakukan terhadap sepuluh individu hasil persilangan antara ICCRI3 dengan DR1 menunjukkan adanya pengelompokan yang terdiri atas dua
kelompok pada koefisien keragaman 0.5. Individu nomor 1 berada pada satu kelompok dengan nomor 4, 5 dan 10, sedangkan nomor 2, 6, 8, dan 9 pada kelompok lain. Individu nomor 3 dan 7 terpisah dari kedua kelompok (Gambar 16).
Gambar 16. Pengelompokka n aksesi hasil persilangan antara klon ICCRI-3 x DR-1 berdasarkan marker molekuler SSR dan berdasarkan responsnya terhadap infeksi Phytophthora palmivora.
Berdasarkan responnya terhadap penyakit busuk buah yang disebabkan P.palmivora, persilangan dua tetua ini menghasilkan sembilan individu yang tahan, dan satu agak rentan (Gambar 16). Hal ini mengindikasikan bahwa secara umum silangan antara tetua ICCRI3 de ngan DR1 merupaka n tetua yang memiliki potensi kuat untuk menghasilkan keturunan yang tahan terhadap penyakit busuk buah yang disebabkan P.palmivora .
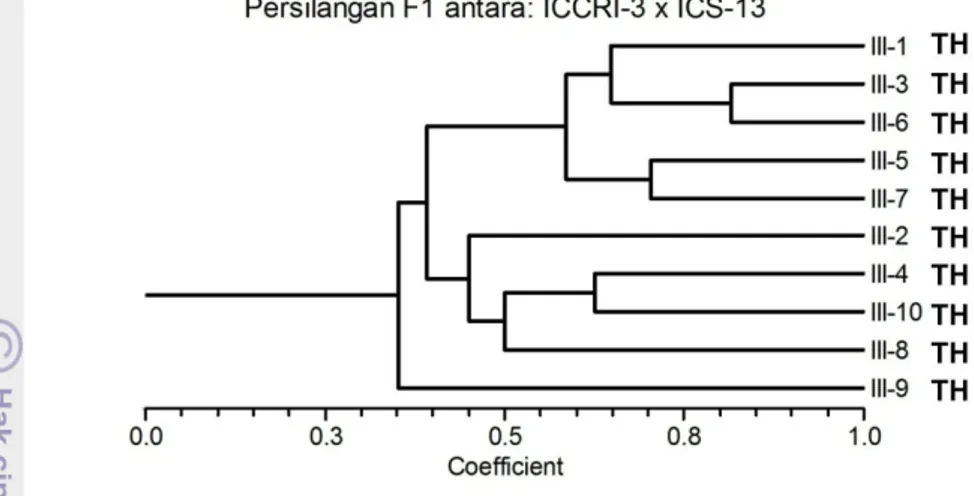
Gambar 17. Pengelompokkan aksesi hasil persilangan antara klon ICCRI-3 x ICS-13 berdasarkan marker molekuler SSR dan berdasarkan responsnya terhadap infeksi Phytophthora palmivora.
Analisis yang dilakuka n terhadap sepuluh ind ividu hasil persilangan antara ICCRI3 dengan ICS13 menunjukkan adanya pengelompokan yang terdiri atas dua kelompok pada koefisien keragaman 0.5. Individu nomor 1,3, 5, 6, dan 7 berada dalam satu kelompok, nomor 2, 4, 8 dan 10 berada pada kelompok yang lain, sedangkan individu nomor 9 tidak termasuk ke dalam dua kelompok tersebut (Gambar 17).
Berdasarkan responnya terhadap penyakit busuk buah yang disebabkan P.palmivora, persilangan dua tetua ini menghasilkan sepuluh individu yang seluruhny a tahan (Gambar 17). Hal ini mengindikasikan bahwa secara umum silangan antara tetua ICCRI3 dengan ICS13 merupaka n tetua yang memiliki potensi kuat untuk menghasilkan keturunan yang taha n terhadap pe nyakit busuk buah yang disebabka n P.palmivora .
Gambar 18. Pengelompokkan aksesi hasil persilangan antara klon ICCRI-3 x Sca-6 berdasarkan marker molekuler SSR dan berdasarkan responsnya terhadap infeksi Phytophthora palmivora.
Analisis yang dilakukan terhadap sepuluh individu hasil persilangan antara ICCRI3 dengan SCa-6 menunjukkan adanya pengelompokan menjadi dua kelompok pada koefisien keragaman 0.5. Individu nomor 1, 2, 3, 6, dan 8 berada dalam satu kelompok, nomor 4, 7, 9 dan 10 berada pada kelompok yang lain, sedangkan individu nomor 5 tidak termasuk ke dalam dua kelompok tersebut (Gambar 18).
Berdasarkan responnya terhadap penyakit busuk buah yang disebabkan P.palmivora, persilangan dua tetua di atas menghasilkan sepuluh individu yang berragam terdiri atas lima individu tahan, satu agak tahan, satu agak rentan, dua rentan, dan satu sangat rentan (Gambar 18). Keragaman ini nampaknya tidak ada kaitannya dengan pengelompokan karena pada kelompok yang sama ternyata ketahanannya bervariasi. Hal ini mengindikasikan bahwa meskipun kedua tetua memiliki sifat tahan tetapi de ngan jarak genetik yang tinggi dapat menghasilkan hibrida yang memiliki keragaman tinggi. Dengan demikian apabila mengharapkan hasil silangan yang seluruhnya tahan terhadap P. palmivora maka tidak disarankan untuk menyilangkan kedua tetua ini.
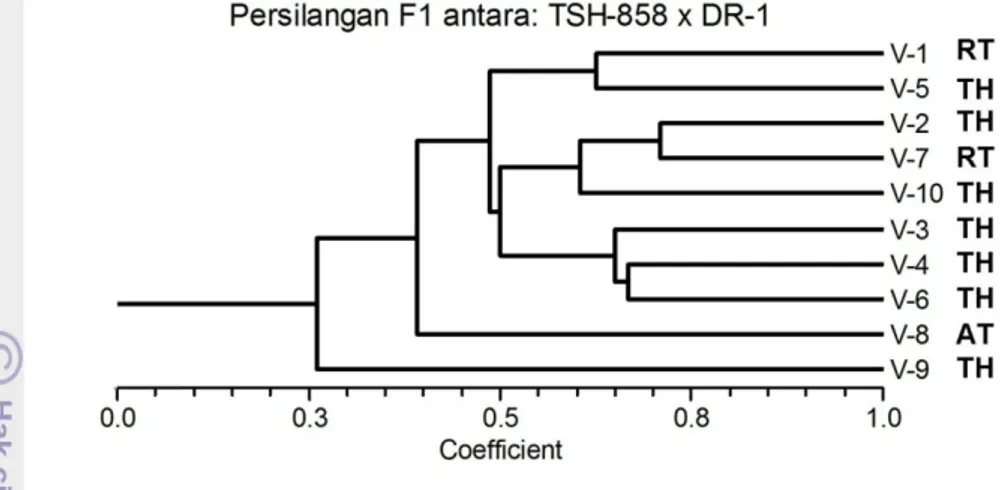
Gambar 19. Pengelompokkan aksesi hasil persilangan antara klon TSH-858 x DR1 berdasarkan marker molekuler SSR dan berdasarkan responsnya terhadap infeksi Phytophthora palmivora.
Analisis yang dilakukan terhadap sepuluh individu hasil persilangan antara TSH 858 de ngan DR1 menunjukkan adanya pengelompokan yang terdiri atas dua kelompok pada koefisien keragaman 0.5. Individu nomor 1,2, 5, 7, dan 10 berada dalam satu kelompok, nomor 3, 4, da n 6 berada pada kelompok yang lain, sedangkan individu nomor 8 da n 9 tidak termasuk ke dalam dua kelompok tersebut (Gambar 19).
Berdasarkan responnya terhadap penyakit busuk buah yang disebabkan P.palmivora, persilangan dua tetua ini menghasilkan tujuh individu yang tahan, satu agak tahan, da n dua rentan (Gambar 19). Hal ini mengindikasikan bahwa meskipun kedua tetua memiliki sifat tahan terhadap P palmivora, tetapi dengan jarak genetik yang cukup tinggi (0.53) dapat menghasilkan hibrida yang memiliki keragaman tinggi.
Gambar 20. Pengelompokkan aksesi hasil persilangan antara klon TSH-858 x ICS-13 berdasarkan marker molekuler SSR dan berdasarkan responsnya terhadap infeksi Phytophthora palmivora.
Analisis yang dilakukan terhadap sepuluh ind ividu hasil persilangan antara TSH-858 de ngan ICS-13 menunjukkan adanya pengelompokan yang terdiri atas empat kelompok pada koefisien keragaman 0.5. Individu nomor 1, 6, dan 9 berada da lam satu kelompok, nomor 2 dan 3 berada pada kelompok ke dua, individu 4 dan 8 pada kelompok ke tiga, seda ngka n individu nomor 5 dan 7 dalam kelompok yang lain (Gambar 20).
Berdasarkan responnya terhadap penyakit busuk buah yang disebabkan P.palmivora, persilangan dua tetua ini menghasilkan sepuluh individu yang cukup berragam terdiri atas lima individu taha n, dua agak tahan, da n dua rentan (Gambar 18). Hal ini mengindikasikan bahwa meskipun kedua tetua memiliki sifat tahan terhadap P palmivora, tetapi de ngan jarak genetik yang cukup jauh dapat menghasilkan hibrida yang memiliki keragaman tinggi.
Gambar 21. Pengelompokkan aksesi hasil persilangan antara klon TSH-858 x Sca-6 berdasarkan marker molekuler SSR dan berdasarkan responsnya terhadap infeksi Phytophthora palmivora.
Analisis yang dilakukan terhadap sepuluh individu hasil persilangan antara TSH-858 de ngan SCa-6 menunjukkan adanya pengelompokan yang terdiri atas tiga kelompok pada koefisien keragaman 0.5. Individu nomor 1, 6, 8 dan 10 berada dalam satu kelompok, nomor 3, 4, dan 5 berada pada kelompok ke dua, sedangkan individu nomor 2, 7 dan 9 dalam kelompok ke tiga (Gambar 21).
Berdasarkan responnya terhadap penyakit busuk buah yang disebabkan P.palmivora, persilangan dua tetua ini menghasilkan sepuluh individu yang berragam terdiri atas enam individu tahan, satu agak tahan, satu agak rentan, satu rentan, dan satu sangat rentan (Gambar 21). Keragaman ini nampaknya tidak ada kaitannya dengan pengelompokan karena. Hal ini mengindikasikan bahwa meskipun kedua tetua memiliki sifat tahan tetapi dengan jarak genetik yang tinggi (0.68) dapat menghasilkan hibrida yang memiliki keragaman tinggi. Dengan demikian apabila mengharapkan hasil silangan yang seluruhnya tahan terhadap P. palmivora maka tidak disarankan untuk menyilangkan kedua tetua ini.
Gambar 22. Pengelompokka n aksesi hasil persilangan antara klon DR-1 x ICS-13 berdasarkan marker molekuler SSR dan berdasarkan responsnya terhadap infeksi Phytophthora palmivora.
Analisis yang dilakuka n terhadap tujuh individu hasil persilanga n antara DR1 dengan ICS13 menunjukkan adanya pengelompokan yang terdiri atas tiga kelompok pada koefisien keragaman 0.5. Individu nomor 1 dan 5 berada dalam satu kelompok, nomor 2, 6, dan 7 berada pada kelompok ke dua, sedangkan individu nomor 3 dan 4 termasuk ke dalam kelompok ke tiga (Gambar 22).
Berdasarkan responnya terhadap penyakit busuk buah yang disebabkan P.palmivora, persilangan dua tetua ini menghasilkan tujuh individu yang seluruhny a tahan (Gambar 22). Hal ini mengindikasikan bahwa secara umum silangan antara tetua DR1 de ngan ICS13 merupaka n tetua yang memiliki pot ensi kuat untuk menghasilkan keturunan yang tahan terhadap penyakit busuk buah yang disebabkan P.palmivora .
Analisis yang dilakukan terhadap sepuluh ind ividu hasil persilangan antara DR1 de ngan SCa-6 menunjukkan tidak adanya pengelompokan di antara sepuluh individu hasil silangan yang dianalisis (Gambar 23). Hal ini mengindikasikan bahwa silangan ke dua tetua yang memiliki jarak genetik cukup tinggi (0.83) ini menghasilkan keragaman yang cukup tinggi.
Berdasarkan responnya terhadap penyakit busuk buah yang disebabkan P.palmivora, persilangan dua tetua ini menghasilkan delapan individu yang tahan, dan dua agak tahan (Gambar 23).
Gambar 23. Pengelompokkan aksesi hasil persilangan antara klon DR-1 x Sca-6 berdasarkan marker molekuler SSR dan berdasarkan responsnya terhadap infeksi Phytophthora palmivora.
Analisis yang dilakukan terhadap sembilan ind ividu hasil persilangan antara ICS13 dengan SCa 6 menunjukkan adanya pengelompokan yang terdiri atas dua kelompok pada koefisien keragaman 0.5. Individu nomor 1,3, 4, 6, dan 9 berada dalam satu kelompok, sedangkan nomor 3, 5, 7 dan 8 berada pada kelompok yang lain (Gambar 24).
Gambar 24. Pengelompokkan aksesi hasil persilangan antara klon ICS-13 x Sca-6 berdasarkan marker molekuler SSR dan berdasarkan responsnya terhadap infeksi Phytophthora palmivora.
Berdasarkan responnya terhadap penyakit busuk buah yang disebabkan P.palmivora, persilangan dua tetua ini menghasilkan delapan individu yang
tahan dan satu individu agak tahan (Gambar 24). Hal ini mengindikasikan bahwa secara umum silangan antara tetua ICS13 dengan SCa6 merupaka n tetua yang memiliki potensi kuat untuk menghasilka n keturuna n yang tahan terhadap penyakit busuk b uah ya ng disebabka n P. palmivora .
Simpulan
Hasil analisis dengan marka SSR terhadap populasi F1 menunjukkan adanya keragaman genetik yang cukup tinggi di antara individu- individu yang dianalisis. Hal ini terkait dengan perbedaan jarak genetik di antara tetua yang disilangkan. Demikian pula dengan karakter ketahanannya terhadap P. palmivora, hasil silangan menunjukkan keragaman tingkat ketahanannya, meskipun berasal dari dua tetua yang memiliki ketahanan yang sama. Tetua dengan jarak genetik yang tinggi menghasilkan hibrida yang memiliki keragaman tinggi baik pada tingkat alel maupun sifat resistensinya terhadap P. palmivora.
DAFTAR PUSTAKA
Borrone JW, Brown JS, Kuhn DN, Motamayor JC, and Schnell RJ. 2007. Microsatellite markers developed from Theobroma cacao L.expressed sequence tags. Molecular Ecology Notes 7: 236–239
Clement D, Risterucci AM, Grivet L,Motamayor JC, Goran JN, and Lanaud C, 2003. Mapping Q TL for yield components, vigor and resistance to Phytophthora palmivora in Theobroma cacao L. Genome 46:204-212. Drenth A. and B. Sendall. 2004. Economic Impact of Phytophthora Diseases in
Southeast Asia. In Drenth, A. and Guest, D.I., ed. 2004. Diversity and management of Phytop ht hora in Southeast Asia. ACIAR Monograph. P. 10-18
Martinez CC et al. 2006. Combining ability for disease resistance, yield, and horticultural traits of cacao (Theobroma cacao L.) clones. J Amer. Soc. Hor. Sci. 131 (2): 231-241
Pugh T et al. 2004. A new cacao linkage map based on codominant markers: development and integration of 201 new microsatelite markers. Theor Appl Genet 108:1151 – 1161
Rubiyo. 2009. Kajian Genetika Ketahanan Tanaman Kakao (Theobroma cacao L.) terhadap Penyakit Busuk Buah (Phytophthora palmivora Butl.) di Indonesia. Disertasi S3 IPB (tidak diterbitkan), 168p.
Wahyudi T, Raharjo P. 2008. Sejarah dan Prospek. Di dalam: Wahyudi T, editor. Panduan Lengkap Kakao. Jakarta: Penebar Swadaya. hlm 11-26.
Winarno H. 2008. Bahan Tanam. Di dalam: Wahyudi T, editor. Panduan Lengkap Kakao. Jakarta: Penebar Swadaya. hlm 68-73.
Wood GAR. 1985. Establishment. In GAR Wood and RA Lass (Eds.) Cocoa: 119-165. Longman, London