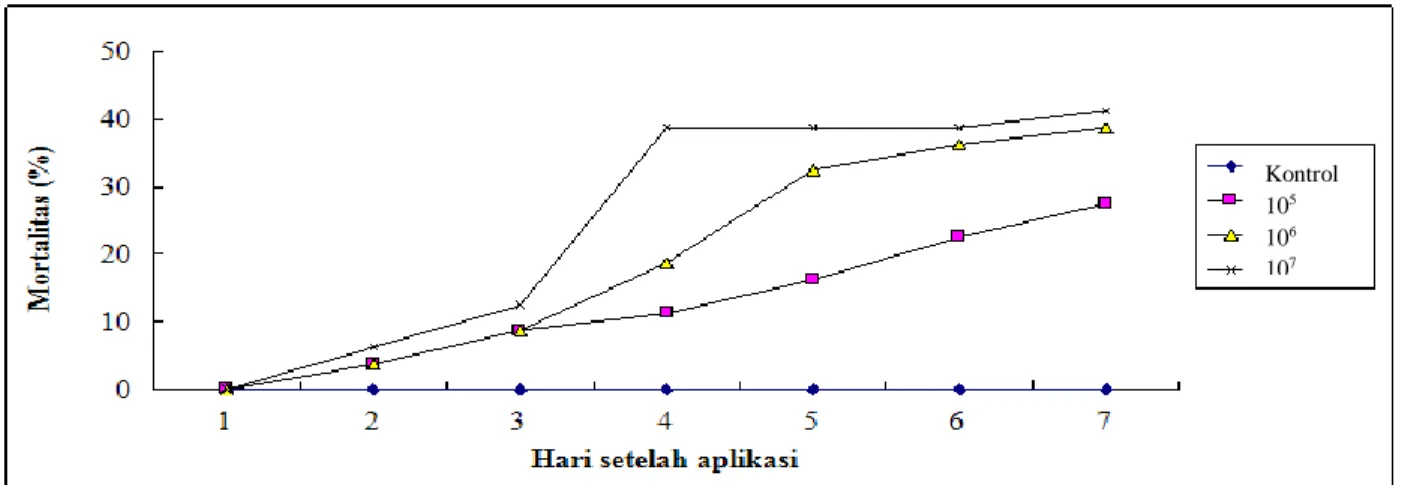
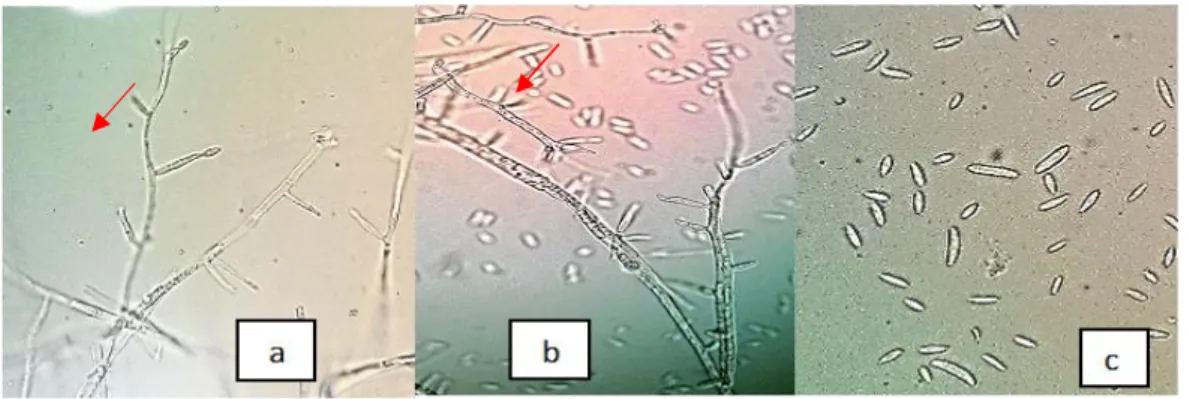
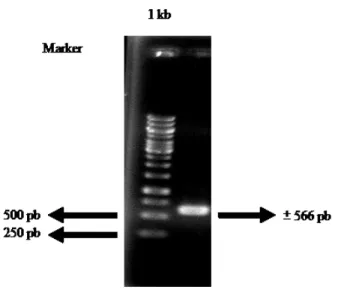
PENGENDALIAN Diaphorina citri KUWAYAMA VEKTOR PENYAKIT CVPD PADA TANAMAN JERUK DENGAN CENDAWAN ENTOMOPATOGEN ISOLAT LOKAL BENGKULU
Bebas
71
0
0
Teks penuh
Gambar

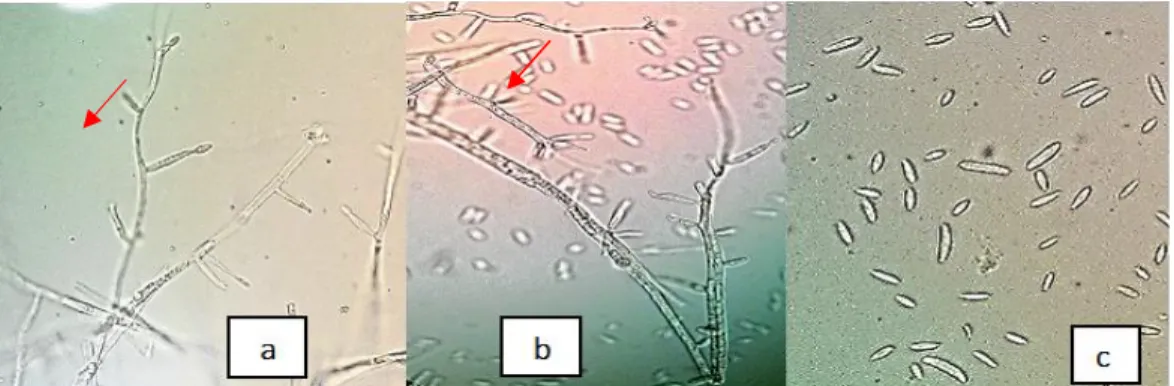
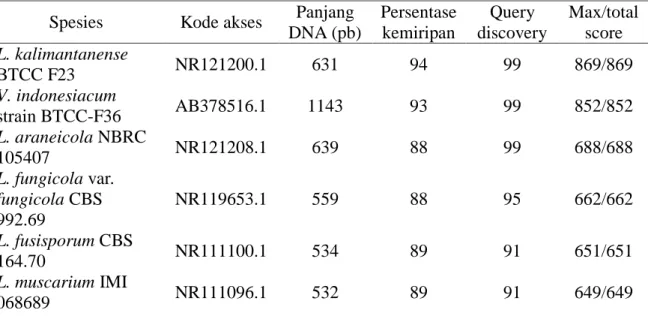
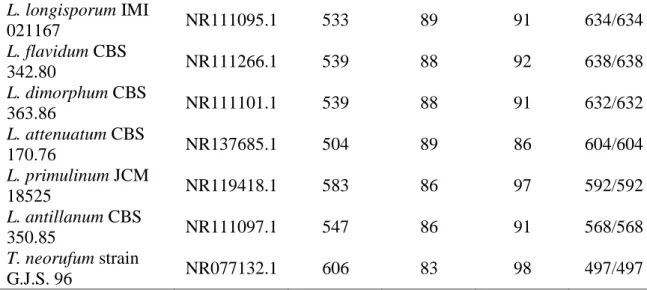
+7


Dokumen terkait
