See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/267744825
Coordination and variability in the elite female
tennis serve
ARTICLE in JOURNAL OF SPORTS SCIENCES · OCTOBER 2014 Impact Factor: 2.25 · DOI: 10.1080/02640414.2014.962569 · Source: PubMed
CITATIONS
4
READS
201
4 AUTHORS, INCLUDING:
Bruce C Elliott
University of Western Australia
297PUBLICATIONS 4,526CITATIONS
SEE PROFILE
Brendan Lay
University of Western Australia
57PUBLICATIONS 530CITATIONS
SEE PROFILE
Machar Reid
Tennis Australia
106PUBLICATIONS 681CITATIONS
SEE PROFILE
All in-text references underlined in blue are linked to publications on ResearchGate, letting you access and read them immediately.
On: 03 Novem ber 2014, At : 12: 37 Publisher: Rout ledge
I nform a Lt d Regist ered in England and Wales Regist ered Num ber: 1072954 Regist ered office: Mort im er House, 37- 41 Mort im er St reet , London W1T 3JH, UK
Journal of Sports Sciences
Publicat ion det ails, including inst ruct ions f or aut hors and subscript ion inf ormat ion: ht t p: / / www. t andf online. com/ loi/ rj sp20
Coordination and variability in the elite female tennis
serve
David Whit esidea, Bruce Clif f ord Elliot ta, Brendan Laya & Machar Reidb
a
School of Sport Science, Exercise and Healt h, Universit y of West ern Aust ralia, Crawley, Aust ralia
b
Sport s Science and Medicine Unit , Tennis Aust ralia, Melbourne, Aust ralia Published online: 30 Oct 2014.
To cite this article: David Whit eside, Bruce Clif f ord Elliot t , Brendan Lay & Machar Reid (2014): Coordinat ion and variabilit y in t he elit e f emale t ennis serve, Journal of Sport s Sciences, DOI: 10. 1080/ 02640414. 2014. 962569
To link to this article: ht t p: / / dx. doi. org/ 10. 1080/ 02640414. 2014. 962569
PLEASE SCROLL DOWN FOR ARTI CLE
Taylor & Francis m akes every effort t o ensure t he accuracy of all t he inform at ion ( t he “ Cont ent ” ) cont ained in t he publicat ions on our plat form . How ever, Taylor & Francis, our agent s, and our licensors m ake no
represent at ions or warrant ies w hat soever as t o t he accuracy, com plet eness, or suit abilit y for any purpose of t he Cont ent . Any opinions and view s expressed in t his publicat ion are t he opinions and view s of t he aut hors, and are not t he view s of or endorsed by Taylor & Francis. The accuracy of t he Cont ent should not be relied upon and should be independent ly verified w it h prim ary sources of inform at ion. Taylor and Francis shall not be liable for any losses, act ions, claim s, proceedings, dem ands, cost s, expenses, dam ages, and ot her liabilit ies w hat soever or how soever caused arising direct ly or indirect ly in connect ion w it h, in relat ion t o or arising out of t he use of t he Cont ent .
This art icle m ay be used for research, t eaching, and privat e st udy purposes. Any subst ant ial or syst em at ic reproduct ion, redist ribut ion, reselling, loan, sub- licensing, syst em at ic supply, or dist ribut ion in any
Coordination and variability in the elite female tennis serve
DAVID WHITESIDE
1, BRUCE CLIFFORD ELLIOTT
1, BRENDAN LAY
1& MACHAR REID
21
School of Sport Science, Exercise and Health, University of Western Australia, Crawley, Australia and2Sports Science and Medicine Unit, Tennis Australia, Melbourne, Australia
(Accepted 1 September 2014)
Abstract
Enhancing the understanding of coordination and variability in the tennis serve may be of interest to coaches as they work with players to improve performance. The current study examined coordinated joint rotations and variability in the lower limbs, trunk, serving arm and ball location in the elite female tennis serve. Pre-pubescent, pubescent and adult players performed maximal effortflat serves while a 22-camera 500 Hz motion analysis system captured three-dimensional body kinematics. Coordinated joint rotations in the lower limbs and trunk appeared most consistent at the time players left the ground, suggesting that they coordinate the proximal elements of the kinematic chain to ensure that they leave the ground at a consistent time, in a consistent posture. Variability in the two degrees of freedom at the elbow became significantly greater closer to impact in adults, possibly illustrating the mechanical adjustments (compensation) these players employed to manage the changing impact location from serve to serve. Despite the variable ball toss, the temporal composition of the serve was highly consistent and supports previous assertions that players use the location of the ball to regulate their movement. Future work should consider these associations in other populations, while coaches may use the currentfindings to improve female serve performance.
Keywords:biomechanics, coaching, compensation, perception-action, motor control
Introduction
A proficient serve is a crucial part of a tennis player’s stroke repertoire as it can be used to gain an advan-tage at the start of each point (McGinnis, 2013). Skilled execution of the serve involves concatenation of the lower limbs, trunk and serving arm to generate racquet head speed and transfer momentum to the ball (Bahamonde, 2000). While the notion of coor-dination has been explored variously in past tennis research, variability in specific coordinative joint rotations (e.g. concurrent extension of the hip, knee and ankle during leg drive; concurrent upper extremity joint rotations during forwardswing) is yet to be comprehensively examined and may provide coaches with useful information.
Investigations of the serve have primarily focused on discrete values such as joint ranges, discrete kine-matic peaks and phase durations. This research has confirmed that flexion at the knee and ankle pre-cedes the vigorous extension at these joints that pro-pels the player into the air (Fleisig, Nicholls, Elliott, & Escamilla,2003; Reid, Elliott, & Alderson,2008). While the player is airborne, momentum is then transferred to the serving arm through transverse,
frontal and sagittal plane trunk rotations (Bahamonde, 2000; Elliott, 2006; Martin, Kulpa, Delamarche, & Bideau, 2013). During the final stages of the serve, vigorous internal rotation at the shoulder and wrist flexion augments racquet head speed (Elliott, 2006; Marshall & Elliott, 2000; Tanabe & Ito,2007), while pronation and extension at the elbow act to orientate the racquet in a manner befitting impact (Bahamonde, 2005; Elliott, Marshall, & Noffal, 1995). Additionally, the direc-tion of the ball toss ultimately produces an impact location forward of, and lateral to, the front foot (Chow et al., 2003; Reid, Whiteside, & Elliott,
2011). From a coaching perspective, these studies provide relevant information regarding mechanics of the service action and how players generate racquet velocity and intercept the ball. There exists scope to extend this work by exploring other aspects of the service action, namely the role of coordinated joint rotations and movement variability, as they relate to performance.
Recent work has petitioned the exploration of movement variability in sports biomechanics research (Bartlett, Wheat, & Robins, 2007).
Correspondence: David Whiteside, School of Sport Science, Exercise & Health, University of Western Australia, 35 Stirling Highway, Crawley, Perth 6009, Australia. E-mail:[email protected]
Journal of Sports Sciences, 2014
http://dx.doi.org/10.1080/02640414.2014.962569
© 2014 Taylor & Francis
Documenting the variability of movement patterns within the serve may uncover how tennis players coordinate joint rotations to produce accurate, high speed serves. Despite a long history of research describing movement variability and its functional relevance to human movement (Hatze, 1986; Winter, 1984), movement variability in sporting motions was often considered noise (Bartlett et al.,
2007). While this viewpoint has been tempered by contemporary movement research, consistency in some parameters related to performance is still con-sidered critical to success.
In hitting and projectile tasks, the launch para-meters critical to success relate to the trajectory and orientation of the end-effector (i.e. the racquet, bat, club, in hitting tasks, or the hand in throwing tasks), as its terminal location, orientation and velocity will ultimately determine the outcome of the task. Indeed, a single study on the tennis serve has reported that consistency in speed and location of the serving hand around impact is positively related to serve speed and accuracy (Antúnez, Hernández, García, Vaíllo, & Arroyo,2012). Other launch para-meters that directly govern the outcome of the serve include the impact height, ball projection angle and racquet velocity, the combination of which are thought to determine serve outcome (Whiteside, Elliott, Lay, & Reid, 2013b). Though these launch parameters are known to be important, the coordi-nated joint rotations that regulate their consistency are not well understood.
Contemporary motor control research in sport describes a particular form of coordinated joint rota-tion, referred to as mechanical compensation. In human movement, this mechanism helps to regulate the performance parameters that ultimately deter-mine the outcome of the task (Dupuy, Mottet, & Ripoll, 2000; Kudo, Tsutsui, Ishikura, Ito, & Yamamoto,2000; Smeets, Frens, & Brenner,2002). More explicitly, inadvertent variations in a given execution parameter (e.g. a joint angle) are counter-acted by the actions of other execution parameters (Davids, Glazier, Araujo, & Bartlett, 2003). In this way, errors that are introduced into the movement system during performance can be managed, thus preventing any negative influence on the task out-come that they would otherwise produce. Mechanical compensation has been highlighted in numerous projectile and striking sports including golf drives (Horan, Evans, & Kavanagh, 2011), free throw shooting in basketball (Button, MacLeod, Sanders, & Coleman, 2003; Mullineaux & Uhl,
2010), underarm throwing (Dupuy et al., 2000; Kudo et al., 2000), overarm throwing (Wagner, Pfusterschmied, Klous, Von Duvillard, & Müller,
2012), table tennis forehands (Bootsma & Van Wieringen, 1990), tennis forehands (Knudson,
1990), pistol shooting (Scholz, Schoner, & Latash,
2000) and dart throwing (Smeets et al.,2002). Since mechanical compensation is an inherent biomechani-cal response to the demands presented when enacting a particular movement task, it does not follow a con-sistent pattern. Therefore, variations in joint rotations (or other mechanics) are no longer considered to be indicative of movement system ineptitude. Rather, movement variability is now considered critical to the stabilisation of the performance parameters that directly govern the outcome of the task (e.g. launch parameters in the serve). In other words, variability in the movement system is not detrimental so long as the critical end point parameters remain stable. This notion is reflected in recent tennis research, where Whiteside et al. (2013b) showed that increased varia-bility in coordinated elbow and wrist joint rotations did not reduce serve accuracy. Given the whole-body nature of the serve, other mechanics may also con-tribute to the compensatory process but are yet to be characterised.
Movement variability in sporting actions is not restricted to the magnitudes of discrete joint rota-tions. Research in other striking skills has highlighted the importance of consistent temporal patterns (i.e. the times at which specific movements occur) in interceptive (batting) skills in cricket (Renshaw, Oldham, Davids, & Golds, 2007) and baseball (Katsumata, 2007), as well as the volleyball serve (Davids, Kingsbury, Bennett, & Handford, 2001). It has been suggested that these athletes utilise infor-mation from the incoming ball (i.e. its location) to regulate the initiation of key propulsive movements, hence ensuring that they intercept it at the appropri-ate moment. Similar strappropri-ategies have been found in the tennis serve, where players regulate their move-ments such that their arrival in the trophy position coincides with ball zenith (Reid, Whiteside, & Elliott, 2010; Whiteside, Elliott, Lay, & Reid,
2013a). Likewise, the timing of peak forward racquet velocity is considered critical to accuracy in thefirst serve (Antúnez et al.,2012). Ultimately, coordinated joint rotations and/or temporal consistency may be thought of as techniques that players exploit to sim-plify complex sporting movements. With this in mind, it is expected that tennis players simplify those aspects of the serve that are most important to performance. At present, the particular methods employed to do so are somewhat unclear as this topic is yet to be comprehensively examined in the tennis serve.
It has been suggested that, in learning movement skills, inexperienced performers initially reduce the skill complexity by restraining degrees of freedom (Anderson & Sidaway, 1994; Stergiou & Decker,
2011). These degrees of freedom are then gradually released as players become more familiar with the
task and explore alternative solutions to the move-ment problem (Gentile, 2000; Newell, Deutsch, Sosnoff, & Mayer-Kress, 2006). In this sense, chil-dren are often considered novices owing to their motor inexperience (Guarrera-Bowlby & Gentile,
2004). Indeed, the popular “ten year (10,000 h) rule” (Ericsson, Krampe, & Tesch-Römer, 1993) virtually precludes any attempt to categorise children as expert performers. This position is offered partial support by Newell’s constraints model (Newell,
1986), which states that organismic factors (e.g. anthropometry, strength, anticipatory ability) infl u-ence performance. As such, more physically and mentally mature adults are expected to exhibit more skilled performance than children. However, “elite child athletes” present a unique cohort whose motor proficiency may exceed their age-related expectations and whose practice regimes are often meticulous and intensive. Consequently, these ath-letes may not conform to traditional descriptions of child motor performance and deserve investigation in their own right. With movement patterns expected to change throughout development, it seems neces-sary to separate performers according to level of development when investigating movement in devel-oping populations.
At present, there exists scope to supplement exist-ing tennis literature with research focused on coor-dinated joint rotations and variability in the serve. Therefore, the aims of this study were to examine coordinated joint rotations and variability in the lower limbs, trunk, serving arm and ball in the tennis serves of elite female players at three stages of devel-opment. It was expected that functional movement variability in the distal joints would manifest during serve performance to preserve a successful outcome, though its disposition would change with age. However, the temporal composition of the serve was expected to be comparatively more consistent as players stabilised the timing of critical postures with respect to impact.
Methods
Participants
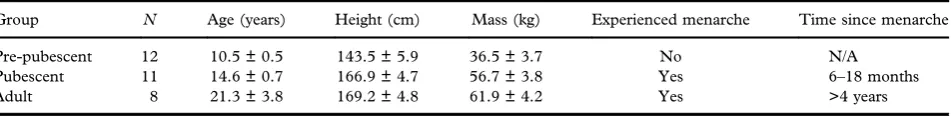
The relevant Human Ethics Committee approved this study prior to recruitment. Thirty-one elite female tennis players provided informed consent
and were arranged into pre-pubescent, pubescent and adult groups based on their age and menarchial status (Table I). At the time of testing, players in the pre-pubescent and pubescent groups held a top 8 national ranking for their respective age groups, while the adult players possessed a professional (Women’s Tennis Association: WTA) ranking higher than 325. This cohort was the same as that used in a previous study (Whiteside et al.,2013a).
Protocol
The protocol was completed at an indoor biomecha-nics laboratory, where a full-size tennis court was constructed. Sixty retro-reflective markers, 14 mm in diameter, were affixed to each player according an established, calibrated anatomical systems (CAST)-based, full body marker set (Besier, Sturnieks, Alderson, & Lloyd, 2003; Chin, Elliott, Alderson, Lloyd, & Foster, 2009; Lloyd, Alderson, & Elliott,
2000). Three hemispherical markers, composed of ultra-light foam (radius 7 mm), were placed on the racquet and three more on ball (Whiteside, Chin, & Middleton, 2013) to create coordinate systems therein. Prior to the serving protocol, dynamic cali-bration of a≈5.5 (deep) 4 (wide) 4 (high) m capture volume yielded a mean residual calibration error smaller than 0.002 m.
Each player completed a standard 10 min warm up and used their own racquet to complete the protocol. Verbal confirmation of preparedness prompted the initiation of the test protocol in which players performed maximal effort flat (i.e. “power”) serves. Players were instructed to aim for a familiar 1 1 m target (Elliott et al., 1995; Martin et al., 2013; Reid et al., 2008; Sakurai, Reid, & Elliott, 2012) bordering the T of the ser-vice box (right-handers: deuce court; left-handers: advantage court) and to hit all serves with the same effect (i.e. to emphasise speed as opposed to spin). Five blocks of eight serves were performed with a 2 min rest period separating successive blocks. Three-dimensional (3D) marker trajectories were recorded using a 22-camera VICON MX system (VICON Motion Systems, Oxford, UK) operating at 500 Hz. The global reference frame originated at the centre of the baseline, where positive x pointed rightward along the baseline, positive y pointed to the net and positive z pointed up. Five of each
Table I. Mean (± standard deviation) age, physical and menarchial characteristics of participants.
Group N Age (years) Height (cm) Mass (kg) Experienced menarche Time since menarche
Pre-pubescent 12 10.5 ± 0.5 143.5 ± 5.9 36.5 ± 3.7 No N/A
Pubescent 11 14.6 ± 0.7 166.9 ± 4.7 56.7 ± 3.8 Yes 6–18 months
Adult 8 21.3 ± 3.8 169.2 ± 4.8 61.9 ± 4.2 Yes >4 years
Coordination and variability in the tennis serve 3
player’s fastest serves landing in the target area were selected for analysis.
Data processing
Gaps in the raw marker trajectories were interpolated using a cubic spline within the VICON Nexus soft-ware. A second-order polynomial extrapolation spe-cific to tennis limited the distortion of kinematic data around impact (Knudson & Bahamonde, 2001; Reid, Campbell, & Elliott, 2012). Data were subse-quently filtered using a Woltring filter (Woltring,
1986) with the optimal mean squared error of 2 mm determined by a residual analysis, and then modelled using the University of Western Australia’s full body (Besier et al., 2003; Lloyd et al., 2000), racquet and ball (Whiteside et al., 2013) models. Joint rotations were expressed using the Euler ZXY sequence. Trunk rotations were expressed relative to a virtual anatomical reference frame (xpointing for-ward tofor-wards the net, y pointing up andz pointing right) originating at the global origin. To maintain consistency in the statistical analyses, kinematics for the left-handed players were inverted where appro-priate such that all players could be considered together as right-hand dominant (Campbell, Straker, O’Sullivan, Elliott, & Reid, 2013; Whiteside et al., 2013b).
Relevant events of the service action
Figure 1denotes how the service action was deemed to begin at the instant the ball was released from the
hand. Ball zenith represented the peak vertical dis-placement of the ball during its toss. The subsequent nadir of vertical racquet displacement was the rac-quet low point, which is usually coincident with a player leaving the ground (Bonnefoy, Slawinski, Leveque, Riquet, & Miller, 2009). Impact was defined as the frame (i.e. 0.002 s) prior to racquet-ball contact. The duration of the serve was consid-ered as the time period between ball release and impact. Leg drive was defined as the period from ball zenith to racquet low point, while racquet low point to impact was considered the forwardswing phase of the serve.
Variables of interest
To gauge inter-limb coordination during leg drive, the relative bilateral flexion-extension motion of the ankles was compared, as was the relative bilateral flexion-extension motion of the knees. The relative transverse (twist) and frontal plane (shoulder-over-shoulder) trunk rotations prior to impact indicated how players manipulated the trunk during this time, while relative extension and pronation denoted the same for the elbow in the serving arm. The location of the ball during the toss was expressed relative to the front toe (Chow et al., 2003; Reid et al.,2011), and its spatial variability measured at both ball zenith and impact. Finally, the timing of both ball zenith and racquet low point (expressed as a percentage of the serve), and also duration of forwardswing pro-vided an insight into the temporal pattern of the service action.
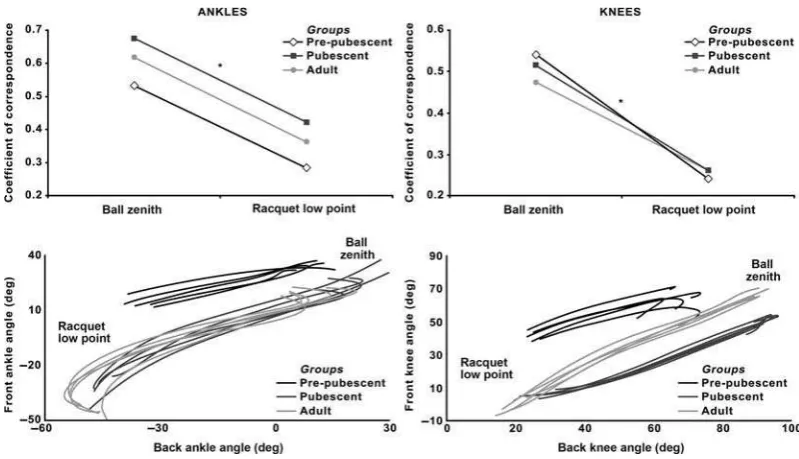
Figure 1. Key time points of interest in the tennis serve.
Assessment of variability in joint mechanics and the ball toss
Angle-angle plots provided a qualitative insight into relative joint rotations. The variability of these traces was quantified using the coefficient of correspon-dence – a vector coding technique that assesses the repeatability of several variable–variable traces (Tepavac & Field-Fote, 2001). Unlike alternative vector coding methods that consider only vector direction, the coefficient of correspondence is advan-tageous in that it also considers vector magnitude (Wheat & Glazier, 2006). Ultimately, the coefficient of correspondence quantified the magnitude of variability in coordinated joint rotations using a value between 0 and 1 (0 = no variability; 1 = max-imum variability). Akin to previous work (Horan et al., 2011), the average coefficient of correspon-dence value ±5% of the relevant event represented the variability at that time point.
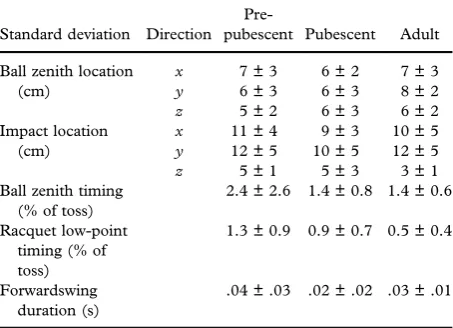
The 3D (xσ,yσ andzσ) standard deviations of ball displacement at ball zenith and impact were calcu-lated for each player across their five trials. The mean and standard deviations of these standard deviations provided a gauge of ball toss variability for each group (Davids et al., 2001; Reid et al.,
2010). Further, each standard deviation was doubled (effectively creating error bar representing one stan-dard deviation either side of the mean, in each dimension) and then multiplied (2xσ 2yσ 2zσ) to yield the“variability volume”: a singular quantity of the ball’s 3D spatial variability for each player, across their five serves.
Statistical analyses
Four split plot analyses of variance (SPANOVAs) were used to examine between group (i.e. differences between each group), within group (i.e. differences between each time point) and interaction (i.e. group time) effects in the coefficient of corre-spondence. The same procedure was used to com-pare the variability volumes in each group at ball zenith and impact. The ranges of motion at the ankles, knees, trunk and elbow were compared between groups using one-way analyses of variance. Where significant main effects existed, Bonferroni-corrected post-hoc tests were employed to find the source of the difference. In total, 13 analyses of variance were performed, thus inflating the risk of encountering type I errors. To account for this, the significance level was adjusted sequentially for each test according to the correction method proposed by Holm (1979). The p-value was not sufficient to reject the null hypothesis in the eleventh test, indi-cating that all significance in this study was reported using ap-value of 0.0125. The temporal variability of ball zenith, racquet low point and forwardswing duration were interpreted descriptively.
Results
Ankle mechanics
The representative traces inFigure 2represent how all groups utilised simultaneous plantar flexion at both ankles and simultaneous extension in both
Figure 2. Mean coefficients of correspondence and representative angle–angle plots for the ankles and knees. Note: *Significant difference between ball zenith and racquet low point in all groups.
Coordination and variability in the tennis serve 5
knees during leg drive. The coefficient of correspon-dence for ankle motion displayed a significant main effect for group (F2,28 = 16.499; P < .001), which post-hoctests revealed to be a product of significantly higher overall variability in the pubescent group compared with the pre-pubescent group (mean dif-ference (MD): 0.14; 95% confidence interval (CI): 0.08–0.20;P< .001). From a temporal perspective, the bilateral coupling of ankle plantarflexion was significantly more consistent at racquet low point compared with ball zenith and independent of group (F1,28 = 151.111; MD: 0.25; CI: 0.21–0.29; P < .001).
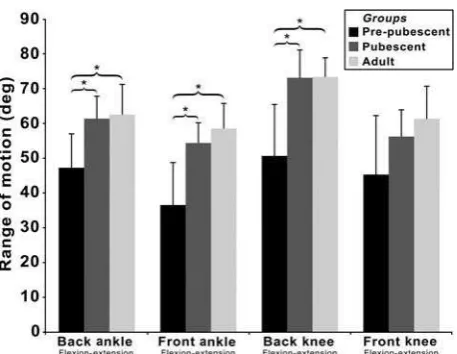
The range of plantarflexion at the back ankle was significantly (F2,28= 10.913;P< .001) smaller dur-ing leg drive in the pre-pubescent group compared with the pubescent (MD: 14.12°; CI: 5.10–23.14°; P = .001) and adult (MD: 15.24°; CI: 5.38–25.11°; P = .002) groups. Likewise, the range of plantar flexion at the front ankle (F2,28 = 17.299;P< .001) was significantly smaller during leg drive in the pre-pubescent group compared with the pre-pubescent (MD: 17.83°; CI: 8.12–27.53°;P< .001) and adult (MD: 21.91°; CI: 11.29–32.52°; P < .001) groups (Figure 3).
Knee mechanics
Regarding the coefficient of correspondence for bilateral knee extension, the main effect for group was not significant (F2,28= .162;P= .852), although a significant main effect for time revealed that rela-tive knee extension was significantly more consistent at racquet low point compared with ball zenith (F1,28 = 86.376; MD: 0.26; CI: 0.20–0.31; P < .001).
During leg drive, the back knee extended through a significantly (F2,28 = 15.954; P < .001) greater range in the pubescent (MD: 22.46°; CI: 10.97– 33.95°; P < .001) and adult (MD: 22.59°; CI: 10.02–35.15°; P < .001) groups compared with the pre-pubescent group, though range of motion (RoM) at the front knee was not significantly differ-ent between groups (F2,28= 4.364; P= .022).
Trunk mechanics
The variability of concurrent frontal and transverse plane trunk rotations revealed a significant main effect for group (F2,28 = 10.530;P< .001), wherein relative trunk rotations were significantly more con-sistent in the pubescent (MD: 0.08; CI: 0.03–0.13; P = .001) and adult (MD: 0.08; CI: 0.02–0.13; P = .005) groups compared with the pre-pubescent group. A post-hoc decomposition of the significant main effect for time point (F2,28 = 67.672; P < .001) revealed that relative trunk motion was significantly different at all time points (most vari-able at ball zenith, most consistent at racquet low point, in between at impact).
The range of trunk twist rotation did not differ with age (F2,28= .383;P= .686); however, the range of shoulder-over-shoulder rotation was significantly (F2,28 = 13.744; P < .001) larger in the pubescent (MD: 20.12°; CI: 9.45–30.74°;P< .001) and adult (MD: 18.04°; CI: 6.43–29.65°; P = .001) groups compared with the pre-pubescent group.
Elbow mechanics
A main effect for group (F2,28= 7.606;P= .002) was discovered for the coefficient of correspondence of the relative elbow joint rotations (flexion and radio-ulnar pronation). Further analyses revealed that the pubescent group was significantly more consistent than the adult group (MD: 0.12; CI: 0.04–0.21; P= .002) over the period in question. Additionally, post-hoc analyses of the significant time effect (F2,28 = 70.069; P < .001) showed that variability was significantly different at each of the three time points (becoming progressively more variable between ball zenith and impact). Importantly, an interaction effect was also noted (F2,28 = 24.089; P < .001) and was found to relate to the fact that the coupled elbow joint rotations only became sig-nificantly more variable between racquet low point and impact in the adult group (MD: 0.47; CI: 0.31– 0.64;P< .001) (Figure 4).
The range of flexion-extension at the elbow between ball zenith and impact did not differ between groups (F2,28= .953; P = .398). However, during the same period, the range of radio-ulnar pronation was significantly (F2,28 = 12.116;
Figure 3. Mean ranges offlexion-extension motion at the ankles and knees during leg drive.
Note: *Significant difference between groups.
P < .001) greater in the adult group compared with the pubescent (MD: 17.33°; CI: 8.12–26.55°; P < .001) and pre-pubescent (MD: 17.78°; CI: 4.14–23.18°;P= .002) groups (Figure 5).
Spatial variability of the ball toss
The variability volumes were not significantly affected by group (F2,28 = .371; P = .694). However, a main effect for time (F1,28 = 23.065; P < .001) revealed that the spatial location of the ball was significantly more variable at impact
compared with ball zenith (MD: 1672 cm3; CI: 959–2386 cm3) (Table II). Descriptively, ball loca-tion was most consistent in the vertical direcloca-tion at impact compared with the left-right and forward-backward directions (Table III).
Temporal variability of the ball zenith and racquet low-point events
Qualitatively, although the timing of racquet low point was comparatively more consistent than ball zenith in all groups, both events displayed impressive temporal consistency. The duration of forwardswing was equally consistent, displaying an average stan-dard deviation <.04 s in all three groups.
Discussion
Coaching texts often stress the importance of coordi-nation and rhythm in the serve (Bollettieri, 2001; Elliott & Saviano,2001; Yandell, 1999). Developing the understanding of coordinated joint rotations and movement variability may help coaches to conceptua-lise specific aspects of coordination and translate them to practice. The present study extends the understand-ing of the service action in these respects, across three developmentally different groups of elite female players. For example, prepubescent players were observed to extend their ankles and knees through a smaller range than pubescent and adult players during leg drive. More universally, the variability of lower limb and trunk postures, in all players, decreased between ball zenith and the moment that they left the ground. Thereafter, significant increases in the variability of
Figure 4. Mean coefficients of correspondence and representative angle–angle plots for the trunk and elbow.
Note: *Significant difference between the time points in all groups.†Significant difference between racquet low point and impact in the adult group.
Figure 5. Mean ranges of motion at the trunk and elbow between ball zenith and impact.
Note: *Significant difference between groups.
Coordination and variability in the tennis serve 7
trunk and/or elbow joint rotations appeared to be a functional response to the varying impact location, thereby allowing these players to achieve a suitable racquet-ball impact as the ball location changed from serve to serve. In other words, the timing at and pos-ture in which these players left the ground was rela-tively consistent; however, while airborne, their trunk and elbow mechanics generally became more variable. Practically, thesefindings encourage coaches to incor-porate deliberate (though not extreme) perturbations of the service action to cultivate appropriate coordina-tive joint rotations and perception-action coupling in the tennis serve.
Lower limb mechanics
During leg drive, the angle–angle traces illustrate simultaneous plantar-flexion at both ankles and simultaneous extension at both knees. This is not surprising and demonstrates how elite players are able to coordinate bilateral extension at the lower limb joints to propel their bodies off the ground (Bahamonde, 2000; Girard, Micallef, & Millet,
2005; Reid et al.,2008). It should be noted, however,
that the range through which these joints extended was significantly reduced in the pre-pubescent group (AnkleBack≈ 47°; AnkleFront≈37°; KneeBack ≈51°) compared with the pubescent (AnkleBack ≈ 61°; AnkleFront ≈ 54°; KneeBack ≈ 73°) and adult (AnkleBack≈ 63°; AnkleFront≈58°; KneeBack ≈73°) groups. This may offer a supplementary explanation for why a reduced leg drive magnitude has been reported in prepubescent females (Whiteside et al.,
2013a). Given that lower limb motions contribute
significantly to the force required in the tennis serve (Kibler,1995), junior coaches should be mindful of this potential limitation in the immature service action. More explicitly, it has been hypothesised that leg drive is a precursor to trunk rotations (Bahamonde, 2000), pre-stretching of the shoulder (Bahamonde,1997) and racquet speed (Reid et al.,
2008), all of which would then be limited in the pre-pubescent serve.
In all three groups, variability in bilateral exten-sion at the ankles and knees decreased significantly between the start (ball zenith) and end (racquet low point) of leg drive. With racquet low point representing the time at which players generally leave the ground (Bonnefoy et al., 2009), it appears that players of all ages act to sequentially reduce variability in lower limb motion up to this time. In doing so, players may find it easier to produce a more consistent vertical propulsion. At both ball zenith and racquet low point, the pub-escent group exhibited significantly more variable relative ankle motions than the pre-pubescent and adult groups and may represent an intermediate stage of motor development. To this point, their disparate ranges of motion at the ankle denote how the pre-pubescent (less RoM) and adult (more RoM) groups used different motor patterns at the ankles, each of which was highly consistent in its own right. In the pre-pubescent group, reduced ankle involvement (i.e. “freezing”) may be indica-tive of a deliberate attempt to reduce the complex-ity of the movement, allowing these players to develop a more repeatable action. In contrast, additional years of practice and greater lower limb strength may have afforded the adult players more masterful coordination of the ankles through
Table II. Two-way mixed ANOVA results for ball variability volumes at ball zenith and impact.
Within group Between group Interaction
Group Ball zenith Impact F P F P F P
Pre-pubescent 1596 ± 1273 4373 ± 2179
Pubescent 1520 ± 860 3264 ± 2476
Adult 2509 ± 1997 3006 ± 2336
Overall 1805 ± 1396 3627 ± 2330 23.065 <.001* .371 .694 3.431 .046
Note: *Significant difference between time points.
Table III. Mean (± standard deviations) within-individual stan-dard deviations (n= 5 serves) for the spatial and temporal para-meters of the ball toss.
Standard deviation Direction
a more expansive range, providing a different explanation for their consistency. The pubescent group may represent an intermediate and explora-tory phase of motor development where the degrees of freedom (i.e. ankle plantar-flexion) have been fully released but lack mastery, resulting in more variable performance. These movement discrepancies may also help to explain why the magnitude of prepubescent and pubescent leg drive has been reported as being 21–22% and 15–16% smaller than professional female players, respectively (Whiteside et al., 2013a).
Trunk mechanics
Simultaneous frontal and transverse plane trunk rotations were evident from the angle–angle traces in all groups, though their respective magnitudes differed. Independent of group, twist and shoulder-over-shoulder rotations initially occurred together, before the latter motion was restrained in the two younger groups as they approached impact. With leg drive generating momentum that may be transferred to the trunk (Bahamonde,
2000), the smaller range of frontal plane trunk rotation in the pre-pubescent group is not unex-pected in light of the lower limb results. These findings support previous work where the magni-tude of shoulder-over-shoulder rotation has been shown to be significantly (5–9%) smaller in junior players compared with their adult counterparts (Whiteside et al., 2013a).
Despite being significantly more variable in the pre-pubescent group, the trunk orientation at rac-quet low point was the most repeatable mechan-ical feature of each group’s service action. That this trunk posture was relatively variable at the commencement of leg drive (ball zenith), but then extremely repeatable at the end of leg drive (racquet low point), points to a functional impor-tance of trunk postures at the time these players left the ground. Similar phenomena have been described for elite golfers, where the attainment of consistent postures at certain critical time points of the golf swing are considered features of skilled performance (Bradshaw et al., 2009). Therefore, the significant reduction in variability towards a highly repeatable trunk posture at rac-quet low point may be indicative of an equivalent posture in the tennis serve. Subsequent to racquet low point, relative trunk motion became signifi -cantly more variable in all players and likely represents postural adjustments required to man-oeuvre the body correctly for impact.
Serving arm mechanics
During forwardswing, the range of elbow extension did not differ with age; however, the adult group rotated through a significantly greater range of pro-nation (≈45°) compared with the pre-pubescent (≈28°) and pubescent (≈31°) groups. What might be interpreted as a liberated degree of freedom (pro-nation) may have been responsible for the adult’s unique and significant increase in elbow joint rota-tion variability prior to impact. Where previous work in adults has suggested that pronation prior to impact acts to rotate the racquet face to contact the ball (Elliott,2006; Tanabe & Ito,2007), their lack of pronation indicates that this motion may be restrained in junior players. Instead, junior players appear to commence forwardswing in a more pro-nated posture, thereby reducing the need to manage this degree of freedom thereafter. Restraining prona-tion effectively reduces the elbow joint to a single degree of freedom and offers an explanation for the lower coefficient of correspondence at the elbow in junior players. This mechanism may be an attempt to reduce the complexity of the movement and, in turn, the margin for error at impact. A potential drawback of this approach is that the face of the racquet is more “open” during the forwardswing, likely providing greater aerodynamic resistance com-pared with adult players who cut through the air with the edge of their“closed”racquets. Further, restrict-ing this degree of freedom logically limits the com-pensatory potential of the system and, in turn, its adaptability. It follows that this would leave their movement systems more susceptible to perturba-tions. However, it is also possible that young players rely on other degrees of freedom (e.g. at the shoulder and/or wrist) for compensatory purposes. Combined with relatively lower muscle strength, this fact may be relevant to upper extremity pathologies and pro-vides an avenue for future work.
The diverging angle–angle curves at impact clearly show that variable and abrupt, low-magnitude pro-nation or supipro-nation movements were common immediately prior to impacting the ball. Interestingly, previous work has noted similar varia-tions in pronation-supination mechanics immedi-ately prior to impact (Elliott et al., 1995; Sprigings, Marshall, Elliott, & Jennings, 1994; Tanabe & Ito,
2007; Van Gheluwe, De Ruysscher, & Craenhals,
1987). Tanabe and Ito (2007) proposed that, in faster servers, the emergence of pronation-supina-tion was dependent on shoulder mechanics and helps to orient the racquet for impact. In other words, joint motions were not independent of one another and emerged synergistically, according to the unique needs of each serve. The variable Coordination and variability in the tennis serve 9
magnitudes and directions of the elbow joint rota-tions around impact in this study are consistent with compensatory function that may help to ensure a suitable racquet-ball impact. These compensatory motions seem to intensify after the onset of puberty, coinciding with an increased involvement of prona-tion in the serve. The locaprona-tion of the ball at impact may help to further explain the mechanical compen-sation in question.
Spatial variability of the ball toss
After the ball leaves the hand, the server has no con-trol over its trajectory and must move appropriately to intercept it. For this reason, body and racquet move-ments are heavily dependent on the ball toss. It could be argued that a repeatable ball toss would mitigate its influence on biomechanics; however, the values in
Table III suggest that perhaps the ball toss is less consistent than many coaches appreciate. From the variability volume, it can be conservatively assumed that the ball was placed anywhere within a ≈1500– 2500 cm3area of space at ball zenith and, by impact, this volume had increased to≈3000–4400 cm3. This endorses the growing body of scientific work that has demonstrated how elite players do not use a highly consistent ball toss (Reid et al., 2010, 2011; Whiteside, Giblin, & Reid,2014). More importantly, a changing impact location from serve to serve man-dates movement variability in the service action. Given that the ball location (a) varies between serves and (b) ultimately shapes the mechanics of the service action, it appears more important for players to learn how to perceive and respond to the varying ball loca-tion rather than develop a perfectly consistent acloca-tion (which may be impossible anyhow; Whiteside et al.,2013b,2014). While the absolute ball location was more variable at impact–akin to previous work in tennis (Reid et al.,2010,2011)–its vertical location was most consistent at this time. This seemingly con-firms that impact height is a critical launch parameter that is highly consistent in successful serves (Elliott, Marsh, & Blanksby,1986; Reid, Elliott, & Alderson,
2007; Whiteside et al., 2013b) and which coaches should aim to refine.
Temporal variability of the ball zenith and racquet low-point events
The occurrence of ball zenith, though significantly ear-lier in the pre-pubescent group, was highly consistent within the groups as evidenced by the respective stan-dard deviations (pre-pubescent group: 2.4% ± 2.6% of serve; pubescent: 1.4% ± 0.8%; adult: 1.4% ± 0.6%). The standard deviations of racquet low-point timing were more consistent still (pre-pubescent: 1.3% ± 0.9%; pubescent: 0.9% ± 0.7%; adult:
0.5% ± 0.4%). Therefore, not only are lower limb and trunk postures repeatable at racquet low point, but the event itself occurs at a highly repeatable time point of the service action. This temporal stability is consistent with other striking actions, where the tem-poral initiation of propulsive movements has been shown to be highly repeatable in baseball (Katsumata, 2007) and cricket batting (Renshaw et al., 2007) as well as volleyball serving (Davids et al., 2001). Indeed, it has already been suggested that tennis players use the location of the ball to reg-ulate their preparatory movements in the serve such that their arrival in the trophy position coincides with ball zenith (Reid et al.,2010; Whiteside et al.,2013a). The players in this study may have used the same method to ensure that racquet low point occurs at a relatively consistent juncture of the serve. Considering that racquet low point represents the beginning of for-wardswing, regulating the occurrence of this event also stabilises the duration of forwardswing. This strategy effectively prevents the movement system from collap-sing in the presence of a variable ball toss since action and perception are coupled. For example, if the ball toss is slightly higher than intended, the player com-mensurately protracts the initial phase of their service action, thus delaying the initiation of forwardswing. Having ensured that they will arrive at impact at an appropriate time, the player’sfinal dilemma is achiev-ing an appropriate racquet orientation at impact. The solution appears to lie in the subtle mechanical adjust-ments (i.e. mechanical compensation) in the distal joints that help to manoeuvre the racquet to intercept the ball.
Future work may wish to explore smaller targets to determine whether the size of the target contributed to any of the variability recorded in this study. Studying their disposition in unsuccessful serves could also reveal how the mechanics examined in this study relate to accuracy. Similarly, exploring other joint mechanics and/or couplings would help to develop a more comprehensive understanding of coordination in the serve. Instructional methods aimed at developing coordinated joint rotations and/or perception-action coupling in the serve could also be appraised to improve coaching techni-ques. Further, perception-action couplings in the serve could be evaluated more directly by quantify-ing gaze behaviour durquantify-ing the ball toss.
Conclusion
The elite female tennis players in this study appeared to reduce movement variability in the lower limbs and trunk between the trophy position (i.e. ball zenith) and the time that they leave the ground (i.e. racquet low point). They also left the ground at a highly consistent time point of the service action,
which acted to stabilise the duration of forward-swing. Given that this temporal consistency persisted in spite of a variable ball toss location, these players seemingly used information from the ball to deter-mine the appropriate times to initiate appropriate body movements. After becoming airborne, subtle mechanical adjustments in the trunk and serving arm acted to manoeuvre the racquet to intercept the ball. Variability in these adjustments across serves was mandatory since the location of the ball at impact changed from serve to serve. There was a reduced potential for this mechanical compensation in junior players as they restrained elbow pronation in the serving arm during forwardswing to reduce their margin for error at impact. In light of these points, tennis players may benefit from instructional techniques that emphasise perception-action cou-pling as it relates to the serve. More specifically, deliberate perturbations of the service action (e.g. manipulating the ball toss, constraining joint motions, utilising dynamic targets) expose a player to a wider variety of performance contexts than they would otherwise experience. This would logically help players to become more attuned to the move-ment errors that present during performance and enhance their ability to manage these errors by developing a more adaptable movement system.
References
Anderson, D., & Sidaway, B. (1994). Coordination changes asso-ciated with practice of a soccer kick. Research Quarterly for Exercise and Sport,65(2), 93–99.
Antúnez, R., Hernández, F., García, J., Vaíllo, R., & Arroyo, J. (2012). Relationship between motor variability, accuracy, and ball speed in the tennis serve. Journal of Human Kinetics,33, 45–53. doi:10.2478/v10078-012-0043-3
Bahamonde, R. (1997).Joint power production duringflat and slice tennis serves. Paper presented at the XV International sympo-sium on biomechanics in sports, Denton, TX.
Bahamonde, R. (2000). Changes in angular momentum during the tennis serve.Journal of Sports Sciences,18, 579–592. Bahamonde, R. (2005). Review of the biomechanical function of
the elbow during tennis strokes.International SportMed Journal, 6(2), 42–63.
Bartlett, R., Wheat, J., & Robins, M. (2007). Is movement varia-bility important for sports biomechanists?Sports Biomechanics, 6(2), 224–243.
Besier, T., Sturnieks, D., Alderson, J., & Lloyd, D. (2003). Repeatability of gait data using a functional hip joint centre and a mean helical knee axis.Journal of Biomechanics, 36(8), 1159–1168.
Bollettieri, N. (2001). Sonic Serves Bollettieri’s tennis handbook. Champaign, IL: Human Kinetics.
Bonnefoy, A., Slawinski, J., Leveque, J. M., Riquet, A., & Miller, C. (2009). Relationship between the vertical racquet head height and the lower limb motions of elite players’ flat serve. Computer Methods in Biomechanics and Biomedical Engineering, 12(1), 55–57.
Bootsma, R., & Van Wieringen, P. (1990). Timing an attacking forehand drive in table tennis. Journal of Experimental Psychology,16(1), 21–29.
Bradshaw, E., Keogh, J., Hume, P., Maulder, P., Nortje, J., & Marnewick, M. (2009). The effect of biological movement variability on the performance of the golf swing in high- and low-handicapped players. Research Quarterly for Exercise and Sport,80(2), 185–196.
Button, C., MacLeod, M., Sanders, R., & Coleman, S. (2003). Examining movement variability in the basketball free-throw action at different skill levels. Research Quarterly for Exercise and Sport,74(3), 257–269.
Campbell, A., Straker, L., O’Sullivan, P., Elliott, B., & Reid, M. (2013). Lumbar loading in the elite adolescent tennis serve: A link to low back pain.Medicine and Science in Sports and Exercise, 45(8), 1562–1568.
Chin, A., Elliott, B., Alderson, J., Lloyd, D., & Foster, D. (2009). The off-break and“doosra”: Kinematic variations of elite and sub-elite bowlers in creating ball spin in cricket bowling.Sports Biomechanics,8(3), 187–198.
Chow, J., Carlton, L., Lim, Y., Chae, W., Shim, J., Kuenster, A., & Kokubun, K. (2003). Comparing the pre- and post-impact ball and racquet kinematics of elite tennis players’ first and second serves: A preliminary study.Journal of Sports Sciences, 21(7), 529–537.
Davids, K., Glazier, P., Araujo, D., & Bartlett, R. (2003). Movement systems as dynamical systems.Sports Medicine,33 (4), 245–260.
Davids, K., Kingsbury, D., Bennett, S., & Handford, C. (2001). Information-movement coupling: Implications for the organization of research and practice during acquisition of self-paced extrinsic timing skills.Journal of Sports Sciences, 19(2), 117–127.
Dupuy, M., Mottet, D., & Ripoll, H. (2000). The regulation of release parameters in underarm precision throwing.Journal of Sports Sciences,18, 375–382.
Elliott, B. (2006). Biomechanics and tennis. British Journal of Sports Medicine,40(5), 392–396.
Elliott, B., Marsh, T., & Blanksby, B. (1986). A three-dimensional cinematographic analysis of the tennis serve. International Journal of Sport Biomechanics,2, 260–271.
Elliott, B., Marshall, N., & Noffal, G. (1995). Contributions of upper limb segment rotations during the power serve in tennis. Journal of Applied Biomechanics,11, 433–442.
Elliott, B., & Saviano, N. (2001). Serves and returns. In E. P. Roetert & J. Groppel (Eds.), World class tennis technique. Champaign, IL: Human Kinetics.
Ericsson, K. A., Krampe, R., & Tesch-Römer, C. (1993). The role of deliberate practice in the acquisition of expert perfor-mance.Psychological Review,100(3), 363–406.
Fleisig, G., Nicholls, R., Elliott, B., & Escamilla, R. (2003). Kinematics used by world class tennis players to produce high-velocity serves.Sports Biomechanics,2(1), 51–64. Gentile, A. (2000). Skill acquistion: Action, movement, and
neu-romotor process. In J. Carr & R. Shepherd (Eds.),Movement science: Foundation for physical therapy in rehabilitation. Rockville, MD: Aspen.
Girard, O., Micallef, J., & Millet, G. (2005). Lower-limb activity during the power serve in tennis: Effects of perfor-mance level. Medicine and Science in Sports and Exercise, 37(6), 1021–1029.
Guarrera-Bowlby, P., & Gentile, A. (2004). Form and variability during sit-to-stand transitions: Children versus adults.Journal of Motor Behaviour,36(1), 104–114.
Hatze, H. (1986). Motion variability–its definition, quantification, and origin.Journal of Motor Behavior,18(1), 5–16.
Holm, S. (1979). A simple sequentially rejective multiple test procedure.Scandinavian Journal of Statistics,6(2), 65–70. Horan, S., Evans, K., & Kavanagh, J. (2011). Movement
varia-bility in the golf swing of male and female skilled golfers. Medicine and Science in Sports and Exercise,43(8), 1474–1483.
Coordination and variability in the tennis serve 11
Katsumata, H. (2007). A functional modulation for timing a movement: A coordinative structure in baseball hitting. Human Movement Science,26(1), 27–47.
Kibler, W. B. (1995). Biomechanical analysis of the shoulder during tennis activities.Clinics in Sports Medicine,14(1), 79–85. Knudson, D. (1990). Intrasubject variability of upper extremity angular kinematics in the tennis forehand drive. International Journal of Sport Biomechanics,6, 415–421.
Knudson, D., & Bahamonde, R. (2001). Effect of endpoint con-ditions on position and velocity near impact in tennis.Journal of Sports Sciences,19, 839–844.
Kudo, K., Tsutsui, S., Ishikura, T., Ito, T., & Yamamoto, Y. (2000). Compensatory coordination of release parameters in a throwing task. Journal of Motor Behaviour, 32(4), 337–345.
Lloyd, D., Alderson, J., & Elliott, B. (2000). An upper limb kinematic model for the examination of cricket bowling: A case study of Mutiah Muralitharan. Journal of Sports Sciences, 18(12), 975–982.
Marshall, R., & Elliott, B. (2000). Long-axis rotation: The missing link in proximal-to-distal segmental sequencing. Journal of Sports Sciences,18, 247–254.
Martin, C., Kulpa, R., Delamarche, P., & Bideau, B. (2013). Professional tennis players’serve: Correlation between segmen-tal angular momentums and ball velocity.Sports Biomechanics, 12(1), 2–14.
McGinnis, P. (2013). Qualititative biomechanical analysis to improve techniqueBiomechanics of sport and exercise(3rd ed). Champaign, IL: Human Kinetics.
Mullineaux, D., & Uhl, T. (2010). Coordination-variability and kinematics of misses versus swishes of basketball free throws. Journal of Sports Sciences,28(9), 1017–1024.
Newell, K. (1986). Constraints on the development of coordina-tion. In M. Wade & H. Whiting (Eds.),Motor development in children: Aspects of coordination and control (pp. 341–360). Boston, MA: Martinus Nijhoff.
Newell, K., Deutsch, K., Sosnoff, J., & Mayer-Kress, G. (2006). Variability in motor output as noise: A default and erroneous proposition? In K. Davids, S. Bennett, & K. Newell (Eds.), Movement system variability. Champaign, IL: Human Kinetics.
Reid, M., Campbell, A., & Elliott, B. (2012). Comparison of endpoint data treatment methods for estimation of kinematics and kinetics near impact during the tennis serve. Journal of Applied Biomechanics,28(1), 93–98.
Reid, M., Elliott, B., & Alderson, J. (2007). Shoulder joint loading in the high performance flat and kick tennis serves. British Journal of Sports Medicine,41(12), 884–889.
Reid, M., Elliott, B., & Alderson, J. (2008). Lower-limb coordination and shoulder joint mechanics in the tennis serve. Medicine and Science in Sports and Exercise, 40(2), 308–315.
Reid, M., Whiteside, D., & Elliott, B. (2010). Effect of skill decomposition on racket and ball kinematics of the elite junior tennis serve.Sports Biomechanics,9(4), 296–303.
Reid, M., Whiteside, D., & Elliott, B. (2011). Serving to different locations: Set-up, toss, and racket kinematics of the profes-sional tennis serve.Sports Biomechanics,10(4), 407–414. Renshaw, I., Oldham, A., Davids, K., & Golds, T. (2007).
Changing ecological constraints of practice alters coordination
of dynamic interceptive actions. European Journal of Sport Science,7(3), 157–167.
Sakurai, S., Reid, M., & Elliott, B. (2012). Ball spin in the tennis serve: Spin rate and axis of rotation.Sports Biomechanics,12(1), 23–29. doi:10.1080/14763141.2012.671355
Scholz, J., Schoner, G., & Latash, M. (2000). Identifying the control structure of multijoint coordination during pistol shoot-ing.Experimental Brain Research,135(3), 382–404.
Smeets, J., Frens, M., & Brenner, E. (2002). Throwing darts: Timing is not the limiting factor.Experimental Brain Research, 144(2), 268–274.
Sprigings, E., Marshall, R., Elliott, B., & Jennings, L. (1994). A three-dimensional kinematic method for determining the effec-tiveness of arm segment rotations in producing racquet-head speed.Journal of Biomechanics,27(3), 245–254.
Stergiou, N., & Decker, L. M. (2011). Human movement varia-bility, nonlinear dynamics, and pathology: Is there a connec-tion?Human Movement Science,30(5), 869–888.
Tanabe, S., & Ito, A. (2007). A three-dimensional analysis of the contributions of upper limb joint movements to horizontal racket head velocity at ball impact during tennis serving. Sports Biomechanics,6(3), 418–433.
Tepavac, D., & Field-Fote, E. (2001). Vector coding: A technique for quanitification of intersegmental coupling in multicyclic behaviours.Journal of Applied Biomechanics,17, 259–270. Van Gheluwe, B., De Ruysscher, I., & Craenhals, J. (1987).
Pronation and endorotation of the racket arm in a tennis serve. In B. Jonsson (Ed.),Biomechanics X-B (pp. 667–672). Champaign, IL: Human Kinetics.
Wagner, H., Pfusterschmied, J., Klous, M., Von Duvillard, S. P., & Müller, E. (2012). Movement variability and skill level of various throwing techniques. Human Movement Science, 31, 78–90.
Wheat, J., & Glazier, P. (2006). Measuring coordination and variability in coordination. In K. Davids, S. Bennett, & K. Newell (Eds.), Movement system variability. Champaign, IL: Human Kinetics.
Whiteside, D., Chin, A., & Middleton, K. (2013). The validation of a three-dimensional ball rotation model.Proceedings of the Institution of Mechanical Engineers, Part P: Journal of Sports Engineering and Technology,227(1), 49–56.
Whiteside, D., Elliott, B., Lay, B., & Reid, M. (2013a). The effect of age on discrete kinematics in the elite female tennis serve. Journal of Applied Biomechanics,29(5), 573–582.
Whiteside, D., Elliott, B., Lay, B., & Reid, M. (2013b). A kine-matic comparison of successful and unsuccessful tennis serves across the elite development pathway. Human Movement Science,32(4), 822–835.
Whiteside, D., Giblin, G., & Reid, M. (2014, July 12–16). Redefining spatial consistency in the ball toss of the profes-sional female tennis serve. In K. Sato, W. Sands, & S. Mizuguchi (Eds.),Proceedings of the XXXII international con-ference on biomechanics in sports, Johnson City, TN.
Winter, D. (1984). Kinematic and kinetic patterns in human gait: Variability and compensating effects.Human Movement Science, 3(1–2), 51–76.
Woltring, H. (1986). A fortran package for generalized, cross-validatory spline smoothing and differentiation. Advances in Engineering Software (1978),8(2), 104–113.
Yandell, J. (1999).Visual tennis. Champaign, IL: Human Kinetics.