KARAKTERISASI SECARA MORFOLOGI DAN BIOMOLEKULER BAKTERI
PENYEBAB VIBRIOSIS PADA UDANG PENAEID
Ince Ayu Khairana Kadriah, Koko Kurniawan, dan Muharijadi Atmomarsono Balai Penelitian dan Pengembangan Budidaya Air Payau
Jl. Makmur Dg. Sitakka No. 129, Maros 90512, Sulawesi Selatan E-mail: inceayu@gmail.com
ABSTRAK
Pendekatan konvensional untuk karakterisasi, dan identifikasi bakteri Vibrio berpendar antara lain didasarkan pada karakterisasi sifat-sifat fenotipik yang berbasis kultur. Karakterisasi yang berbasis biomolekuler dilakukan dengan mengamati karakterisasi gen 16S-rRNA isolat berbasis PCR dan dilanjutkan dengan analisis urutan basa-basanya dengan metode sekuensing. Penelitian ini bertujuan untuk menganalisis keragaman morfologi dan genetik bakteri Vibrio. sp. berpendar yang diisolasi dari tambak-tambak udang di Maros, Takalar dan Pinrang serta dari panti pembenihan di Barru, Banyuwangi dan Bali. Karakteristik morfologi yang diamati adalah bentuk, warna, bau dan kemampuan pendaran koloni. Perbedaan karakteristik kemudian dikonfirmasi secara biomolekuler berdasarkan sekuensing gen 16S-rRNA yang selanjutnya disejajarkan dengan sekuen bakteri yang terdeposit pada NCBI. Hasil pengamatan morfologi menunjukkan bakteri Vibrio patogen memiliki karakteristik morfologi warna koloni hijau, bentuk koloni bundar dengan elevasi cembung dan dapat memendarkan cahaya bila diamati di ruang gelap. Tiga isolat Vibrio patogen (By-1, M-120 dan P-275) teridentifikasi secara biomolekuler sebagai V. harveyi berdasarkan sekuensing gen 16Sr-RNA. Sedangkan isolat T-170 teridentifikasi sebagai V. parahaemolyticus.
KATA KUNCI: karakter morfologi, biomolekuler, gen 16S-rRNA, vibriosis PENDAHULUAN
Kejadian serangan vibriosis yang sering mematikan benur pada panti-panti pembenihan secara massal biasanya diawali dengan fenomena air bercahaya. Pendaran cahaya yang terlihat di air dan di benur disebabkan adanya infeksi bakteri Vibrio berpendar. Bakteri Vibrio berpendar ini biasanya menyerang sistem pembenihan baik telur, larva maupun lingkungan pemeliharaan (Baticados et al., 1990). Populasi bakteri Vibrio yang sudah mencapai quorum bisa berubah sifatnya dari saprofit menjadi patogen dan kemudian bersepakat untuk bersama-sama “menyalakan lampu (biolumenescence)” sambil melepaskan toksin yang dapat mematikan udang dengan cepat.
Pada studi mengenai infeksi bakteri patogen terhadap udang, kegiatan pertama yang biasa dilakukan jika terjadi serangan adalah melakukan analisis gejala klinis hewan budidaya yang terserang penyakit. Setelah itu dilanjutkan dengan analisis patologi anatomi. Analisis ini penting untuk mengetahui jenis bakteri yang menyerang dari ciri morfologi udang yang terserang maupun ciri-ciri morfologi koloni bakteri yang diisolasi dari air, sedimen dan udang sakit.
Pesatnya perkembangan teknik biologi biomolekuler menawarkan peluang baru yang signifikan untuk penelitian diagnosis penyakit ikan. Menggunakan asam nukleat sebagai target, dan metode baru yang menganalisis polimorfisme dalam asam nukleat, dapat meningkatkan spesifisitas, sensitivitas dan kecepatan diagnosis serta menawarkan cara baru untuk memeriksa hubungan antara genotip dan fenotip dari berbagai patogen. Kemajuan dalam teknik PCR sangat membantu studi epidemiologi serta identifikasi penyebab wabah penyakit atau deteksi patogen (Cunningham, 2002). Penelitian ini bertujuan untuk mengetahui jenis bakteri Vibrio berpendar yang sering ditemukan menjadi penyebab utama Vibriosis di tambak-tambak budidaya udang penaeid di Indonesia khususnya di Sulawesi Selatan. Karakterisasi jenis bakteri Vibrio dilakukan dengan dengan mengamati morfologi dan dilanjutkan uji biomolekuler berbasis gen 16S-rRNA.
) dalam tabung reaksi dengan metode gores dan diinkubasi pada suhu 28C. Karakterisasi Secara Morfologi
Isolat terpilih kemudian diamati karakter morfologinya dengan mengamati bentuk koloni, warna, bau dan kemampuan berpendarnya. Pengamatan karakter morfologi isolat bakteri dilakukan menggunakan agar TCBS. Seleksi awal isolat bakteri dilakukan berdasarkan pertumbuhan dan kemampuan pendarannya. Isolat bakteri yang tetap dapat tumbuh dengan baik serta tetap berpendar setelah diremajakan kemudian dipilih untuk uji karakterisasi secara biomolekuler.
Karakterisasi Secara Biomolekuler
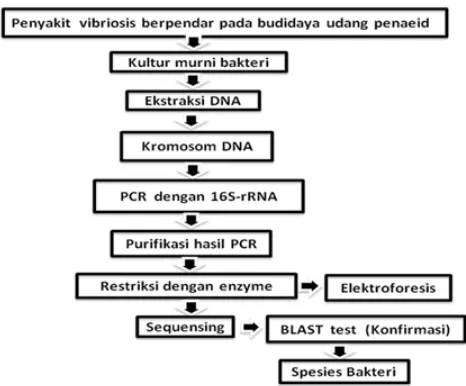
Karakterisasi secara biomolekuler dilakukan dengan membandingkan sekuen gen 16S-rRNA isolat bakteri dengan sekuen gen 16S-rRNA yang terdeposit di NCBI. Pada uji karakterisasi dengan teknik biomolekuler ini dilakukan pengujian untuk tujuan identifikasi spesies. Pada tahap ini diketahui karakter gen 16S-rRNA dari isolat bakteri Vibrio patogen yang diisolasi dari lokasi berbeda berdasarkan urutan DNA hasil sekuensing yang dilanjutkan dengan uji BLAST (Basic Local Alignment Search Tool). Hasil alignment kemudian dibuat pohon filogeninya menggunakan perangkat lunak dari situs European Bioinformatics Institute (EBI) (http://www.ebi.ac.uk/Tools/msa/clustalw2) untuk mengetahui kekerabatan isolat bakteri koleksi dengan bakteri sejenis yang terdeposit pada NCBI (National Center for Biotechnology Information) (Gambar 1).
Sebelum dilakukan karakterisasi secara biomolekuler dengan amplifikasi DNA menggunakan metode PCR, terlebih dahulu dilakukan isolasi genom menggunakan metode Phenol-Chloroform yang telah dikembangkan oleh Parenrengi et al. (2000). Tahapan ekstraksinya diawali dengan kultur bakteri. Bakteri dikultur di dalam Nutrient broth selama 4 jam kemudian dipanen dengan cara sentrifugasi. Sebanyak 1 mL biakan bakteri dipindahkan kedalam tabung mikro steril 1,5 mL dan disentrifugasi (6.000 rpm selama 10 menit). Proses sentrifugasi diulang sebanyak dua kali dan kemudian dilakukan pencucian dengan larutan fisiologis juga dengan sentrifugasi. Pelet bakteri yang dihasilkan kurang lebih 50 mg kemudian dicampur dengan 500 µL lysis bufer, 20 µL proteinase-K (stok 20 mg/mL) dan 40 µL sodium dodecyl sulfate (SDS) 10%. Setelah itu, biakan bakteri diinkubasi dalam waterbath selama 1–3 jam pada suhu 550C. Penambahan 12,5 µL RNAse dilakukan sebelum biakan disimpan pada suhu ruang selama 15-30 menit. Selanjutnya ditambahkan Phenol:Chloroform:Isoamyl alcohol (PCIA 25:24:1) sebanyak 500 µL. Tabung mikro divortex secara perlahan sampai homogen dan disimpan pada suhu ruang selama 10 menit. Selanjutnya disentrifugasi pada kecepatan 13.000 rpm selama 8 menit. Lapisan paling atas diambil dan dipindahkan ke tabung mikro baru dan dilakukan penambahan PCIA seperti sebelumnya. Setelah lapisan paling atas dipindahkan ke tabung mikro baru, dilakukan penambahan satu bagian larutan Chloroform:Isoamyl alcohol (CIA 24:1) dan disentrifugasi selama 4 menit dengan kecepatan 13.000 rpm. Lapisan paling atas dipindahkan ke tabung mikro baru secara hati-hati dengan menghindari interphase. Kemudian dua bagian ethanol absolut dingin ditambahkan ke dalam tabung mikro yang berisi larutan DNA dan dicampur perlahan sampai homogen. Dilakukan sentrifugasi dengan kecepatan 6.000 rpm selama 30 menit. Cairan dibuang kemudian pellet DNA
dicuci dengan 1 mL ethanol 70% kemudian disentrifugasi dengan kecepatan 6.000 rpm selama 15 menit. Pelet DNA dikeringkan selama satu malam dan setelah kering ditambahkan 100 µL Bufer Tris-Etilen Diamin Tetra Aceticacid (EDTA) (TE) dan selanjutnya disimpan pada suhu -200C sampai digunakan. Kualitas DNA genom hasil ekstraksi dapat dilihat melalui analisis elektroforesis pada gel agarosa 0,8%.
Setelah genom bakteri berhasil diisolasi, proses selanjutnya adalah amplifikasi gen 16S-rRNA dari isolat bakteri hasil koleksi dengan metode PCR. Proses PCR dilakukan menggunakan Kit PCR Ready To Go (RTG) dengan pasangan primer 0008F dan 1492RH (Yuhana, 2008). Proses PCR untuk memperbanyak DNA melibatkan serangkaian siklus suhu yang berulang dan masing-masing siklus terdiri atas tiga tahapan yaitu denaturasi, annealing dan elongasi.
HASIL DAN BAHASAN
Isolasi dan Karakterisasi Secara Morfologi Bakteri Vibrio Berpendar
Proses isolasi bakteri Vibrio yang dilakukan dari sampel benur sakit, sedimen tambak, air tambak dan air laut di sekitar Sulawesi Selatan, Banyuwangi dan Bali menghasilkan 35 isolat Vibrio berpendar yang dapat terus tumbuh dengan baik tanpa mengalami kontaminasi dan perubahan morfologi (Tabel 1). Koloni tunggal isolat Vibrio ditumbuhkan pada media agar TSA miring dan disimpan pada suhu 370C. Bakteri hasil isolasi ditumbuhkan pada media selektif Vibrio (TCBSA) untuk diamati karakteristik morfologinya. Koloni bakteri hijau dengan bentuk bundar, tidak beraturan, elevasi cembung dan agak rata, tepian licin dan berlekuk (Gambar 2). Morfologi koloni bakteri Vibrio patogen yang menyerang pada kasus Vibriosis umumnya berwarna hijau (green colony) jika ditumbuhkan di media agar-agar TCBS dengan bentuk koloni bulat, cembung mengkilap, elevasi rata dan terlihat memendarkan cahaya bila diamati di ruang gelap (Moriarty, 1998).
Seleksi awal isolat bakteri dilakukan berdasarkan pertumbuhan dan kemampuan pendarannya. Isolat bakteri yang tetap dapat tumbuh dengan baik serta tetap berpendar setelah diremajakan kemudian dipilih untuk uji karakterisasi secara biomolekuler. Dari 35 isolat bakteri Vibrio berpendar yang diisolasi dan diamati karakter morfologinya, dipilih 12 isolat untuk diuji karakterisasinya secara biomolekuler.
Karakterisasi Secara Biomolekuler Bakteri Vibrio Berpendar
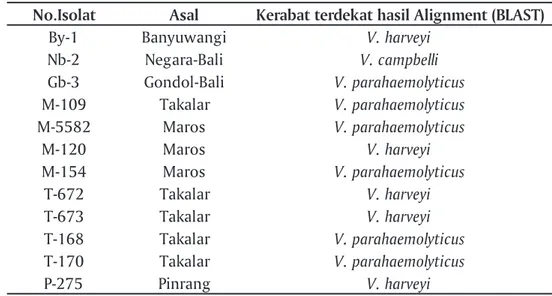
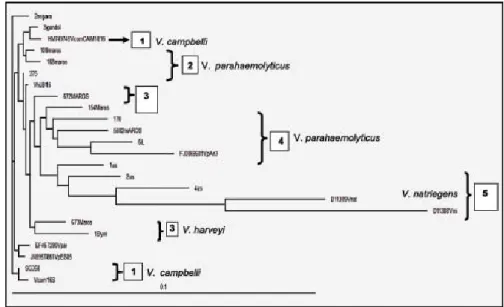
Karakterisasi secara biomolekuler dilakaukan dengan membandingkan sekuen gen 16S-rRNA isolat bakteri dengan sekuen gen 16S-rRNA yang terdeposit di NCBI. Hasil penelusuran sekuen gen Vibrio patogen di NCBI akan dijadikan rujukan untuk menentukan spesies dari koloni bakteri yang dikoleksi. Pada Tabel 2 ditampilkan hasil penelusuran di laman NCBI berupa beberapa sekuen gen 16S-r RNA spesies bakteri Vibrio. Hasil alignment dari 12 isolat terpilih digunakan untuk membuat pohon filogeni dari isolat bakteri Vibrio berpendar menggunakan perangkat lunak EBI (Gambar 3 dan Tabel 3). Pada Gambar 3 dapat dilihat 5 isolat teridentifikasi sebagai V. harveyi (isolat-isolat: By-1, P-275, M-120, T-672, dan T-673), 1 isolat V. campbelli (isolat Nb-2) dan 6 isolat diidentifikasi sebagai V. parahaemolyticus
M-119 Maros Air tambak
M-120 Maros Air tambak
M-128 Maros Air tambak
M-130 Maros Air tambak
M-133 Maros Air tambak
M-137 Maros Air tambak
M-139 Maros Air tambak
M-144 Maros Air tambak
M-145 Maros Air tambak
M-154 Maros Air tambak
M-159 Maros Air tambak
T-671 Takalar Air tambak
T-672 Takalar Air tambak
T-673 Takalar Air tambak
T-675 Takalar Air tambak
T-676 Takalar Air tambak
M-5575 Maros Air tambak
M-5582 Maros Sedimen tambak
M-5584 Maros Sedimen tambak
M-5585 Maros Sedimen tambak
T-168 Takalar Air tambak
T-170 Takalar Air tambak
T-173 Takalar Air tambak
P-275 Pinrang Air tambak
Gambar 2. Karakterisasi morfologi bakteri Vibrio patogen pada media TCBSA. V. harveyi, B. V. campbelli
(isolat-isolat: Gb-3, M-109, M-154, M-5582, T-168 dan T-170) (Tabel 3). Hasil karakterisasi secara molekuler memperlihatkan adanya beberapa perbedaan spesies bakteri yang diperoleh dengan hasil identifikasi spesies dari uji biokimia. Beberapa spesies bakteri yang memiliki kekerabatan genetik yang cukup dekat dengan V. harveyi seperti V. parahaemolyticus, V. alginolyticus dan V. campbellii sulit untuk dibedakan dengan menggunakan metode uji biokimia (Kita-Tsukamoto et al., 1993). Identifikasi
No. Kode Spesies
1. HM749745VcomCAIM1816 V. parahaemolyticus 2. VhLB15 V. harveyi 3. FJ3869581VpAn3 V. parahaemolyticus 4. D11309Vnat V. natriegens 5. D11308Vmi V. mimicus 6. EF467290Vpar V. parahaemolyticus 7. JN5957861VpES05 V. parahaemolyticus 8. SG358 V. campbelli 9. Vcam16S V. campbelli
Tabel 2. Sekuen DNA gen 16S r-RNA dari NCBI yang menjadi rujukan pada Gambar 3
No.Isolat Asal Kerabat terdekat hasil Alignment (BLAST)
By-1 Banyuwangi V. harveyi
Nb-2 Negara-Bali V. campbelli Gb-3 Gondol-Bali V. parahaemolyticus M-109 Takalar V. parahaemolyticus M-5582 Maros V. parahaemolyticus M-120 Maros V. harveyi M-154 Maros V. parahaemolyticus T-672 Takalar V. harveyi T-673 Takalar V. harveyi T-168 Takalar V. parahaemolyticus T-170 Takalar V. parahaemolyticus P-275 Pinrang V. harveyi
spesies dilakukan berdasarkan hasil pensejajaran sekuen DNA isolat bakteri uji dengan sekuen DNA yang terdeposit pada NCBI.
Hasil uji BLAST yang dilanjutkan dengan pembuatan pohon filogeni (Gambar 3) menunjukkan ada lima grup bakteri. Grup 1 menunjukkan isolat Nb-2 secara genetik memiliki kedekatan dengan V. campbelli. Pada grup 2 terlihat bahwa isolat Gb-3 memiliki kedekatan dengan V. parahaemolyticus. Pada grup 3 isolat-isolat P-275, By-1, T-672, dan T-673 teridentifikasi memiliki kedekatan secara genetis dengan bakteri V. harveyi. Pada grup 4 isolat-isolat T-170, M-5582, M-154 dan 6iL (isolat Vibrio patogen dari ikan kerapu) teridentifikasi memiliki kedekatan secara genetik dengan bakteri V. parahaemolyticus. Sedangkan grup 5 isolat 4us dan 2us terlihat memiliki kedekatan dengan V. natriegens dan V. mimicus.
KESIMPULAN
Karakterisasi secara morfologi tidak dapat membedakan antara beberapa spesies bakteri Vibrio berpendar patogenik. Hasil uji karakterisasi secara biomolekuler menunjukkan terdapat 5 kelompok spesies Vibrio berdesarkan kedekatannya secara genetis. Terdapat hubungan kekerabatan yang cukup dekat antara V. harveyi (673) yang diisolasi dari Maros dengan V. harveyi yang diisolasi dari Banyuwangi. Demikian pula dengan V.parahaemolyticus yang diisolasi dari Gondol dengan V.parahaemolyticus yang diisolasi dari Maros (109).
DAFTAR ACUAN
Baticados, MCL., Lavilla-Pitogo, C.R., Cruz-Lacierda, E.R., de la Pena, L.D., & Sunaz, N.A. (1990). Studies on the chemical control of luminous bacteria Vibrio harveyi and V. splendidus isolated from diseased Penaeus monodon larvae and rearing water. Dis Aqua Org, 9, 133-139.
Cunningham, C.O. (2002). Molecular diagnosis of fish and shellfish diseases: present status and potential use in disease control. Aquaculture, 20, 19-55.
Kita-Tsukamoto, K., Oyaizu, H., Nanba, K., & Simidu, U. (1993). Phylogenetic relationships of marine bacteria, mainly members of the family Vibrionaceae, determined on the basis of 16S rRNA sequences. Int J Syst Bacteriol, 43, 8–19.
Moriarty, D.J.W. (1998). Control of luminous Vibrio species in penaeid aquaculture ponds. Aquaculture, 168, 351-358.
Parenrengi, A. (2000). Studies on genetic variability of groupers (Genus: Epinephelus) from Indo-Malaysian waters using PCR-RAPD Analysis. [Thesis master of Science], Terengganu: Kolej University Terengganu, Universiti Putra Malaysia.