Tanggap Genotipe Jagung terhadap Cekaman
Kekeringan: Peranan Akar---Maize Genotypes
Responses to Drought Stress: Roots’ Roles
DATASET · MARCH 2014 READS262
2 AUTHORS, INCLUDING: Roy Efendi Indonesian Agency for Agricultural Research … 4 PUBLICATIONS 0 CITATIONS SEE PROFILETanggap Genotipe Jagung terhadap Cekaman Kekeringan:
Peranan Akar
Roy Efendi dan M. Azrai Balai Penelitian Tanaman Serealia Jl. Dr. Ratulangi 274, Maros, Sulawesi Selatan
ABSTRACT. Maize Genotypes Responses to Drought
Stress: Roots’ Roles. A glasshouse and field studies were
conducted, aiming to evaluate direct responses and tolerances of maize genotypes based on yields and to determine the role of roots in supporting the tolerance of maize plants against drought stress. A split plot design with four replications was used in the field experiment. The main plots were drought stresses and the sub plots were 15 maize genotypes. Drought stress was given from 35 days after planting until harvest. A factorial in split plot design was used in the glasshouse experiment. The treatments consisted of combination between maize genotypes and a solution of polyethylene glycol (PEG) 6000 at 10% concentration. The PEG stress was applied at 10-day-old plants. The laboratory experiment used a completely randomized factorial design with treatment combination of 15 maize genotypes and two concentrations of PEG solution (0% and 15%). Seeds of each maize genotype were germinated using paper towel roll method and germination media corresponded with the appropriate treatments of PEG solution. Results of the experiment showed that drought stress caused decreases in plant height by 32%, leaf area by 30%, leaf chlorophyll content by 45%, percentage of fertile plants by 84%, increases in plant wilting (leaf rolling), time intervals of male and female flowerings of 10 days longer, as well as increases in the root proline contents to 862%. Maize genotypes sensitive to drought condition experienced higher yield decreases, ranged from 68 to 89% as compared to the medium tolerant genotypes, of only 54 to 60%. The largest direct effect of drought stress was on the root dry weight, as was indicated by a significant coefficient of correlation, r = 0.63.
Keywords: Maize, root growth, drought stresses, proline contents
ABSTRAK. Penelitian dilakukan di laboratorium, rumah kaca, dan
lapangan dengan tujuan untuk (1) mengetahui respon dan mengevaluasi toleransi 15 genotipe jagung secara langsung berdasarkan hasil, dan (2) mengetahui peranan akar dalam mendukung toleransi tanaman jagung terhadap cekaman kekeringan. Percobaan lapang dilakukan dengan rancangan petak terpisah dan empat ulangan. Sebagai petak utama adalah cekaman kekeringan, sedangkan anak petak adalah 15 genotipe jagung. Cekaman kekeringan diberikan sejak tanaman berumur 35 hari setelah tanam hingga panen. Percobaan rumah kaca dilakukan dengan rancangan acak kelompok faktorial. Perlakuannya adalah kombinasi antara 15 genotipe jagung dengan pemberian larutan 10% polyethylene glycol (PEG) 6000. Cekaman PEG dilakukan pada saat tanaman berumur 10 hari setelah tanam. Percobaan laboratorium menggunakan rancangan acak lengkap faktorial dengan mengkombinasikan 15 genotipe jagung dan dua konsentrasi larutan PEG (0% dan 15%). Benih masing-masing genotipe jagung dikecambahkan dengan metode uji kertas gulung dalam plastik dan media perkecambahan yang digunakan telah diberi larutan PEG sesuai perlakuan. Hasil penelitian menunjukkan bahwa kondisi cekaman kekeringan menyebabkan penurunan tinggi tanaman 32%, luas daun 30%, kandungan klorofil daun 45%, persentase tanaman
fertil 84%, serta mengakibatkan peningkatan kelayuan tanaman (penggulungan daun), interval waktu berbunga jantan dan betina (ASI) 10 hari lebih lama dan kandungan prolin akar hingga 862%. Genotipe jagung yang peka terhadap cekaman kekeringan mengalami penurunan hasil yang lebih besar, berkisar antara 68-89% daripada genotipe yang agak toleran 54-60%. Pengaruh langsung yang besar terhadap hasil jagung pada kondisi cekaman kekeringan adalah pada bobot kering akar dengan nilai koefisien lintas 0,63.
Kata kunci: Jagung, pertumbuhan akar, cekaman kekeringan, kandungan prolin
B
entuk morfologi, anatomi, dan metabolisme yang berbeda menyebabkan tanaman jagung memiliki respon yang beragam terhadap cekaman kekeringan. Karakter morfologi dan fisiologi yang dilaporkan terkait dengan sifat toleran terhadap cekaman kekeringan antara lain pertumbuhan dan perkembangan akar (Bohn et al. 2006; Vadez 2007), efisiensi penggunaan air, laju kehilangan air melalui transpirasi, densitas stomata (Blum 2005), dan kemampuan melindungi aparatus kloroplas dari kerusakan (Prochazkova et al. 2001; Mittler 2002).Pendekatan utama yang sering digunakan untuk melihat kemampuan tanaman menghadapi cekaman kekeringan adalah (a) kemampuan mengabsorbsi air secara maksimal dengan memperluas dan memper-dalam sistem perakaran, serta (b) kemampuan tanaman mempertahankan turgor melalui penurunan potensial osmotik (Tardieu 1997 dalam Sopandie 2006). Menurut Dubrovsky dan Go´mez-lomeli (2003), strategi tanaman dalam menghadapi cekaman kekeringan, dimulai dari fase perkecambahan dan pertumbuhan vegetatif dengan membentuk formasi akar yang dalam dan percabangan akar yang banyak. Hal ini merupakan ciri penting dari sifat tanaman yang toleran kekeringan. Hasil penelitian menunjukkan bahwa beberapa varietas jagung yang memiliki akar primer yang lebih dalam mampu mengabsorbsi air lebih banyak (Weele et al. 2000). Selain memodifikasi perakaran pada kondisi cekaman kekeringan, tanaman jagung juga mengatur mekanisme tekanan osmotik sel dengan cara mengakumulasi solut kompatibel (osmotic adjustment, OA) di dalam sel. Salah satu senyawa OA yang paling banyak diproduksi di akar adalah prolin (Sharp and Davies 1979).
Evaluasi toleransi tanaman terhadap cekaman kekeringan dapat dilakukan dengan dua pendekatan, yaitu secara langsung berdasarkan penurunan biji relatif yang dihasilkan pada kondisi cekaman kekeringan dibanding kondisi optimum, dan secara tidak langsung dengan mengamati berbagai peubah morfologi dan fisiologi yang terkait dengan sifat toleransi terhadap cekaman kekeringan (Banziger et al. 2000). Penelitian ini bertujuan untuk melihat respon dan mengevaluasi toleransi genotipe jagung secara langsung berdasarkan hasil dan mengetahui peranan akar dalam mendukung toleransi tanaman jagung terhadap cekaman kekeringan.
BAHAN DAN METODE
Percobaan Cekaman Kekeringan di Lapang
Percobaan dilaksanakan di Kebun Percobaan Muneng, Probolinggo, Jawa Timur pada bulan Juli-November 2007. Ketinggian lokasi percobaan adalah 10 m di atas permukaan laut, jenis tanah alfisol, kepadatan tanah 0,92 g/cm3, porositas tanah 65,24%. Kondisi iklim saat
percobaan berlangsung adalah: suhu minimum berkisar 17-240C dan maksimum 30-350C, kelembaban
udara minimum 35-50% dan maksimum 70-86%, evaporasi harian berkisar antara 4,8-8,1 ml. Selama percobaan berlangsung tidak turun hujan, sehingga tanaman tercekam kekeringan sesuai rencana penelitian. Percobaan menggunakan rancangan petak-terpisah dengan empat ulangan. Petak utama adalah kondisi cekaman kekeringan dan optimum, sedangkan anak petak adalah 15 genotipe jagung. Petak terdiri atas satu baris dengan panjang 5 m. Tiap genotipe ditanam dengan jarak tanam 75 cm x 20 cm, satu tanaman per lubang. Pemupukan pertama diaplikasikan pada 10 hari setelah tanam (HST) dengan takaran pupuk urea 100 kg, SP36 150 kg, dan KCl 100 kg/ha. Pemupukan kedua diaplikasikan pada saat tanaman berumur 40 HST dengan 200 kg urea/ha.
Perlakuan kondisi optimum adalah pemberian air pada setiap 12 hari sekali dengan cara dileb. Untuk perlakuan cekaman kekeringan, pemberian air dihentikan pada saat tanaman berumur 35 HST sampai panen.
Pengamatan dilakukan terhadap tinggi tanaman, klorofil daun yang diukur dengan Klorofil Meter SPAD 502, luas daun tongkol, umur berbunga jantan dan betina, skor penggulungan daun (skor 1-5) yang diamati pada kondisi cekaman kekeringan pada pukul 12.00-14.00, persentase tanaman fertil, bobot pipilan biji/ tanaman, dan indeks sensitivitas cekaman kekeringan
(S) berdasarkan bobot biji/tanaman yang dihitung menggunakan rumus yang dikemukakan oleh Fischer dan Maurer (1978): ) X / Xp ( 1 ) Y / Yp ( 1 S − − =
di mana Yp adalah rata-rata bobot biji/tanaman suatu genotipe yang mendapat cekaman kekeringan, Y adalah rata-rata bobot biji/tanaman suatu genotipe yang tidak mendapat cekaman kekeringan, Xp adalah rata-rata bobot biji/tanaman dari seluruh genotipe yang mendapat cekaman kekeringan dan X adalah rata-rata bobot biji/tanaman dari seluruh genotipe yang tidak mendapat cekaman kekeringan. Kriteria untuk penentuan tingkat toleransi terhadap cekaman kekeringan adalah jika nilai S < 0,5 maka genotipe tersebut toleran, jika 0,5 < S < 1,0 maka genotipe tersebut medium toleran, dan jika S>1,0 maka genotipe dinilai peka.
Percobaan Cekaman Kekeringan pada Fase Awal Vegetatif di Rumah Kaca
Seleksi genotipe jagung toleran kekeringan dilakukan pada fase awal vegetatif di rumah kaca Balai Besar Penelitian dan Pengembangan Bioteknologi dan Genetika Pertanian (BB Biogen) pada bulan Oktober 2007-Januari 2008. Penelitian menggunakan rancangan acak kelompok faktorial. Faktor pertama adalah 15 genotipe jagung, sedangkan faktor kedua pemberian larutan polyethylen glycol (PEG) 6000 dengan konsentrasi 10% yang setara dengan -0.19 MPa (Mexal et al. 1975). Sebagai pembanding adalah tanpa pemberian PEG.
Media tanam yang digunakan adalah campuran
cocopeat dan arang sekam padi dengan perbandingan
1:1 (v/v) yang dimasukkan ke dalam pot (diameter 9 cm dan tinggi 20 cm). Cekaman kekeringan dikondisikan pada saat tanaman berumur 10 HST dengan me-nyiramkan larutan PEG pada media tanam. Frekuensi penyiraman PEG adalah dua hari sekali masing-masing 50 ml. Pupuk diberikan dengan cara melarutkan hara pada larutan PEG dengan frekuensi empat hari sekali. Pupuk yang digunakan adalah Growmore dengan dosis 2 g/l. Pengamatan dilakukan terhadap panjang akar primer dan bobot kering akar pada saat tanaman berumur 30 HST.
Percobaan Cekaman Kekeringan pada Fase Perkecambahan di Laboratorium
Percobaan dilakukan di laboratorium benih Institut Pertanian Bogor pada bulan Agustus-September 2007 dengan rancangan acak lengkap faktorial. Faktor
pertama adalah 15 genotipe jagung, sedangkan faktor kedua pemberian PEG 6000 dengan konsentrasi 15% yang setara dengan -0,41 MPa (Mexal et al. 1975). Sebagai pembanding adalah tanpa pemberian PEG. Benih dari masing-masing genotipe dikecambahkan dengan metode uji kertas gulung dalam plastik (UKDdp).
Benih jagung sebanyak 20 biji disusun di atas tiga lembar kertas merang berukuran 30 cm x 20 cm dan ditutup dengan tiga lembar kertas merang yang telah dilembabkan dengan larutan PEG dengan konsentrasi sesuai perlakuan. Kertas merang yang berisi benih digulung dan diinkubasi dalam germinator tipe IPB 72-1. Pengamatan dilakukan pada saat tanaman berumur 5 hari setelah tanam terhadap panjang dan kandungan prolin pada akar primer yang dianalisis dengan metode yang dikembangkan oleh Bates et al. (1973) menggunakan spektrometer dan prolin murni sebagai standar. Sekitar 0,2 g akar primer jagung digerus dalam mortar porselin, dihomogenisasi dengan 10 ml asam sulfosalsik 3%, kemudian disentrifusi dengan kecepatan 6.000 rpm selama 5 menit dan diambil supernatannya. Supernatan ditera sebanyak 10 ml, 2 ml cairan sampel diambil dan direaksi dengan 2 ml asam ninhidrin dan 2 ml asam asetat glasial dalam tabung reaksi, kemudian dipanaskan selama 1 jam pada suhu 100oC. Kemudian
didinginkan dalam air es selama 5 menit dalam ice bath. Campuran tersebut diekstraksi dengan 4 ml toluen. Kromofor yang terbentuk diukur absorbansinya pada panjang gelombang 520 nm dengan spektrofotometer.
HASIL DAN PEMBAHASAN
Respon Pertumbuhan dan Hasil Jagung terhadap Kondisi Kekeringan
Indikator genotipe jagung toleran kekeringan dapat dievaluasi secara langsung berdasarkan penurunan relatif hasil biji dibanding kondisi optimum (Banziger et
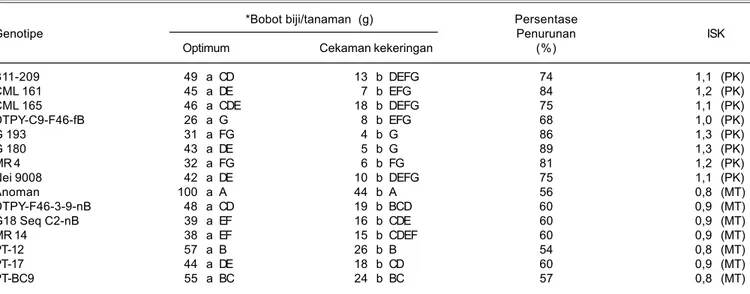
al. 2000). Nilai indeks sensitivitas cekaman kekeringan yang dihitung berdasarkan bobot biji/tanaman menunjukkan bahwa genotipe Anoman, DTPY-F46-3-9-nB, G18 Seq C2-DTPY-F46-3-9-nB, MR 14 PT-12, PT-17, dan PT-BC9 merupakan genotipe medium toleran dengan tingkat penurunan bobot biji berkisar antara 54,1-60,3%, sedangkan genotipe B11-209, CML 161, CML 165, DTPY-C9-F46-fB, G 193, G 180, MR 4, dan Nei 9008 merupakan genotipe peka kekeringan dengan tingkat penurunan bobot biji yang lebih besar, berkisar 68-89% (Tabel 1).
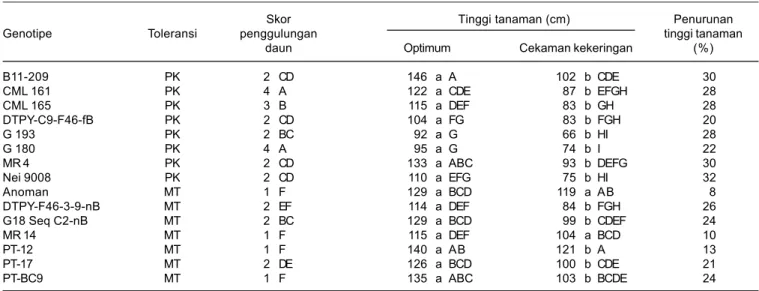
Pada kondisi cekaman kekeringan, tanaman jagung mengalami kelayuan atau penggulungan daun yang merupakan gejala kritis tanaman dan defisit air di mana laju kehilangan air melalui transpirasi lebih besar dibanding laju absorbsi air oleh akar (Banziger et al. 2000). Pada kondisi lengas tanah 12,6%, seluruh genotipe jagung umumnya mengalami tingkat kelayuan (penggulungan daun) yang beragam, hanya MR 14 yang belum menunjukkan gejala kelayuan. Genotipe CML 161 dan G 180 paling peka terhadap cekaman kekeringan dengan gejala penggulungan daun paling besar dengan skor 3,8 dan 4,0, sedangkan genotipe lainnya berkisar antara 1,3-2,5 (Tabel 2).
Tabel 1. Pengaruh cekaman kekeringan terhadap bobot biji jagung dan nilai indeks sensitivitas pada kondisi cekaman kekeringan (ISK). *Bobot biji/tanaman (g) Persentase
Genotipe Penurunan ISK
Optimum Cekaman kekeringan (%)
B11-209 49 a CD 13 b DEFG 74 1,1 (PK) CML 161 45 a DE 7 b EFG 84 1,2 (PK) CML 165 46 a CDE 18 b DEFG 75 1,1 (PK) DTPY-C9-F46-fB 26 a G 8 b EFG 68 1,0 (PK) G 193 31 a FG 4 b G 86 1,3 (PK) G 180 43 a DE 5 b G 89 1,3 (PK) MR 4 32 a FG 6 b FG 81 1,2 (PK) Nei 9008 42 a DE 10 b DEFG 75 1,1 (PK) Anoman 100 a A 44 b A 56 0,8 (MT) DTPY-F46-3-9-nB 48 a CD 19 b BCD 60 0,9 (MT) G18 Seq C2-nB 39 a EF 16 b CDE 60 0,9 (MT) MR 14 38 a EF 15 b CDEF 60 0,9 (MT) PT-12 57 a B 26 b B 54 0,8 (MT) PT-17 44 a DE 18 b CD 60 0,9 (MT) PT-BC9 55 a BC 24 b BC 57 0,8 (MT)
Angka yang diikuti oleh huruf kapital yang sama pada kolom atau oleh huruf kecil yang sama pada baris tidak berbeda nyata berdasarkan uji Duncan 5%, *pada kadar air biji 14%, MT = medium toleran, dan PK = peka.
tinggi tanaman paling besar berkisar antara 30,1%-31,7% (Tabel 2).
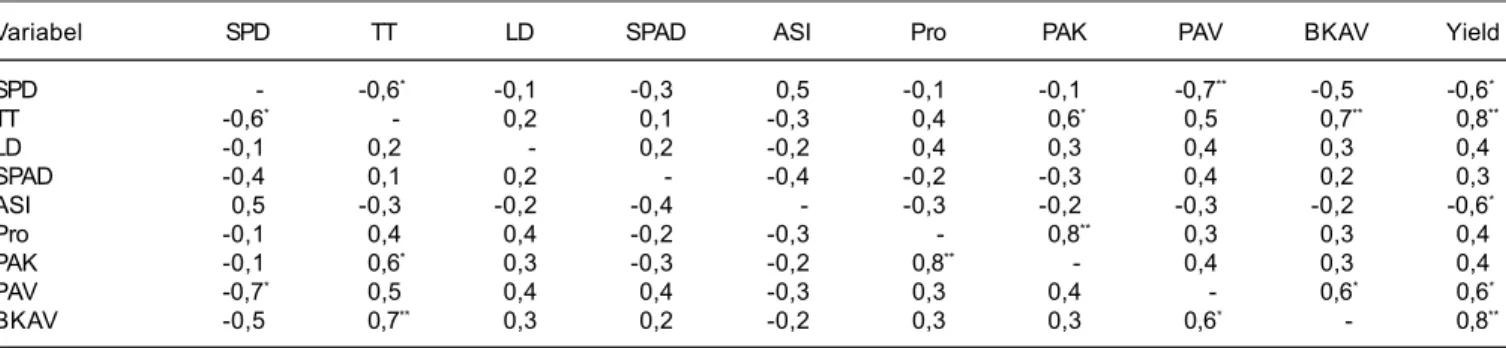
Analisis menunjukkan bahwa skor penggulungan daun berkorelasi negatif sangat nyata dengan panjang akar (R = -0,7), sedangkan tinggi tanaman berkorelasi positif sangat nyata dengan bobot kering akar (R = 0,7) (Tabel 8). Hal ini menunjukkan bahwa kemampuan tanaman menjaga tekanan turgor daun dan Kondisi cekaman kekeringan nyata menekan
pertumbuhan tanaman, namun genotipe medium toleran seperti Anoman dan MR 14 relatif mampu mempertahankan laju pertumbuhan dengan tingkat penurunan tinggi tanaman paling rendah, masing-masing hanya 0,6% dan 8,0%, tidak berbeda nyata antarkedua genotipe pada kondisi optimum. Genotipe peka B11-209, MR 4, dan Nei 9008 mengalami penurunan
Tabel 2. Pengaruh cekaman kekeringan terhadap tinggi tanaman dan penggulungan daun pada saat tanaman jagung berumur 52 hari dengan lengas tanah 12,6%.
Skor Tinggi tanaman (cm) Penurunan
Genotipe Toleransi penggulungan tinggi tanaman
daun Optimum Cekaman kekeringan (%)
B11-209 PK 2 CD 146 a A 102 b CDE 30 CML 161 PK 4 A 122 a CDE 87 b EFGH 28 CML 165 PK 3 B 115 a DEF 83 b GH 28 DTPY-C9-F46-fB PK 2 CD 104 a FG 83 b FGH 20 G 193 PK 2 BC 92 a G 66 b HI 28 G 180 PK 4 A 95 a G 74 b I 22 MR 4 PK 2 CD 133 a ABC 93 b DEFG 30 Nei 9008 PK 2 CD 110 a EFG 75 b HI 32 Anoman MT 1 F 129 a BCD 119 a AB 8 DTPY-F46-3-9-nB MT 2 EF 114 a DEF 84 b FGH 26 G18 Seq C2-nB MT 2 BC 129 a BCD 99 b CDEF 24 MR 14 MT 1 F 115 a DEF 104 a BCD 10 PT-12 MT 1 F 140 a AB 121 b A 13 PT-17 MT 2 DE 126 a BCD 100 b CDE 21 PT-BC9 MT 1 F 135 a ABC 103 b BCDE 24
Angka yang diikuti oleh huruf kapital yang sama pada kolom atau oleh huruf kecil yang sama pada baris tidak berbeda nyata berdasarkan uji Duncan 5%, MT = medium toleran, dan PK = peka.
Tabel 3. Pengaruh cekaman kekeringan terhadap penurunan luas daun dan degradasi klorofil daun pada saat tanaman jagung berumur 75 hari.
Luas daun (cm2) Nilai klorofil daun (unit) Genotipe Toleransi
Optimum Cekaman Persentase Optimum Cekaman Persentase
kekeringan penurunan (%) kekeringan penurunan(%)
B11-209 PK 383 360 6 42 a DE 33 b CDE 22 CML 161 PK 390 382 2 45 a CD 31 b DEF 32 CML 165 PK 398 357 10 50 a ABC 31 b DEF 39 DTPY-C9-F46-fB PK 336 302 10 46 a BCD 39 b AB 14 G 193 PK 385 371 4 52 a AB 43 b A 17 G 180 PK 331 296 11 39 a E 26 b FG 33 MR 4 PK 457 321 30 32 a F 21 b G 35
Nei 9008 PK 391 365 7 50 a ABC 32 b CDE 35
Anoman MT 498 438 12 51 a ABC 38 b ABC 25
DTPY-F46-3-9-nB MT 318 306 4 51 a ABC 41 b A 20 G18 Seq C2-nB MT 363 346 5 49 a ABC 35 b BCD 30 MR 14 MT 432 340 21 45 a CD 33 b BCDE 27 PT-12 MT 335 320 4 54 a A 38 b ABC 29 PT-17 MT 409 302 26 51 a ABC 28 b EF 45 PT-BC9 MT 357 337 6 51 a ABC 33 b BCDE 35 Rata-rata 386 a 343 b
Angka yang diikuti oleh huruf kapital yang sama pada kolom atau oleh huruf kecil yang sama pada baris tidak berbeda nyata berdasarkan uji Duncan 5%, MT = medium toleran, dan PK = peka.
penurunan hasil yang lebih rendah, karena peluang berhasilnya penyerbukan lebih besar dibanding genotipe yang ASI-nya lebih besar. Menurut Earl dan Davis (2003) ASI yang besar mengakibatkan lemahnya perkembangan ovari menjadi sink, sehingga tingkat kesuburan bunga betina menjadi menurun dan berdampak pada penurunan hasil.
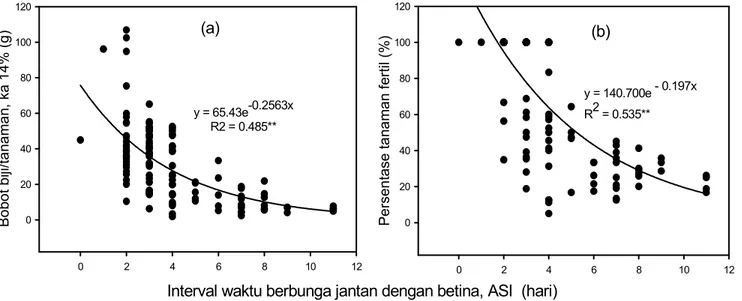
Penurunan hasil pada kondisi cekaman kekeringan disebabkan menurunnya jumlah fertil. Tingkat penurunan terbesar fertilitas pada kondisi cekaman kekeringan terdapat pada genotipe peka G 193 dengan penurunan 84,2%, sedangkan tingkat penurunan terkecil terdapat pada genotipe medium toleran PT-BC9 sebesar 42,4% (Tabel 5). Besarnya penurunan fertilitas berhubungan dengan besarnya ASI. Penurunan tersebut berbentuk eksponensial. Penurunan tajam terjadi pada ASI sekitar 4-8 hari dengan jumlah tanaman fertil berkisar antara 20-60% (Gambar 1).
Larutan PEG dalam media perkecambahan menyebabkan tanaman mengalami cekaman kekeringan, hal ini disebabkan PEG dapat mengikat air sehingga menjadi kurang tersedia bagi tanaman. Dari sifat tersebut maka PEG dapat digunakan untuk simulasi cekaman kekeringan yang meniru tingkat potensial air tanah (Verslues et al., 2006).
Akumulasi prolin pada akar tanaman jagung merupakan respon terhadap kondisi cekaman kekeringan dan merupakan salah satu mekanisme ketahanan terhadap cekaman kekeringan. Pada kondisi cekaman PEG (kekeringan), seluruh genotipe pertumbuhan tanaman (tajuk) ditunjang oleh perakaran
yang dalam dan besar. Perluasan akar yang lebih besar (panjang akar dan bobot kering akar besar) memberi peluang untuk mengabsorbsi air lebih banyak pada lapisan tanah yang lebih dalam dengan lengas tanah lebih besar dibanding di permukaan tanah. Absorbsi air yang cukup oleh akar pada kondisi cekaman kekeringan berpengaruh terhadap kelangsungan pertumbuhan tajuk tanaman.
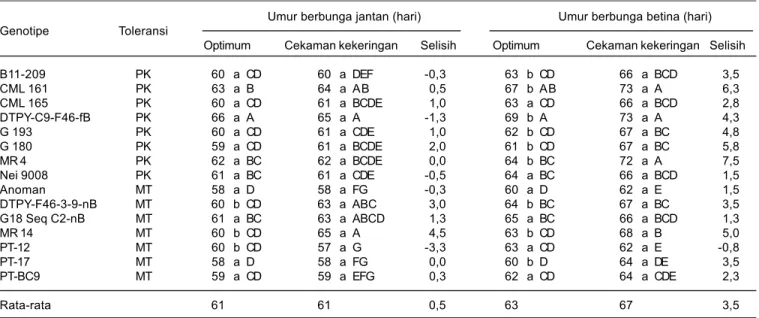
Kondisi cekaman kekeringan menyebabkan pembentukan bunga jantan dan betina menjadi lebih lama, dan pemunculan bunga betina lebih lama dibanding bunga jantan. Pada kondisi cekaman kekeringan, waktu berbunga betina rata-rata 3,5 hari lebih lama dibanding kondisi optimum, sedangkan waktu berbunga jantan hanya 0,4 hari (Tabel 4). Hal ini memperpanjang interval munculnya bunga jantan dan betina (anthesis silking interval, ASI).
Besarnya ASI pada kondisi cekaman kekeringan dipengaruhi oleh genotipe. ASI pada genotipe peka B11-209, CML 161, DTPY-C9-F46-fB, G 180, G 193, dan MR 4 lebih besar, berkisar antara 6– 9,8 hari, sedangkan ASI pada genotipe medium toleran Anoman, G18 Seq C2-nB, dan MR 14 hanya 3,3–3,5 hari (Tabel 5).
Pada kondisi cekaman kekeringan, ASI nyata berkorelasi negatif terhadap hasil dengan nilai koefisen korelasi -0,6 (Tabel 9). Hal ini menunjukkan bahwa ASI yang besar menyebabkan penurunan hasil (bobot biji/ tanaman) secara eksponensial (Gambar 1). Genotipe medium toleran dengan ASI yang lebih kecil mengalami
Tabel 4. Pengaruh cekaman kekeringan terhadap waktu keluarnya bunga betina dan bunga jantan tanaman jagung.
Umur berbunga jantan (hari) Umur berbunga betina (hari) Genotipe Toleransi
Optimum Cekaman kekeringan Selisih Optimum Cekaman kekeringan Selisih
B11-209 PK 60 a CD 60 a DEF -0,3 63 b CD 66 a BCD 3,5 CML 161 PK 63 a B 64 a AB 0,5 67 b AB 73 a A 6,3 CML 165 PK 60 a CD 61 a BCDE 1,0 63 a CD 66 a BCD 2,8 DTPY-C9-F46-fB PK 66 a A 65 a A -1,3 69 b A 73 a A 4,3 G 193 PK 60 a CD 61 a CDE 1,0 62 b CD 67 a BC 4,8 G 180 PK 59 a CD 61 a BCDE 2,0 61 b CD 67 a BC 5,8 MR 4 PK 62 a BC 62 a BCDE 0,0 64 b BC 72 a A 7,5 Nei 9008 PK 61 a BC 61 a CDE -0,5 64 a BC 66 a BCD 1,5 Anoman MT 58 a D 58 a FG -0,3 60 a D 62 a E 1,5 DTPY-F46-3-9-nB MT 60 b CD 63 a ABC 3,0 64 b BC 67 a BC 3,5 G18 Seq C2-nB MT 61 a BC 63 a ABCD 1,3 65 a BC 66 a BCD 1,3 MR 14 MT 60 b CD 65 a A 4,5 63 b CD 68 a B 5,0 PT-12 MT 60 b CD 57 a G -3,3 63 a CD 62 a E -0,8 PT-17 MT 58 a D 58 a FG 0,0 60 b D 64 a DE 3,5 PT-BC9 MT 59 a CD 59 a EFG 0,3 62 a CD 64 a CDE 2,3 Rata-rata 61 61 0,5 63 67 3,5
Angka yang diikuti oleh huruf kapital yang sama pada kolom atau oleh huruf kecil yang sama pada baris tidak berbeda nyata berdasarkan uji Duncan 5%, MT = medium toleran, dan PK = peka.
Tabel 5. Pengaruh cekaman kekeringan terhadap interval waktu berbunga jantan dengan betina (anthesis silking interval, ASI) dan persentase tanaman jagung yang fertil.
Interval waktu berbunga jantan
dengan betina atau ASI (hari) Persentase tanaman fertil (%) Genotipe Toleransi
Optimum Cekaman kekeringan Optimum Cekaman kekeringan Penurunan (%)
B11-209 PK 2,5 b A 6,3 a BC 100,0 a A 28,3 b BCD 71,7 CML 161 PK 3,5 b A 9,3 a A 100,0 a A 27,6 b BCD 72,4 CML 165 PK 2,8 a A 4,5 a CD 100,0 a A 50,9 b A 49,0 DTPY-C9-F46-fB PK 2,5 b A 8,0 a AB 100,0 a A 27,7 b BCD 72,4 G 193 PK 2,5 b A 6,3 a BC 100,0 a A 15,8 b E 84,2 G 180 PK 2,3 b A 6,0 a C 100,0 a A 21,6 b DE 78,4 MR 4 PK 2,3 b A 9,8 a A 100,0 a A 22,8 b CDE 77,2 Nei 9008 PK 3,0 a A 5,0 a CD 100,0 a A 21,2 b DE 78,8 Anoman MT 1,8 a A 3,5 a D 100,0 a A 56,2 b A 43,9 DTPY-F46-3-9-nB MT 3,3 a A 3,8 a D 100,0 a A 48,7 b A 51,4 G18 Seq C2-nB MT 3,3 a A 3,3 a D 100,0 a A 35,7 b B 64,4 MR 14 MT 2,8 a A 3,3 a D 100,0 a A 36,6 b B 63,4 PT-12 MT 2,5 b A 5,0 a CD 100,0 a A 56,1 b A 43,9 PT-17 MT 2,3 b A 5,8 a C 100,0 a A 33,8 b BC 66,2 PT-BC9 MT 2,5 a A 4,5 a CD 100,0 a A 57,6 b A 42,4
Angka yang diikuti oleh huruf kapital yang sama pada kolom atau oleh huruf kecil yang sama pada baris tidak berbeda nyata berdasarkan uji Duncan 5%, MT = medium toleran, dan PK = peka.
y = 65.43e-0.2563x R2 = 0.485**
Interval waktu berbunga jantan dengan betina, ASI (hari)
0 2 4 6 8 10 12 Bo b o t b iji /t a n a ma n , k a 1 4 % ( g ) 0 20 40 60 80 100 120 (a) y = 140.700e - 0.197x R2 = 0.535** 0 2 4 6 8 10 12 Pe rs e n ta s e t a n a ma n f e rt il (% ) 0 20 40 60 80 100 120 (b)
Gambar 1. Hubungan antara interval waktu berbunga betina dan jantan (ASI) dan (a) bobot biji/tanaman dan (b) persentase tanaman jagung yang fertil dengan hasil.
mengalami peningkatan kandungan prolin pada akar primer kecambah, namun peningkatan akumulasi prolin tiap genotipe berbeda-beda. Peningkatan kandungan prolin paling tinggi terdapat pada genotipe medium toleran PT-17 dan PT-BC9, berkisar antara 815,6-862,4 %, sedangkan pada genotipe peka G 180 dan MR 4 hanya 126,4-205,4%. Hal tersebut sejalan dengan hasil penelitian Raymond dan Smirnoff (2002); Sharp et al. (2004); Ogawa dan Yamauchi (2006); Mohammadkhan dan
Heidari (2008) yang menunjukkan bahwa pada kondisi cekaman kekeringan terjadi peningkatan akumulasi prolin yang lebih besar pada ujung akar primer kecambah genotipe toleran dibanding genotipe peka.
Hasil analisis korelasi antarvariabel menunjukkan bahwa pada kondisi cekaman kekeringan, kandungan prolin berkorelasi nyata dengan panjang akar (Tabel 8). Hubungan tersebut berbentuk linier dengan nilai koefisien determinasi 0,6 (Gambar 2). Genotipe yang
Gambar 2. Hubungan antara kandungan prolin pada akar primer dengan panjang akar primer kecambah jagung umur 5 hari setelah tanam.
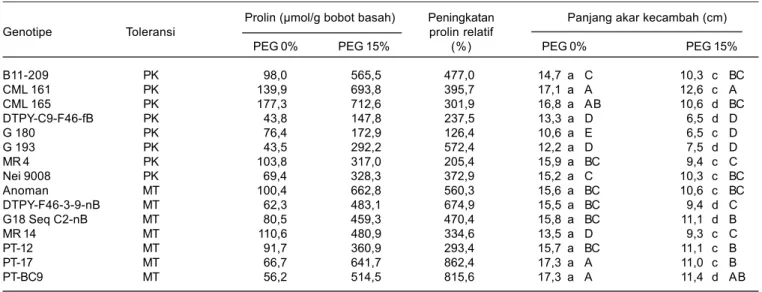
Tabel 6. Kandungan prolin dan panjang akar primer kecambah jagung pada umur lima hari setelah tanam
Prolin (μmol/g bobot basah) Peningkatan Panjang akar kecambah (cm)
Genotipe Toleransi prolin relatif
PEG 0% PEG 15% (%) PEG 0% PEG 15%
B11-209 PK 98,0 565,5 477,0 14,7 a C 10,3 c BC CML 161 PK 139,9 693,8 395,7 17,1 a A 12,6 c A CML 165 PK 177,3 712,6 301,9 16,8 a AB 10,6 d BC DTPY-C9-F46-fB PK 43,8 147,8 237,5 13,3 a D 6,5 d D G 180 PK 76,4 172,9 126,4 10,6 a E 6,5 c D G 193 PK 43,5 292,2 572,4 12,2 a D 7,5 d D MR 4 PK 103,8 317,0 205,4 15,9 a BC 9,4 c C Nei 9008 PK 69,4 328,3 372,9 15,2 a C 10,3 c BC Anoman MT 100,4 662,8 560,3 15,6 a BC 10,6 c BC DTPY-F46-3-9-nB MT 62,3 483,1 674,9 15,5 a BC 9,4 d C G18 Seq C2-nB MT 80,5 459,3 470,4 15,8 a BC 11,1 d B MR 14 MT 110,6 480,9 334,6 13,5 a D 9,3 c C PT-12 MT 91,7 360,9 293,4 15,7 a BC 11,1 c B PT-17 MT 66,7 641,7 862,4 17,3 a A 11,0 c B PT-BC9 MT 56,2 514,5 815,6 17,3 a A 11,4 d AB
Angka yang diikuti oleh huruf kapital yang sama pada kolom atau oleh huruf kecil yang sama pada baris, tidak berbeda nyata berdasarkan uji Duncan 5%, MT = medium toleran, dan PK = peka.
memiliki kandungan prolin tinggi cenderung memiliki akar yang lebih panjang dibanding genotipe yang memiliki kandungan prolin yang rendah (Gambar 2).
Pada kondisi cekaman PEG, pertumbuhan akar genotipe peka DTPY-C9-F46-fB, G 180, dan G 193 lebih tertekan. Panjang akar genotipe tersebut hanya berkisar 6,5-7,5 cm sedangkan genotipe lainnya akar berkisar 9,30-12,55 cm. Hal ini karena genotipe tersebut meng-akumulasi prolin yang lebih rendah pada akar, berkisar 147,8-292,2 μmol/g dibanding genotipe lainnya yang berkisar antara 317,0-693,8 μmol/g bobot segar (Tabel
6). Menurut Sharp et al. (1988); Ogawa danYamauchi (2006) kandungan prolin pada akar primer jagung berperan penting dalam mengatur tekanan osmotik sel sehingga keberlangsungan absorbsi air dapat terjaga untuk menstimulasi pertumbuhan panjang akar pada kondisi cekaman kekeringan.
Panjang dan Bobot Kering Akar Primer pada Fase Awal Vegetatif
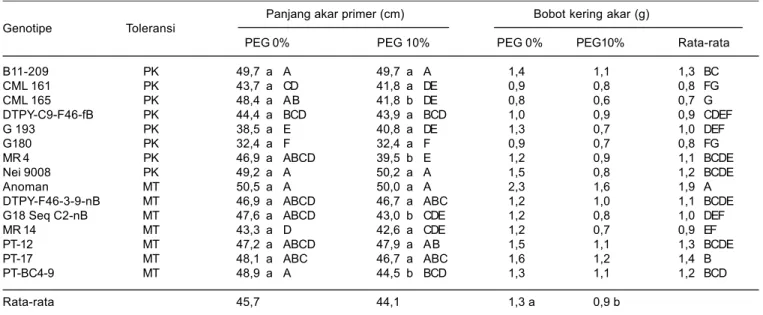
Pada kondisi cekaman PEG, pertumbuhan bobot kering akar mengalami penurunan yang nyata, sedangkan panjang akar beragam. Genotipe Anoman B11-2009, G 193, Nei 9008, dan PT-12 mampu mempertahankan dan meningkatkan panjang akar primer sekitar 1,4-5,8% dibanding kondisi optimum, sedangkan genotipe lainnya mengalami penurunan panjang akar. Tingkat penurunan panjang akar terbesar terdapat pada genotipe peka CML 165 dan MR 4 masing-masing 13,6% dan 21,5% (Tabel 7).
Tingkat penurunan bobot kering akar terbesar pada kondisi cekaman PEG terdapat pada genotipe peka G 193 sebesar 41,3%, sedangkan terkecil terdapat pada genotipe medium toleran yaitu DTPY-C9-F46-fB yang hanya 12,8% (Tabel 7). Bobot kering akar terbesar pada kondisi cekaman PEG terdapat pada genotipe medium Anoman sebesar 1,6 g, sedangkan terkecil pada genotipe peka CML 165 sebanyak 0,6 g (Tabel 23).
Kemampuan meningkatkan luas perakaran pada kondisi cekaman kekeringan dengan cara meningkatkan panjang akar dan memperbanyak cabang akar merupakan kemampuan tanaman dalam
Kandungan prolin akar (µg/gram bobot basah)
0 100 200 300 400 500 600 700 800 900 P a n ja n g a k a r (c m ) 4 6 8 10 12 14 16 18 PEG 0% PEG 15% y = 13.310 + 0.020x R2= 0,141 (ns) y = 6.3140 + 0.007x R2= 0,627 (**)
Kandungan prolin akar (µg/gram bobot basah)
0 100 200 300 400 500 600 700 800 900 P a n ja n g a k a r (c m ) 4 6 8 10 12 14 16 18 PEG 0% PEG 15% y = 13.310 + 0.020x R2= 0,141 (ns) y = 6.3140 + 0.007x R2= 0,627 (**)
Tabel 7. Pengaruh cekaman kekeringan dan penyiraman PEG ke dalam media tanam terhadap panjang dan bobot kering akar pada saat tanaman berumur 30 hari setelah tanam.
Panjang akar primer (cm) Bobot kering akar (g)
Genotipe Toleransi
PEG 0% PEG 10% PEG 0% PEG10% Rata-rata
B11-209 PK 49,7 a A 49,7 a A 1,4 1,1 1,3 BC CML 161 PK 43,7 a CD 41,8 a DE 0,9 0,8 0,8 FG CML 165 PK 48,4 a AB 41,8 b DE 0,8 0,6 0,7 G DTPY-C9-F46-fB PK 44,4 a BCD 43,9 a BCD 1,0 0,9 0,9 CDEF G 193 PK 38,5 a E 40,8 a DE 1,3 0,7 1,0 DEF G180 PK 32,4 a F 32,4 a F 0,9 0,7 0,8 FG MR 4 PK 46,9 a ABCD 39,5 b E 1,2 0,9 1,1 BCDE Nei 9008 PK 49,2 a A 50,2 a A 1,5 0,8 1,2 BCDE Anoman MT 50,5 a A 50,0 a A 2,3 1,6 1,9 A
DTPY-F46-3-9-nB MT 46,9 a ABCD 46,7 a ABC 1,2 1,0 1,1 BCDE
G18 Seq C2-nB MT 47,6 a ABCD 43,0 b CDE 1,2 0,8 1,0 DEF
MR 14 MT 43,3 a D 42,6 a CDE 1,2 0,7 0,9 EF
PT-12 MT 47,2 a ABCD 47,9 a AB 1,5 1,1 1,3 BCDE
PT-17 MT 48,1 a ABC 46,7 a ABC 1,6 1,2 1,4 B
PT-BC4-9 MT 48,9 a A 44,5 b BCD 1,3 1,1 1,2 BCD
Rata-rata 45,7 44,1 1,3 a 0,9 b
Angka yang diikuti oleh huruf kapital yang sama pada kolom atau oleh huruf kecil yang sama pada baris tidak berbeda nyata berdasarkan uji Duncan 5%, MT = medium toleran, dan PK = peka.
memperluas jangkauan akar ke lapisan tanah yang lebih dalam dengan lengas tanah yang lebih besar dibanding di lapisan permukaan tanah, sehingga akar berpeluang besar mengabsorbsi air tanah lebih banyak. Hasil penelitian Grzesiak dan Skoczowski (2004) menunjukkan bahwa genotype jagung yang toleran pada kondisi cekaman kekeringan memiliki pertumbuhan akar yang lebih dalam dibanding genotipe peka. Genotipe medium toleran Anoman memiliki panjang dan bobot kering akar terbesar pada kondisi cekaman kekeringan dengan panjang akar primer 50,0 cm dan bobot kering akar 1,6 g. Hal tersebut me-mungkinkan genotipe Anoman mampu mengabsorbsi air lebih banyak sehingga hanya mengalami kelayuan yang rendah dan mampu mempertahankan pertumbuhan tajuk (Tabel 2) dengan tingkat penurunan hasil yang lebih rendah dibanding genotipe peka (Tabel 1).
Peranan Akar terhadap Toleransi Genotipe Jagung pada Kondisi Cekaman Kekeringan
Analisis antarvariabel menunjukkan bahwa bobot kering akar dan tinggi tanaman berpengaruh sangat nyata terhadap hasil jagung pada kondisi cekaman kekeringan dengan nilai koefisien korelasi masing-masing 0,8 dan 0,78. Namun variabel tersebut juga berkorelasi nyata dengan variabel lainnya sehingga sulit melihat variabel mana yang berpengaruh besar secara langsung terhadap hasil.
Analisis sidik lintas dapat digunakan untuk mengetahui variabel pertumbuhan yang mempunyai pengaruh langsung maupun tidak langsung terhadap hasil jagung pada kondisi cekaman kekeringan. Pengaruh langsung yang besar terhadap hasil jagung pada kondisi cekaman kekeringan adalah bobot kering akar dengan nilai koefisien lintas 0,6 sedangkan pengaruh langsung tinggi tanaman terhadap hasil sangat kecil dengan nilai koefisien lintas 0,2 (Tabel 9). Namun tinggi tanaman berpengaruh tak langsung paling besar terhadap hasil yaitu 0,4 terhadap hasil melalui akar. Hal ini menunjukkan bahwa tinggi tanaman atau bobot biomas yang tinggi berkontribusi besar terhadap pertumbuhan akar pada kondisi cekaman kekeringan. Menurut Moller dan Pellerin (1999); Hebert et al. (2001); Amos dan Walters (2006), pada kondisi cekaman kekeringan bobot biomas berkorelasi secara liner terhadap biomas akar, karena pada kondisi cekaman kekeringan, sebagian besar hasil fotosintat dari tajuk dialokasikan untuk pertumbuhan akar, sehingga jangkauan akar menjadi lebih luas dan dalam untuk mengabsorbsi air pada lapisan tanah yang lebih dalam. Hubungan tersebut juga terlihat pada Tabel 8, di mana hasil analisis korelasi antarvariabel menunjukkan bahwa bobot kering akar berkorelasi positif sangat nyata dengan tinggi tanaman (R = 0,7) (Tabel 8). Beberapa penelitian menunjukkan bahwa kedalaman dan percabangan akar yang banyak (bobot kering akar yang besar) pada kondisi cekaman kekeringan berkorelasi positif dengan kemampuan tanaman untuk mempertahankan
produksi biomas tajuk dan memberi kontribusi nyata dalam menekan kehilangan hasil (Matsura et al. 1996; Wu and Cosgrove 2000; Sinclair and Muchow 2001; Vadez 2007).
Menurut Dubrovsky dan Go´mez-lomeli (2003), strategi tanaman toleran menghadapi cekaman kekeringan dimulai pada fase perkecambahan dan pertumbuhan vegetatif untuk membentuk formasi akar yang dalam dan percabangan akar yang banyak. Perakaran tersebut berpengaruh positif terhadap besarnya absorbsi air oleh akar dan hal ini merupakan ciri penting dari sifat tanaman yang toleran kekeringan. Hasil penelitian menunjukkan pula bahwa beberapa varietas jagung yang memiliki akar primer yang lebih dalam ternyata mampu mengabsorbsi air lebih banyak (Weele et al. 2000).
Dengan mempertimbangkan peranan karakter akar dalam mendukung kemampuan toleransi genotipe jagung terhadap cekaman kekeringan, maka perlu diperhitungkan karakter akar dalam seleksi atau gen pengendali pertumbuhan akar dalam program pemuliaan untuk merakit varietas jagung toleran
cekaman kekeringan. Menurut O’Toole dan Bland 1987, Hochhldinger et al. (2004), keberlangsungan pertumbuhan akar pada kondisi cekaman kekeringan dikendalikan secara genetik, sehingga Grzesiak et al. (1999) menyarankan karakter morfologi akar dapat digunakan secara praktis dalam seleksi toleransi genotipe jagung terhadap cekaman kekeringan secara langsung maupun tidak langsung.
KESIMPULAN
1. Cekaman kekeringan menyebabkan pertumbuhan tanaman, luas daun, kehijauan daun, dan persentase tanaman fertil menurun. Sementara gejala kelayuan (penggulungan daun), interval waktu berbunga jantan dan betina (ASI) serta kandungan prolin akar semakin besar.
2. Genotipe Anoman, DTPY-F46-3-9-nB, G18 Seq C2-nB, MR 14 PT-12, PT-17 dan PT-BC9 merupakan genotipe medium toleran, genotipe B11-209, CML 161, CML 165, DTPY-C9-F46-fB, G 193, G 180, MR 4 Tabel 9. Analisis sidik lintas dari beberapa variabel terhadap hasil jagung pada kondisi cekaman kekeringan.
Variabel Pengaruh SPD TT LD SPAD ASI Pro PAK PAV BKAV
langsung SPD -0,1 -0,1 0,1 0,0 0,1 -0,1 0,0 0,0 0,1 0,1 TT 0,2 -0,1 0,2 0,0 0,0 -0,1 0,1 0,1 0,1 0,1 LD 0,1 -01 0,0 0,1 0,0 -0,0 0,1 0,0 0,1 0,0 SPAD 0,2 -0,1 0,0 0,1 0,2 -0,1 -0,1 -0,1 0,1 0,0 ASI -0,3 -0,2 0,1 0,1 0,1 -0,3 0,1 0,1 0,1 0,1 Pro 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 PAK 0,2 -0,0 0,1 0,1 -0,1 -0,1 0,2 0,2 0,1 0,1 PAV -0,3 0,2 -0,1 -0,1 -0,1 0,1 -0,1 -0,1 -0,3 -0,2 BKAV 0,6 -0,3 0,4 0,2 0,1 -0,1 0,2 0,2 0,4 0,6
SPD = skor penggulungan daun, TT = tinggi tanaman, LD = luas daun tongkol, SPAD = nilai klorofil meter daun, ASI = anthesis silking interval, Pro = kandungan prolin pada ujung akar primer kecambah, PAK = panjang akar kecambah, PAV = panjang akar saat tanaman berumur 30 hari setelah tanam (HST), BAKV = bobot kering akar saat tanaman berumur 30 HST
Tabel 8. Analisis korelasi antarvariabel tanaman jagung pada kondisi cekaman kekeringan atau polyethylen glycol (PEG) 6000.
Variabel SPD TT LD SPAD ASI Pro PAK PAV BKAV Yield
SPD - -0,6* -0,1 -0,3 0,5 -0,1 -0,1 -0,7** -0,5 -0,6* TT -0,6* - 0,2 0,1 -0,3 0,4 0,6* 0,5 0,7** 0,8** LD -0,1 0,2 - 0,2 -0,2 0,4 0,3 0,4 0,3 0,4 SPAD -0,4 0,1 0,2 - -0,4 -0,2 -0,3 0,4 0,2 0,3 ASI 0,5 -0,3 -0,2 -0,4 - -0,3 -0,2 -0,3 -0,2 -0,6* Pro -0,1 0,4 0,4 -0,2 -0,3 - 0,8** 0,3 0,3 0,4 PAK -0,1 0,6* 0,3 -0,3 -0,2 0,8** - 0,4 0,3 0,4 PAV -0,7* 0,5 0,4 0,4 -0,3 0,3 0,4 - 0,6* 0,6* BKAV -0,5 0,7** 0,3 0,2 -0,2 0,3 0,3 0,6* - 0,8**
SPD = skor penggulungan daun, TT = tinggi tanaman, LD = luas daun tongkol, SPAD = nilai klorofil meter daun, ASI = anthesis silking interval, Pro = kandungan prolin pada ujung akar primer kecambah, PAK = panjang akar kecambah, PAV = panjang akar saat tanaman berumur 30 hari setelah tanam (HST), BAKV = bobot kering akar saat tanaman berumur 30 HST
dan Nei 9008 merupakan genotipe peka cekaman kekeringan.
3. Karakter akar merupakan variabel yang paling dominan mempengaruhi penurunan hasil pada kondisi cekaman kekeringan.
4. Untuk merakit varietas jagung yang toleran cekaman kekeringan perlu memperhitungkan karakter akar dalam seleksi genotipe toleran cekaman kekeringan atau mencari gen pengendali pertumbuhan akar.
DAFTAR PUSTAKA
Amos, B. and D.T. Walter. 2006. Maize root biomass and net rhizodeposited carbon: an analysis of the literature. Soil Sci.Soc.Am. 70:1489-1503.
Bänziger, M., G.O. Edmeades, D. Beck, and M. Bellon. 2000. Breeding for Drought and Nitrogen Stress Tolerance in Maize From Theory to Practice. Mexico, CIMMYT.
Bates, L.S., R.P. Waldren, and I.D. Teare. 1973. Rapid determination of free proline for water-stress studies. Plant and Soil (39):205-207.
Blum, A. 2005. Drought resistance, water-use efficiency, and yield potential: are they compatible, dissonant, or mutually exclusive?. Aus. Agri. Research 56: 1159–1168.
Bohn, M., J. Novais, R. Fonseca, R. Tuberosa, and T.E. Grift. 2006. Genetic evaluation of root complexity in maize. Acta Agro. Hungarica. 54(3):1-13.
Dubrovsky, J.G. and L.F. Go´mez-lomeli. 2003. Water deficit accelerates determinate developmental program of the primary root and does not affect lateral root initiation in a sonorant desert cactus (Pachycereus pringlei, cactaceae). American J. Botany (90): 823–831.
Earl, H.J. and R.F. Davis. 2003. Effect of drought stress on leaf and whole canopy radiation use efficiency and yield of maize. Agron. J. (95): 688-696.
Fischer, R.A. and R. Maurer. 1978. Drought resistance in spring wheat cultivar: I. Grain yield response. Aust. J. Agric. Res. (29): 897-912.
Grzesiak, M.T. and A. Skoczowski. 2004. Influence of different soil moisture on leaf water content, chlorophyll fluorescence and seedling growth in maize and triticale genotypes differing to drought tolerance. Z.P.P.N. Rolniczych. 496: 229-240. Grzesiak, S., T. Hura, M.T. Grzesiak, and S. Pienkowski. 1999. The
impact of limited soil moisture and waterlogging stress conditions on morphological and anatomical root traits in maize (Zea mays L.) hybrids of different drought tolerance. Acta Physiologiae Plantarum 21 (3): 305-315.
Hebert, Y., E. Guingo, and O. Loudet. 2001. The response of root/ shoot partitioning and root morphology to light reduction in maize genotypes. Crop Sci. 41:363-371.
Hochhldinger, F., K. Woll, M. Sauer, and D. Dembinsky. 2004. Genetic dissection of root formation in maize (Zea mays) reveals root type specific. 93:359-368.
Levitt, J. 1980. Responses of plants environmental stresses. Vol. 2 Academic Press. New York. p 606.
Matsura, A., S. Inanaga, and Y. Sugimoto. 1996. Mechanism of interspecific differences among four graminaceous crops in growth response to soil drying. Japanese J. of Crop Science. (65): 352-360.
Mexal, J., J.T. Fisher, J. Osteryoung, and C.P. Patrick. 1975. Oxygen availability in polyethylene glycol solutions and its implications in plant-water relations. Plant Physiol. (55): 20-24.
Mittler, R. 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. (50): 405–410.
Mohammadkhan, N. and R. Heidari. 2008. Drought-induced accumulation of soluble sugars and proline in two maize varieties. World Applied Sci. J. (3): 448-453.
Moller, A. and S. Pellerin. 1999. Maize root system growth and development as influenced by phosphorus deficiency. Exp. Botany: 50: 487-497.
Ogawa, A. and A. Yamauchi. 2006. Root osmotic adjustment osmotic stress in maize seedling, 1.Transient change of growth and water relation in roots in response to osmotic stress. Plant Prod. Sci. 9 (1): 27-38.
O’Toole, J.C. and W.L. Bland. 1987. Genotypic variation in crop plant root systems. Advances in Agronomy (41): 91–145. Prochazkova, D., R.K. Sairam, G.C. Srivasta, and D.V. Singh. 2001.
Oxidative stress and antioxidant activity as the basis of senescence in maize leaves. Plant Science 161:765-771. Raymond, M.J. and N. Smirnoff. 2002. Proline metabolism and
transport in maize seedling at low water potential. Annals of botany. 89:813-823.
Sharp, R.E. and W.J. Davies. 1979. Solute regulation and growth by roots and shoots of water-stressed maize plants. Planta 147, 43-49.
Sharp, R.E., W.K. Silk, and T.C. Hsiao. 1988. Growth of the maize primary root at low water potentials. I. Spatial distribution of expansive growth. Plant Physiology (87): 50–57.
Sinclair, T.R. and C.R. Muchow. 2001. System analysis of plant traits to increase grain yield on limited water supplies. Agron. J. 93: 263-270.
Sopandie, D. 2006. Perspektif fisiologi dalam pengembangan tanaman pangan di lahan marjinal. Orasi Ilmiah Guru Besar Tetap Fisiologi Tanaman. Fakutas Pertanian, Institut Pertanian Bogor. 16 September 2006.
Vadez, V. 2007. Exploiting the functionality of root systems for dry, saline, and nutrient deficient environments in a changing climate. In International Crops Research Institute for the Semi-Arid Tropics (ICRISAT), India. 4:1-61.
Verslues, P.E., M. Agarwal, K.S. Agarwal, and J. Zhu. 2006. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. The Plant Journal. 45, 523–539.
Weele, C.M., W.G. Spollen, R.E. Sharp, and T.I. Baskin. 2000. Growth of Arabidopsis thaliana seedlings under water deficit studied by control of water potential in nutrient-agar media. J. Exper. Botany (51): 1555–1562.
Wu, Y. and D.J. Cosgrove. 2000. Adaptation of root to low water potentials by changes in cell wall extensibility and cell wall proteins. J. Exper. Botany (51):1543-1553.