“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 540
Profil Sekuens Sebagian Gen Cytochrome C Oxidase Unit I (COI) Ayam
Hutan Merah (
Gallus gallus
) Sulawesi Utara
Decky D. W. Kamagi
Jurusan Pendidikan Biologi FMIPA, Universitas Negeri Manado, Indonesia Email: deckykamagi@yahoo.com
Abstrak
Penelitian ini bertujuan untuk mengungkap profil sebagian sekuens COI mtDNA ayam hutan merah (Gallus gallus) Sulawesi Utara. Sampel diperoleh dari jaringan otot dari spesimen yang dikumpulkan dari beberapa daerah di Sulawesi Utara. Sampel yang dikumpulkan dipreparasi dengan perlakuan alkohol 95% dan disimpan dalam suhu dibawah 50C. Isolasi DNA total menggunakan innuPREP DNA micro kit dengan protokol yang dimodifikasi. Amplifikasi gen COI menggunakan primer BirdR1 dan BirdF1. Isolasi DNA dan amplifikasi DNA target dikerjakan di Laboratorium Molekuler Universitas Negeri Manado, sedangkan amplikon disekuensing di Laboratorium FirstBase Singapura. Sekuens gen target di blast melalui program blast nukleotida NCBI. Sekuens gen target selanjutnya diselaraskan dengan beberapa aksesi Gallus gallus yang diperoleh dari GenBank melalui program Clustal-W. Pohon filogenetik dibangun menggunakan metode Maximum Likelihood, dengan bootstrap 1000x berbantuan program MEGA5.2 dan analisis sekuens lainnya menggunakan DnaSPv5. Panjang sekuens gen target yang teramplifikasi adalah 490 pb. Hasil blast yang diperoleh menunjukkan bahwa sekuens gen target berpadanan 98% dengan sekuens gen COI beberapa aksesi Gallus-gallus yang diambil dari GenBank. Hasil penyelarasan rangkap antara sekuen-sekuens gen target menunjukkan tidak adanya situs polimorfik, sedangkan penyelarasan antara sekuens gen target dan beberapa aksesi Gallus gallus menunjukkan adanya situs polimorfik (S) atau situs bervariasi. Banyak situs polimorfik (S) adalah 11, sedangkan sekuens terlestarikan (C) adalah 386/397 = 0,977. Banyak haplotipe (h) adalah 4, dan diversitas haplotipe (Hd) adalah 0,703. Pohon filogenetik menunjukkan bahwa ayam hutan merah membentuk klade tersendiri bersama-sama dengan Gallus gallus gallus dan terpisah dari kelompok spesies dan aksesi Gallus gallus lainnya.
Kata Kunci: gen COI, ayam hutan merah (Gallus gallus), Sulawesi Utara PENDAHULUAN
Ayam (Gallus gallus) memiliki variasi jenis dan morfolofis paling besar di antara spesies yang ada di bumi. Ayam tersebar dan mudah ditemukan di beberapa daerah di Indonesia. Ayam dengan berbagai ras dan kelebihan mereka dapat dijumpai hampir di semua tempat. Ayam pedaging, ayam petelur, ayam kampung, ayam untuk adu ketangkasan memiliki sifat yang khas sebagai hasil domestifikasi, persilangan dan pemuliaan (breeding) dan pengembangan keragaman genetik. Proses domestifikasi, persilangan dan pemuliaan dan pengembangan keragaman bermula dari sumber genetik utama, yaitu dari ayam hutan yang ditemukan di beberapa wilayah biogeografi. Ayam hutan merah adalah nenek moyang dari ayam domestik, dan tersebar di Asia tenggara yaitu: Sumatera, Jawa sampai ke Bali, Sulawesi, Filipina, Malaysia, India, Pakistan, Thailand. Ayam hutan merah adalah sebagai nenek moyang utama dari seluruh ayam domestik, didukung beberapa bukti meliputi: temuan arkeologis di lembah Indus, Hebei China, yang diperkirakan hidup pada awal 5400 BC (West dan Zhou, 1988), bukti molekuler berupa sekuens region control mitokondria (Fumihito et al. 1996) dan data mikrosatelit dari berbagai populasi ayam yang ada.
Analisis karakterisrik sekuen gen-gen mitokondria ayam (Gallus gallus) jenis liar yang ada di Sulawesi Utara sekarang ini masih terbatas. Isolasi, amplifikasi dan sekuensing gen target yaitu gen-gen mtDNA pada ayam akan menjadi pembuka jalan
“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 541
untuk kepentingan analisis karakter sekuens gen target tersebut. Menemukan metode yang tepat mengisolasi mtDNA akan mempermudah memperoleh isolat, dan selanjutnya dapat ditindaklanjuti melalui amplifikasi gen melalui PCR, dan pada akhirnya profil sekuens dapat diungkapkan untuk menganalisis variasi genetik. Populasi ayam hutan sebagai sumber daya genetik (plasma nutfah), harus tetap dilestarikan agar sumber daya genetik tidak punah. Sekarang ini populasi beberapa spesies liar terancam punah, akibat perburuan secara ilegal, musuh alami dan karena rusaknya habitat alami oleh ulah manusia dan perubahan iklim. Usaha konservasi sumber daya genetik (plasma nutfah) akan lebih terarah dan berhasil guna apabila karakteristik dan keragaman populasi itu telah diketahui dengan pasti. Informasi mengenai keragaman dan kekerabatan suatu populasi dapat ditelusuri dan diperoleh berdasarkan variasi morfologis (fenotipik), tingkah laku dan variasi genetik dari spesies-spesies yang ada. Pengungkapan keragaman genetik telah banyak dilakukan melalui kajian di tingkat DNA. Sampai saat ini telah banyak dilakukan kajian keragaman genetik dan biologi populasi dengan menggunakan gen DNA mitokondria (mtDNA) (Avise et al., 1979, Brown et al., 1982).
Karakteristik genom mtDNA jenis ayam liar (wild type) Sulawesi Utara sampai sekarang masih terbatas, sehingga perlu dilakukan penelitian dan pengkajian secara mendasar, agar diperoleh sejumlah informasi yang dapat digunakan untuk mengungkapkan keragaman genetik masing-masing jenis, yang nantinya berguna untuk strategi dan program penangkaran atau domestikasi. Pengungkapan keragaman genetik telah banyak dilakukan melalui kajian protein dan isoenzim, namun keragaman genetik yang dapat diungkap tidak maksimal, karena rendahnya polimorfisme protein. Untuk mengatasi masalah tersebut banyak peneliti mengalihkan perhatian pada pengkajian keragaman genetik melalui kajian DNA. Kajian DNA dapat diperoleh hasil yang mampu mengungkap perbedaan yang lebih teliti dalam membedakan intra dan interspesies menyangkut struktur, komposisi, dan organisasi pada tingkat DNA. Materi DNA yang digunakan pada umumnya adalah DNA inti atau DNA total. Sampai saat ini telah banyak dilakukan kajian keragaman genetik dan biologi populasi dengan menggunakan gen atau DNA mitokondria (mtDNA) (Avise et al., 1979, Brown et al., 1982). Studi keragaman genetik interspesifik yang didasarkan pada perbedaan dan persamaan mtDNA dapat menghasilkan rekonstruksi filogenetik beberapa spesies yang saling berdekatan. Pola variasi pada mtDNA dapat digunakan untuk mengelola spesies atau penyidikan spesies yang terancam punah (Moritz, et al. 1987).
Hasil analisis sekuens gen-gen mtDNA dapat ditempatkan pada dua area kepentingan berbeda yaitu: a) konservasi gen, berupa identifikasi dan managemen keragaman genetik, dan b) ekologi gen, yang mana variasi mtDNA digunakan sebagai penuntun dan bantuan untuk kajian demografi dari populasi. Konservasi gen dapat dilakukan berdasarkan informasi filogenetik, dan secara umum sangat relevan untuk rencana jangka panjan, sedangkan ekologi gen yang mengisyaratkan tentang frekuensi alel akan menyediakan informasi yang berguna bagi managemen populasi dalam jangka pendek. Berdasar kajian komparatif, analisis mtDNA dapat mengisyaratkan sinyal kualitatif pada perubahan populasi, yang merupakan target kajian ekologi. Jadi dari beberapa penggunaan mtDNA terutama dipadukan dengan kajian variasi gen inti, akan memberikan kontribusi yang signifikan dalam rencana
Gen-“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 542
gen DNA mitokondria (mtDNA) secara luas telah digunakan untuk menduga hubungan filogenetik diantara primata (Master et al., 2007, Randi (1996), Roos et al., 2008). DNA mitokondria dapat digunakan sebagai indikator yang sensistif pada analisis keragaman genetik di tingkat populasi. Kajian berbagai mtDNA dapat digunakan untuk mengungkapkan kelompok-kelompok geografik dari molekul atau individu yang berkerabat atau hubungan matrilineal (kerabat dari satu ibu) di dalam populasi. mtDNA juga banyak digunakan dalam pengkajian ulang tentang hubungan filogeni di antara taksa yang berkerabat dekat (Moritz et al., 1987). Gen-gen pada mitokondria diketahui sebagai gen yang lebih cepat berkembang dan dapat lebih bervariasi baik di dalam maupun antar spesies serta dapat digunakan untuk mengkaji filogeni dan biogeografi (Roos et al., 2008; Lim, et al., 2010; Randi, 1996; Karanth et al., 2008).
Genom lengkap gen-gen mitokondria Gallus gallus telah dipetakan (Kornegay, et al., 1993) sehingga beberapa gen mitokondria tersebut dapat digunakan sebagai penanda genetik dan dapat digunakan sebagai bahan kajian untuk mengungkap sejarah evolusi sekuens, hubungan kekerabatan dan sebaran biogeografi. Berbagai primer mtDNA untuk keperluan amplifikasi DNA berbasis PCR terutama pada kelompok burung dan vertebrata lainnya telah tersedia (Sorenson, et al., 1999), sehingga sekarang ini sintesis berbagai primer untuk amplifikasi melalui PCR hampir semua gen mtDNA kelompok burung sudah dengan mudah dapat dilakukan, bahkan dapat dirancang sendiri dengan berbagai kepentingan seperti yang telah diuraikan. Sekuens mtDNA ayam Gallus gallus secara umum telah diketahui (Desjardins dan Morais, 1990), namun sekuens itu pada ayam jenis liar (ayam hutan) di Sulawesi Utara masih sangat terbatas bahkan kemungkinan belum pernah diidentifikasi. Sekuens mtDNA yang terlestarikan dari ayam secara umum itu dapat digunakan sebagai pembanding untuk identifikasi dan penelusuran perjalanan evolusioner jenis-jenis ayam yang hidup di Sulawesi Utara. Demikian pula, berdasarkan sekuens beberapa gen dari mtDNA dari ayam secara umum itu dapat dipertimbangkan dalam rekonstruksi filogenetik untuk mengungkapkan hubungan kekerabatan ayam-ayam yang ada di Sulawesi Utara dan beberapa aksesi ayam yang diperoleh dari genBank.
METODE PENELITIAN
Pengambilan dan Preparasi Sampel
Sampel dikumpulkan dari beberapa jaringan otot dan dipreparasi dengan perlakuan alkohol 95% dan disimpan dalam temperatur dibawa 50C.
Ekstraksi DNA
DNA total diekstraksi menggunakan innuPREPDNA micro Kit. Protokol isolasi bersadarkan petunjuk manual dalam kit, dengan modifikasi tertentu.
Amplifikasi Gen Target
Amplifikasi masing-masing gen target menggunakan primer dan Bird R1 dan BirdF1 untuk gen COI. Preparasi komponen dan kondisi PCR dilakukan berdasarkan komponen dan kondisi PCR secara umum dengan modifikasi pada beberapa tahapan tertentu.
Sekuensing
“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 543
Penyelarasan Sekuens (Sequences Alignment)
Sekuens homolog masing-masing sekuens gen target diselaraskan
menggunakan program Clustal-W berbantuan program MEGA5.2. Analisis Karakter Sekuens
Analisis karakter profil masing-masing sekuens gen target menggunakan piranti lunak MEGA5.2, dan DnaSPv5. Analisis karakter profil sekuens masing-masing gen target meliputi pemeriksaan pola dan laju substitusi, jarak genetik dan rekonstruksi pohon filogenetik.
HASIL PENELITIAN
Preparasi Sampel dan Isolasi DNA Total
Spesimen Ayam hutan merah telah diperoleh dari beberapa lokasi di daerah Sulawesi Utara, kawasan Kab. Minahasa. Sampel diperoleh dari sayatan jaringan otot bagian dada beberapa spesimen. Sampel yang diperoleh diberi perlakuan dengan larutan alkohol 95%, dan setiap 4 jam larutan alkohol yang digunakan untuk merendam diganti baru. Sampel tiap spesimen disimpan dalam suhu rendah minimal 50C dibawah titik nol, dan dalam keadaan tanpa larutan alkohol.
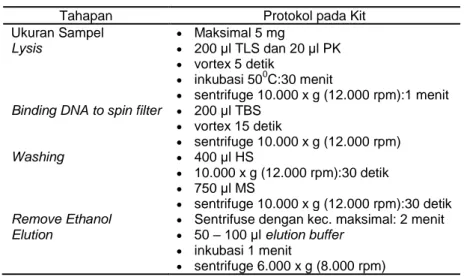
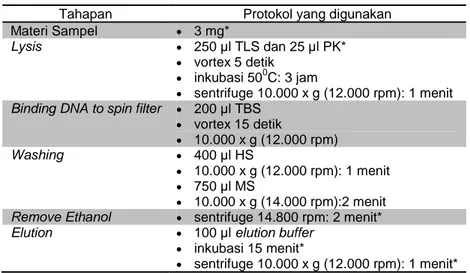
Isolasi DNA total menempuh cara berdasarkan petunjuk manual seperti yang tertera pada protokol kit yang tersedia, namun dengan modifikasi tertentu. Tahapan isolasi DNA total standar yang sesuai protokol pada kit ditunjukkan pada Tabel 1. Modifikasi perlakuan yang telah dilakukan pada tahapan isolasi DNA total dari protokol kit yang tersedia adalah seperti ditunjukkan pada Tabel 2.
Tabel 1. Tahapan isolasi DNA standar dalam protokol kit
Tahapan Protokol pada Kit Ukuran Sampel Maksimal 5 mg
Lysis 200 µl TLS dan 20 µl PK
vortex 5 detik
inkubasi 500C:30 menit
sentrifuge 10.000 x g (12.000 rpm):1 menit Binding DNA to spin filter 200 µl TBS
vortex 15 detik sentrifuge 10.000 x g (12.000 rpm) Washing 400 µl HS 10.000 x g (12.000 rpm):30 detik 750 µl MS sentrifuge 10.000 x g (12.000 rpm):30 detik
Remove Ethanol Sentrifuse dengan kec. maksimal: 2 menit
Elution 50 – 100 µl elution buffer
inkubasi 1 menit
“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 544
Tabel 2. Modifikasi Perlakuan pada Tahapan Isolasi DNA
Tahapan Protokol yang digunakan Materi Sampel 3 mg*
Lysis 250 µl TLS dan 25 µl PK*
vortex 5 detik
inkubasi 500C: 3 jam
sentrifuge 10.000 x g (12.000 rpm): 1 menit Binding DNA to spin filter 200 µl TBS
vortex 15 detik 10.000 x g (12.000 rpm) Washing 400 µl HS 10.000 x g (12.000 rpm): 1 menit 750 µl MS 10.000 x g (14.000 rpm):2 menit
Remove Ethanol sentrifuge 14.800 rpm: 2 menit*
Elution 100 µl elution buffer
inkubasi 15 menit*
sentrifuge 10.000 x g (12.000 rpm): 1 menit* Catatan: Tahapan protokol yang diberi tanda asterix adalah modifikasi perlakuan.
Amplifikasi Gen Target
Preparasi komponen dan kondisi PCR untuk mengamplifikasi gen target dilakukan dengan cara yang sama yaitu mengikuti cara seperti yang ditunjukkan pada Tabel 3 dan Tabel 4. Amplifikasi DNA target dilakukan sebanyak 35 kali.
Tabel 3. Komponen Reaksi PCR
Komponen PCR Konsentrasi Volume Keterangan
DNA template - 2 µL
ddH2O - 34 µL
PCR Mix 5 X 10 µL
MgCl2 1.5 Mm 1.5 µL
dNTPs 20 Mm 1.5 µL
Primer forward (L) 10 pmol µL-1 2 µL
Primer reverse (H) 10 pmol µL-1 2 µL
Tag DNA polymerase 4 – 6 U µL-1
2 µL
Tabel 4. Kondisi Reaksi PCR
Banyaknya siklus Durasi (menit) Temperatur 0C Tahap
35 2 95 Denaturasi awal 0.5 95 Denaturasi 0.5 52 Annealing 0.5 72 Elongasi 1 72 Pasca elongasi
“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 545
Pengukuran Konsentrasi amplikon mtDNA
Hasil pengukuran masing-masing amplikon mtDNA melalui Gene Quant Pro, yaitu suatu alat pengukuran konsentrasi DNA yang menggunakan spectrophotometer absorban ultraviolet pada panjang gelombang 260 nm dan 280 nm ditunjukkan pada Tabel 5.
Tabel 5. Hasil pengukuran amplikon gen COI
Spesimen Sampel Rasio λ260/ λ280 Konsentrasi
Ayam hutan merah AH 1 1,807 75,8
2 1,877 101
Ayam hutan merah AK 1 1,880 135
2 1,822 87,4
Hasil pengukuran amplikon masing-masing gen target melalui elektroforesis gel agarosa 1,5% ditunjukkan pada Gambar 1 sebagai berikut:
Gambar 1. Separasi amplikon gen COI pada gel agarosa 1,5%
AH1dan AH2= Ayam hutan Minahasa Selatan , AK1 dan AK2 =Ayam hutan Minahasa utara Sekuensing
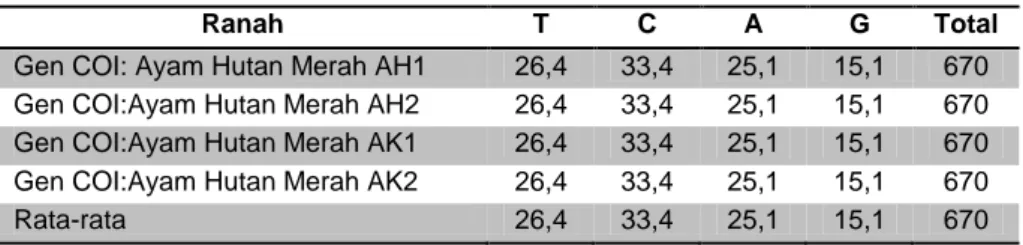
Panjang sekuens gen target yang teramplifikasi adalah 490 pb. Sekuens sebagian gen COI secara lengkap tidak disertakan. Proporsi basa-basa sekuens target ditunjukkan pada Tabel 6. Proporsi basa-basa secara berturut-turut dimulai dari yang paling besar adalah C=33,4%, T=26,4%, A=25,1% dan G=15,1%. Basa-basa pada sekuens semua sampel gen target tidak menunjukkan kejadian substitusi, dengan demikian gen target tersebut 100% identik.
Tabel 6. Proporsi Basa Gen COI (%)
Ranah T C A G Total
Gen COI: Ayam Hutan Merah AH1 26,4 33,4 25,1 15,1 670 Gen COI:Ayam Hutan Merah AH2 26,4 33,4 25,1 15,1 670 Gen COI:Ayam Hutan Merah AK1 26,4 33,4 25,1 15,1 670 Gen COI:Ayam Hutan Merah AK2 26,4 33,4 25,1 15,1 670 Rata-rata 26,4 33,4 25,1 15,1 670
Penyelarasan Rangkap Sekuens
Potongan sekuens gen target dengan panjang 395nt diselaraskan dengan
potongan sekuens-sekuens yang berpadanan dengan sekuens beberapa aksesi Gallus
gallus yang diambil dari GenBank. Hasil penyelarasan ditunjukkan pada Tabel 7. Hasil penyelarasan mengungkapkan ada 11 situs polimorfik atau situs bervariasi. Banyak haplotipe (h) adalah 5, dan diversitas haplotipe (Hd) adalah 0,703 Rata-rata jarak
“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 546
genetik keseluruhan 0,008. Situs-situs polimorfik dan kejadian substitusi basa adalah situs nomor 145 (A↔G), 141 (G↔C), 268 (C↔A), 295 (C↔A), 348 (T↔C), 381(A↔T), 384 (T↔C), 390 (T↔A), 393 (C↔T), 394 (T↔C)dan 395 (C↔T). Substitusi pada situs-situs polimorfik tersebut terdiri atas substitusi transisi dan tranversi. Rasio kejadian substitusi transisi dan transversi (Ts/Tv) = 1.4056. Kejadian beberapa substitusi tersebut terletak pada basa di posisi pertama dan ketiga pada kodon. Kejadian substitusi di situs-situs polimorfik tersebut sebagian menyebabkan pertukaran asam amino ketika ditranslasi. Pertukaran asam amino itu terjadi pada kodon nomor 49, 81, 90, 99 dan 130.
Tabel 7. Penyelarasan Rangkap Sekuens
Ayam Hutan Merah (AH1) aca gga tgg aca gtt tac ccc cct tta gcc ggc aac cta gcc cac gct (48)
Ayam Hutan Merah (AH2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48)
Ayam Hutan Merah (AK1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48)
Ayam Hutan Merah (AK2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48)
Jinhu silky (EU626083.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48)
Dagu (EU626078.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48) EU257689.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48) EU257688.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48) Dagu (EU626072.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48) AUJF1 (JN793568.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48) AULH1 (JN793565.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48)
Gallus gallus bankiva (AB044985.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48)
Gallus gallus gallus(JX178039.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48)
Gallus gallus spadiceus (AB044987.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (48)
Ayam Hutan Merah (AH1) ggc gca tca gta gac cta gcc atc ttt tca tta cac tta gca ggt gtt (96) Ayam Hutan Merah (AH2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96)
Ayam Hutan Merah (AK1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96)
Ayam Hutan Merah (AK2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96)
Jinhu silky (EU626083.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96)
Dagu (EU626078.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96) EU257689.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96) EU257688.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96) Dagu (EU626072.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96) AUJF1 (JN793568.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96) AULH1 (JN793565.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96)
Gallus gallus bankiva (AB044985.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96)
Gallus gallus gallus(JX178039.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96)
Gallus gallus spadiceus (AB044987.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (96)
Ayam Hutan Merah (AH1) tcc tcc att cta gga gcc atc aac ttt atc act acc atc atc aac ata (144) Ayam Hutan Merah (AH2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144)
Ayam Hutan Merah (AK1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144)
Ayam Hutan Merah (AK2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144)
Jinhu silky (EU626083.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144)
Dagu (EU626078.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144) EU257689.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144) EU257688.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144) Dagu (EU626072.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144) AUJF1 (JN793568.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144) AULH1 (JN793565.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144)
Gallus gallus bankiva (AB044985.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144)
Gallus gallus gallus(JX178039.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144)
Gallus gallus spadiceus (AB044987.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (144)
Ayam Hutan Merah (AH1) aaa ccc ccc gca ctg tca caa tac caa aca ccc cta ttc gta tga tcc (192) Ayam Hutan Merah (AH2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192)
Ayam Hutan Merah (AK1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192)
Ayam Hutan Merah (AK2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192)
Jinhu silky (EU626083.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192)
Dagu (EU626078.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192) EU257689.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192) EU257688.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192) Dagu (EU626072.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192) AUJF1 (JN793568.1) g.. ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192) AULH1 (JN793565.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192)
Gallus gallus bankiva (AB044985.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192)
Gallus gallus gallus(JX178039.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (192)
“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 547
Ayam Hutan Merah (AH1) gtc ctc att act gcc atc cta cta ctc ctc tcc tta ccc gtc cta gca (240)
Ayam Hutan Merah (AH2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240)
Ayam Hutan Merah (AK1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240)
Ayam Hutan Merah (AK2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240)
Jinhu silky (EU626083.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240)
Dagu (EU626078.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240) EU257689.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240) EU257688.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240) Dagu (EU626072.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240) AUJF1 (JN793568.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240) AULH1 (JN793565.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240)
Gallus gallus bankiva (AB044985.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240)
Gallus gallus gallus(JX178039.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240)
Gallus gallus spadiceus (AB044987.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (240)
Ayam Hutan Merah (AH1) gct ggg att acc ata cta ctt acc gac cgc aac ctt aac acc aca ttc (288) Ayam Hutan Merah (AH2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288)
Ayam Hutan Merah (AK1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288)
Ayam Hutan Merah (AK2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288)
Jinhu silky (EU626083.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288)
Dagu (EU626078.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288) EU257689.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288) EU257688.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288) Dagu (EU626072.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288) AUJF1 (JN793568.1) c.. ... ... ... ... ... ... ... ... a.. ... ... ... ... ... ... (288) AULH1 (JN793565.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288)
Gallus gallus bankiva (AB044985.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288)
Gallus gallus gallus(JX178039.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288)
Gallus gallus spadiceus (AB044987.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (288)
Ayam Hutan Merah (AH1) ttc gac cca gct gga gga gga gac cca atc cta tac caa cac cta ttc (336) Ayam Hutan Merah (AH2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336)
Ayam Hutan Merah (AK1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336)
Ayam Hutan Merah (AK2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336)
Jinhu silky (EU626083.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336)
Dagu (EU626078.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336) EU257689.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336) EU257688.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336) Dagu (EU626072.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336) AUJF1 (JN793568.1) ... ... a.. ... ... ... ... ... ... ... ... ... ... ... ... ... (336) AULH1 (JN793565.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336)
Gallus gallus bankiva (AB044985.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336)
Gallus gallus gallus(JX178039.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336)
Gallus gallus spadiceus (AB044987.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (336)
Ayam Hutan Merah (AH1) tga ttc ttc ggt cac ccc gaa gtt tac atc ctc atc ctc cca gga ttt (384) Ayam Hutan Merah (AH2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (384)
Ayam Hutan Merah (AK1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (384)
Ayam Hutan Merah (AK2) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (384)
Jinhu silky (EU626083.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ..t ..c (384) Dagu (EU626078.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ..t ..c (384) EU257689.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ..t ..c (384) EU257688.1 ... ... ... ... ... ... ... ... ... ... ... ... ... ... ..t ..c (384) Dagu (EU626072.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ..t ..c (384) AUJF1 (JN793568.1) ... ... ... ..c ... ... ... ... ... ... ... ... ... ... ..t ..c (384) AULH1 (JN793565.1) ... ... ... ..c ... ... ... ... ... ... ... ... ... ... ..t ..c (384) Gallus gallus bankiva (AB044985.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ..t ..c (384) Gallus gallus gallus(JX178039.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... ... (384)
Gallus gallus spadiceus AB044987.1) ... ... ... ... ... ... ... ... ... ... ... ... ... ... ..t ..c (384) Ayam Hutan Merah (AH1) gga att atctc (395) Ayam Hutan Merah (AH2) ... ... ... .. (395)
Ayam Hutan Merah (AK1) ... ... ... .. (395)
Ayam Hutan Merah (AK2) ... ... ... .. (395)
Jinhu silky (EU626083.1) ... ..a ..t .. (395)
Dagu (EU626078.1) ... ..a ..t .. (395) EU257689.1 ... ..a ..t .. (395) EU257688.1 ... ..a ..t .. (395) Dagu (EU626072.1) ... ..a ..t .. (395) AUJF1 (JN793568.1) ... ..a ..t .. (395) AULH1 (JN793565.1) ... ..a ..t .. (395)
Gallus gallus bankiva (AB044985.1) ... ..a ..t .. (395)
Gallus gallus gallus(JX178039.1) ... ... ... ct (395) Gallus gallus spadiceus (AB044987.1) ... ..a ..t .. (395)
“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 548
Rekonstruksi Pohon Filogeni
Berdasarkan estimasi pola substitusi yang terjadi, maka model HKY (Hasegawa-Kishino-Yano) merupakan pola substitusi yang terbaik pada kejadian substitusi pada
sekuens gen COI. Berdasarkan pola substitusi HKY, metode Maximum Likelihood dan
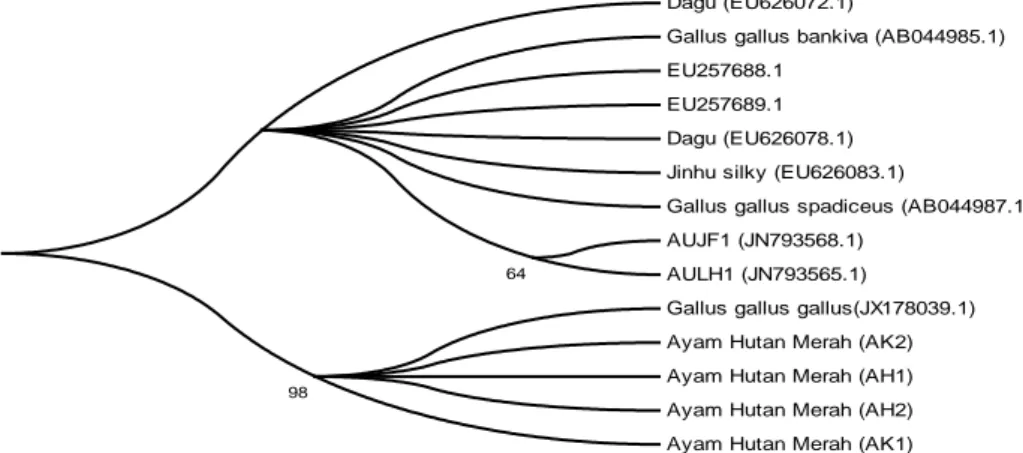
uji Bootstrapping 1000x, rekonstruksi pohon filogeni dapat ditunjukkan pada Gambar 2. Topologi pohon menunjukkan pohon tanpa akar, dimana semua ayam hutan merah sampel bergerombol bersama-sama dengan Gallus gallus gallus dan terpisah dari kelompok-kelompok aksesi lainnya.
Gambar 2. Rekonstruksi pohon filogeni Ayam hutan merah dan beberapa aksesi Gallus gallus yang diambil dari GenBank berbasis gen COI, metode Maximum Likelihood, dan
uji Bootstrapping 1000x.
PEMBAHASAN Profil sekuens
Gen COI (Cytochrome C Oxydase unit I), adalah salah satu gen penyandi protein pada mtDNA. Produk gen COI adalah sitokrom oxidase sub unit I. Pada genom mitokondria ayam, gen COI berlokasi diantara sekuens ke 6651 sampai 8201 dengan panjang 1550 pb, diapit gen tRNA-Tyr dan gen tRNA-Ser (Nishibori et al. 2005). Gen COI mempunyai start codon GTG dan stop codon AAG. Gen COI mengandung kelompok karakter diskrit (masing-masing posisi kodon) yang memaparkan laju mutasi, sehingga dapat dipergunakan sebagai suatu penanda filogenetik (marker phylogenetic) atau sebagai barcode (Desjardins, and Morais. 1990).
Telah diketahui secara umum bahwa gen individual maupun genom keseluruhan secara signifikan beragam dalam komposisi nukleotida. Keragaman komposisi nukleotisa secara luas ditemukan pada DNA inti maupun DNA ekstrakromosomal seperti mtDNA. Keragaman komposisi nukleotida telah terbukti secara signifikan berhubungan erat dengan komposisi asam amino dalam protein (Singer and Hickey, 2002). Keragaman komposisi nukleotida suatu gen atau genom tercermin dari sekuen basa-basa nitrogen yang terkandung di kedua untai nukleotida.
Genom mitokondria vertebrata dan mamalia adalah untai sirkuler tertutup dan memiliki 13 gen penyandi protein, dua gen rRNA, dan 22 gen tRNA. Dua untai yang membentuk genom dikenal sebagai untai H (heavy strand) dan untai L (light strand). Dari 13 gen penyandi protein (coding), 12 berada di untai H dan hanya satu yang di
Dagu (EU626072.1)
Gallus gallus bankiva (AB044985.1) EU257688.1
EU257689.1 Dagu (EU626078.1) Jinhu silky (EU626083.1)
Gallus gallus spadiceus (AB044987.1) AUJF1 (JN793568.1)
AULH1 (JN793565.1)
Gallus gallus gallus(JX178039.1) Ayam Hutan Merah (AK2) Ayam Hutan Merah (AH1) Ayam Hutan Merah (AH2) Ayam Hutan Merah (AK1)
98
“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 549
untai L. Daerah non-coding terutama terbatas pada daerah yang disebut D-loop, dianggap memiliki peran fungsional dalam replikasi dan transkripsi, dan awal mula replikasi untai L (Clayton, 2000). Keragaman komposisi basa pada nukleotida biasanya paling menonjol terdapat pada posisi basa pada kodon sinonim dari gen, karena redundansi dalam kode genetik, dan variasi dalam kandungan DNA yang mungkin memiliki sedikit efek pada kandungan asam amino yang menyusun protein (Singer and Hickey, 2000).
Hasil penyelarasan rangkap sekuens sampel ditambah sekuens yang diambil dari GenBank, ditemukan beberapa situs yang polimorfik sebagai akibat dari kejadian substitusi basa, dimana melibatkan kejadian substitusi transisi maupun transversi. Berdasarkan rasio transisi dan transversi Ts/Tv > 1, maka kejadian substitusi transisi ternyata lebih banyak terjadi daripada substitusi tranversi. Perbandingan kejadian substitusi transisi yang jauh lebih besar daripada substitusi transversi, sejalan dengan yang dikatakan Brown et. al., (1982) bahwa pada gen-gen mitokondria kejadian substitusi transisi lebih dominan daripada substitusi transversi. Tingginya kejadian substitusi transisi dibandingkan transversi adalah biasa terjadi pada sekuen mtDNA eutharian (Brown, et al, 1984). Kocher et. al., (1989) mengemukakan bahwa biasanya pada mtDNA substitusi transisi lebih dominan daripada substitusi transversi, dan saturasi T↔C akan jauh lebih besar dibanding A↔G. Lebih lanjut Kocher, et al., (1989), mengemukakan bahwa substitusi nukleotida pada tingkat spesies sebagian besar adalah transisi sedangkan pada tingkat genus adalah transversi. Hal ini menegaskan bahwa ayam hutan merah dan beberapa aksesi yang diambil dari Gen Bank sesungguhnya berkerabat dekat, karena masih dalam hierarki taksa spesies.
Brown et. al., (1982) mengemukakan bahwa timbulnya kejadian transisi tinggi pada mtDNA adalah sebagai manisfestasi dari meningkatnya tekanan mutasi, namun demikian laju substitusi nukleotida tidak berkorelasi dengan laju perubahan struktural genom dari organela. Pada Hewan tingkat tinggi, mtDNA memang pesat dalam substitusi nukleotida, tetapi susunan dan ukuran gen dari genom tetap untuk setiap spesies (Castro, et al., 1998). Brown et. al., (1982) mengatakan bahwa faktor yang bertanggung jawab sehingga tingginya laju mutasi mtDNA melampaui DNA inti antara lain: a) kecenderungan rusaknya sistim replikasi, b) kurang efisiensinya fungsi pengeditan dan c) tingginya tingkat pertukaran (turnover).
Profil sekuens yang dicirikan oleh proporsi masing-masing basa yang dimiliki masing-masing sampel yang dikaji, ternyata kandungan basa C lebih besar daripada basa-basa lainnya, sedangkan yang paling kecil adalah basa G. Begitu pula proporsi basa-basa yang dimiliki oleh masing-masing sampel ditambah dengan beberapa aksesi yang siambil dari Gen Bank, proporsi basa C adalah yang terbesar dan basa G yang paling kecil. Berkenaan dengan itu Avise (1994) mengemukakan bahwa ciri khas gen penyandi protein pada mitokondria adalah komposisi basa G yang rendah dan C yang tinggi. Temuan Irwin et al., (1991) pada beberapa vertebrata mamalia ternyata menunjukkan bahwa frekuensi basa G di semua posisi pada kodon adalah rendah dan pada manusia frekuensi basa G adalah paling rendah.
Filogeni
Domestikasi ayam dari jenis liar berawal di Asia tenggara, kemudian menyebar ke China, dan selanjutnya menyebar sampai ke Eropa (West and Zhou, 1988). Ayam
“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 550
hutan yang hidup di Asia Tenggara terdiri atas terdiri atas: Gallus gallus gallus (ayam hutan merah), Gallus lafayettei (ayam hutan La Feyette) dan Gallus varius (ayam hutan hijau)(Sawai et al. 2010). Ayam hutan merah memiliki dimorfik seksual mencolok dengan jantan yang memiliki pial berdaging merah, dan tersebar hampir di semua daerah. Ayam La Feyette secara morfologis menyerupai ayam hutan merah, tapi hanya ditemukan di Sri Lanka. Ayam hutan hijau hanya ditemukan di pulau Jawa, Bali dan Lombok. Dua subspesies ayam hutan merah yaitu Gallus gallus gallus dan Gallus gallus spadieus, adalah nenek moyang dari ayam domestik, lain halnya dengan Gallus gallus bankiva yang tidak berkontribusi (Fumihito et al.1996). Hasil rekonstruksi pohon filogenetik berbasis gen COI mtDNA, dimana ayam hutan merah Sulawesi Utara yang berkelompok dengan Gallus gallus gallus dan terpisah dengan kelompok subspesies lainnya, memberikan bukti dan dukungan bahwa ayam hutan merah Sulawesi Utara berkerabat dekat dengan Gallus gallus gallus, atau dengan kata lain ayam hutan merah Sulawesi Utara terhimpun dalam rumpun subspesies Gallus gallus gallus. PENUTUP
Kesimpulan dan Saran
Sekuens sebagian gen COI ayam hutan merah Sulawesi Utara menunjukkan profil yang spesifik. Berdasarkan sebagian gen COI ayam hutan merah Sulawesi Utara berkerabat dekat dengan Gallus gallus gallus. Perlu dilakukan penelitian lebih lanjut untuk mendapatkan informasi lebih lengkap mengenai profil sekuens dan sejarah evolusioner ayam hutan merah Sulawesi Utara melalui pengungkapan seluruh genom mtDNA atau melalui penggunaan penanda genetik lainnya.
DAFTAR RUJUKAN
Avise, J. C., R. A., Lansman, and R. O., Shade, 1979. Use endonuclease to Measure Mitochondrial DNA Sequence Relatedness in Natural Population. I. Population Structure and Evolution in Genus Peromyscus. Genetics 92:279-295.
Brown, W. M., E.M. Prager, A.Wang, and A.C.Wilson, 1982. Mitochondrial DNA sequences of Primates: Tempo and Mode of Evolution. Journal of Molecular Evolution. 18:225-239.
Desjardins, P., & R. Morais. 1990. Sequence and gene organization of the chicken mitochondrial genome. Journal of Molecular Biology 212: 599-634.
Fumihito A, Miyake T, Takada M, Shingu R, Endo T, et al. (1996) Monophyletic origin and unique dispersal patterns of domestic fowls. Proc Natl Acad Sci U S A 93: 6792-6795.
Karanth, K.P., S. Lalji, R. V. Collura and S.C. Beth. 2008. Molecular phylogeny and biogeography of langurs and leaf monkey of South Asia. Molecular Phylogenetic Evolution. 46:683-694.
Kornegay,J.R., Kocher,T.D., Williams,L.A. and Wilson,A.C. 1993. Pathways of lysozyme evolution inferred from the sequences of cytochrome b in birds. Journal Molecular Evolution. 37 (4), 367-379.
Lim, L. S., K. C. Ang, M.C. Mahani, A.W. Shahrom and B.M. Md-Zain. 2010. Mitochondrial DNA polymorphism and phylogenetic relationships of proto malays in peninsular Malaysia. Journal of Biology Science. 10:71-83.
Master, J. C., M. Boniotto, S. Crovella, C. Roos, L. Pozzi, and M. Delpero, 2007. Phylogenetic relationships Among the Lorisoidea As Indicates by Craniodental
“Menyiapkan Generasi Cerdas Berwawasan Lingkungan di Abad 21” 551
Morphology and Mitochondrial Sequence data, American Journal of Primatology
69:6-15.
Moritz, C., T. E. Dowling and W. M. Brown. 1987. Evolution of animal mitochondrial DNA. Annual Review of Ecology and Systematics. 18: 269-292.
Nishibori M, Shimogiri T, Hayashi T, Yasue H (2005) Molecular evidence for hybridization of species in the genus Gallus except for Gallus varius. Animal Genetics 36: 367-375.
Randi, E. (1996). A mitochondrial cytochrome b phylogeny of the Alectoris partridges. Mol. Phylogenet. Evol. 6: 214–227.
Sawai H., Kim H. L., Kuno K., Suzuki S., Gotoh H., Takada M. , Takahata N., Satta Y., Fumihito A. 2010. The Origin and Genetic Variation of Domestic Chickens with Special Reference to Junglefowls Gallus g. gallus and G. varius. PlosOne, Volume 5-5.
Roos, C., T. Nadler and L. Walter, 2008. Mitochondrial phylogeny, taxonomy and biogeography of silver langur spesies group (Trachypithecus cristatus). Molecular, Phylogenetics Evolution 47:629-636.
Sorenson, M.D., J.C. Ast, D.E. Dimcheff, T. Yuri and D.P. Mindell. 1999. Primers for a PCR based approach to mitochondrial genome sequencing in birds and other vertebrates. Molecular Phylogenetics and Evolution 12: 105-114.
West B. and Zhou B. (1988) Did chickens go north? New evidence for domestication. Journal of Archaeological Science 15: 515-33.