www.elsevier.com / locate / bres
Short communication
Upregulation of glial clusterin in brains of patients with AIDs
a ,
*
b a a˜
Jorge E. Torres-Munoz
, Maximino Redondo , Catherine Czeisler , Brenda Roberts ,
a a
Noe Tacoronte , Carol K. Petito
a
Department of Pathology, University of Miami Medical School, Miami FL 33136, USA
b
Hospital Costa del Sol, the Department of Immunology, Marbella, Spain
Accepted 3 October 2000
Abstract
Since clusterin (CLU) production in reactive astrocytes may be neuroprotective, we examined its distribution in AIDS brains where brain injury and reactive astrocytosis are common. The relative area and number of CLU-positive astrocytes, as well as their percent total of all white matter glia, significantly increased in AIDS brains with and without HIV encephalitis (P,0.05). Proliferation markers were absent. In contrast, the relative area and number of GFAP-positive astrocytes and their percent of all white matter glia, increased in some cases but the mean increases were not significant. Clusterin is sensitive marker of glial reactivity in AIDS brains and its enhanced expression was not dependent on increases in GFAP. 2001 Elsevier Science B.V. All rights reserved.
Theme: Development and regeneration
Topic: Glial and other non-neuronal cells
Keywords: Astrocyte; Glial fibrillary acidic protein; AIDS; Gliosis; Clusterin
Clusterin (CLU) is an amphiphilic, heterodimeric CLU following experimental brain injury [22,33] and in glycoprotein that is widely distributed in systemic organs, disease states such as inflammation and neurodegeneration brain, plasma and cerebrospinal fluid (for review, see [18,32]. Clusterin expression in human oligodendrocytes is [3,10]). An initially diverse nomenclature has been re- controversial although increases following Wallerian de-placed with the term ‘clusterin’ for all members of this generation in rat brain [16,17] suggests a reparative
family of glycoproteins [8]. process in these cells [17].
Clusterin’s multiple functions include cell differentiation Potential mechanisms of CLU neuroprotection include and maturation, complement inhibitory effects, cell aggre- its membrane stabilizing properties [32]; its proposed role gation, lipid transport, membrane stabilization, and facilita- in synaptic plasticity and remodeling [2,4,18], its protec-tion of cell to cell contact. More recently, the role of CLU tion against complement-mediated cell death [7] and its as a heat shock-like protein that stabilizes stressed proteins role as a functional heat shock protein [19,31]. Because of
has been proposed [19,31]. the neuroprotective properties of CLU, and its increased
In brain, CLU contributes to development, injury and production in reactive astrocytes, we examined CLU repair. It progressively increases in neurons during brain distribution in brains of patients with the acquired im-maturation [21] and localizes to astrocytes, ependyma and munodeficiency syndrome (AIDS) since brain injury and certain neuronal populations in the adult [5,22]. Clusterin reactive astrocytosis are common in gray and white matter has anti-apoptotic properties (for review, see [13]) and is of these patients [26].
expressed in cells that survive in vitro programmed cell For this present study, we used immunohistochemistry death [6]. Reactive astrocytes are the major producers of to identify CLU, glial fibrillary acidic protein (GFAP), and Ki67 proliferation antigen (MIB-1) in deparaffinized fron-tal lobe sections of four AIDS brains with HIVE, four
*Corresponding author. Tel.: 11-305-243-3584; fax: 1
1-305-243-AIDS cases with normal brains (HIVnE) and four
HIV-4086.
˜
E-mail address: [email protected] (J.E. Torres-Munoz). negative controls. All four HIVE cases were men; two of
298 J.E. Torres-Munoz et al. / Brain Research 888 (2001) 297 –301
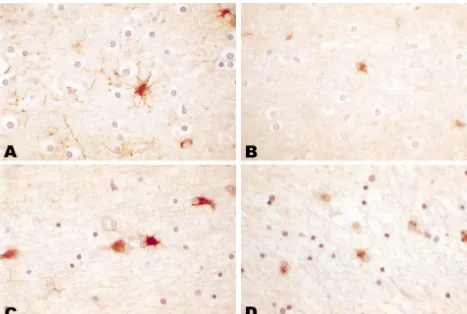
four HIVnE cases were men; and one of four controls were morphology (Fig. 1B). Most co-localized with GFAP, men. A preliminary report has been published [27]. We confirming their astrocytic identify (data not shown). In the determined the relative number and percent total of GFAP- AIDS cases, white matter GFAP-positive astrocytes had positive astrocytes and CLU-positive glial cells in cerebral slightly larger nuclei and more defined cell bodies,
espe-2
white matter by point counting, using a 10 mm grid cially in the HIVE patients with gemistocytic astrocytosis, ocular reticule at 4003 or 6003 magnification and count- characterized by large eosinophilic cell bodies from which ing all immunoreactive cells whose nuclei lay under the radiated cytoplasmic processes (Fig. 1C).
Clusterin-posi-2
intersection of two grid lines per mm . We estimated tive glia also appeared to have slightly larger more relative size of GFAP and CLU-positive cells also by point vesicular nuclei and as well as larger cell bodies with counting and expressed the results as percent area of thicker processes (Fig. 1D). In the cerebral cortex,
as-2
GFAP and CLU-positive cytoplasm per mm . Results were trocytes showed mild increases in size of GFAP and expressed as means of twenty or more microscopic fields CLU-positive cells in AIDS when compared with controls. and group differences were determined with analysis of These changes generally confined to subpial and perivascu-variance and the Kruskal–Wallis non-parametric test; if lar locations. Neuronal CLU in both controls and AIDS statistical differences were found, we then applied the cases was intense and had a perinuclear particulate stain-Mann–Whitney test to compare groups (AIDS with HIVE, ing, consistent with a Golgi apparatus localization. In both AIDS without HIVE and controls). We evaluated CLU controls and AIDS patients, MIB-1 immunoreactive nuclei mRNA expression by in situ hybridization (ISH) as were confined to intravascular and perivascular spaces and previously described [24] on deparaffinized sections of absent or rare in gray and white matter.
frontal cortex and white matter, using digoxygenin (DIG)- CLU mRNA was present in neurons and astrocytes in labeled cRNA probe for CLU [12]. Dig-labeled Poly T three of three controls and four of six AIDS brains. Its controlled for RNA integrity and normal rat brain con- distribution and intensity was variable and paralleled that trolled for CLU mRNA. Sections were evaluated blindly. of poly T ISH; differences between controls and AIDS Two controls had normal brains and one each had were not detected. Co-localization of CLU mRNA with metastatic adenocarcinoma and lacunar infarcts in remote GFAP-positive cells confirmed the astrocytic origin of brain regions. Frontal lobe sections were normal. All four CLU-positive glia.
HIVnE cases had normal brains, including the frontal lobe Quantitative estimates of the relative cytoplasmic area sections. Two HIVE cases had lymphoma or cryptococcal and number of white matter CLU-positive glia were meningitis in remote brain regions. Frontal lobe sections approximately 10-fold lower than those of GFAP-positive were normal in one and contained focal or diffuse white glia in controls as well as in the AIDS cases (Table 1). matter gemistocytic astrocytosis in three. Table 1 summa- However, the relative cytoplasmic area, total number and rizes patient age, post-mortem intervals and means6S.D. the percent total of CLU-immunoreactive glia in white for relative numbers and percent total numbers of CLU and matter was significantly increased in the two AIDS groups GFAP immunoreactive cells and of their relative cyto- when compared with the controls (P#0.05) (Fig. 2). The
2
plasmic area per mm . relative numbers of CLU-positive were higher in the
In controls, white matter astrocytes had small vesicular HIVnE cases than the HIVE cases and the relative cyto-nuclei surrounded by thin GFAP-positive cell bodies and plasmic area of the CLU-positive glia was higher in the thin, delicate processes (Fig. 1A). Clusterin-positive glia HIVE cases than the HIVnE cases; however, these differ-also had vesicular nuclei surrounded by smaller cell bodies ences were not significant (P.0.05). In contrast to the and shorter thin processes, consistent with an astrocyte CLU results, the relative cytoplasmic area, relative number
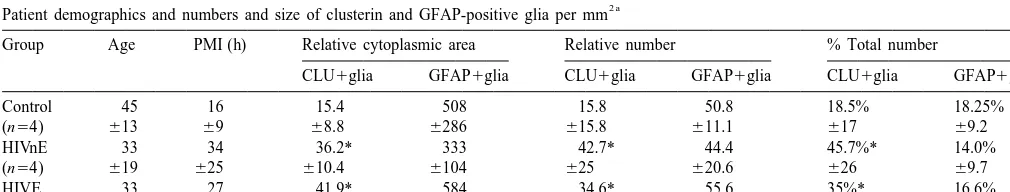
Table 1
2 a
Patient demographics and numbers and size of clusterin and GFAP-positive glia per mm
Group Age PMI (h) Relative cytoplasmic area Relative number % Total number
CLU1glia GFAP1glia CLU1glia GFAP1glia CLU1glia GFAP1glia
Control 45 16 15.4 508 15.8 50.8 18.5% 18.25%
(n54) 613 69 68.8 6286 615.8 611.1 617 69.2
HIVnE 33 34 36.2* 333 42.7* 44.4 45.7%* 14.0%
(n54) 619 625 610.4 6104 625 620.6 626 69.7
HIVE 33 27 41.9* 584 34.6* 55.6 35%* 16.6%
(n53) 617 67 617.7 6492 625 628.6 627 616
a
Abbreviations: GFAP: glial fibrillary acidic protein; PMI: post-mortem interval; HIVnE: AIDS patients with normal brains; HIVE: AIDS patients with HIV Encephalitis. Relative number and percent total represent the relative number of immunoreactive glia and their percent total of all glial cells per 1
2 2
J.E. Torres-Munoz et al. / Brain Research 888 (2001) 297 –301 299
Fig. 1. In controls, GFAP-positive astrocytes have vesicular nuclei, small cell bodies, and thin, delicate processes (A). Clusterin-positive cells have a similar nuclear morphology but smaller cell bodies and fewer processes (B). In AIDS brains, GFAP-positive astrocytes had larger cell bodies but no apparent increase in numbers (C). In AIDS brains, clusterin-positive cells are increased in size and number and have the nuclear and cellular morphology of astrocytes (D). Original magnification3600. hematoxylin counterstain.
and the percent total of GFAP-positive astrocytes were astrocytic in origin. It is likely that the CLU increase similar in the three patient groups (P.0.05), (Fig. 2). represents induced or enhanced expression in astrocytes Although those cases with gemistocytic astrocytes had that, in controls, had little or no clusterin. However, we increased GFAP-positive cytoplasmic area, the high stan- can not exclude the possibility that the AIDS-related dard deviations in AIDS and controls precluded differences increase in CLU-positive cells was due, in part, to induced
between groups. or enhanced expression in oligodendrocytes. The increase
Alterations in GFAP are a common means to assess in relative numbers and percent of all white matter CLU-astrocytic reactivity since its mRNA and protein increase positive cells could not be due to proliferation of these within 1–2 days after an acute injury such as transient cells since MIB-1-immunoreactivy, a cell-proliferation ischemia in rat brain (see [20] for review). Equally rapid marker, was absent in glial cells.
translational increases in astrocyte CLU also occurs [33] The stimulus for, and the consequences of, the increased and provide an independent way of determining reactive CLU expression in AIDS brains are not known.
Upregula-astrocytosis in vivo. tion of astrocyte production and secretion of CLU in vitro
300 J.E. Torres-Munoz et al. / Brain Research 888 (2001) 297 –301
in astrocytes: Implications for AIDS dementia complex, Proc. Natl. Acad. Sci. USA 94 (1994) 494–498.
[2] N. Bruchovsky, R. Snock, P.S. Rennie, K. Akahura, L.S. Golden-berg, M. Geave, Control of tumor progression by maintenance of apoptosis, Prost. Suppl. 6 (1996) 13–21.
[3] M. Calero, T. Tokuda, A. Rostagno et al., Functional and structural properties of lipid-associated apolipoprotein J (clusterin), Biochem. J. 344 (1999) 375–383.
[4] H.W. Cheng, T. Jiang, S.A. Brown, G.M. Pasinetti, C.F. Finch, T.H. McNeil, Response of striatal astrocytes to neuronal differentiation: an immunocytochemical and ultrastructural study, Neuroscience 62 (1994) 425–439.
[5] M. Danik, J.G. Chabot, D. Hassan-Gonzalez, M. Suh, R. Quirion, Localization of sulfated glycoprotein-2 / clusterin mRNA in the rat brain by in situ hybridization, J. Comp. Neurol. 334 (1993) 209– 227.
[6] L.E. French, A. Wohlwend, A.-P. Sappino, J. Tschopp, J.A. Schif-ferli, Human clusterin gene expression is confined to surviving cells during in vitro programmed cell death, J. Clin. Invest. 93 (1994) 877–884.
[7] L.E. French, L.L. Polla, J. Tschopp, J.A. Schifferli, Membrane attack complex (MAC) deposits in skin are not always accompanied by S-protein and clusterin, J. Invest. Dermatol. 98 (1992) 758–763. [8] I.B. Frotz, B.F. Murphy, Clusterin: Insights into a multifunctional
protein, Trends Endocrinol. Metab. 4 (1993) 41–54.
[9] P. Gasque, M. Fontaine, B.P. Morgan, Complement expression in human brain. Biosynthesis of terminal pathway components and regulators in human glial cells and cell lines, J. Immunol. 154 (1995) 4726–4733.
[10] D.E. Jenne, J. Tschopp, Clusterin: the intriguing guises of a widely expressed glycoprotein, Trends Biochem. Sci. 17 (1992) 154–159. [11] S. Krajewski, H.J. James, J. Ross et al., Expression of pro- and anti-apoptosis gene products in brains from pediatric patients with HIV-1 encephalitis, Neuropathol. Appl. Neurobiol. 23 (1997) 242– 253.
[12] L. Kirszbaum, J.A. Sharpe, B. Murphy et al., Molecular cloning and characterization of the novel, human complement-associates protein, SP-40,40: a link between the complement and reproductive systems, EMBO J. 8 (1989) 711–718.
Fig. 2. Relative cytoplasmic area (A), relative numbers of immuno- [13] C. Koch-Brandt, C. Morgans, Clusterin: a role in cell survival in the reactive cells (B) and % total of all glial cells for clusterin immuno- face of apoptosis?, Prog. Mol. Subcell. Biol. 16 (1996) 130–149. reactive glia and GFAP-immunoreactive astrocytes in controls (white [14] N.J. Laping, T.E. Morgan, R.N. Nichols et al., Transforming growth bars) AIDS without HIV encephalitis (HIVnE) (stippled bars) and AIDS factor b1 induces neuronal and astrocyte genes: tubulin A1, glial with HIV encephalitis (HIVE) (black bars). Relative cytoplasmic area and fibrillary acidic protein, and clusterin, Neuroscience 58 (1994)
2 – 1
cell numbers are expressed per mm310 for CLU-positive glia and per 563–572.
2
mm for GFP-positive astrocytes. *: P#0.05 when compared with [15] G. Levi, M. Patrizio, A. Bernardo, T.C. Petrucci, C. Agresti, Human controls. immunodeficiency virus coat protein gp120 inhibits theb-adrenergic
regulation of astroglial and microglial functions, Proc. Natl. Acad. Sci. USA 90 (1993) 1541–1545.
[16] L. Liu, J.K. Persson, M. Svensson, H. Aldskogius, Glial cell responses, complement, and clusterin in the central nervous system Acknowledgements
following dorsal root transection, Glia 23 (1998) 221–238. [17] L. Liu, M. Svensson, H. Aldskogius, Clusterin upregulation
follow-The National Institutes of Health (RO1 NS27416 and ing rubrospinal tract lesion in the adult rat, Exp. Neurol. 157 (1999)
R01 NS39177, CKP) supported this work. The anti-clus- 69–76.
[18] P.C. May, C.E. Finch, Sulfated glycoprotein 2: new relationships of
terin antibody was a generous gift of Dr. B.F. Murphy,
this multifunctional protein to neurodegeneration, Trends Neurosci.
Department of Medicine, St. Vincent’s Hospital, University
15 (1992) 391–396.
of Melbourne, Parkville, Australia. Dr. M.D. Norenberg [19] D. Michel, G. Chatelain, S. North, G. Brun, Stress-induced tran-kindly reviewed the manuscript. scription of the clusterin / apoJ gene, Biochem. 328 (1997) 45–50.
[20] M.D. Norenberg, Astrocytes responses to CNS injury: a review, J. Neuropath. Exp. Neurol. 53 (1994) 213–220.
[21] M.K. O’Bryan, S.S. Cheema, P.F. Bartlett, B.F. Murphy, M.J.
References Pearse, Clusterin levels increase during neuronal development, J.
Neurobiol. 24 (1993) 421–432.
J.E. Torres-Munoz et al. / Brain Research 888 (2001) 297 –301 301
expression in astrocytes and neurons of the adult rat brain, J. Comp. [29] C. Tornatore, R. Chandra, J.R. Berger, E.O. Major, HIV-1 infection Neurol. 339 (1994) 387–400. of subcortical astrocytes in the pediatric central nervous system, [23] L. Pulliam, D. West, N. Haigwood, R.A. Swanson, HIV-1 envelope Neurology 44 (1994) 481–487.
gp120 alters astrocytes in human brain cultures, AIDS Res. Hum. [30] S.M. Wahl, J.B. Allen, N. McCartney-Francis et al., Macrophage and Retroviruses 9 (1993) 439–444. astrocyte-derived transforming growth factor b as a mediator of
˜
[24] M. Redondo, E. Villa, J. Torres-Munoz, T. Tellez, M. Masell, C.K. central nervous system dysfunction in acquired immune deficiency Petito, Overexpression of clusterin in human breast carcinoma, Am. syndrome, J. Exp. Med. 173 (1991) 981–991.
J. Pathol. 157 (2000) 393–399. [31] M.R. Wilson, S.B. Easterbrook-Smith, Clusterin is a secreted [25] Y. Saito, L. Sharer, L. Epstein et al., Overexpression of nef as mammalian chaperone, Trends Biochem. Sci. 25 (2000) 95–98.
marker for restricted HIV-1 infection of astrocytes in postmortem [32] E. Wu, C.F. Brosnan, C.S. Raine, SP-40,40 immunoreactivity in pediatric central nervous system, Neurology 44 (1994) 474–480. inflammatory CNS lesions displaying astrocyte / oligodendrocyte [26] L.R. Sharer, Pathology of HIV-1 infection of the central nervous interactions, J. Neuropathol. Exp. Neurol. 52 (1993) 129–134.
system: a review, J. Neuropathol. Exp. Neurol. 51 (1992) 3–11. [33] M. Zoli, F. Ferraguti, I. Zini, S. Bettuzzi, L.F. Agnati, Increases in ˜
[27] J.E. Torres-Munoz, M. Redondo, N. Tacoronte, C.K. Petito, Clus- sulphates glycoprotein-2 mRNA levels in the rat brain after transient terin expression in astrocytes of human AIDS brains, Neuroscience forebrain ischemia or partial mesodiencephalic hemitransection,
25 (1999) 44. Brain Res. Mol. Brain Res. 18 (1993) 163–177.