www.elsevier.com / locate / bres
Research report
Effects of CART peptides on food consumption, feeding and
associated behaviors in the goldfish, Carassius auratus: actions on
neuropeptide Y- and orexin A-induced feeding
*
´ `
Helene Volkoff, Richard Ector Peter
Department of Biological Sciences, University of Alberta, Edmonton, Alb., Canada T6G 2E9
Accepted 19 September 2000
Abstract
CART (cocaine- and amphetamine-regulated transcript) peptides are novel brain neuropeptides that have been shown to have a role in the control of feeding behavior in mammals. The effects of intracereboventricular (i.c.v.) administration of two CART fragments, CART (62–76) and CART (55–102) on feeding and behavioral activity of goldfish (Carassius auratus) were examined. Both CART peptide fragments inhibited food intake in goldfish. Co-injection of CART peptide and neuropeptide Y (NPY) reduced the increase in feeding caused by injection of NPY alone. CART (55–102) was more potent than CART (62–76). Co-treatment with CART (55–102) and orexin A inhibited the feeding response induced by orexin A alone. Feeding behavior was stimulated relative to saline controls following injection of CART (62–76) alone and co-injection of NPY and CART (62–76), but was not affected by treatment with CART (55–102) alone, NPY and CART (55–102) or orexin A and CART (55–102). Total behavioral activity was increased with NPY, orexin A, and both CART fragments alone as compared to saline controls, as well as in fish co-treated with NPY and CART (62–76) or NPY and CART (55–102) as compared to saline controls and NPY-treated fish. Tremors were seen in fish treated with CART (55–102) alone, and in fish co-treated with NPY and CART (55–102) and orexin A and CART (55–102). Co-treatment of the fish with NPY but not with orexin A significantly lowered the frequency of fish showing tremors as compared to fish treated with CART alone. These results indicate that CART peptides are involved in the regulatory pathways of feeding and behavioral activity in goldfish. 2000 Elsevier Science B.V. All rights reserved.
Theme: Neural basis of behavior
Topic: Neuropeptides and behavior
Keywords: CART peptide; Neuropeptide Y; Orexin A; Feeding behavior; Behavioral activity; Goldfish
1. Introduction adrenals [5,7,8,15,24,27,28,39]. The mature CART peptide contains either 102 (long form) or 89 (short form) amino CART (cocaine- and amphetamine-regulated transcript) acid residues and has several potential cleavage sites [14]. peptides are recently characterized neuropeptides that have The processing of CART peptides is tissue-specific been implicated in the control of appetite in mammals. [31,44]. To date, at least six CART peptides have been CART was initially isolated using PCR differential display identified in mammals [1,31].
as mRNA produced primarily in the rat striatum and In mammals, CART is expressed in hypothalamic areas transcriptionally up-regulated by acute administration of implicated in the control of feeding behavior, such as the psychomotor stimulants such as cocaine and amphetamine arcuate and paraventricular nucleus [2,3,9,14,16,20,28] and [13]. CART was later found to be present throughout its mRNA expression is lowered in fasting conditions mammalian brain and in pituitary, gut, pancreas, and [2,29]. In addition, the gastrointestinal tract contains CART immunoreactivity [8]. In rodents, intracerebroven-tricular injections of CART fragments have been shown to *Corresponding author. Tel.: 11-780-492-5685; fax: 1
1-780-492-inhibit both normal and starvation-induced feeding in a 7033.
E-mail address: [email protected] (R.E. Peter). dose-dependent manner [3,29,32,47]. CART (55–102), a
naturally occurring fragment isolated from ovine hypo- pellets (Moore Clark, St. Andrews, NB, Canada). Fish thalamus [40] appears to be the most potent fragment. The were acclimated under these standard conditions for ap-numbers in the fragments derive from the predicted signal proximately 2 weeks before the start of an experiment. peptide cleavage site in the long form of CART [14]. Other Forty-eight hours prior to experimentation, two fish were CART fragments including CART (54–105), CART (60– moved into an observation tank and starved.
105), CART (61–105) [43], CART (55–76) and CART (62–76) [32] have also been shown to cause a
dose-2.2. Intracerebroventricular (i.c.v.) injections dependent feeding inhibition in rats. Immunoneutralization
of CART by administration of rabbit anti-CART (55–102)
Brain i.c.v. injections were administered following
results in higher food intake in rats, suggesting that CART .
procedures described by Peter and Gill [37] Briefly, is an endogenous satiety factor [32,42].
following deep anesthesia, a three-sided flap was cut in the NPY-containing terminals are present around CART
roof of the skull using a dentist drill equipped with a peptide-positive cell bodies in the paraventricular nucleus
circular saw. The flap was then folded to the side, exposing of the hypothalamus [3,9,32] suggesting a regulation of
the brain. Fish were then placed in a specially designed NPY on CART output. Decreased levels of the adipocyte
stereotaxic apparatus. The needle of a 5-ml microsyringe hormone leptin, as seen during fasting or in obese rats with
was stereotaxically placed in the preoptic region of the impaired leptin action, cause a fall in the expression of
brain third ventricle according to coordinates (11.0, M, D CART mRNA [2,48] which can be restored by peripheral
1.2) taken from the stereotaxic atlas of the goldfish brain administration of leptin [29]. Leptin treatment activates
[37]. Following injection of 2 ml of test solution, the CART hypothalamic neurons [16]. In turn,
intracereb-needle was withdrawn and the space in the cranial cavity roventricular injection of recombinant CART inhibits
filled with teleost physiological saline [4]. The skull flap NPY-induced feeding [32].
was put back in place, and secured by surgical thread. Fish In fish, there is limited information on the neural control
were then returned to their tanks, and normally recovered of feeding behavior. Recent studies show that orexigenic
from anesthesia within 2–5 min. All experiments were peptides such as NPY [33,35], galanin [10], and orexins
carried out in accordance with the principles published in [46], and anorexigenic peptides such as
corticotropin-re-the Canadian Council on Animal Care’s ‘‘guide to corticotropin-re-the care leasing factor (Bernier, Andrusky, Volkoff and Peter,
and use of experimental animals’’. unpublished; [11]), cholecystokinin [20], serotonin [12]
and bombesin [19] are involved in the regulation of
feeding in goldfish. 2.3. Observational experiments
To determine if CART peptides are involved in feeding
in goldfish, we tested the effects of different doses of two Fish were tested in random order in terms of treatment recombinant CART fragments, CART (62–76) and CART and days. For each experiment, two fish were placed in a (55–102), on the feeding behavior of food-restricted fish single observation tank to avoid stress due to isolation and and in fish subjected to NPY- or orexin A (OXA)-induced to allow for an accurate observation of feeding behavior feeding. As CART peptides might have various physiolog- and food consumption. Observations were made for each ical functions and influence other behaviors besides feed- individual fish. An approximate 4% bw ration of pellets ing, we also examined the effects of CART fragments on per fish was administered at 15 min post-injection.
Be-behavioral activity. havioral observations and measurement of food
by counting the times fish mouthed, picked up and spat gravel, or ‘bumped’ any object in the tank (air stone, wall) or their tank mate. A tremor was defined as any rapid change in direction or rapid shaking of the head or the body.
2.4. Reagents
Human CART (62–76) (Tyr–Gly–Val–Pro–Met–Cys– Asp–Ala–Gly–Glu–Gln–Cys–Ala–Val), human CART (55–102) (Val–Pro–Ile–Tyr–Glu–Lys–Lys–Tyr–Gly– Gln –Val – Pro – Met – Cys – Asp – Ala–Gly–Glu–Gly–Cys– Ala – Val – Arg – Lys – Gly–Ala–Arg–Ile–Gly – Lys–Leu – Cys–Asp – Cys – Pro – Arg – Gly–Thr–Ser–Cys–Asn–Ser – Phe–Leu–Leu–Lys–Cys–Leu) and human orexin A (Glp–Pro–Leu–Pro – Asp–Cys–Cys–Arg–Gln–Lys–Thr – Cys – Ser – Cys – Arg–Leu – Tyr–Glu–Leu–Leu–His–Gly – Ala – Gly – Asn – His – Ala – Ala – Gly – Ile–Leu–Thr–Leu – NH2) were purchased from American Peptide Company (Sunnyvale, CA). The numbers in CART correspond to the number in the long form of the pro-CART protein with 102 amino acids. Throughout this manuscript, unless specified otherwise, the term ‘CART’ refers to peptides. Goldfish NPY was a gift from Dr. J.E. Rivier (Clayton Foundation Laboratories for Peptide Biology, The Salk Institute for Biological Studies, La Jolla, CA, USA). Stock
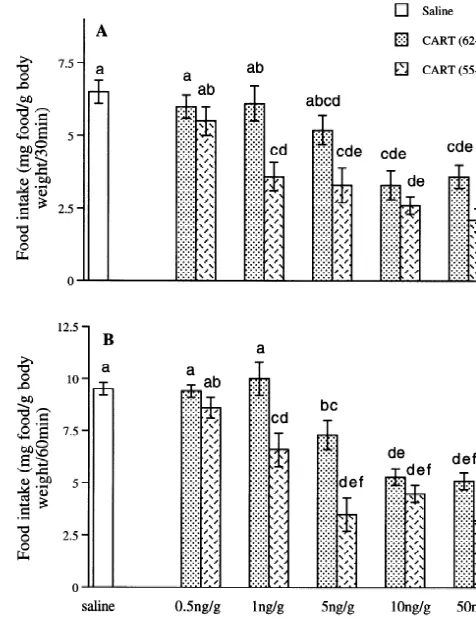
solutions of peptides were made and subsequently diluted Fig. 1. Effects of i.c.v. injection of recombinant CART peptides on food in fish physiological saline [4]. intake of food-restricted goldfish 30 (A) and 60 min (B) following presentation of food. Fish were i.c.v. injected with either saline (n520), CART (62–76) (0.5 ng / g, n58; 1 ng / g, n59; 5 ng / g, n59; 10 ng / g, n512; 50 ng / g, n58), or CART (55–102) (0.5 ng / g, n510; 1 ng / g, 2.5. Statistics n59; 5 ng / g, n510; 10 ng / g, n59; 50 ng / g, n511). Fish received food 15 min post-i.c.v. injection, and were observed for 1 h. Data are Statistical analyses for food intake, number of acts and mean6S.E.M. Bars with dissimilar superscripts indicate groups that differ
significantly. number of tremors were conducted using ANOVA
fol-lowed by Student–Newman–Keuls range test. Significance was considered at P,0.05. All error bars indicate standard
2
error of means (S.E.M.). Two tailedx and Fisher’s exact
test were used for comparison of the frequencies of after 30 min. Doses of 10 and 50 ng / g induced a tremors between saline controls and treated fish. Data significant decrease in food intake at 30 and 60 min. At 60 analyses were performed with Instat 2.0 software for min, the feeding level of fish treated with 10 and 50 ng / g
MacIntosh. CART (62–76) was significantly lower than in fish treated
with a dose of 5 ng / g.
At 0.5 ng / g, CART (55–102) had no significant effects on food consumption compared to the saline group. CART 3. Results (55–102) significantly decreased food consumption at all other doses tested, during all time intervals, compared to 3.1. Effects of human CART (62 –76) and CART (55 – saline-injected controls. At 30 and 60 min after the 102) i.c.v. injections on food intake of food-restricted presentation of food, there was a significant difference goldfish between the number of pellets consumed by 1 and 50 ng / g
treated fish.
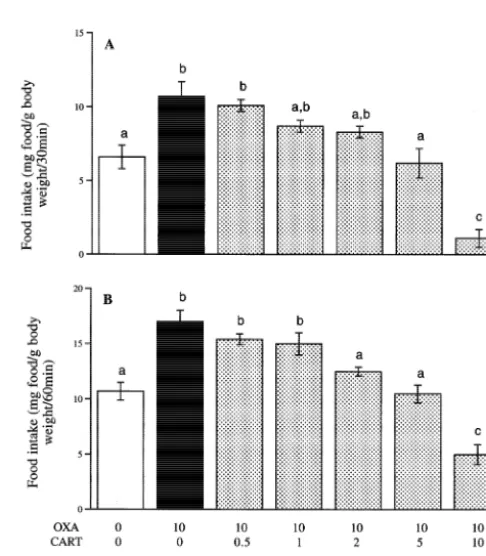
3.2. Effects of human CART (62 –76) and CART (55 – ng / g CART (55–102) caused a greater reduction in food 102) i.c.v. injections on food intake of NPY-treated intake than co-treatment with NPY and CART (62–76) at goldfish the same dosages. At 60 min, co-treatment with 5 ng / g NPY and 2.5 ng / g CART (55–102) caused a greater Fish treated with 5 ng / g NPY exhibited a higher food reduction in food intake than co-treatment with NPY and intake compared to the saline group at both 30 and 60 min CART (62–76) at the same dosages. There was no
(Fig. 2). difference in food intake between the two fragments at any
At 30 min, NPY-induced feeding was significantly other dose at either time interval. reduced by co-treatment with CART (62–76) at doses of
2.5, 5 and 10 ng / g. At 60 min, NPY-induced feeding was
reduced by co-treatment with CART (62–76) at all doses 3.3. Effects of human CART (55 –102) i.c.v. injections tested. Fish co-treated with 5 ng / g NPY and CART (62– on food intake of orexin A-treated goldfish
76) at doses of 5 and 10 ng / g had levels of feeding similar
to the saline group at 30 and 60 min. Fish treated with 10 ng / g OXA alone exhibited a higher NPY-induced feeding was reduced by CART (55–102) food intake compared to the saline group at both 30 and 60 at all doses tested, at both 30 and 60 min. All groups of min (Fig. 3).
fish co-treated with NPY and CART (55–102) had levels At 30 min, orexin A-induced feeding was reduced by of feeding similar to the saline group. There were no co-treatment with CART (55–102) at doses of 5 and 10 significant differences between the five co-treatment ng / g. At 60 min, OXA-induced feeding was reduced by groups at either 30 or 60 min. co-treatments with of CART (55–102) at doses of 2, 5 At 30 and 60 min, co-treatment with 5 ng / g NPY and and10 ng / g. At 30 and 60 min, only fish co-treated with 10 0.5 ng / g CART (55–102), and with 5 ng / g NPY and 1 ng / g OXA and 10 ng / g CART (55–102) exhibited a food
intake significantly lower than the saline controls.
Fig. 2. Effects of i.c.v. injection of recombinant CART peptides on food intake of NPY-treated goldfish 30 (A) and 60 min (B) following
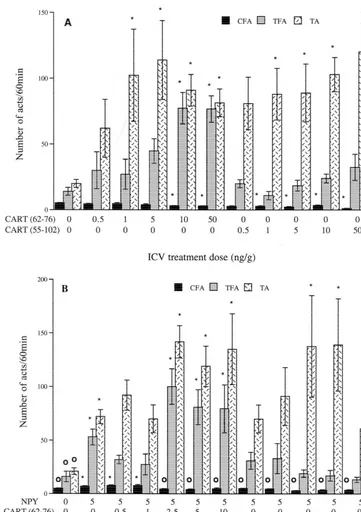
3.4. Effects of human CART (62 –76) and CART (55 – feeding acts was significantly reduced by treatment with 102) i.c.v. injections on feeding behavior and behavioral CART (62–76) at 10 and 50 ng / g and with CART activity of goldfish (55–102) at 1, 5, 10 and 50 ng / g (Fig. 4A). Feeding behavior, as indicated by the total number of feeding acts In food-restricted goldfish, the number of complete in 60 min, was increased with CART (62–76) at doses of
Fig. 4. (continued )
10 and 50 ng / g. CART (55–102) had no significant effects significant tremor activity. Co-treatment with 10 ng / g on feeding behavior. The total number of acts was OXA and CART (55–102) at doses of 5 and 10 ng / g significantly increased with both CART fragments at 1, 5, induced a significant increase in the number of fish 10 and 50 ng / g as compared to saline controls.
In NPY-treated goldfish, the number of complete feed-ing acts was significantly increased only in fish treated with 5 ng / g NPY and in fish co-treated with 5 ng / g NPY and 0.5 or 1 ng / g CART (62–76) as compared to saline controls (Fig. 4B). The total number of feeding acts was increased in fish treated with NPY alone and in fish co-treated with 5 ng / g NPY and CART (62–76) at doses of 2.5, 5 and 10 ng / g relative to saline controls, but was not significantly different from saline controls in any group treated with CART (55–102). The total number of acts was increased in fish treated with NPY alone, in fish co-treated with 5 ng / g NPY and CART (62–76) at doses of 2.5, 5 and 10 ng / g, and in fish co-treated with NPY 5 ng / g and CART (55–102) at doses of 2.5 and 5 ng / g, as compared to the saline controls.
In OXA-treated goldfish, the number of complete feed-ing acts was higher in fish treated with OXA alone and in fish co-treated with 10 ng / g OXA and 0.5 or 1 ng / g CART (55–102) as compared to the saline controls (Fig. 4C). The total number of feeding acts and the total number of acts were both increased in fish treated with OXA alone relative to saline controls and were significantly lower in fish co-treated with 10 ng / g OXA and CART (55–102) at
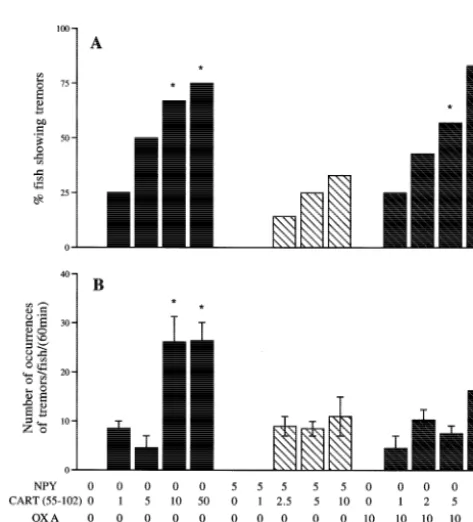
all doses tested, as compared to OXA alone. Fig. 5. Effects of i.c.v. injection of CART(55–102) on the occurrence of Tremors were induced by CART (55–102) alone at tremors in food-restricted, NPY-treated and orexin A-treated goldfish. The data is presented as the frequency of fish displaying tremors in each doses of 1 ng / g and higher, but this induction was only
treatment group (A, mean6S.E.M.) and the number of occurrences of significant at doses of 10 and 50 ng / g as compared to the
showing tremors as compared to saline controls. There was of feeding, respectively, as compared to the saline group. no significant difference in the number of occurrences of The stimulatory effects of i.c.v. injected NPY were sup-tremors per fish when co-treated with NPY and CART pressed by CART in a dose-dependent manner, with (55–102) or OXA and CART (55–102) compared to CART (55–102) being more potent. In rats, simultaneous saline controls or to fish treated with 10 ng / g CART i.c.v. injection of 4 nmol NPY and 2.4 nmol CART (55–102). Tremors were not observed in fish treated with (55–76) attenuates feeding [32] and co-injection of 5 mg CART (62–76) alone or in combination with NPY or OXA NPY and CART (55–102) at doses of 0.5, 1 and 2mg also
(data not shown). causes a dose-dependent inhibition of NPY-stimulated
feeding [29,47]. When administered alone at moderate doses (0.5 ng / g), neither fragment affected food intake, 4. Discussion suggesting that the inhibition of NPY-induced food intake was not due to a pharmacological non-specific effect. This Intracerebroventricular injection of either human CART further suggests that the inhibition of NPY-induced feeding (62–76) or CART (55–102) into the brain third ventricle by CART peptides does not result merely from additive of goldfish decreased food consumption in food restricted effects of two peptides with opposite effects, but might be fish, as well as in NPY- and OXA-treated fish with evidence of an interaction between NPY and CART increased food intake. The decrease in food intake was not systems. Such interactions, both anatomical and functional, due to stress, since injected fish were responsive to food, have been demonstrated in mammals [29,30,32,47]. but ingested lesser amounts of pellets. Fish were also very As previously reported [46], we found orexin A to be a active and displayed feeding and searching behavior. potent orexigenic factor in goldfish. In this study, we show To date, the effects of CART fragments have only been that CART (55–102) dose-dependently suppresses the tested on rodents. In rats, several CART fragments have orexigenic effects of orexin A. To our knowledge, this is been reported to inhibit food consumption in a dose- the first report on the actions of CART peptides on orexin-dependent fashion when injected i.c.v. [29,32,42,47]. induced feeding behavior. A pronounced inhibition of Among all fragments, the full length CART (55–102) orexin-induced feeding behavior occurs at doses of CART appears to be the most potent. Kristensen et al. [29] (55–102) that do not affect normal feeding behavior, or reported a 60–70% decrease in food intake 1 h after normal and orexin-induced food intake, suggesting the injection of 1–2 mg CART (55–102), similar to the existence of interactions between orexin and NPY systems. present findings obtained in goldfish. The effects of CART Whereas synaptic appositions have been demonstrated (55–102) are rapid, as a significant decrease in food intake between CART and NPY fibers, and between orexin and was observed as soon as 15 min post-injection (data not NPY cells in mammals, no neuroanatomical link between shown). Similar results were reported in mice, in which CART and orexins has been described to date [3]. Perhaps i.c.v. injection of 2–4 mg of CART (55–102) caused a the effects of CART on orexin-induced feeding are me-decrease in feeding 10 min post-injection [43]. CART diated by NPY systems. This hypothesis is strengthened by (62–76) also elicited a decrease in food consumption of recent reports that Y1 receptors are involved in the goldfish but at higher i.c.v. dosages than CART (55–102). stimulation of feeding by orexins in rats [23,49] as well as At higher doses, CART (62–86) caused a similar inhibi- in goldfish (H. Volkoff and R.E. Peter, unpublished data). tion of feeding as that caused by CART (55–102). CART Further studies are needed to clarify if CART acts directly (62–76) has also been shown to cause a similar inhibition on orexin neurons or via NPY systems.
of feeding in rats [32]. When compared to other anorex- Feeding behavior was enhanced in fish co-treated with igenic agents in goldfish, CART fragments are similarly NPY and CART (62–76) as compared to saline controls, potent to bombesin, which causes a 80% decrease in 30 but was not affected by co-treatments with NPY and min food intake at 60 ng / g [19], and cholecystokinin, CART (55–102) or OXA and CART (55–102). The fact which causes a 95% decrease in 30 min food intake at 50 that CART (55–102) inhibits both NPY- and
OXA-in-ng / g [20]. duced feeding behavior and that CART (62–76) has no
To date, only two CART fragments, CART (55–76) and effect on NPY-induced feeding is further evidence of the CART (55–102), have been studied for their interaction higher potency of the long fragment.
effects of CART on locomotor behavior [26] and anxiety- peptides act as potent satiety factors and stimulate be-like reactions [25] have previously been reported in havioral activity in goldfish. Our results demonstrate for mammals. Associated with i.c.v. injection of high doses of the first time that CART peptides are involved in the CART (62–76) was an increase in incomplete feeding regulation of feeding in a non-mammalian vertebrate, and acts. Interestingly, injections of CART (55–102) did not that they affect NPY- and orexin A-elicited behaviors. The induce an increase in feeding behavior specifically. Total stimulation of behavioral activity as well as the induction behavioral activity was enhanced in fish treated with NPY of tremors by CART fragments indicates that there are and greatly increased in orexin A-treated fish as compared multiple functions of these peptides in vertebrates, not to saline controls. In mammals, NPY mostly induces a limited to the regulation of food intake. The results of the suppression of behavioral activity. However, NPY has also present study provide indirect evidence for the existence of been reported to enhance searching behavior in rats [22]. CART in lower vertebrates, and suggests an early The increase in overall activity in NPY-treated goldfish is evolutionary origin for this peptide transmitter and its mainly the result of an increased feeding behavior as the behavioral actions in the nervous system of vertebrates. fish displayed relatively few non-feeding acts. In orexin
A-treated fish, non-feeding acts largely contribute to the increased behavioral activity. Orexin A increases arousal
Acknowledgements and locomotor activity in rats [17,22,34] and disruption of
the prepro-orexin gene induces a behavioral arrest in mice
This work was supported by grant A6371 from the [6]. The increase in activity was much higher in the case of
Natural Sciences and Engineering Research Council orexin-treated fish (averaging 300) than in the case of
(NSERC) of Canada to R.E.P. CART (55–102) or NPY (averaging 100). Fish co-treated
with NPY and CART (62–76) and with NPY and CART (55–102) were equally or more active than NPY-treated
References fish. Fish co-treated with OXA and CART (55–102) were
less active than OXA-treated fish, but as active as fish
[1] L.D. Adams, W. Gong, S. Dall Vechia, R.G. Hunter, M.J. Kuhar, treated with CART alone. The increase in activity in
CART: from gene to function, Brain Res. 848 (1999) 137–140. OXA-treated fish was due mainly to an increase in the
[2] R.S. Ahima, J. Kelly, J.K. Elmquist, J.S. Flier, Distinct physiologic number of feeding acts, whereas the increase in activity in
and neuronal responses to decreased leptin and mild hyperlep-CART-treated fish was the result of numerous non-feeding tinemia, Endocrinology 140 (1999) 4923–4931.
acts. As CART (55–102) given alone has no significant [3] C. Broberger, Hypothalamic cocaine- and amphetamine-regulated transcript (CART) neurons: histochemical relationship to thyro-effect on the number of feeding acts, it is probable that
tropin-releasing hormone, melanin-concentrating hormone, orexin / CART (55–102) attenuates OXA-induced increase in
hypocretin and neuropeptide Y, Brain Res. 848 (1999) 101–113. activity mainly by inhibiting OXA-induced feeding
be-[4] G. Burnstock, Saline for fresh-water fish, J. Physiol. 141 (1958)
havior. 35–45.
Most fish treated with CART (55–102) displayed ‘tre- [5] Y. Charnay, C. Perrin, P.G. Vallet, B. Greggio, E. Kovari, C. Bouras, mors’. Similar behavioral effects were not seen in saline-, Mapping of cocaine and amphetamine regulated transcript (CART) mRNA expression in the hypothalamus of elderly human, J. Chem. NPY-, OXA- or CART (62–76)-treated fish. The tremors
Neuroanat. 17 (1999) 123–128. were intermittent and the affected fish were clearly able to
[6] R.M. Chemelli, J.T. Willie, C.M. Sinton, J.K. Elmquist, T. Scam-swim and eat. Overdosing of central nervous system with mell, C. Lee, J.A. Richardson, S.C. Williams, Y. Xiong, Y. Kisanuki, stimulants such as cocaine and amphetamine-related drugs T.E. Fitch, M. Nakazato, R.E. Hammer, C.B. Saper, M. Yanagisawa, results in a dose-dependent increase in the frequency of Narcolepsy in orexin knockout mice: molecular genetics of sleep
regulation, Cell 98 (1999) 437–451. behavioral seizures [18]. As CART peptide gene
expres-[7] P.R. Couceyro, E.O. Koylu, M.J. Kuhar, Further studies on the sion is up-regulated by treatment by such drugs, it is not
anatomical distribution of CART by in situ hybridization, J. Chem. surprising that high doses of CART induce seizure-like Neuroanat. 12 (1997) 229–241.
behavior. In rats, i.c.v. injection of CART (55–102) at [8] P. Couceyro, M. Paquet, E. Koylu, M.J. Kuhar, Y. Smith, Cocaine-high doses (1–2 mg) induces movement-associated tre- and amphetamine-regulated transcript (CART) peptide immuno-reactivity in myenteric plexus neurons of the rat ileum and co-mors. We observed tremors at all doses of CART (55–102)
localization with choline acetyltransferase, Synapse 30 (1998) 1–8. tested, with a higher number of occurrences at higher
[9] S. Dall Vechia, P.D. Lambert, P.C. Couceyro, M.J. Kuhar, Y. Smith, doses. Co-treatment of the fish with NPY but not with CART peptide immunoreactivity in the hypothalamus and pituitary orexin A significantly attenuated the number of fish in monkeys: analysis of ultrastructural features and synaptic con-showing tremors, as compared to fish treated with CART nections in the paraventricular nucleus, J. Comp. Neurol. 416 (2000)
291–308. alone. These results are in accordance with the fact that
[10] N. De Pedro, M.V. Cespedes, M.J. Delgado, M. Alonso-Bedate, The NPY has been shown to have an anticonvulsant role both
galanin-induced feeding stimulation is mediated via a-2-adrenergic in vivo and in vitro [36,45], and that high doses of orexin receptors in goldfish, Regul. Pept. 57 (1995) 77–84.
41] on feeding in goldfish: involvement of cortisol and catechol- [30] M.J. Kuhar, S.E. Dall Vechia, CART peptides: novel addiction- and amines, Behav. Neurosci. 111 (1997) 398–403. feeding-related neuropeptides, Trends Neurosci. 22 (1999) 316–320. [12] N. De Pedro, M.L. Pinillos, A.I. Valenciano, M. Alonso-Bedate, M.J. [31] M.J. Kuhar, L.L. Yoho, CART peptide analysis by Western blotting,
Delgado, Inhibitory effect of serotonin on feeding behavior in Synapse 33 (1999) 163–171.
goldfish: involvement of CRF, Peptides 19 (1998) 505–511. [32] P.D. Lambert, P.R. Couceyro, K.M. McGirr, S.E. Dall Vechia, Y. [13] J. Douglass, A.A. Mckinzie, P. Couceyro, PCR differential display Smith, M.J. Kuhar, CART peptides in the central control of feeding identifies a rat brain mRNA that is transcriptionally regulated by and interactions with neuropeptide Y, Synapse 29 (1998) 293–298.
˜
cocaine and amphetamine, J. Neurosci. 15 (1995) 2471–2481. [33] M.A. Lopez-Patino, A.I. Guijarro, E. Isorna, M.J. Delgado, M. [14] J. Douglass, S. Daoud, Characterization of the human cDNA and Alonso-Bedate, N. de Pedro, Neuropeptide Y has a stimulatory genomic DNA encoding CART: a cocaine- and amphetamine- action on feeding behavior in goldfish, Eur. J. Pharmacol. 377 regulated transcript, Gene 169 (1996) 241–245. (1999) 147–153.
[15] N.J. Dun, S.L. Dun, E.H. Kwok, J. Yang, J. Chang, Cocaine- and [34] T. Nakamura, K. Uramura, T. Nambu, T. Yada, K. Goto, M. amphetamine-regulated transcript-immunoreactivity in the rat sym- Yanagisawa, T. Sakurai, Orexin-induced hyperlocomotion and patho-adrenal axis, Neurosci. Lett. 283 (2000) 97–100. stereotypy are mediated by the dopaminergic system, Brain Res. 873 [16] C.F. Elias, C. Lee, J. Kelly, C. Aschkenasi, R.S. Ahima, P.R. (2000) 181–187.
Couceyro, M.J. Kuhar, C.B. Saper, J.K. Elmquist, Leptin activates [35] Y.K. Narnaware, P.P. Peyon, X. Lin, R.E. Peter, Regulation of food hypothalamic CART neurons projecting to the spinal cord, Neuron intake ny neuropeptide Y in goldfish, Am. J. Physiol. Integrative
21 (1998) 1375–1385. Comp. Physiol. 279 (2000) R1025–R1034.
[17] J.J. Hagan, R.A. Leslie, S. Patel, M.L. Evans, T.A. Wattam, S. [36] R.D. Palmiter, J.C. Erickson, G. Hollopeter, S.C. Baraban, M.W. Holmes, C.D. Benham, S.G. Taylor, C. Routledge, P. Hemmati, R.P. Schwartz, Life without neuropeptide Y, Recent Prog. Horm. Res. 53 Munton, T.E. Ashmeade, A.S. Shah, J.P. Hatcher, P.D. Hatcher, D.N. (1998) 163–199.
Jones, M.I. Smith, D.C. Piper, A.J. Hunter, R.A. Porter, N. Upton, [37] R.E. Peter, V.E. Gill, A stereotaxic atlas and technique for forebrain Orexin A activates locus coeruleus cell firing and increases arousal nuclei of the goldfish, Carassius auratus, J. Comp. Neurol. 159 in the rat, Proc. Natl. Acad. Sci. USA 96 (1999) 10911–10916. (1975) 69–102.
[18] G.R. Hanson, M. Jensen, M. Johnson, H.S. White, Distinct features [38] O. Rugarn, M. Hammar, A. Theodorsson, E. Theodorsson, C. of seizures induced by cocaine and amphetamine analogs, Eur. J. Stenfors, Sex differences in neuropeptide distribution in the rat Pharmacol. 377 (1999) 167–173. brain, Peptides 20 (1999) 81–86.
[19] B.A. Himick, R.E. Peter, Bombesin acts to suppress feeding [39] Y. Smith, J. Kieval, P.R. Couceyro, M.J. Kuhar, CART peptide-behavior and alter serum growth hormone in goldfish, Physiol. immunoreactive neurones in the nucleus accumbens in monkeys: Behav. 55 (1994) 65–72. ultrastructural analysis, colocalization studies, and synaptic interac-[20] B.A. Himick, R.E. Peter, CCK / gastrin-like immunoreactivity in tions with dopaminergic afferents, J. Comp. Neurol. 407 (1999)
brain and gut, and CCK suppression of feeding in goldfish, Am. J. 491–511.
Physiol. 267 (1995) R841–R851. [40] J. Spiess, J. Villarreal, W. Vale, Isolation and sequence analysis of a [21] Y.L. Hurd, P. Svensson, M. Ponten, The role of dopamine, dynor- somatostatin-like polypeptide from ovine hypothalamus,
Biochemis-phin, and CART systems in the ventral striatum and amygdala in try 20 (1981) 1982–1988.
cocaine abuse, Ann. NY Acad. Sci. 877 (1999) 499–506. [41] S. Taheri, M. Mahmoodi, J. Opacka-Juffry, M.A. Ghatei, S.R. [22] T. Ida, K. Nakahara, T. Katayama, N. Murakami, M. Nakazato, Bloom, Distribution and quantification of immunoreactive orexin A
Effect of lateral cerebroventricular injection of the appetite-stimulat- in rat tissues, FEBS Lett. 457 (1999) 157–161.
ing neuropeptide, orexin and neuropeptide Y, on the various [42] L. Thim, P. Kristensen, P.J. Larsen, B.S. Wulff, CART, a new behavioral activities of rats, Brain Res. 821 (1999) 526–529. anorectic peptide, Int. J. Biochem. Cell Biol. 30 (1998) 1281–1284. [23] M.R. Jain, T.L. Horvath, P.S. Kalra, S.P. Kalra, Evidence that NPY [43] L. Thim, P.F. Nielsen, M.E. Judge, A.S. Andersen, I. Diers, M. Y1 receptors are involved in stimulation of feeding by orexins Egel-Mitani, S. Hastrup, Purification and characterisation of a new (hypocretins) in sated rats, Regul. Pept. 87 (2000) 19–24. hypothalamic satiety peptide, cocaine and amphetamine regulated [24] P.B. Jensen, P. Kristensen, J.T. Clausen, M.E. Judge, S. Hastrup, L. transcript (CART), produced in yeast, FEBS Lett. 428 (1998)
Thim, B.S. Wulff, C. Foged, J. Jensen, J.J. Holst, O.D. Madsen, The 263–268.
hypothalamic satiety peptide CART is expressed in anorectic and [44] L. Thim, P. Kristensen, P.F. Nielsen, B.S. Wulff, J.T. Clausen, non-anorectic pancreatic islet tumors and in the normal islet of Tissue-specific processing of cocaine- and amphetamine-regulated Langerhans, FEBS Lett. 447 (1999) 139–143. transcript peptides in the rat, Proc. Natl. Acad. Sci. USA 96 (1999) [25] A. Kask, H.B. Schioth, F. Mutulis, J.E. Wikberg, L. Rago, Anorex- 2722–2727.
igenic cocaine- and amphetamine-regulated transcript peptide inten- [45] A. Vezzani, G. Sperk, W.F. Colmers, Neuropeptide Y: emerging sifies fear reactions in rats, Brain Res. 857 (2000) 283–285. evidence for a functional role in seizure modulation, Trends [26] H.L. Kimmel, W. Gong, S.D. Vechia, R.G. Hunter, M.J. Kuhar, Neurosci. 22 (1999) 25–30.
Intra-ventral tegmental area injection of rat cocaine and amphet- [46] H. Volkoff, J.M. Bjorklund, R.E. Peter, Stimulation of feeding amine-regulated transcript peptide 55–102 induces locomotor activi- behavior and food consumption in the goldfish, Carassius auratus, ty and promotes conditioned place preference, Pharmacol. Exp. by orexin-A and orexin-B, Brain Res. 846 (1999) 204–209. Ther. 294 (2000) 784–792. [47] N. Vrang, M. Tang-Christensen, P.J. Larsen, P. Kristensen, Recombi-[27] E.O. Koylu, P.R. Couceyro, P.D. Lambert, M.J. Kuhar, Cocaine- and nant CART peptide induces c-Fos expression in central areas amphetamine-regulated transcript peptide immunohistochemical involved in control of feeding behaviour, Brain Res. 818 (1999) localization in the rat brain, J. Comp. Neurol. 391 (1998) 115–132. 499–509.
[28] E.O. Koylu, P.R. Couceyro, P.D. Lambert, N.C. Ling, E.B. DeSouza, [48] Z.W. Wang, Y.T. Zhou, T. Kakuma, Y. Lee, M. Higa, S.P. Kalra, M.J. Kuhar, Immunohistochemical localization of novel CART M.G. Dube, P.S. Kalra, R.H. Unger, Comparing the hypothalamic peptides in rat hypothalamus, pituitary and adrenal gland, J. and extrahypothalamic actions of endogenous hyperleptinemia, Proc. Neuroendocrinol. 9 (1997) 823–833. Natl. Acad. Sci. USA 96 (1999) 10373–10378.