SKRIPSI
Oleh
RENA TRI HERNAWATI
B1J008095
KEMENTERIAN PENDIDIKAN DAN KEBUDAYAAN
UNIVERSITAS JENDERAL SOEDIRMAN
FAKULTAS BIOLOGI
PURWOKERTO
Oleh
RENA TRI HERNAWATI
B1J008095
Untuk Memenuhi Persyaratan Memperoleh Gelar Sarjana Sain pada Fakultas Biologi
Universitas Jenderal Soedirman Purwokerto
Disetujui dan disahkan Pada tanggal...
Pembimbing I, Pembimbing II,
Dr. Agus Nuryanto, S.Si., M.Si. Drs. Indarmawan, M.S. NIP. 19690825 199702 1 001 NIP. 19531028197903 1 002
Mengetahui: Dekan Fakultas Biologi Universitas Jenderal Soedirman,
Dra. Purnomowati, SU. NIP. 19531021 198103 2 001
iii
SWT yang Maha Kasih dan Maha Pemberi Petunjuk, atas segala berkah dan karunia yang diberikan sehingga penulis dapat menyelesaikan penelitian dan menyusun skripsi tentang Hubungan Kekerabatan Crustacea (Decapoda) di Sungai Cijalu Kecamatan Majenang Kabupaten Cilacap.
Sistematika dan gaya penulisan skripsi ini telah diusahakan memenuhi ketentuan-ketentuan di dalam Pedoman Penulisan Usulan Penelitian dan Skripsi Fakultas Biologi Universitas Jenderal Soedirman hasil revisi tahun 2004.
Penulis mengucapkan terima kasih kepada semua pihak yang telah membantu selama proses penyusunan skripsi ini.
1. Dra. Purnomowati, SU. selaku Dekan Fakultas Biologi yang telah memberikan izin untuk melaksanakan penelitian.
2. Dr. Agus Nuryanto, S.Si., M.Si. selaku Pembimbing I yang telah memberikan bimbingan dan arahan dalam penyusunan laporan penelitian. 3. Drs. Indarmawan, M.Si. selaku Pembimbing II yang telah memberikan
bimbingan dan arahan dalam penyusunan laporan penelitian ini.
4. Drs. M. Nadjmi Abulias, M.Si. dan Dra. A. E. Pulungsari, M.Si. sebagai penelaah yang telah memeberikan kritik dan saran.
Akhirnya penulis berharap semoga skripsi ini dapat memberikan sumbangsih kepada khasanah ilmu pengetahuan, khususnya di bidang kajian Taksonomi Hewan. Amin.
Purwokerto, Februari 2013
iv
Halaman
PRAKATA... ... iii
DAFTAR TABEL... v
DAFTAR GAMBAR... vi
DAFTAR LAMPIRAN... vii
INTISARI... viii
ABSTRACT ... ix
I. PENDAHULUAN... 1
II. MATERI DAN METODE... 8
1. Materi, Lokasi dan Waktu Penelitian... 8
2. Metode Penelitian... 9
3. Metode Analisis... 12
III. HASIL DAN PEMBAHASAN ... 13
IV. KESIMPULAN DAN SARAN ... 42
DAFTAR REFERENSI... 43
LAMPIRAN... 46
v
vi
Halaman
3.1. Morfologi umum udang air tawar genus Macrobrachium... 17
3.2. M. pilimanus tampak lateral... 18
3.3. M. pilimanus tampak dorsal... 19
3.4. Preanal carina pada M. pilimanus... 20
3.5. Morfologi M. sintangense tampak lateral dan dorsal... 21
3.6. Skema pubescence dan chela pada M. sintangense... 22
3.7. Morfologi M. lanchesteri... 24
3.8. Skema bentuk karapas genus Parathelphusa... 28
3.9. Skema frontal genus Parathelphusa... 28
3.10. Skema palpus mandibula genus Parathelphusa... 29
3.11. Skema pleopod pertama genus Parathelphusa... 29
3.12. Skema bentuk abdomen ♂ genus Parathelphusa... 30
3.13. Ketebalan karapas P. bogorensis dan P. convexa dan skema pereiopoda pada P. convexa... 31
3.14. Skema pereiopoda pada P. convexa... 33
3.15. Pohon filogeni dengan konsensus aturan mayoritas bootstrap 50%... 35
3.16. Kladogram dengan panjang langkah panjang langkah 48, indeks konsistensi (CI)= 0,98 dan indeks retensi (RI)= 0,95... 36
vii
1. Peta Lokasi Pengambilan Sampel... 46
2. Foto Stasiun Pengambilan Sampel... 47
3. Data Kualitas Fisika dan Kimia Sungai Cijalu... 53
4. Kunci determinasi... 54
5. Hasil pengukuran morfometri species anggota ordo Decapoda yang tertangkap di Sungai Cijalu Kecamatan Majenang... 59
6. Simplesiomorphy characters dari Satuan Taksonomi Operasional atau OTU (Operational Taxonomy Unit)... 64
viii
hidup di lingkungan perairan. Sebuah penelitian berjudul “Hubungan Kekerabatan Crustacea (Decapoda) di Sungai Cijalu Kecamatan Majenang Kabupaten Cilacap” telah dilakukan pada bulan Juni sampai Nopember 2012. Tujuan dari penelitian ini yaitu untuk mengetahui diversitas dan hubungan kekerabatan species ordo Decapoda yang ditangkap dari Sungai Cijalu. Sampel diambil menggunakan metode survei dengan teknik pengambilan sampel secara acak kelompok atau Cluster Random Sampling dengan tiga kali ulangan. Karakter morfologi yang diamati dari udang dan kepiting berupa rostrum, karapas, pereiopoda, pubescence, preanal carina, karpus, merus, abdomen, telson, dan uropod. Variabel yang diperoleh ditransformasi ke data biner dan dianalisis menggunakan uji parsimoni. Pohon filogenetik direkonstruksi menggunakan software PAUP version 4.b10. dengan outgroup species dari ordo Isopoda (Pseudotyphloscia pallida) yang dijadikan sebagai pembanding. Proses identifikasi, determinasi dan verifikasi mendapatkan lima species anggota Decapoda, tiga species udang dengan nama Macrobrachium sintangense, M. pilimanus, dan M. lanchesteri dan dua species kepiting dengan nama Parathelphusa bogorensis, dan P. convexa. Analisis kladistik menghasilkan kladogram dengan panjang langkah 48, indeks konsistensi (CI)= 0,98 dan indeks retensi (RI)= 0,95. Hubungan kekerabatan filogenetik Decapoda menunjukkan bahwa species dari ordo Decapoda bersifat monofiletik dengan tiga cabang pohon. M. sintangense sebagai spesies primitif yang berada di cabang pohon pertama, disusul oleh cabang pohon kedua oleh M. pilimanus dan M. lanchesteri. Terdapat dua spesies kepiting Parathelphusa bogorensis dan P. convexa pada cabang pohon ketiga. Berdasarkan kladogram, P.convexa sebagai spesies yang paling maju (derived species).
Kata Kunci: Decapoda, Sungai Cijalu, kekerabatan filogenetik, diversitas, kladistik.
ix
environment. A study entitled “The Relationships of the Crustaceans (Decapods) at Cijalu River at Majenang Subdistric, Cilacap Regency” have been performed at the periode of June to November 2012”. The aims of the study were to know the species diversity and relationships of the order Decapods collected at Cijalu River. The samples were taken using survey method with Cluster Random Sampling Technique and tree replications. The observed morphological characters of prawns and crabs were rostrum, carapace, pereiopod, pubescence, preanal carina, carpus, merus, abdomen, telson, and uropod. The obtained variables were transformed into binary data and analyzed by using parsimony test. Phylogenetic tree was reconstructed using PAUP software version 4.b10. with outgroup species from the order Isopoda (Pseudotyphloscia pallida) as comparisson. The identification, determination and verification resulted five species of Decapods, consisted of three species of shrimps, namely Macrobrachium sintangense, M. pilimanus, and M. lanchesteri and two species of crabs, that is Parathelphusa bogorensis, and P. convexa. The cladistic analysis produced cladogram with length of 48 steps, consistency index (CI) of 0.98, and retention index (RI) of 0.95. Phylogenetic relationships of Decapods indicated that the species of the order Decapods created a monophyletic group with three branches compared to outgroup species. Cladogram indicated that M. sintangense was as a primitive species on the first branches of the tree, followed by the second branches by M. pilimanus and M. lanchesteri. There were two species of crabs Parathelphusa bogorensis and P. convexa on the third branch of the tree. According the cladogram, P.convexa was the most advanced species (derived species).
Keywords: Decapods, Cijalu River, phylogenetic relationships, diversity, cladistic.
iii
Sungai merupakan salah satu perairan yang mengalir dari hulu ke muara. Di sepanjang aliran sungai terjadi penggabungan oleh beberapa sungai dan parit. Aliran tersebut akan melalui daerah-daerah yang memiliki topografi berbeda, seperti daerah curam, landai, dan ada yang relatif datar. Perbedaan topografi yang dilalui oleh sungai akan menyebabkan terjadinya perbedaan kecepatan arus pada bagian-bagian sungai tersebut. Bagian sungai yang melalui daerah yang datar kecepatan arusnya akan lambat dan relatif tenang (Duya, 2008).
Sungai dan anak-anak sungai tersebut berfungsi untuk menampung, menyimpan dan mengalirkan air yang berasal dari curah hujan serta sumber air lainnya. Penyimpanan dan pengaliran air dihimpun dan ditata berdasarkan hukum alam di sekelilingnya sesuai dengan keseimbangan daerah tersebut (Rahayu et al., 2009).
Sungai Cijalu merupakan anak sungai yang berada di Kecamatan Majenang Kabupaten Cilacap yang bermuara ke DAS Citanduy. Sungai Cijalu yang deras bermata air di Gunung Padontelu atau Gunung Tiga yang berada di Desa Sepatnunggal. Sebagian besar masyarakat desa tersebut memanfaatkan air Sungai Cilaju untuk irigasi dan sumur resapan. Aliran air Sungai Cijalu berdasarkan ordo sungai mencakup Sungai Cijalu, Sungai Cileumeuh, dan Sungai Cikawung yang bermuara ke Sungai Citanduy (Departemen Pekerjaan Umum, 2010).
Bagian hulu memiliki karakteristik dasar sungai berupa batu besar, agak curam, serta di samping kanan kiri masih terdapat banyak pohon. Bagian tengah
2
dari Sungai Cijalu mempunyai struktur berbatu kecil dan pasir yang berada di Desa Mulyadadi. Hilir Sungai Cijalu berada di Desa Pahonjean Kecamatan Majenang yang merupakan salah satu daerah yang memiliki topografi cekung sehingga pada saat musim hujan sangat berpotensi banjir. Aliran air Sungai Cijalu bergabung dengan Sungai Cileumeuh dan Cikawung bermuara di Sungai Citanduy (Departemen Pekerjaan Umum, 2010).
Kondisi hidrologi Pulau Jawa dan Indonesia umumnya dicirikan oleh meningkatnya kejadian ekstrem seperti banjir dan kekeringan dengan kandungan cemaran yang tinggi di badan-badan air seperti sungai dan danau (Pawitan, 2004). Adanya bencana alam seperti banjir seringkali menjadi ancaman bagi keberadaan species yang ada didalamnya. Aktivitas manusia yang terkadang mengeksploitasi sungai secara berlebihan juga menjadi ancaman kehidupan hewan-hewan air seperti ikan dan udang sungai. Disamping itu, metode penangkapan hasil sungai yang tidak ramah lingkungan oleh pencari lokal seperti menggunakan racun dan bahan kimia lain juga dapat mengancam keberadaan ikan dan udang serta dapat mempengaruhi hasil tangkapan komoditas tersebut. Oleh karena itu, pemahaman biologi, parameter lingkungan, dan struktur populasi merupakan hal-hal yang perlu dioptimalkan dalam menjaga ketersediaan hasil alam liar (Deekae and Abowei, 2010).
Crustacea merupakan kelompok subphylum terbesar dalam phylum Arthropoda yang merupakan organisme dengan alat gerak (appendages) bersendi. Crustacea termasuk ke dalam species benthos utama yang terdiri atas udang, kepiting, dan udang karang. Crustacea hidup pada daerah tepian danau, sungai, dan estuarin (Goldman and Horne, 1983).
3
Sebagian besar Crustacea bersifat akuatik, dan bernapas dengan insang. Crustacea memiliki pelindung eksoskeleton yang terbuat dari chitin yang merupakan sebuah residu polysaccharide acetylglucosamine. Eksoskeleton menjadi keras dengan kalsium karbonat atau kuat namun fleksibel oleh proses penyamakan kulit. Klasifikasi Crustacea dilakukan berdasarkan pada potongan dari eksoskeleton dan jumlah segmen serta appendage (Goldman and Horne, 1983). Alat-alat tambahan bersifat biramus (bercabang dua). Kepala terbentuk oleh segmen-segmen yang menyatu, kadang-kadang bersatu dengan dada membentuk chephalothorax, contoh: Cambarus sp. (udang air tawar), Panilirus sp. (udang karang atau lobster), Scylla serrata (kepiting), Portunus sp. (rajungan) dan Uca sp. (ketam), Mitela sp. dan Balanus sp. (barnakel), Cyclops sp. dan Daphnia sp. (pinjal air) (Brotowidjoyo, 1994).
Berbeda dengan ordo lain dalam classis Malacostraca, pada ordo Decapoda terdapat tiga pasang apendik pada ruas thoraks pertama yang termodifikasi menjadi maksiliped dan 5 pasang apendik thoraks berikutnya sebagai kaki jalan atau pereiopoda. Pasangan kaki jalan pertama seringkali berukuran besar dan bercapit, disebut cheliped. Eksopodit pada kaki jalan biasanya tidak ada. Kepala tumbuh menyatu dengan semua ruas thoraks di bagian dorsal, tepi lateral karapas menutup seluruh insang yang terletak dalam rongga insang (Suwignyo et al., 2005).
Berdasarkan bentuk, cara hidup, dan habitatnya, ordo Decapoda dibagi menjadi dua subordo yaitu Natantia dan Reptantia. Subordo Natantia terdiri atas udang air tawar dan laut. Kepiting, udang karang (lobster), klomang, dan rajungan
4
termasuk ke dalam subordo Reptantia. Natant yang berarti berenang, sedangkan Reptant berarti merayap (Suwignyo et al., 2005).
Subordo Natantia dibagi menjadi 3 familia yaitu Palaemonidae, Atyidae, dan Alpheidae. Familia Palaemonidae dan familia Atyidae mempunyai bagian mata tidak ditutupi oleh karapas, panjang rostrum normal, pereiopoda pertama lebih kecil daripada pereiopoda kedua, pada pereiopoda kedua terdapat satu karpus. Familia Alpheidae mempunyai bagian mata yang tertutupi oleh karapas, rostrum pendek, pereiopoda pertama lebih besar daripada pereiopoda kedua, pereiopoda kedua mempunyai karpus yang terdiri atas lima bagian (ruas). Familia Palaemonidae mempunyai duri hepatik, pereiopoda pertama dan kedua tanpa setae. Familia Atyidae tidak mempunyai duri hepatik, pereiopoda pertama dan kedua mempunyai setae yang bertumpuk tebal pada ruas jari terakhir (Wowor et al., 2004 dalam Yule and Sen, 2004).
Kepiting termasuk sub ordo Reptantia dan termasuk ke dalam familia Brachyura, yang sama-sama mempunyai karakteristik lima pasang kaki dada atau thoracic legs (pereiopoda). Pada kepiting sungai, pereiopoda pertama termodifikasi sebagai capit (cheliped), dan empat pasang kaki sisanya termodifikasi sebagai kaki jalan (walking legs). Bagian tubuh kepiting sungai terdiri atas kepala, thoraks dan abdomen. Kepala dan thoraks ditutupi oleh karapaks yang lebar, abdomen mereduksi, memipih dan lentur di bawah thoracic sternum. Pada kepiting sungai dewasa, abdomen jantan ramping dan sempit serta berbentuk triangular atau bentuk T. Abdomen betina lebar dan membulat serta menutupi seluruh sternum. Jantan dewasa mempunyai dua pasang abdominal appendage (pleopoda) yang termodifikasi menjadi alat kopulasi yang dikenal
5
dengan pleopoda. Bentuk pleopoda sangat penting dalam taksonomi, terutama karena morfologi eksternal dari kepiting sungai cenderung agak konservatif (Ng, 1988; Cumberlidge, 1999; dalam Yeo et al., 2008).
Menurut Wowor (Komunikasi Pribadi, 5 Mei 2012), sebagian besar udang air tawar yang biasa ditemukan di sungai-sungai Pulau Jawa yaitu familia Palaemonidae dan sebagian kecil dari familia Atyidae. Pulau Jawa masuk ke dalam daerah Dataran Sunda. Menurut Chia dan Ng (2006), jenis kepiting air tawar yang biasa ditemukan di Dataran Sunda (Sumatra, Semenanjung Malaysia, Jawa, Borneo (Kalimantan), Palawan, Mindoro, Bali) berasal dari genus Sundathelphusa dan Parathelphusa, sedangkan genus Perbrinckia merupakan genus yang terpisah distribusinya, hanya ada di Jawa, Borneo dan Srilanka (Ng, 1995).
Hubungan kekerabatan antara kelompok hewan Crustacea (Decapoda) diteliti menggunakan metode analisis kladistik. Analisis kladistik (cladistic analysis) telah menjadi sinonim dengan sistematika filogenetik. Suatu klad atau clade (Bahasa Yunani clados, yang berarti “cabang”) adalah suatu cabang evolusi. Analisis kladistik mengelompokkan organisme menurut urutan waktu munculnya percabangan itu disepanjang pohon filogenetik yang bercabang dua (dikotomi). Masing-masing titik percabangan dalam suatu pohon didefinisikan atau ditentukan oleh homologi baru yang unik bagi beberapa species pada cabang tersebut (Campbell et al., 2003). Metode kladistik secara langsung menyatakan hubungan kekerabatan filogenetik dari organisme dengan menggunakan diagram bercabang yang disebut kladogram (Winston, 1999).
6
Menurut Campbell et al. (2003), analisis kladistik menggunakan suatu konsep yang disebut outgroup comparisson untuk mengenali karakter primitif bagi semua anggota kelompok yang ingin diteliti dan untuk memantapkan titik permulaan untuk membangun sebuah pohon filogenetik. Outgroup adalah suatu species atau kelompok species yang relatif masih berkerabat dekat dengan kelompok species yang sedang dipelajari, tetapi jelas-jelas hubungannya tidak sedekat hubungan antar anggota species yang sedang dipelajari. Semua anggota ingroup yang dipelajari dibandingkan dengan outgroup tadi. Karakter yang dimiliki bersama oleh outgroup dan ingroup kemungkinan besar telah dimiliki juga oleh nenek moyang bersama dan dengan demikian dinamakan karakter primitif (plesiomorfi) yang dimiliki bersama.
Analisis kladistik mencari sinapomorfi (karakter turunan yang dimiliki bersama) untuk membangun pohon filogenetik. Karakter tersebut merupakan homologi yang berkembang dari nenek moyang bersama semua species pada satu cabang suatu percabangan pohon tersebut, tetapi tidak ditemukan pada nenek moyang cabang lain. Percabangan pertama pohon itu menggambarkan nenek moyang yang sama-sama dimiliki oleh cabang yang mengandung outgroup maupun ingroup. Cabang-cabang pohon itu ditentukan dengan cara membandingkan jumlah karakter yang diturunkan pada masing-masing takson (Campbell et al., 2003). Ingroup yang dipelajari hubungan kekerabatannya dalam penelitian ini adalah kepiting dan udang yang ditangkap di Sungai Cijalu merupakan, sedangkan outgroup-nya adalah Isopoda.
7
Berdasarkan latar belakang yang telah dikemukakan di atas, maka permasalahan yang muncul adalah sebagai berikut:
1. Bagaimana diversitas species ordo Decapoda yang tertangkap di Sungai Cijalu. 2. Bagaimana hubungan kekerabatan species ordo Decapoda yang tertangkap di
Sungai Cijalu.
Tujuan penelitian ini adalah untuk mengetahui:
1. Diversitas species ordo Decapoda yang tertangkap di Sungai Cijalu.
2. Hubungan kekerabatan species ordo Decapoda yang tertangkap Sungai Cijalu. Manfaat yang dapat diperoleh dari penelitian ini diharapkan mampu memberikan informasi dasar tentang species dari ordo Decapoda yang ada di Sungai Cijalu sehingga diharapkan berguna dalam kepentingan manajemen badan air Sungai Cijalu, penelitian lanjutan, upaya konservasi, maupun upaya budidaya.
8
II. MATERI DAN METODE
1. Materi, Lokasi, dan Waktu Penelitian
1.1. Materi Penelitian
1.1.1. Bahan Penelitian
Bahan-bahan yang digunakan dalam penelitian ini adalah alkohol 70%, kertas label, kertas HVS, udang dan kepiting yang ditangkap di Sungai Cijalu, Majenang serta Isopoda.
1.1.2. Alat Penelitian
Alat yang digunakan dalam penelitian ini adalah botol koleksi, ice box, bak preparat, sarung tangan, alat tulis, electric shocker, seser, kantong plastik, milimeter blok, kamera digital, kaca pembesar, penggaris, jangka sorong, pinset, mikroskop stereo, cawan petri, kertas kalkir dan gunting.
1.2. Lokasi dan waktu penelitian
Penelitian ini dilakukan di sepanjang aliran Sungai Cijalu Kecamatan Majenang Kabupaten Cilacap. Identifikasi dilakukan di Laboratorium Taksonomi Hewan Fakultas Biologi Universitas Jenderal Soedirman Purwokerto dan dilakukan verifikasi ke Laboratorium Crustacea Pusat Penelitian Biologi Bidang Zoologi LIPI Cibinong. Penelitian ini dilaksanakan selama 6 bulan, mulai pada bulan Juni-Nopember 2012.
9
2. Metode Penelitian 2.1 Teknik Pengambilan Sampel
Penelitian ini menggunakan metode survei dengan teknik pengambilan sampel secara acak kelompok atau Cluster Random Sampling dengan lokasi yang ditetapkan yaitu di bagian hulu, tengah, dan hilir sungai.
2.2 Cara Kerja Penelitian
2.1.1. Pengambilan Sampel udang dan kepiting
Pengambilan sampel species ordo Decapoda dilakukan di tiga lokasi Sungai Cijalu yaitu hulu (Desa Sepatnunggal), tengah (Desa Mulyadadi), dan hilir (Desa Pahonjean). Lokasi penangkapan species ordo Decapoda dilakukan di 11 tempat, yaitu di bagian hulu sebanyak 4 stasiun, bagian tengah 4 stasiun dan bagian hilir 3 stasiun. Sampel udang dan kepiting diambil menggunakan electric shocker dan seser sebanyak 3 kali dengan bergeser ke titik lain pada setiap stasiun, dalam interval waktu 20 menit.
2.1.2. Penanganan Spesimen di Lapangan
Spesimen species dari ordo Decapoda yang didapatkan langsung diawetkan ke dalam botol koleksi atau kantung plastik yang berisi larutan alkohol 70%, kemudian diberi label yang berisi data lokasi stasiun pengambilan sampel.
10 2.1.3. Penanganan Spesimen di Laboratorium
Spesimen Crustacea yang didapatkan dari lapangan dicuci menggunakan air mengalir untuk menghilangkan kotoran yang terbawa, kemudian larutan alkohol yang sebelumnya diganti dengan larutan alkohol 70 % yang baru.
2.1.4. Pengamatan dan Pengukuran Sampel
Pengamatan yang dilakukan pada sampel udang, dan kepiting yaitu rostrum, bentuk post antenula, gigi pada tepi anterolateral, duri pada pereiopoda pertama, merus pada pereiopoda pertama, jumlah pereiopoda, bentuk karpus pereiopoda pertama, ukuran chela pereiopoda pertama, ukuran besar kedua pereiopoda pertama, chela pereiopoda kedua, ukuran besar kedua pereiopoda kedua, bentuk karpus pereiopoda kedua, letak pubescence pada chela pereiopoda kedua, panjang dan pendeknya pubescence pada chela pereiopoda kedua, kepadatan pubescence pada chela pereiopoda kedua, celah chela pada pereiopoda kedua, tuberkel pada chela, preanal carina, abdomen, uropod, telson. Pengukuran yang dilakukan pada sampel udang, dan kepiting yaitu panjang rostrum, panjang antenula peduncle, tebal dan lebar karapas, pereiopoda pertama (panjang karpus, merus, dan chela), pereiopoda kedua (panjang karpus, merus, dan chela).
2.1.5. Identifikasi dan Determinasi
Sampel udang dan kepiting diidentifikasi dan dideterminasi berdasarkan Kunci Determinasi Brachyura oleh Ng (2004) dalam Yule and Sen (2004), Caridea oleh Wowor et al. (2004) dalam Yule and Sen (2004) dan Kepiting Air Tawar oleh Bott (1970), dengan dilakukan verifikasi ke Laboratorium Crustacea Pusat Penelitian Biologi Bidang Zoologi LIPI Cibinong.
11 2.1.6. Penentuan Hubungan Kekerabatan
Penentuan hubungan kekerabatan species Crustacea (Decapoda) yang diperoleh secara kladistik dilakukan dengancara kerja sebagai berikut.
a. Karakter morfologi dari OTU ditentukan dan disusun dalam bentuk matrik. b. Karakter morfologi yang dapat dibandingkan diseleksi agar menghasilkan
gambaran secara umum. Karakter yang digunakan merupakan multi state karakter dengan nilai skor angka 0, 1, 2, 3, dan 4. Angka 0 merupakan nilai skoring yang paling rendah (primitif), angka 1, 2, 3, dan 4 merupakan nilai tinggi pada skoring yang menunjukkan kemajuan karakter.
c. Hubungan kekerabatan ditentukan berdasarkan karakter morfologi yang dibandingkan pada semua species ordo Decapoda atau antara species ingroup dan species outgroup yang termasuk ke dalam taksonomi opersional.
3. Metode Analisis
3.1. Data berupa karakter morfologi, skoring, dan kondisi outgroup dan ingroup Crustacea yang tertangkap di sungai Cijalu yang disusun dalam matriks dan dianalisis secara cladistic menggunakan algoritma maximum parsimony dengan bantuan program Phylogenetic Analysis Using Parsimony (PAUP) versi 4.0b10 menurut Swofford (1998) untuk mengetahui hubungan kekerabatannya.
3.2. Langkah-langkah dalam melakukan analisis filogenetik dan merekonstruksi pohon filogenetik menurut Anonymous (2002):
1. Membuat hasil pengamatan karakter morfologi dan pemberian skoring, untuk tiap karakter dan disusun dalam bentuk matrik.
12
2. Memindahkan matriks data ke dalam program PAUP dalam bentuk
nexus.files.
3. Menyimpan data matriks dalam posisi yang benar (tidak terdapat kesalahan pemasukan matriks skoring, sesuai urutan dan karakter yang diamati) dengan klik save.
4. Ketik nama file dengan menambahkan “.nex” kemudian klik Ok . 5. Buka program PAUP.
6. Buka file yang telah disimpan, klik file kemudian klik kembali open. 7. Ketik “set criterion=parsimony;” pilih execute.
8. Ketik “hsearch;” pilih execute kemudian klik close.
9. Selanjutnya untuk memunculkan pohon ketik “showtrees;” pilih execute. 10. Ketik “describe all/chglist apolist;” untuk memunculkan keterangan
indeks konsistensi (CI) maupun indeks retensi (RI).
11. Ketik “pscores/CI=yes” untuk melihat daftar nilai CI pada tiap branch. 12. Ketik “pscores/RI=yes” untuk melihat daftar nilai RI pada tiap branch. 13. Ketik “bootstrap nreps=1000” untuk mengetahui tingkat kepercayaan
hasil pohon filogeni yang terbentuk.
14. Simpan pohon dengan ketik “savetrees;” pilih execute.
15. Buka software Treeview (Treev32) untuk membuka pohon yang telah didapatkan.
16. Setelah programnya terinstal klik icon treeview. 17. Klik open file nexus hasil show trees.
18. Bentuk kladogram dapat disesuaikan dengan keinginan, slanted cladogram atau rectangular cladogram.
13
III. HASIL DAN PEMBAHASAN
3.1. Deskripsi Lokasi Pengambilan Sampel
Sungai Cijalu terletak di Kecamatan Majenang Kabupaten Cilacap. Panjang sungai lebih kurang 15 km dengan lebar sungai antara 15-42 m dan kedalaman antara 0,3-0,9 m. Substrat dasar sungai berupa batu, kerikil, pasir, dan lumpur. Sungai tersebut mempunyai fungsi sosial ekonomi bagi masyarakat di sekitarnya anatra lain untuk kepentingan sehari-hari seperti mandi, mencuci, kakus, membuang limbah domestik, dan sumber air untuk kepentingan pertanian.
Pembagian lokasi dan gambar lokasi pengambilan sampel disajikan pada Lampiran 1 dan Lampiran 2. Kondisi secara umum lokasi pengambilan sampel adalah sebagai berikut.
Bagian hulu Sungai Cijalu dibagi menjadi 4 lokasi pengambilan sampel yaitu stasiun I, II, III, dan IV. Stasiun I terletak di Kampung Larangan Desa Sepatnunggal, Stasiun II terletak di Kampung Kutangsa Desa Bener, Stasiun III terletak di Kampung Gintung Desa Bener, dan Stasiun IV terletak di Kampung Lampeng Desa Jenang. Lebar sungai berkisar antara 20-42 m, kecepatan arusnya antara 1,42-1,62 m/s, dan kedalaman 0,5-0,9 m. Substrat dasar berupa batuan besar, kerikil, dan pasir (Lampiran 3). Kondisi lingkungan masih alami, disekitar sungai terdapat hutan pinus, bambu, semak-semak, persawahan dan jauh dari pemukiman penduduk. Akses menuju lokasi pengambilan sampel sangat sulit dengan kondisi jalan yang terjal, berbatu, dan curam.
Bagian tengah Sungai Cijalu dibagi menjadi 4 lokasi sampling yaitu stasiun V, VI, VII, dan VIII. Stasiun V terletak di Kampung Selis Desa Jenang, Stasiun VI terletak di Kampung Mulyadadi Desa Mulyadadi, Stasiun VII terletak di
14
Kampung Mulyasari Desa Mulyadadi, dan Stasiun VIII terletak di Kampung Rawalo Desa Pahonjean. Lebar sungai berkisar antara 17-32 m, kecepatan arusnya antara 0,39-1,34 m/s, dan kedalaman 0,3-0,7 m. Substrat dasar berupa batuan kecil, pasir, dan lumpur (Lampiran 3). Kondisi lingkungan disekitar sungai semak-semak, persawahan, perkebunan, dan pemukiman penduduk. Akses menuju lokasi pengambilan sampel mudah dijangkau dari jalan kemudian turun ke sungai.
Bagian hilir Sungai Cijalu dibagi menjadi 3 yaitu stasiun IX, X, dan XII. Stasiun IX terletak di Kampung Mbale Kambang Desa Pahonjean, Stasiun X terletak di Kampung Mbale Kambang Desa Pahonjean, dan Stasiun XI terletak di Kapung Cilanggir Kecamatan Wanareja. Lebar sungai berkisar antara 15-17 m, kecepatan arusnya antara 0,2-0,53 m/s, dan kedalaman 0,4-0,9 m. Substrat dasar berupa pasir dan lumpur (Lampiran 3). Kondisi lingkungan disekitar sungai merupakan persawahan dan perkebunan. Akses menuju lokasi pengambilan sampel pada stasiun yang terakhir dijangkau dengan berjalan kaki sekitar 2 km yang melewati jembatan bambu dan perkebunan singkong.
3.2. Perolehan Sampel Udang dan Kepiting
Berdasarkan hasil identifikasi, determinasi, dan verifikasi didapatkan 5 species anggota ordo Decapoda yang ditangkap di sungai Cijalu yaitu 3 species udang dari genus Macrobrachium dan 2 species kepiting yang termasuk dalam genus Parathelphusa. Species udang yang ditemukan di sungai Cijalu yaitu Macrobrachium pilimanus, M. lanchesteri, dan M. sintangense. Species udang yang tertangkap di sungai Cijalu berbeda dengan yang ditemukan di sungai
15
Cileumeuh, yaitu species M. cowlesi, M. idae, M. idella, dan M. oenone (Arifiyanto, 2008).
Species kepiting yang ditemukan di sungai Cijalu yaitu Parathelphusa convexa dan P. bogorensis. Menurut Ng (2004), kepiting air tawar yang sesungguhnya yaitu kepiting yang tidak perlu kembali ke laut untuk melepaskan telurnya. Anggota infraordo Brachyura yang termasuk kepiting air tawar yaitu species dari familia Parathelphusidae, Potamidae, dan Gecarcinucidae. Menurut Wowor (2010), P. bogorensis dan P. convexa merupakan species yang umum ditemukan di sungai-sungai di Pulau Jawa. Penelitian di DAS Ciliwung ditemukan empat species kepiting yaitu Malayopotamon javanense, Parathelphusa bogorensis, P. convexa, Geosesarma sp.
Daftar selengkapnya species ordo Decapoda yang tertangkap di Sungai Cijalu disajikan pada Tabel 3.1.
Tabel. 3.1 Data Perolehan Sampel Udang dan Kepiting
Lokasi Stasiun Udang Kepiting Jumlah Jenis Jumlah Jenis
Hulu I 2 M. pilimanus (2♂) 11 P. bogorensis (3♂; 5♀) P. convexa (2♂; 1♀) II - - 5 P. bogorensis (1♂; 2♀) P. convexa (2♂) III - - - - IV 61 M. sintangense (20♂; 41♀) 2 P. convexa (2♂) Tengah V 25 M. sintangense (8♂;17♀) 11 P. bogorensis (1♂; 3♀)
16
Lokasi Stasiun Udang Kepiting Jumlah Jenis Jumlah Jenis
Tengah V 7 P. convexa (2♂; 5♀) Tengah VI 11 M. sintangense (10♂; 1♀) 7 P. bogorensis (1♀) P. convexa (3♂; 3♀) VII 34 M. sintangense (24♂; 10♀) 5 P. convexa (1♂; 4♀) VIII 28 M. sintangense (12♂; 12♀) M. lanchesteri (2♂; 2♀) - - Hilir IX 33 M. sintangense (12♂; 3♀) M. lanchesteri (2♂; 16♀) 7 P. convexa (2♂; 5♀) X 38 M. sintangense (6♂; 5♀) M. lanchesteri (25♂; 2♀) - - XI 120 M. sintangense (15♂; 9♀) M. lanchesteri (57♂; 29♀) - -
17
3.3. Deskripsi Species Udang dan Kepiting yang Ditemukan
3.3.1. Species udang
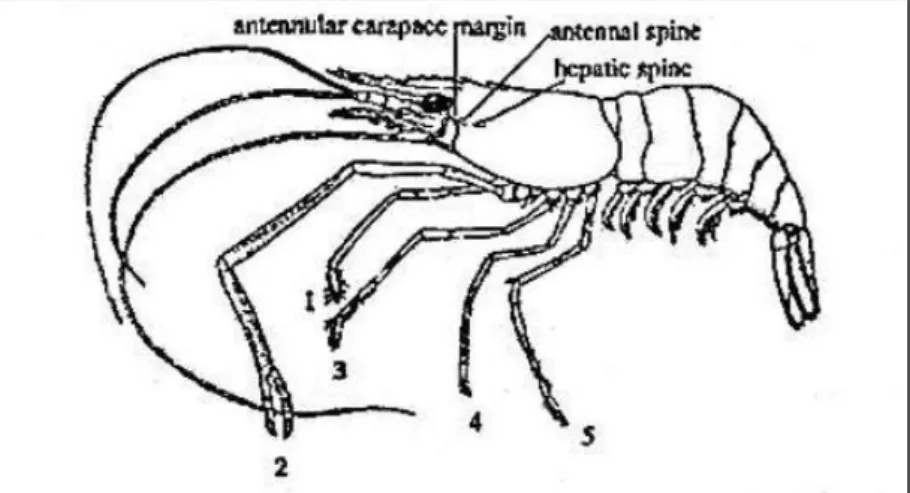
Gambar 3.1. Morfologi umum udang air tawar genus Macrobrachium (Sumber: Carpenter and Niem, 1998 dalam Wowor et al., 2004 dalam Yule and Sen, 2004).
Klasifikasi udang Macrobrachium (Gambar 3.1.) yang tertangkap di Sungai Cijalu menurut Wowor (2004) dalam Yule and Sen (2004) adalah sebagai berikut: Regnum : Animalia Phylum : Arthropoda Subphylum : Crustacea Classis : Malacostraca Ordo : Decapoda Infraordo : Caridea Familia : Palaemonidae Genus : Macrobrachium
Species : Macrobrachium pilimanus (De Man, 1879); Macrobrachium sintangense (De Man, 1898); Macrobrachium lanchesteri (De Man, 1911)
18 3.3.1.a. Macrobrachium pilimanus
Macrobrachium pilimanus hanya ditemukan di Stasiun I yang lokasinya berada di bagian hulu sungai (Tabel 3.1). Jumlah yang ditemukan adalah sebanyak 2 individu. M. pilimanus ditangkap di tepian Sungai Cijalu yang terdapat banyak sampah daun dan memiliki substrat berbatu, dan berpasir. Menurut Ou and Yeo (1995), M. pilimanus dapat ditemukan di aliran sungai yang deras dengan substrat berpasir dan berbatu dengan sampah daun sepanjang tepian sungai, tetapi tanpa vegetasi yang padat. Berikut adalah gambar morfologi M. pilimanus yang tertangkap di Sungai Cijalu:
Gambar 3.2. M. pilimanus tampak lateral
Keterangan:
1. Rostrum pendek 2. Pereiopoda pertama
3. Pereiopoda kedua ukuran keduanya tidak sama dengan pubescence lembut yang padat 4. Pereiopoda ketiga licin
5. Pereiopoda keempat licin
6. Pereiopoda kelima licin (panjangnya kira-kira sama dengan pereiopoda keempat) 2 4 5 6 3 6 1
19
Gambar 3.3. M. pilimanus tampak dorsal
Keterangan:
1. Panjang rostrum tidak mencapai segmen ketiga antenula peduncle 2. Antenula peduncle
3. Merus
4. Karpus berbentuk cangkir dan lebih pendek dari merus 5. Palm
6. Propodus 7. Jari
8. Chela seluruhnya ditutupi oleh pubescence
Berdasarkan hasil pengamatan dan pengukuran morfometri, M. pilimanus memiliki karakter morfologi (lihat Gambar 3.3.) yang berbeda dibandingkan dengan anggota Crustacea dalam ordo Decapoda yang tertangkap di Sungai Cijalu lainnya dari ciri-ciri bentuk post antenula lurus, ukuran besarnya kedua pereiopoda kedua tidak sama, bentuk karpus pereiopoda kedua silinder, panjang karpus pereiopoda kedua lebih pendek dari merus, chela pereiopoda kedua lebih panjang dari karpus, letak pubescence pada chela pereiopoda kedua ada di sepanjang chela, pubescence pada chela pereiopoda kedua panjang dan padat, terdapat celah pada pereiopoda kedua, tuberkel terdapat di tengah chela, dan memiliki preanal carina atau duri kecil sebelum bagian anal seperti Gambar 3.4. dibawah ini. 4 6 6 3 5 6 7 8 6 1 6 2
20
(a) (b)
Gambar 3.4. (a) Skema Preanal carina (Sumber: Wowor et al., 2004 dalam Yule and Sen, 2004); (b) M. pilimanus (perbesaran 4x10)
Keterangan: 1. Preanal carina
M. pilimanus memiliki pereiopoda kedua dengan karpus yang pendek dan berbentuk cangkir, permukaan karpus, dan chela halus atau sedikit bergranula. Setae yang lembut dan panjang terdapat pada chela. Gigi dorsal rostrum letaknya berdekatan satu sama lain (Ng, 1995). DistribusiM. pilimanus ditemukan di sepanjang paparan Sunda, dari utara Vietnam dan selatan Yunnan di China sampai Jawa dan Kalimantan (Holthuis, 1950, 1984; Dang and Nguyen, 1972; Dai, 1984; Choong and Khoo, 1987a, 1987b; Choong, 1989; Ng, 1995; Ou and Yeo, 1995; Cai and Dai, 1999; Cai et al., 2003; dalam Wowor and Short, 2007).
3.3.1.b. Macrobrachium sintangense
Species M. sintangense ditemukan di banyak stasiun pengambilan sampel (Stasiun IV, V, VI, VII, VIII, IX, X, dan XI). Jumlah individu M. sintangense yang ditemukan paling melimpah berada di stasiun IV (Tabel 3.1). Sebagian besar sampel species yang ditangkap adalah individu betina yang sedang bertelur dan
21
ditangkap di aliran sungai yang teduh. Menurut Sabar (1979) dalam Cai, (2004), M. sintangense umumnya ditemukan di aliran sungai yang lebar dan sungai yang relatif lebih cepat aliran airnya. Gambar morfologi M. sintangense yang tertangkap di Sungai Cijalu disajikan pada gambar dibawah ini.
Gambar 3.5. (a) M. sintangense tampak lateral; (b) skema bentuk post
antenula (Sumber: Wowor et al., 2004 dalam Yule and Sen,
2004); (c) M. sintangense tampak dorsal; (d) foto bentuk post
antenula M. sintangense
Keterangan: (a) 1: rostrum, 2: pereiopoda pertama, 3: pereiopoda kedua ukuran keduanya sama dengan pubescence 2/3 proksimal jari, 4: pereiopoda ketiga licin, 5: pereiopoda keempat licin, 6: pereiopoda kelima licin; (b) skema post antenula; (c) 1: panjang rostrum mencapai segmen ketiga antenula peduncle, 2: antenula peduncle, 3: merus, 4: karpus pereiopoda kedua berbentuk subsilindris, 5: propodus, 6: palm, 7: jari, 8: chela; (d) 1: Duri hepatik, 2: Post antenula membulat dengan tepi karapas
Berdasarkan pengamatan dan pengukuran morfometri (lihat Gambar 3.5.), M. sintangense dapat dibedakan dengan anggota Crustacea dalam ordo Decapoda yang tertangkap di Sungai Cijalu lainnya dengan melihat karakter morfologi dari
(d) (c) 1 6 5 4 2 3 4 5 7 3 8 6 1 2 1 2 (a) (b)
22
bentuk post antenula membulat, tidak memiliki preanal carina, ukuran besarnya kedua pereiopoda kedua sama, dan bentuk karpus pereiopoda kedua subsilinder. Panjang karpus pereiopoda kedua lebih panjang dari merus, chela pereiopoda kedua lebih pendek dari karpus. Letak pubescence pada chela pereiopoda kedua hanya terdapat jari, pubescence pada chela pereiopoda kedua pendek dan padat, kepadatan pubescence pada chela pereiopoda kedua hanya 2/3 proksimal jari, jari pereiopoda kedua tidak bercelah, terdapat 2 baris tuberkel di tepi pemotong chela seperti Gambar 3.6. dibawah ini.
(a) (b)
Gambar 3.6. (a) Skema Pubescence pada M. sintangense (Sumber: Wowor et
al., 2004 dalam Yule and Sen, 2004); (b) chela pada M. sintangense (perbesaran 4x10)
Keterangan:
1. Dua baris tubercle di tiap tepi pemotong (dactylus)
Saat proses identifikasi terdapat banyak species yang meragukan untuk disebut sebagai M. sintangense, terutama jika dilihat dari ciri dua baris tuberkel di kedua tepi pemotong dan adanya pubescence yang kadang tidak telihat terutama pada saat mengidentifikasi species betina. Menurut Holthuis, 1950; Chong et al., 1987 dalam Cai et al. (2004), pubescence pada jari yang dapat bergerak rupanya hanya jelas terlihat dominan pada species jantan. Begitu pula, dengan dua baris
23
tuberkel pada kedua tepi pemotong dari dactylus yang biasanya kosong, hal itu mungkin terjadi karena species tersebut belum dewasa.
M. sintangense memiliki ciri jari ditutupi dengan pubescence sepanjang 2/3 proksimal jari. Selain itu, terdapat satu baris tubercle diantara tepi pemotong dari dactylus (tubercle pada sisi tengah lebih menonjol) dan pada sisi tengah dari jari yang tidak dapat bergerak (Chong et al.,1987). M. sintangense pertama ditemukan dari daerah Sintang di Lembah Sungai Kapuas, kemudian banyak dilaporkan ditemukan di negara-negara Asia Tenggara (Holthuis, 1950; Ng, 1990 dalam Ng, 1995).
3.3.1.c. Macrobrachium lanchesteri (De Man, 1911)
Macrobrachium lanchesteri merupakan jenis udang yang termasuk dalam familia Palaemonidae yang ditemukan di Stasiun VIII, IX, X, dan XI. Jika diperhatikan udang jenis tersebut banyak ditangkap dari hilir Sungai Cijalu. Semakin ke hilir, jumlah individu dari species makin melimpah (Tabel 3.1). Udang tersebut ditangkap pertama kali di stasiun VIII yang termasuk diakhir bagian tengah Sungai Cijalu dan memiliki topografi perairan yang tenang dan lebar (Lampiran 2). M. lanchesteri juga ditemukan di stasiun-stasiun berikutnya yang merupakan hilir Sungai Cijalu.
Berdasarkan hasil pengamatan dan pengukuran morfometri, M. lanchesteri dapat dibedakan dengan anggota Crustacea dalam ordo Decapoda lainnya dengan melihat karakter morfologi dari bentuk post antenula membulat, ukuran besarnya kedua pereiopoda kedua sama, bentuk karpus pereiopoda kedua subsilinder, panjang karpus pereiopoda kedua lebih panjang dari merus, chela pereiopoda kedua lebih pendek dari karpus, chela pereiopoda kedua hanya terdapat jari,
24
pubescence pada chela pereiopoda kedua pendek dan tersebar, kepadatan pubescence pada chela pereiopoda kedua yaitu tersebar di sepanjang jari, jari pereiopoda kedua tidak bercelah, tidak terdapat tuberkel pada chela, dan memiliki preanal carina seperti Gambar 3.7. dibawah ini.
Gambar 3.7. (a) M. lanchesteri tampak lateral, (b) post antenula pada
M. lanchesteri (perbesaran 4x10) dan (c) M. lanchesteri
tampak lateral dorsal, (d) preanal carina M. lanchesteri (e) skema pubescence pada M. lanchesteri (Sumber: Wowor et al., 2004 dalam Yule and Sen, 2004); (f) chela pada M. lanchesteri (perbesaran 4x10)
Keterangan: (a) 1: rostrum memiliki 1-2 gigi apikal, 2: pereiopoda pertama, 3: pereiopoda kedua ukuran keduanya sama, karpus lebih panjang daripada merus, karpus berbentuk subsilindris, jari Pereiopoda kedua di tutupi pubescence lembut seluruhnya 4: pereiopoda ketiga licin, 5: pereiopoda keempat licin, 6: pereiopoda kelima licin; (b) 1: duri hepatik, 2: post antenula membulat dengan tepi karapas; (c) M. lanchesteri tampak lateral dorsal; (d) 1: Preanal carina; (e);(f) 1: tanpa dua baris tubercle di tepi pemotong (dactylus)
1 3 1 2 4 6 5 1 2 (a) (b) 1 (c) (d) (e) (f)
25
Menurut Chong and Khoo (1988), M. lanchesteri mempunyai karakter morfologi berupa rostrum lurus melebihi tepi anterior dari antenula peduncle, pada jantan yang sangat besar, rostrum kadang melebihi tepi anterior dari scapocerit, tepi dorsal conveks pada proksimal, dan datar pada bagian distal; 6-11 gigi (biasanya 8-9) ada pada dorsal, dengan 1 atau 2 gigi yang terpisah pada ujung rostrum dengan daerah kosong yang terlihat jelas, dan 1-2 gigi yang terletak di belakang orbit karapas; tepi ventral lurus dengan 2-7 gigi (biasanya 3/4). Styloceite dan duri antero-lateral antenula lemah dan kecil. Pereiopoda kedua ukurannya sama, sangat ramping; karpus terlihat jelas lebih panjang dari merus dan chela; jantan yang sangat besar dengan karpus sama atau sedikit lebih pendek dari chela, jari sedikit lebih pendek dari palm (terlihat jelas lebih pendek pada jantan dewasa), dengan 2 gigi yang sangat kecil pada ¼ proksimal dari tepi pemotong; capit licin atau halus pada larva dan betina, jantan besar memiliki duri pada seluruh segmen (kecuali jari). Duri yang dapat digerakkan pada exopod dari uropod lemah, tidak terlihat jelas, dan lebih pendek dari gigi tetap bagian luar yang tidak dapat digerakkan.
Cai et al. (2004), mengatakan bahwa M. lanchesteri merupakan species yang hampir beradaptasi dengan baik pada seluruh jenis badan air tawar. Johnson (1963); Cranbrook and Furtado (1988) dalam Othman et al. (2006), menyebutkan bahwa M. lanchesteri umumnya menetap di aliran sungai yang tenang seperti waduk, kolam, parit saluran irigasi, sawah, dan aliran air buatan lainnya yang termasuk badan air tawar. M. lanchesteri terdapat di perairan yang sedikit asam, tetapi jika air terlalu asam dan substrat mengandung alkali akan kurang baik.
26
Menurut Lanchester, 1902; De Man, 1911 dalam Cai et al. (2004), M. lanchesteri merupakan species endemik dan pertama kali ditemukan di Thailand Selatan, dan Thailand Tengah. Akan tetapi, udang air tawar Macrobrachium dari Thailand dan Semenanjung Malaysia tersebut, juga ditemukan di Sabah, Pulau Kalimantan, dan catatan keberadaannya ini pada awalnya diragukan (Suvatti, 1937; Johnson, 1961, 1968; Chong and Khoo, 1988; Ng, 1995 dalam Cai et al., 2004). Selain itu, M. lanchesteri merupakan species umum di Thailand dan diperjualbelikan di hampir seluruh pasar di Thailand, sebagian besar untuk makanan (Cai et al., 2004).
Menurut Wowor (2010), M. lanchesteri secara tidak sengaja masuk ke perairan indonesia bersama dengan bibit ikan ekonomis penting yang bukan asli Indonesia seperti ikan Patin, Lele Dumbo, Mujair yang kemudian ditebar di kolam-kolam dan situ-situ. Udang ini tahan akan kondisi perairan yang ekstrim, seperti suhu air yang relatif tinggi. Selain itu, udang ini juga ditemukan di selokan-selokan sawah yang airnya hangat. Hal itulah yang menyebabkan M. lanchesteri dapat bertahan hidup di daerah lain. Cai et al. (2004), menyatakan bahwa baru-baru ini M. lanchesteri telah ditemukan di Myanmar, China Selatan dan Jawa, Indonesia.
27
27 3.3.2. Species Kepiting
Klasifikasi kepiting Parathelphusa menurut Bott (1970) adalah sebagai berikut: Regnum : Animalia Phylum : Arthropoda Subphylum : Crustacea Classis : Malacostraca Ordo : Decapoda Infraordo : Brachyura Superfamilia : Gecarcinucoidea Familia : Parathelphusidae Genus : Parathelphusa
Species : Parathelphusa convexa (De Man, 1879); Parathelphusa bogorensis.
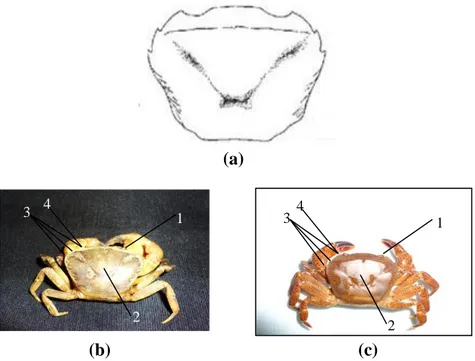
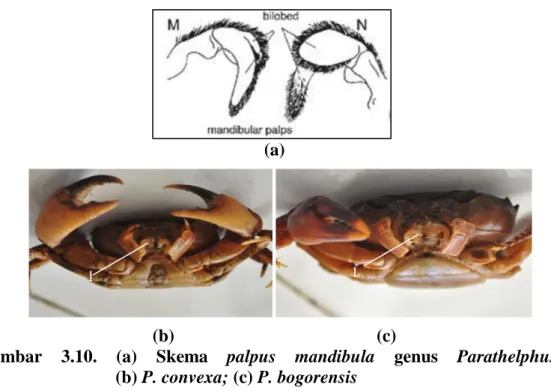
Berdasarkan hasil pengamatan, kepiting genus Parathelphusa memiliki ciri-ciri yaitu mata kecil jika dibandingkan dengan karapas, kornea tidak pernah menutupi hingga tepi karapas. Tepi anterolateral dengan 3 gigi (termasuk gigi orbit eksternal) (Gambar 3.8.). Maxilliped ketiga menutup rapat atau tidak membentuk celah berbentuk jajar genjang, daerah pterygostomial gundul (Gambar 3.9.). Bagian ujung palpus mandibula memiliki dua lobus (Gambar 3.10.). Pleopoda pertama kaku sedangkan pleopoda kedua memiliki ujung seperti benang (Gambar 3.11.). Abdomen jantan berbentuk T dan bertingkat (Gambar 3.12.). Morfologi species genus Parathelphusa disajikan pada gambar dibawah ini.
4 5
7 3
8 1
28
(a)
(b) (c)
Gambar 3.8. (a) Skema bentuk karapas genus Parathelphusa (Sumber: Wowor et al., 2004 dalam Yule and Sen, 2004); (b) P. convexa; (c) P. bogorensis
Keterangan:
1. Mata kecil dibandingkan dengan karapas, kornea tidak pernah menutupi tepi karapas 2. Karapas berbentuk persegi panjang
3. Tepi Anterolateral dengan 3 gigi yang jelas (termasuk gigi orbit eksternal) 4. Gigi orbit eksternal
(a)
(b) (c)
Gambar 3.9. (a) Skema frontal genus Parathelphusa; (b) P. convexa; (c) P. bogorensis
Keterangan:
1. Maxilliped ketiga menutup rapat, tidak membentuk celah berbentuk jajar genjang 2. Wilayah pterygostomial tanpa setae (gundul)
1 1 2 2 1 2 3 4 3 2 1 4
29
29
(a)
(b) (c)
Gambar 3.10. (a) Skema palpus mandibula genus Parathelphusa; (b) P. convexa; (c) P. bogorensis
Keterangan:
1. Palpus mandibula mempunyai 2 lobus
(a)
(b) (c)
Gambar 3.11. (a) Skema pleopoda pertama genus Parathelphusa; (b) P. convexa; (c) P. bogorensis Keterangan: 1. Pleopoda pertama 2. Pleopoda kedua 1 1 1 2 1 2
30
(a)
(b) (c)
Gambar 3.12. (a) Skema bentuk abdomen ♂ genus Parathelphusa (Sumber: Wowor et al., 2004 dalam Yule and Sen, 2004); (c) P. convexa; (b) P. bogorensis
Keterangan: 1. Abdomen ♂
3.3.2.a. Parathelphusa convexa
P. convexa ditemukan di stasiun I, II, IV, VI, VII, dan IX. Species ini ditemukan di tepian sungai, dibalik batu yang berpasir, dan tergenang air. P. convexa ditemukan merata sepanjang Sungai Cijalu, yaitu ditemukan di hulu, tengah dan hilir sungai. Akan tetapi, mulai bagian tengah sampai hilir sungai jumlahnya meningkat dibandingkan bagian hulu sungai (Tabel 3.1). Menurut Wowor (2010), P. convexa memiliki penyebaran yang luas dan dapat ditemukan di sawah dan selokan yang airnya tergenang atau berarus lambat di daerah hulu sampai dengan selokan-selokan maupun situ-situ di daerah hilir.
Berdasarkan hasil pengamatan dan pengukuran morfometri, P. convexa memiliki karakter morfologi yang khas dibandingkan dengan anggota Crustacea
1 1
31
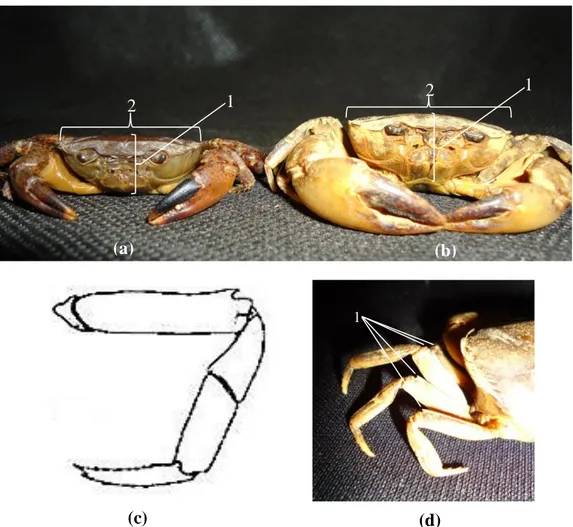
ordo Decapoda yang tertangkap di Sungai Cijalu lainnya dari ciri-ciri karapas tebal dan melengkung. Tebal karapas ½ dari lebar karapas. Duri pereiopoda pertama yang tajam. Merus pereiopoda pertama berduri. Morfologi karapas dan pereiopoda disajikan pada Gambar 3.13.b.c.d. dibawah ini.
Gambar 3.13. (a) ketebalan karapas P. bogorensis kurang dari 1/2 lebar karapas; (b) ketebalan karapas P. convexa kira-kira 1/2 lebar karapas (c) skema pereiopoda P. convexa (Sumber: Wowor et al., 2004 dalam Yule and Sen, 2004); (d) foto pereiopoda P. convexa
Keterangan: (a dan b) 1: tebal karapas, 2: lebar karapas; (d) 1: merus memiliki duri.
Menurut Bott (1970), P. convexa memiliki karapas rata yang kemudian melengkung, tebalnya kira-kira ½ lebar karapas. Ujung gigi epibranchial diarahkan ke dalam, merus pereiopoda pertama sampai kelima dengan duri
1 2 1 2 (a) (b) 1 (c) (d)
32
pradistal yang runcing. Abdomen berbentuk T yang terdiri atas 5 ruas. Hewan relatif kecil. Pleopoda pertama ramping, pendek dan berbentuk tombak. Pleopoda pertama pada bagian distal sedikit menekuk ke dalam. Pleopoda kedua dengan filamen terminal. Pereiopoda pertama kuat, dengan jari capit yang pendek, kuat, hampir edentulous, dan bagian basal sangat menganga, serta mempunyai dactylus yang bengkok. Karpus dengan duri tajam di bagian dalam, merus dari pereiopoda pertama sampai kelima dengan duri pradistal yang runcing. Pereiopoda kedua sampai kelima pendek dan kuat. Distribusi geografis P. convexa sejauh ini sangat dikenal di Pulau Jawa seperti Bogor, Garut, Jawa tengah.
3.3.2.b. Parathelphusa bogorensis
P. bogorensis ditemukan di stasiun I, II, V, dan VI. Species ini ditemukan di hulu Sungai Cijalu yaitu di stasiun I, dan II, tidak ditemukan lagi di stasiun III dan IV yang masih termasuk hulu sungai. Akan tetapi, masih ditemukan di stasiun V, dan VI yang sudah termasuk ke dalam sungai bagian tengah. Hal ini terjadi mungkin karena kedua stasiun V dan VI berada dibawah jembatan dan bendungan yang terdapat batu-batu besar sehingga berarus kencang. Menurut Wowor (2010), P. bogorensis merupakan krustasea yang terdapat di daerah hulu dan penghuni sungai berarus deras.
Berdasarkan hasil pengamatan dan pengukuran morfometri, P. bogorensis memiliki karakter morfologi yang khas dibandingkan dengan anggota Crustacea dalam ordo Decapoda yang tertangkap di Sungai Cijalu lainnya dari ketebalan karapas < ½ dari lebar karapas (Gambar 3.13.a). Duri pereiopoda pertama tumpul. Merus pereiopoda pertama tidak berduri. Morfologi pereiopoda P. bogorensis disajikan pada Gambar 3.14. dibawah ini.
33
(a) (b)
Gambar 3.14. (a) Skema (Sumber: Wowor et al., 2004 dalam Yule and Sen, 2004) dan (b) foto pereiopoda P. bogorensis
Keterangan:
1. Merus memiliki duri.
Menurut Bott (1970), karapas datar dan halus atau licin, kasar pada bagian samping. Dahi datar pada bagian atas, sedikit menekuk, segitiga pada bagian frontal membatasi dengan tajam dimana segitiga tersebut lebih pendek dari daerah kosong yang lebar dari akhir tepian karapas. Cervikalfurche dalam dan luas, dengan sulcus semi bundar yang dihubungkan tanpa halangan. Gigi exorbital tumpul membulat, rendah, tepi sebelah luar yang berliku. Gigi relatif kecil, ujung diarahkan ke dalam. Luasan terbesar dari karapas berada dibelakang gigi epibranchial. Cervikalfurche mencapai diantara tepi anterolateral gigi epibranchial dan gigi tengah. Merus dari Maxiliped ketiga berbentuk bulat telur mendatar. Ischium berlekuk longitudinal mendekati bagian dalam tepi yang terdapat exopodit. Abdomen diantara segmen ke-6 dan kelima sedikit dapat digerakkan. Pleopoda pertama ramping dan panjang, relatif kecil meruncing, bagian distal membuka, dan berbentuk S. Pleopoda kedua memiliki filamen terminal yang panjang. Pereiopoda pertama pada jantan dewasa kuat dan tidak
34
rata permukaannya. Pemotong yang besar dengan dactylus membelok kuat dan tebal, pada bagian bonggol basal terdapat duri. Jari dengan gigi besar pada tengah dari tepi pemotong. Jari lebih pendek dari tinggi palma, jarang yang panjang jarinya separuh panjang palma. Karpus dengan tonjolan yang keluar, merus tanpa duri pradistal. Pereiopoda kedua sampai kelima lebar, merus datar tanpa duri pradistal, tepian yang lebih rendah permukaannya halus. Distribusi P. bogorensis di Pulau Jawa antara lain ditemukan di Jawa Tengah, Djelok, Salatiga, Bogor, Garut, dan di katinggian 5000 dpl Gunung Arjuna (Bott, 1970).
3.3 Kunci determinasi species udang dan kepiting yang ditemukan
Kunci determinasi dan data morfometri udang dan kepiting dari ordo Decapoda yang tertangkap di Sungai Cijalu disajikan pada Lampiran 4 dan Lampiran 5.
3.4 Hubungan Filogenetik Species Ordo Decapoda
Hubungan filogenetik antar species ordo Decapoda dibedakan menggunakan 28 simplesiomorphy characters dari Satuan Taksonomi Operasional atau OTU (Operational Taxonomy Unit) yang diamati dan dianalisis dalam bentuk skoring yang disajikan pada Lampiran 6. Proses skoring dilakukan dengan membandingkan karakter morfologi ingroup dengan karakter morfologi yang dimiliki outgroup. Pemilihan outgroup ditentukan berdasarkan asumsi bahwa ordo Isopoda merupakan ordo primitif yang mememiliki kekerabatan terdekat dengan species anggota ordo Decapoda. Hasil analisis kladistik terhadap data karakter morfologi, skoring, dan kondisi ingroup dan outgroup diperoleh kladogram pada Gambar 3.15.
35
Gambar 3.15. Pohon filogeni dengan konsensus aturan mayoritas bootstrap 50%.
Berdasarkan analisis kladistik pada Gambar 3.15. menggunakan software PAUP.4.b10 dengan bootstrap 1000 kali ulangan diperoleh hasil bahwa terdapat dua sub-clade monofiletik yang terbentuk signifikan dengan didukung nilai bootstrap > 50 %. P. bogorensis, dan P. convexa berada pada sub-clade pertama, sedangkan M. sintangense, M. lanchesteri, dan M. pilimanus berada pada sub-clade kedua. Hasil analisis PAUP.4.b10 selengkapnya disajikan pada Lampiran 7.
Metode maximum parsimony adalah metode yang memprediksikan pohon evolusi / evolutionary tree yang meminimalkan jumlah langkah yang dibutuhkan untuk menghasilkan variasi yang diamati dalam karakter morfologi atau sekuen.
Analisis bootstrap adalah metode yang menguji seberapa baik set data model. Selain itu, bootstrap didukung oleh sebagian besar paket software yang menguji cabang-cabang yang dapat dipercaya (Dharmayanti, 2011). Menurut Felsenstein (1985), cabang pohon filogeni ditolak jika nilai bootstrap di bawah 50%.
36
Berdasarkan analisis kladistik, kladogram yang terbentuk membutuhkan panjang langkah 48, dengan indeks konsistensi (CI)= 0,98 dan indeks retensi (RI)= 0,95 (lihat Lampiran 7). Perubahan karakter apomorfi seperti pada Gambar 3.16. dibawah ini mempunyai level homoplasi yang rendah.
Gambar 3.16. Kladogram dengan panjang langkah panjang langkah 48, indeks konsistensi (CI)= 0,98 dan indeks retensi (RI)= 0,95. Disertai dengan perubahan apomorfinya.
Panjang langkah adalah jumlah nomor perubahan keadaan karakter yang dibutuhkan untuk mendukung hubungan kekerabatan taksa pada sebuah pohon. Homoplasi dan nomor perubahan keadaan karakter yang lebih rendah dalam sebuah pohon sangat diharuskan. Oleh karena itu, pohon dengan panjang langkah yang lebih rendah lebih baik dari pada pohon dengan panjang langkah yang lebih tinggi. Pohon dengan panjang langkah terendah dianggap mempunyai homoplasi yang rendah dan lebih parsimonious (Lipscomb, 1998).
Indeks konsistensi dalam penelitian ini mendekati 1, hal ini berarti tingkat homoplasi pada kladogram yang terbentuk rendah. Menurut Lipscomb (1998), level homoplasi relatif dapat diukur dengan menggunakan indeks konsistensi (sering disingkat CI). Hal ini dihitung sebagai jumlah langkah yang diharapkan
37
memberikan jumlah penentuan karakter dalam data, yang dibagi dengan jumlah sebenarnya dari tahapan kemudian dikalikan dengan 100. Menurut Sanderson dan Donoghue; 1989 dalam Ariati et al., (2000), menyatakan bahwa suatu studi yang banyak menggunakan karakter ordered multistate cenderung mempunyai nilai CI yang rendah, karena karakter tersebut akan bertambah satu langkah lebih panjang (misal dari 1 ke 3).
Indeks retensi pada analisis penelitian ini yaitu 0,95. Nilai tersebut mendekati 1, hal ini berarti tingkat homoplasi pada kladogram yang terbentuk juga rendah. Menurut Lipscomb (1998), pengukuran lain mengenai level homoplasi yang relatif dibutuhkan untuk membuat sebuah pohon adalah indeks retensi (RI). Indeks retensi itu mengukur jumlah synapomorphy yang diharapkan dari sebuah kumpulan data yang disimpan sebagai synapomorphy pada sebuah kladogram.
Hasil CI dan RI tersebut sesuai dengan Sanderson dan Donoghue (1989), menyatakan bahwa nilai indeks konsistensi (CI) berkisar antara 0-1. Bila nilai CI mendekati atau sama dengan 1 berarti dalam kladogram tersebut homoplasinya sangat rendah atau tidak sama sekali, dan bila mendekati atau sama dengan 0 berarti homoplasinya sangat banyak. Sementara itu, fungsi nilai RI sama dengan CI yaitu untuk mengetahui level homoplasi, bahkan adanya nilai RI akan memperkuat informasi tentang adanya homoplasi dalam pohon filogeni (Lipscomb, 1998).
Ingroup (udang dan kepiting; ordo Decapoda) dan outgroup (ordo Isopoda) pada kladogram ini mempunyai nenek moyang yang sama yaitu Crustacea. Hal ini ditunjukkan dengan adanya persamaan morfologi diantara
38
ketiganya yaitu bagian tubuhnya terdiri atas 2 bagian pokok, yaitu cephalothorax (kepala dan dada yang menyatu), dan badan belakang/ perut (abdomen). Setiap ruas tubuhnya terdapat sepasang kaki. Pada bagian kepala dilindungi oleh kulit keras (karapas). Bagian kepala-dada terdapat sepasang antena, sepasang rahang atas (maxilla), dan sepasang rahang bawah (mandibula) (Anonim, 2011).
Isopoda yang digunakan sebagai outgroup pada penelitian ini jenis Pseudotyphloscia pallida Verhoeff (1928). ordo Isopoda sebagai outgroup species ordo Decapoda karena memiliki ciri primitif (plesiomorfi) yaitu ukuran pereiopoda yang seragam, pereiopoda berjumlah 14, dan karpus pada pereiopoda pertama sama panjangnya dengan merus. Karakter derivat dari ciri primitif tersebut kemudian menjadi karakter tersendiri bagi kelompok species ordo Decapoda dengan ciri memiliki ukuran pereiopoda yang tidak seragam (karakter no. 8), pereiopoda berjumlah 10 (karakter no. 9), dan panjang karpus pada pereiopoda pertama lebih pendek dari merus. Ciri tersebut merupakan sinapomorfi dari udang dan kepiting. Distribusi species dari ordo Isopoda ini ditemukan di Pulau Jawa, Sulawesi, Krakatau, Bali dan Taiwan (Green et al., 1990).
Sinapomorfi ciri udang dan kepiting membentuk dua cabang baru yang signifikan (didukung dengan nilai bootstrap > 50%). Cabang pohon pertama didukung dengan nilai bootstrap 100 % yang merupakan kelompok species kepiting familia Brachyura. Banyak karakter sinapomorfi yang dimiliki kelompok ini, diantaranya yaitu: tepi anterolateral karapas (karakter no. 5), bentuk karpus pereiopoda pertama (karakter no. 10), Ukuran chela pereiopoda pertama (karakter no. 12), panjang chela pereiopoda pertama (karakter no. 13), Ukuran kedua pereiopoda pertama (karakter no. 14), bentuk karpus pereiopoda kedua (karakter
39
no. 17), letak tuberkel pada chela (karakter no. 24), bentuk abdomen (karakter no. 26), adanya uropod (karakter no. 27), adanya telson (karakter no. 28).
Kelompok kepiting ini kemudian bercabang lagi menjadi species P. bogorensis dan P. convexa yang telah memiliki karakter derivat
masing-masing. P. bogorensis memiliki karapas yang tipis < ½ lebar karapas (karakter no. 4), dan duri pada pereiopoda pertama yang tumpul (karakter no. 6). P. convexa memiliki karapas yang tebal sekitar ½ dari lebar karapas (karakter no. 4), dan duri pereiopoda pertama tajam (karakter no. 6). P. bogorensis menjadi species yang lebih dulu muncul dibandingkan P. convexa, karena P. bogorensis memiliki kesamaan ciri dengan ciri-ciri yang dimiliki outgroup (Pseudotyphloscia pallida) yaitu merus pereiopoda pertama yang pada awalnya tidak berduri, kemudian pada P. convexa berubah menjadi berduri (karakter no. 7).
Cabang pohon kedua didukung dengan nilai bootstrap 88 % yang merupakan kelompok species udang familia Palaemonidae yang kladogramnya
menunjukkan bahwa familia Palaemonidae bersifat monofiletik dengan M. sintangense sebagai basal species (paling primitif) dan diakhiri dengan M. lanchesteri dan M. pilimanus sebagai species yang lebih maju (derived). Karakter bersama yang dimiliki oleh ketiga jenis udang tersebut yaitu tebal karpus = lebar karpus (karakter no. 4), karpus pereiopoda pertama subsilisdris (karakter no. 10), karpus pereiopoda pertama lebih panjang dari merus (karakter no. 11), chela pereiopoda pertama kecil (karakter no. 12), chela pereiopoda pertama lebih pendek dari karpus (karakter no. 13), chela pereiopoda kedua besar (karakter no. 15).
40
Terdapat banyak karakter yang sama-sama dimiliki oleh M. sintangense dan M. lanchesteri yang kemudian menjadi karakter yang lebih maju (apormorfi) pada M. pilimanus, diantaranya yaitu: ujung rostrum (karakter no. 2), post antenula (karakter no. 3), bentuk karpus pereiopoda kedua (karakter no. 17), panjang karpus pereiopoda kedua (karakter no. 18), chela pereiopoda kedua
(karakter no. 19), dan pubescence pada chela pereiopoda kedua (karakter no. 20). M. pilimanus dan M. lanchesteri berkerabat dekat karena sinapomorfi ciri dari
adanya preanal carina (karakter no. 25) dan dengan didukung nilai bootstrap 65 %.
Analisis ini mendukung hipotesis bahwa ordo ini berasal dari satu nenek moyang (monophyly), cabang pohon kelompok species kepiting merupakan sister group dari cabang pohon kelompok species udang. Berdasarkan Gambar 3.16. kladogram yang terbentuk dimulai dari percabangan pohon nomor 10, 9, 8, dan 7 dapat dikatakan bahwa, species ordo Decapoda yang mungkin pertama kali muncul sesuai evolusi atau runtutan waktu yaitu species udang yang diwakili oleh M. sintangense sebagai species yang paling primitif (basal species), yang kemudian diikuti dengan munculnya species yang lain yaitu M. lanchesteri dan M. pilimanus. Sejalan dengan berjalannya waktu, ada beberapa species udang yang berevolusi menjadi kepiting karena adanya proses reduksi dari anggota tubuhnya maka species tersebut menjadi tidak memiliki uropod (karakter no. 27), tidak memiliki telson (karakter no. 28) dan bentuk abdomen yang menekuk menutupi thoraks (karakter no. 26), yang kemudian diturunkan pada keturunannya. Species yang paling muda yaitu P. convexa.
41
Menurut Glaessner (1960), kecenderungan evolusi Decapoda adalah dari banyaknya lekukan pada garis melintang yang mereduksi yang terjadi melalui penggabungan atau penghilangan secara bertahap. Proses evolusi karapas Glypheocarida, Anomocarida, Brachyura and Palinura terjadi dengan mereduksinya bagian postorbital dan semakin memanjangnya rostrum dan perubahan bentuk lekukan pada gatis melintang, lambat laun mulai terbentuk duri hepatik dan gigi pada rostrum. Bagian cephalo-thorax menjadi rata pada bagian dorsoventral dan terjadi pelebaran pada karapas untuk kebutuhan ruang bagi insang dan otot.
Decapoda perenang memiliki pleopoda yang merupakan organ utama untuk bergerak. Selain itu, pleopoda juga digunakan sebagai tempat melekatnya telur pada Decapoda betina. Tidak megherankan hal tersebut menjadi mereduksi pada Decapoda reptantia. Mereduksinya pleopoda berpengaruh terhadap ukuran abdomen. Sebenarnya tidak sepenuhnya hilang, pada terminal somite (telson) masih terdapat sisanya dan bergabung pada bagian kecil (Glaessner, 1960).
42
IV. KESIMPULAN DAN SARAN
A. Kesimpulan
Berdasarkan hasil identifikasi, determinasi, verifikasi, analisis, dan pembahasan dapat disimpulkan bahwa:
1. Terdapat 5 species dari ordo Decapoda yang ditangkap di sungai Cijalu yaitu 2 species kepiting yang termasuk dalam genus Parathelphusa (Parathelphusa convexa dan P. bogorensis) dan 3 species udang yang
termasuk genus Macrobrachium (Macrobrachium pilimanus, M. sintangense, dan M. lanchesteri).
2. Hubungan filogenetik species anggota ordo Decapoda bersifat monofiletik dengan Macrobrachium sintangense sebagai species yang lebih primitif, sedangkan Parathelphusa convexa merupakan species yang paling maju.
B. Saran
Data yang lebih banyak sangat dibutuhkan untuk pembuktian hubungan filogenetik lebih lanjut, misalnya data molekuler maupun anatomi.