Pengujian Ketahanan Klon-klon Hasil Silangan Tanaman
Kentang Transgenik dengan Nontransgenik terhadap Penyakit
Hawar Daun Phytophthora infestans di Lapangan Uji Terbatas
Ambarwati, AD1), Herman, M1), Listanto, E1), Suryaningsih, E2), dan Sofiari, E2)
1)Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian, Jl. Tentara Pelajar No. 3A, Bogor 16111 2)Balai Penelitian Tanaman Sayuran, Jl. Tangkuban Parahu 517, Lembang, Bandung 40391
Naskah diterima tanggal 27 Desember 2011 dan disetujui untuk diterbitkan tanggal 26 Maret 2012
ABSTRAK. Tanaman kentang transgenik Katahdin event SP904 dan SP951 mengandung gen RB, yang diisolasi dari spesies liar
kentang diploid Solanum bulbocastanum. Gen RB mempunyai ketahanan yang bersifat durable dengan spektrum yang luas terhadap ras-ras Phytophthora infestans di Amerika Serikat. Dalam perakitan tanaman kentang tahan penyakit hawar daun P. infestans di Indonesia, transgenik Katahdin dijadikan sebagai donor tahan dalam persilangan dengan varietas rentan Atlantik dan Granola. Klon-klon hasil silangan dianalisis secara molekuler mengandung gen RB. Penelitian dilakukan untuk menguji ketahanan Klon-klon-Klon-klon hasil silangan tanaman kentang transgenik dengan nontransgenik terhadap isolat P. infestans di lapangan uji terbatas (LUT) yang berlokasi di Balai Penelitian Tanaman Sayuran (Balitsa), Lembang dari bulan Oktober 2009 sampai dengan Maret 2010. Klon-klon yang diuji ialah 12 klon hasil silangan Atlantik x transgenik Katahdin SP904 (A); 15 klon hasil silangan Atlantik x transgenik Katahdin SP951 (B); 17 klon hasil silangan Granola x transgenik Katahdin SP904 (C); dan 20 klon hasil silangan Granola x transgenik Katahdin SP951 (D). Atlantik dan Granola digunakan sebagai kontrol rentan, sedangkan transgenik Katahdin sebagai kontrol tahan. Pengamatan dimulai ketika muncul gejala awal, yaitu pada 26, 32, 39, 46, dan 53 hari setelah tanam. Ketahanan tanaman semakin menurun dengan bertambahnya periode pengamatan, diikuti meningkatnya intensitas penyakit dan AUDPC. Semua klon yang diuji menunjukkan keragaman dalam ketahanan fenotipik terhadap hawar daun P. infestans. Klon-klon hasil silangan Atlantik x transgenik Katahdin SP951 mempunyai nilai AUDPC 697, yang hampir sama dengan transgenik Katahdin SP904 yaitu 698,5. Klon-klon Granola x transgenik Katahdin SP951 mempunyai nilai AUDPC 687,5 lebih kecil dibandingkan transgenik Katahdin SP904. Hasil penelitian ini juga menunjukkan bahwa klon-klon tersebut mempunyai ketahanan yang lebih tinggi dibandingkan transgenik Katahdin SP904.Pada pengamatan 46 hari setelah tanam atau 20 hari setelah infeksi diperoleh tiga klon tahan yaitu B49 (skor 7,5), C111 (skor 7,1), dan D26 (skor 7,3). Ketahanan ini lebih tinggi daripada transgenik Katahdin SP904 (skor 5,1) dan transgenik Katahdin SP951 (skor 6,4).
Katakunci: Kentang transgenik; Persilangan; Gen RB; Hawar daun
ABSTRACT. Ambarwati, AD, Herman, M, Listanto, E, Suryaningsih, E and Sofiari, E 2012. Resistance Testing on Transgenic and Nontransgenic Potato Clones Against Late Blight Phytophthora infestans in Confined Field Trial. Transgenic
potato Katahdin event SP904 and SP951 containing RB gene, which were isolated from a wild diploid potato species, Solanum
bulbocastanum. RB gene showed durable resistance with broad spectrum to all known races of P. infestans in the USA. In
development of potato resistant to late blight P. infestans in Indonesia, Katahdin transgenic were used as a resistant donor and crossed with susceptible varieties i.e. Atlantic and Granola. Clones derived from the crossing were molecularly analyzed and had RB gene contain. Experiment was conducted to assess the resistance of the clones derived from crossing of Katahdin transgenic and nontransgenic to P. infestans in confined field trial (CFT), located at the Indonesian Vegetable Research Institute (IVEGRI), Lembang from October 2009 to March 2010. Several clones tested were 12 clones of Atlantic x Katahdin transgenic SP904 (A); 15 clones of Atlantic x Katahdin transgenic SP951 (B); 17 clones of Granola x Katahdin transgenic SP904 (C); and 20 clones of Granola x Katahdin transgenic SP951 (D). Atlantic and Granola were used as susceptible control whereas Katahdin transgenic as resistant control. Observation was started as late blight symptoms and detected at 26, 32, 39, 46, and 53 days after planting. Plant resistance decreases with increasing period of observation, followed by increasing disease intensity and AUDPC. All clones tested showed variation in phenotypic resistance to late blight P. infestans. Clones derived from crossing of Atlantic x Katahdin transgenic SP951 had AUDPC score 697 and almost similar to Katahdin transgenic SP904 (698.5). Clones derived from crossing of Granola x Katahdin transgenic SP951 had AUDPC score 687.5 and smaller than Katahdin transgenic SP904. The results also indicated that these clones had higher resistance than Katahdin transgenic SP904. Observation at 46 days after planting or 20 days after infection resulted three resistant clones i.e. B49 (score 7.5), C111 (score 7.1); and D26 (score 7.3). This resistance was higher than Katahdin transgenic SP904 (score 5.1) and Katahdin transgenic SP951 (score 6.4).
Keywords: Transgenic potato; Crossing; RB gene; Late blight
Penyakit hawar daun (late blight) pada tanaman kentang yang disebabkan oleh cendawan Phytophthora infestans dapat menyebabkan kehilangan hasil 50– 80%, bahkan mengakibatkan 100% gagal panen pada tanaman rentan, terutama apabila didukung oleh kondisi lingkungan dengan curah hujan dan kelembaban yang
tinggi (Ojiambo et al. 2000, Kusmana 2003). Sampai saat ini pengendalian penyakit hawar daun dilakukan secara intensif dengan penyemprotan fungisida, sehingga dapat membahayakan kesehatan manusia dan kelestarian lingkungan, serta memicu munculnya isolat-isolat tahan dan lebih virulen (Leonard-Schipper
biaya produksi yang lebih banyak untuk pembelian fungisida.
Salah satu cara yang paling efektif untuk melindungi tanaman dari epidemi penyakit hawar daun ialah dengan merakit varietas tahan. Program pemuliaan diarahkan untuk mendapatkan varietas dengan ketahanan lapangan, karena ketahanan ini bersifat lebih durable dibandingkan dengan ketahanan spesifik-ras. Gen ketahanan yang bersifat durable dan berspektrum luas terhadap ras-ras P. infestans terdapat pada spesies kentang liar diploid Solanum bulbocastanum (2n = 2x = 24) yang disebut sebagai gen RB (Resistance dari Bulbocastanum) (Naess et al. 2000, Song et al. 2003). Gen RB diintroduksikan ke dalam varietas kentang tetraploid Katahdin (2n = 4x = 48) menggunakan teknik Agrobacterium tumefaciens (Song et al. 2003) dan menghasilkan beberapa transforman, antara lain ialah event SP904 dan SP951. Tanaman transgenik Katahdin tersebut telah diuji di lapangan uji terbatas (LUT) di Michigan Agricultural Experimental Station (Kuhl et al. 2007) serta di Minnesota (Bradeen et al. 2009), dan menunjukkan ketahanan yang berspektrum luas terhadap ras-ras P. infestans.
Tanaman transgenik Katahdin RB yang tahan terhadap hawar daun P. infestans dapat digunakan sebagai sumber ketahanan untuk mendukung pemuliaan tanaman. Persilangan antara tanaman transgenik Katahdin SP904 maupun SP951 dengan varietas rentan, seperti Atlantik maupun Granola telah dilakukan untuk mendapatkan sifat ketahanan terhadap penyakit hawar daun. Atlantik dan Granola ialah varietas kentang yang banyak diminati oleh petani dan mendominasi areal pertanaman kentang di Indonesia, tetapi mempunyai kelemahan yaitu rentan terhadap penyakit hawar daun (Kusmana 2004, Basuki et al. 2005).
Klon-klon hasil silangan diuji secara molekuler untuk melihat integrasi gen RB dan segregasinya (Ambarwati et al. 2009). Pengujian ekspresi gen RB untuk melihat efikasinya terhadap P. infestans dapat dilakukan di fasilitas uji terbatas (FUT) maupun di lapangan uji terbatas (LUT). Penanaman di LUT harus memenuhi beberapa persyaratan, seperti pembatasan atau pengamanan genetik untuk mencegah pemindahan gen, dengan memperhatikan isolasi waktu, isolasi biologis, dan isolasi fisik (Herman 1999). Pengujian ketahanan klon-klon hasil silangan yang mengandung gen RB dilakukan di LUT di Pasir Sarongge, Cianjur, Jawa Barat pada tahun 2008 (Ambarwati et al. 2011). Untuk mengetahui apakah gen RB juga berspektrum luas terhadap isolat-isolat P. infestans di Indonesia, perlu dilakukan penelitian di LUT pada berbagai lokasi.
klon hasil silangan antara tanaman transgenik Katahdin SP904 atau SP951 dengan tanaman nontransgenik Atlantik atau Granola, yang telah mengandung gen RB, terhadap isolat P. infestans di LUT di Lembang. Hipotesis penelitian ialah klon-klon kentang transgenik yang berbeda menunjukkan keragaman ketahanan terhadap P. infestans.
BAHAN DAN METODE
Penelitian dilakukan di LUT, yang berlokasi di Balai Penelitian Tanaman Sayuran (Balitsa), Lembang dengan ketinggian 1.250 m dpl., dari bulan Oktober 2009 sampai Maret 2010.
Bahan Tanaman
Bahan tanaman yang dievaluasi sebanyak 64 klon, terdiri atas 12 klon hasil silangan Atlantik x transgenik Katahdin SP904 (A); 15 klon hasil silangan Atlantik x transgenik Katahdin SP951 (B); 17 klon hasil silangan Granola x transgenik Katahdin SP904 (C); dan 20 klon hasil silangan Granola x transgenik Katahdin SP951 (D). Klon-klon ini positif mengandung gen RB melalui analisis PCR. Sebagai kontrol atau pembanding disertakan tetua rentan (Granola dan Atlantik), tetua tahan (transgenik Katahdin SP904 dan SP951), Katahdin nontransgenik, dan spesies liar S. bulbocasatum PT29.
Penanaman di LUT
Penanaman di LUT menggunakan rancangan acak kelompok dengan tiga ulangan. Jarak tanam 30 cm dalam baris dan jarak antarbaris 80 cm. Tanaman border sebagai sumber inokulum yaitu Granola, Atlantik, dan nontransgenik Katahdin ditanam mengelilingi plot percobaan, dan setiap empat klon dalam plot diselingi tanaman border. Tiap klon terdiri atas lima tanaman. Tanaman kentang diekspos pada kondisi alamiahnya dan tidak diberikan inokulasi penyakit buatan. Pemupukan dengan pupuk kandang 30 ha, NPK (15–15–15) 800 kg/ha, dengan dosis ¾ diberikan pada saat tanam dan ¼ bagian pada saat tanaman berumur 30 hari. Pemeliharaan terhadap organisme pengganggu tumbuhan (OPT) dilakukan dengan penyemprotan insektisida dan tidak ada penyemprotan fungisida. Sepanjang percobaan berlangsung tidak ada tanaman sejenis yang ditanam di lapangan (isolasi biologis) dan LUT dikelilingi oleh pagar besi (isolasi fisik).
Evaluasi Ketahanan Tanaman
Pengamatan terhadap serangan penyakit hawar daun P. infestans dilakukan setelah muncul gejala
awal, yaitu dimulai dari 26 hari setelah tanam (HST) yang dianggap sebagai hari kesatu setelah infeksi (HSI) kemudian 32, 39, 46, dan 53 HST. Peubah yang diamati yaitu:
a. Skor ketahanan tanaman (skala 0–9) menggunakan acuan dari Henfling (1979) dan Halterman et al. (2008) berdasarkan persentase daun yang terserang, yaitu: 0 (100%), 1 (> 90%), 2 (81–90%), 3 (71– 60%), 4 (61–70%), 5 (41– 60%), 6 (26 – 40%), 7 (11–25%), 8 (<10%), dan 9 (0%).
b. Tingkat ketahanan tanaman ditentukan berdasarkan skor yang dimiliki tiap tanaman uji, di mana skor ≥ 7 (≤ 25% infeksi) termasuk ke dalam kategori tahan (Song et al. 2003, Colton et al. 2006, Halterman et al. 2008) dan skor ≤6,9 (>25% infeksi) termasuk kategori rentan (Song et al. 2003).
c. Intensitas penyakit (%) dihitung menggunakan rumus:
di mana:
IP = Intensitas penyakit;
n = Jumlah tanaman dari tiap kategori serangan; v = Nilai skala dari tiap kategori serangan (0-9); N = Jumlah tanaman contoh;
Z = Skor dari kategori serangan tertinggi.
d. Kumulatif serangan hawar daun dihitung menggunakan metode AUDPC (area under diseases progress curve), menurut Landeo (1999) yaitu:
di mana :
Xt = Persentase serangan penyakit
hawar daun, pengamatan pada waktu ke t;
Xt+1 = Persentase serangan penyakit
hawar daun pada pengamatan t +1 pengamatan berikutnya;
(Dt+1–Dt) = interval pengamatan dari
p e n g a m a t a n p e r t a m a k e pengamatan kedua. Semakin tinggi nilai AUDPC suatu klon menunjukkan semakin rentan terhadap penyakit.
e. Analisis pengelompokan (hierarchical clusters) klon-klon hasil silangan menggunakan program SPSS versi 13.
HASIL DAN PEMBAHASAN
Pengamatan serangan penyakit hawar daun pada klon-klon hasil silangan dimulai ketika muncul gejala awal yaitu pada 26 HST. Daun yang terserang P. infestans menunjukkan adanya bercak basah, yang kemudian meluas ke permukaan daun. Tampak masa sporangia berwarna putih pada permukaan daun bagian bawah dan segera seluruh daun yang terinfeksi menjadi layu dan mati.
Tingkat serangan penyakit hawar daun untuk berbagai periode pengamatan ditampilkan pada Tabel 1. Semua klon yang diuji (64 klon) dan tetua yang digunakan dalam persilangan (Atlantik, Granola, transgenik Katahdin SP904, dan SP951) serta Katahdin nontransgenik dan S. bulbocastanum PT29 pada pengamatan 26 HST menunjukkan skor ketahanan berkisar dari 8,5–9. Namun demikian, pada 32 HST atau 6 hari setelah infeksi (HSI) Atlantik dan Granola sudah terserang hawar daun dengan intensitas penyakit >25% dan dikategorikan sebagai rentan. Sampai 39 HST (13 HSI), rerata klon silangan mempunyai skor 7,1 dengan intensitas penyakit 21,2%, sedangkan tetua transgenik Katahdin SP904 dan SP951 masing-masing dengan skor 7,5 dan 7,6. Dapat dikatakan bahwa tanaman masih menunjukkan kategori tahan, sesuai dengan kriteria dari Song et al. (2003), Colton et al. (2006), dan Halterman et al. (2008), yaitu skor ≥7 atau intensitas penyakit ≤25% termasuk dalam kategori
tahan.Tanaman yang sudah terserang penyakit hawar
daun dapat menjadi sumber inokulum baru bagi tanaman sehat lainnya. Ketahanan tanaman semakin berkurang, yang ditandai dengan menurunnya skor ketahanan, dan intensitas penyakit >25%, sehingga dapat dikatakan bahwa tanaman sudah menunjukkan tingkat kategori rentan pada 46 HST. Penurunan tingkat ketahanan dan meningkatnya intensitas penyakit semakin nyata pada akhir pengamatan yaitu pada 53 HST.
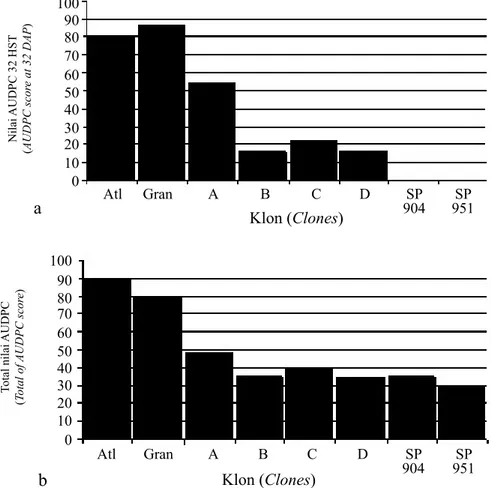
Menurunnya ketahanan diikuti dengan semakin tingginya intensitas penyakit dan nilai AUDPC. Pada 32 HST (Gambar 1a), klon-klon hasil silangan Atlantik x transgenik Katahdin SP904 (A), Atlantik x transgenik Katahdin SP951 (B), Granola x transgenik Katahdin SP904 (C), dan Granola x transgenik Katahdin SP951 (D) mempunyai nilai AUDPC, masing-masing 53,9; 15,9; 22; dan 16,5. Transgenik Katahdin SP904 dan SP951 belum terserang penyakit hawar daun, sementara Atlantik dan Granola menunjukkan nilai AUDPC masing-masing sebesar 79,2 dan 86,5. Nilai AUDPC klon-klon silangan ini lebih kecil dari Atlantik dan Granola tetapi lebih besar daripada transgenik Katahdin SP904 dan SP951. Hal ini menunjukkan bahwa secara umum, klon-klon hasil silangan mempunyai ketahanan yang lebih tinggi Σ n x v IP = x 100% N x Z n AUDPC = Σ (Xt+1 + Xt ) (Dt+1 – Dt) i=1
dibandingkan tetua rentan Atlantik dan Granola, tetapi lebih rendah daripada tetua tahan transgenik Katahdin SP904 dan SP951. Hasil yang sama juga diperoleh pada penelitian LUT klon-klon kentang hasil silangan Atlantik atau Granola dengan transgenik Katahdin SP904 atau SP951 di Pasir Sarongge, Cianjur, Jawa Barat (Ambarwati et al. 2011).
Nilai kumulatif AUDPC dari 32 sampai 53 HST (Gambar 1b), secara umum menunjukkan kecenderungan yang sama dengan nilai AUDPC pada 32 HST. Klon-klon hasil silangan Atlantik x transgenik Katahdin SP951 (B) mempunyai nilai AUDPC 697, hampir sama dengan transgenik Katahdin SP904 yaitu 698,5. Berbeda dengan klon-klon Granola x transgenik Katahdin SP951 (D) mempunyai nilai AUDPC 687,5 lebih kecil dibandingkan transgenik Katahdin SP904. Hal ini menunjukkan bahwa klon-klon tersebut mempunyai ketahanan yang lebih tinggi dibandingkan transgenik Katahdin SP904.
Analisis pengelompokan (hierarchical clusters) klon-klon hasil silangan berdasarkan skor ketahanan pada 46 HST disajikan pada Gambar 2. Terdapat tiga kelompok, yaitu kelompok I untuk klon dengan skor ketahanan 5,6-8,4, kelompok II dan III masing-masing untuk klon dengan skor ketahanan 0,3–3,5 dan 3,8–5,2.
Kelompok I meliputi dua varietas kontrol (transgenik Katahdin SP951 dan S. bulbocastanum) serta 21 klon, yang terdiri dari enam klon (40%) hasil silangan Atlantik x transgenik Katahdin SP951 (B), lima klon (31,3%) hasil silangan Granola x transgenik Katahdin SP904 (C), dan 10 klon (50%) hasil silangan Granola x transgenik Katahdin SP951 (D), sedangkan klon hasil silangan Atlantik x transgenik Katahdin SP904 (A) tidak ada yang masuk kategori kelompok I. Dari 21
nontransgenik di LUT Lembang(Level of late blight P. infestans infection on clones from crosses between transgenic and nontransgenic potato in CFT, Lembang)
Periode pengamatan (Observation period) Infeksi pertama (First infection) 26 HST (DAP) 32 HST (DAP)
6 HSI (DAI) 39 HST (DAP) 13 HSI (DAI) 46 HST (DAP) 20 HSI (DAI) 53 HST (DAP) 27 HSI (DAI)
SK IP (%) SK IP (%) SK IP (%) SK IP (%) SK IP (%) Klon-klon silangan 8,9 1,6 8,3 7,4 7,1 21,2 4,6 48,4 2,4 73,3 Atlantik 8,9 1,2 6,7 25,2 1,1 83,0 0,0 100,0 0,0 100,0 Granola 8,8 2,2 6,6 26,6 3,0 66,6 1,2 86,7 0,6 93,3 Katahdin 8,5 5,2 7,1 20,7 1,5 83,0 0,3 96,7 0,3 96,7 SP904 9,0 0,0 9,0 0,0 7,5 16,1 5,1 43,7 1,8 80,0 SP951 9,0 0,0 9,0 0,0 7,6 15,6 6,4 28,9 1,9 79,3 S. bulbocastanum 9,0 0,0 8,9 1,5 8,8 2,2 8,4 6,7 7,8 13,3
Keterangan (Remarks) SK: skor ketahanan (resistance score), IP: intensitas penyakit (disease intensity), HST : hari setelah tanam (days
after planting), HSI: hari setelah infeksi (days after infection)
klon tersebut, ada tiga klon yang dikategorikan sebagai tahan, yaitu klon C111, B49, dan D26 dengan skor ketahanan masing-masing 7,1; 7,5; dan 7,3. Kelompok II meliputi 15 klon, terdiri dari lima klon A (41,7%), empat klon B (26,7%), empat klon C (25%), dan dua klon D (10%). Dalam kelompok ini juga termasuk tetua rentan Atlantik dan Granola serta Katahdin nontransgenik, sedangkan kelompok III terdiri dari 28 klon, yaitu tujuh klon A (58,3%), lima klon B (33,3%), delapan klon C (43,8%), dan delapan klon D (40%), serta tetua transgenik Katahdin SP904.
Dari semua klon yang dievaluasi, klon hasil silangan Atlantik x transgenik Katahdin SP904 menyebar dalam kelompok II dan III, klon hasil silangan Atlantik x transgenik Katahdin SP951 dan Granola x transgenik Katahdin SP951, paling banyak terdapat pada kelompok I, diikuti kelompok III dan kelompok II, sementara klon hasil silangan Granola x transgenik Katahdin SP904 paling banyak menyebar di kelompok III, diikuti kelompok I dan kelompok II. Secara umum, klon-klon hasil silangan dengan transgenik Katahdin SP951 mempunyai ketahanan yang lebih tinggi dibandingkan klon-klon hasil silangan dengan transgenik Katahdin SP904. Persilangan yang melibatkan transgenik Katahdin SP904 dengan tetua Granola menunjukkan ketahanan lebih tinggi dibandingkan dengan tetua Atlantik. Hasil yang diperoleh dalam penelitian ini, didukung oleh hasil LUT tahun 2008 di Pasir Sarongge, Cianjur, Jawa Barat, di mana klon-klon hasil silangan Atlantik dengan transgenik Katahdin SP904 lebih banyak menyebar pada kelompok II, sedangkan 41% klon dari persilangan dengan tetua transgenik Katahdin SP951 berada pada kelompok III (Ambarwati 2010). Dikaitkan dengan total nilai AUDPC (Gambar 1b) dapat dilihat bahwa Atlantik mempunyai nilai kumulatif serangan yang lebih tinggi dibandingkan
Gambar 1. Nilai area under diseases progress curve (AUDPC) a) pada pengamatan 32 HST dan b) total, Atl: Atlantik, Gran: Granola, A: Atlantik x transgenik Katahdin SP904, B: Atlantik x transgenik Katahdin SP951, C: Granola x transgenik Katahdin SP904, D: Granola x transgenik Katahdin SP951, SP904: transgenik Katahdin SP904, SP951: transgenik Katahdin SP951 (AUDPC) a) at 32 DAP and b) total. Atl: Atlantic, Gran: Granola, A: Atlantic x transgenic Katahdin SP904, B: Atlantic x transgenic Katahdin SP951, C: Granola x transgenic Katahdin SP904, D: Granola x transgenic Katahdin SP951, SP904: transgenic Katahdin SP904, SP951: transgenic Katahdin SP951)
Granola, demikian juga dengan transgenik Katahdin SP904 dengan nilai kumulatif yang lebih tinggi daripada transgenik Katahdin SP951. Menurut laporan evaluasi tanaman kentang transgenik di LUT Lembang (ABSP II 2007), transgenik Katahdin SP951 lebih tahan daripada Katahdin SP904 dan Granola juga lebih tahan dibandingkan Atlantik, berdasarkan intensitas serangan yang diamati pada 28 sampai 50 HST.
Pengamatan tingkat ketahanan untuk keempat populasi klon hasil silangan ditampilkan pada Tabel 2. Klon-klon dari keempat silangan yaitu Atlantik x transgenik Katahdin SP904 (A), Atlantik x transgenik Katahdin SP951 (B), Granola x transgenik Katahdin SP904 (C), maupun Granola x transgenik Katahdin SP951 (D), masih menunjukkan tingkat ketahanan dengan kategori tahan pada pengamatan pertama yaitu 26 HST. Pada saat Atlantik dan Granola menunjukkan
kategori rentan dengan skor ketahanan masing-masing 6,7 dan 6,6 pada pengamatan 32 HST, sebanyak 75–95% klon-klon hasil silangan masih tahan. Pada 39 HST, klon-klon hasil silangan lebih banyak menunjukkan kategori tahan, yaitu sebesar 86,7; 58,8; dan 70% masing-masing untuk klon B, C, dan D, sedangkan klon A sudah banyak yang rentan (66,7%). Pada 46 HST, terdapat tiga klon yang masih tahan, yaitu klon B49 (silangan Atlantik x transgenik Katahdin SP951) dengan skor ketahanan 7,5, klon C111 (silangan Granola x transgenik Katahdin SP904) dengan skor ketahanan 7,1, dan klon D26 (silangan Granola x transgenik Katahdin SP951) dengan skor ketahanan 7,3. Pada hari yang sama Atlantik, Granola, nontransgenik Katahdin, dan bahkan transgenik Katahdin SP904 dan SP951 menunjukkan katagori rentan (Tabel 1). Solanum bulbocastanum masih menunjukkan kategori tahan sampai pengamatan terakhir (53 HST). Ketiga klon yang tahan di LUT
0 90 100 80 70 60 50 40 30 20 10 Atl SP 904 D C B A Gran
a
951SP Nilai AUDPC 32 HST (AUDPC scor e at 32 DAP ) Klon (Clones) 0 90 100 80 70 60 50 40 30 20 10 Atl SP 904 D C B A Granb
SP 951 Total nilai AUDPC (T otal of AUDPC scor e) Klon (Clones)Gambar 2. Dendrogram pengelompokan klon-klon hasil silangan berdasarkan skor ketahanan pada 46 HST, A: Atlantik x transgenik Katahdin SP904, B: Atlantik x transgenik Katahdin SP951, C: Granola x transgenik Katahdin SP904, D: Granola x transgenik Katahdin SP951 (Clustering dendrogram of clones of crosses based on resistance score at 46 DAP A: Atlantic x transgenic Katahdin SP904, B: Atlantic x transgenic Katahdin SP951, C: Granola x transgenic Katahdin SP904, D: Granola x transgenic Katahdin SP951) SP904 SP951 C111 B49 D26 S. bulbo Atl Gran Kat 44. D78 44 68. SP951 68 14. B63 14 50. D1 50 58. D38 58 57. D48 57 62. D37 62 15. B143 15 36. C75 36 55. D25 55 17. B35 17 51. D82 51 63. D15 63 28. C208 28 53. D8 53 22. B59 22 33. C137 33 42. C140 42 16. B128 16 49. D26 49 43. C111 43 19. B49 19 70. S.bul 70 11. A135 11 9. A86 9 66. Atl 66 65. Gran 65 10. A94 10 69. Kat 69 32. C157 32 27. B66 27 8. A12 8 35. C181 35 21. B60 60 59. D63 59 34. C71 34 61. D207 61 12. A252 12 26. B162 26 6. A135 6 1. A27 1 18. B64 18 41. C106 41 45. D80 45 13. B76 13 39. C108 39 2. A242 2 5. A91 5 20. B34 20 56. D73 56 64. C105 64 30.C183 30 48. D7 48 60. D32 60 67. SP904 67 4. 4222 4 46. D89 46 54. D12 54 38. C193 38 40. C113 40 25. B11 25 31. C159 31 23. B169 23 24. B163 24 37. C133 37 52. D76 52 29. C186 29 7. A63 7 3. A16 3 47. D88 47 I II III
Lembang tersebut, juga menunjukkan ketahanan pada penelitian LUT tahun 2008 di Pasir Sarongge, Cianjur, Jawa Barat, di mana pada pengamatan ketiga (63 HST atau 7 HSI) klon B49 mempunyai skor 6,8, klon C111 dan D26 masing-masing dengan skor 8,7 dan 7,7.
Menurunnya ketahanan tanaman juga berhubungan dengan bertambahnya umur fisiologis tanaman. Menurut penelitian Millett & Bradeen (2007) serta Millett et al. (2009), pada umur 4 minggu atau periode prapembungaan, tanaman kentang dengan gen RB menunjukkan tingkat ketahanan paling tinggi. Umur 8 minggu atau setelah periode berbunga selesai dan memasuki tahap awal pembentukan umbi, ketahanan tanaman mulai berkurang, sampai tanaman mengalami penuaan atau berumur ± 12 minggu.
Penampilan respons ketahanan klon-klon hasil silangan terhadap P. infestans disajikan pada Gambar 3.
Pada 32-34 HST terlihat bahwa Atlantik dan Granola sudah termasuk kategori rentan dengan skor 6,6 dan 6,7 (Gambar 3B, C). Klon-klon yang termasuk kategori rentan seperti A86 dan B34 ditampilkan pada Gambar 3H dan 3I, dengan intensitas penyakit masing-masing sebesar 74,8 dan 95,6% pada akhir pengamatan (53-56 HST). Klon-klon ini merupakan hasil silangan antara Atlantik dengan transgenik Katahdin SP904 dan Atlantik x transgenik Katahdin SP951. Berbeda dengan klon tahan seperti C111 dengan skor 7,1 masih tampak hijau bahkan setelah berumur 83 HST (Gambar 3G).
Klon-klon hasil silangan menunjukkan keragaman ketahanan terhadap hawar daun P. infestans. Gen RB berspektrum luas terhadap ras-ras P. infestans dengan ketahanan horisontal (Song et al. 2003), tetapi tingkat ketahanan atau ekspresi gen yang dihasilkan tidak terlalu tinggi. Klon yang termasuk kategori tahan
Gambar 3. Respons ketahanan klon-klon hasil silangan di LUT Lembang (A) Border Atlantik, (B) Atlantik (32 HST), (C) Granola (32 HST), (D) transgenik Katahdin SP951 (53 HST), (E) S. bulbocasta-num PT29 (83 HST), (F) Klon B63 (Atlantik x transgenik Katahdin SP951 (53 HST), (G) klon C111 (Granola x transgenik Katahdin SP904 (83 HST), (H) Klon A86 (Atlantik x transgenik Katahdin SP904 (53 HST), (I) klon B34 (Atlantik x transgenik Katahdin SP951(53 HST) (Resis-tance response of clones of crosses in CFT Lembang. (A) Border Atlantic, (B) Atlantic (32 DAP), (C) Granola (32 DAP), (D) Katahdin transgenic SP951(53 DAP), (E) S. bulbocastanum PT29 (83 DAP), (F) Clone B63 (Atlantic x Katahdin transgenic SP951 (53 DAP), (G) Clone C111 (Granola x Katahdin transgenic SP904 (83 DAP), (H) Clone A86 (Atlantic x Katahdin transgenic SP904 (53 DAP), (I) Clone B34 (Atlantic x Katahdin transgenic SP951(53 DAP)
A B C
D E F
J. Hort. V ol. 22 No. 2, 2012 194 Tabel 2. T ingkat ketahanan klon-klon hasil silangan kentang transgenik dengan nontransgenik terhadap penyakit hawar daun P. infestans di LUT , Lem bang (
Resistance level on clones of crosses between transgenic and nontransgenic potato to late blight P
. infestans in CFT Lembang)
Periode
pengamatan
(Observation period)
Klon-klon hasil silangan (Clones of crosses)
A 1 B 2 C 3 D 4 Klon tahan (Resistant clones) Klon r entan (Susceptible clones) Klon tahan (Resistant clones) Klon r entan (Susceptible clones)
Klon tahan (Resistant clones)
Klon r
entan
(Susceptible clones) Klon tahan (Resistant clones)
Klon r
entan
(Susceptible clones)
26 HST
(DAP)
(Infeksi pertama) ( First infection)
12 (100) 0 (0) 15 (100) 0 (0) 17 (100) 0 (0) 20 (100) 0 (0) 32 HST (DAP ) 5 6 HSI (DAI) 9 (75) 3 (25) 14 (93,3) 1 (6,7) 16 (94,1) 1 (5,9) 19 (95) 1 (5) 39 HST (DAP ) 13 HSI (DAI) 4 (33,3) 8 (66,7) 13 (86,7) 2 (13,3) 10 (58,8) 7 (41,2) 14 (70) 6 (30) 46 HST (DAP ) 20 HSI (DAI) 0 (0) 12 (100) 1 (6,7) 14 (93,3) 1 (5,9) 16 (94,1) 1 (5) 19 (95) 53 HST (DAP ) 27 HSI (DAI) 0 (0) 12 (100) 0 (0) 15 (100) 0 (0) 17 (100) 0 (0) 20 (100) Keterangan (Note ): A 1 : Atlantik x transgenik Katahdin SP904, B 2 : Atlantik x transgenik Katahdin SP951, C 3: Granola x transgenik Katahdin SP904, D 4 : Granola x transgenik Katahdin SP951, 5 HST
hari setelah tanam (
days after planting),
HSI: hari setelah infeksi
(days after infection
), angka dalam tanda kurung menyatakan persentase (
numbers in par
entheses expr
ess per
yaitu B49, C111, dan D26 mempunyai skor ketahanan 7,1–7,5 yang berarti tidak bersifat imun dan masih terjadi infeksi patogen sebesar ≤ 25%. Berbeda dengan gen-gen R pada S. demissum, ketahanan yang dimediasi oleh gen RB tidak menunjukkan reaksi hipersensitif dan tidak bersifat mengeliminasi patogen, tetapi menunda dan membatasi perkembangan serta penyebaran patogen (Bradeen et al. 2009).
Hasil penelitian yang diperoleh, sesuai dengan Bradeen et al. (2009) yang mendapatkan bahwa di antara 57 klon transgenik RB yang dievaluasi, 13 (23%) klon termasuk kategori tahan dan 26 (46%) moderat tahan, pada pengamatan 30 HSI di lapangan. Penelitian yang dilakukan oleh Kramer et al. (2009), Millett et al. (2009), dan Bradeen et al. (2009) mendapatkan bahwa klon-klon transgenik RB yang berbeda, menunjukkan perbedaan tingkat ketahanan terhadap hawar daun, yang diduga berkorelasi dengan banyaknya transkripsi gen RB.
Adanya klon-klon yang lebih tahan dari Atlantik, Granola, maupun transgenik Katahdin sangat bermanfaat, khususnya dalam mengurangi penyemprotan fungisida. Atlantik dan Granola terserang hawar daun 25 – 26% pada 32 HST atau 6 HSI, sehingga harus dilakukan penyemprotan fungisida sebelum 32 HST, sementara ada tiga klon hasil silangan yang masih tahan sampai 46 HST atau 20 HSI. Hasil studi sosial ekonomi (ex ante) yang dilakukan oleh Adiyoga (2009) mendapatkan bahwa dalam mengendalikan P. infestans petani menyemprot fungisida sebanyak 20-30 kali per musim tanam, terutama pada musim penghujan. Oleh karena itu mengetahui waktu terjadinya infeksi dapat mengurangi frekuensi penyemprotan fungisida.
KESIMPULAN
1. Semua klon yang diuji yaitu 12 klon hasil silangan Atlantik x transgenik Katahdin SP904 (A), 15 klon hasil silangan Atlantik x transgenik Katahdin SP951 (B), 17 klon hasil silangan Granola x transgenik Katahdin SP904 dan 20 klon hasil silangan Granola x transgenik Katahdin SP951 menunjukkan keragaman dalam ketahanan fenotipik terhadap hawar daun P. infestans di LUT di Lembang. 2. Klon-klon hasil silangan Atlantik x transgenik
Katahdin SP951 mempunyai nilai AUDPC 697, yang hampir sama dengan transgenik Katahdin SP904 yaitu 698,5. Sementara klon-klon Granola x transgenik Katahdin SP951 mempunyai nilai AUDPC 687,5 lebih kecil dibandingkan transgenik Katahdin SP904. Hal ini menunjukkan bahwa
klon-klon tersebut mempunyai ketahanan yang lebih tinggi dibandingkan transgenik Katahdin SP904. 3. Pada pengamatan 46 HST atau 20 HSI diperoleh
tiga klon tahan yaitu B49 (skor 7,5), C111 (skor 7,1), dan D26 (skor 7,3). Ketahanan ini lebih tinggi daripada transgenik Katahdin SP904 (skor 5,1) dan transgenik Katahdin SP951 (skor 6,4).
UCAPAN TERIMA KASIH
Penulis mengucapkan banyak terima kasih kepada Ibu Dedeh Suwarsih dan Bapak Usep di Balai Penelitian Tanaman Sayuran, Lembang, serta Ibu Sukma Wijayanti di BB – Biogen, Bogor, yang telah banyak membantu dalam pelaksanaan penelitian ini.
PUSTAKA
1. Agricultural Biotechnology Support Project II, 2007, Report on
identification of potato resistant to late blight (Phytophthora infestans Mt.de Bary), The Development of Late Blight
Resistant Potato.
2. Adiyoga, W 2009, Costs and benefits of transgenic late blight resistant potatoes in Indonesia, in Norton, GW & Desiree, MH (eds.). Projected Impacts of Agricultural Biotechnologies for
Fruits and Vegetables in the Philippines and Indonesia, ISAAA SEAsia Center, Los Banos Laguna 4030, Philippines, pp.
86-104.
3. Ambarwati, AD, Purwito, A, Herman, M, Sumaraw, SM & Aswidinnoor, H 2009, ‘Analisis integrasi dan segregasi gen ketahanan terhadap hawar daun pada progeni F1 hasil persilangan tanaman kentang transgenik dengan non transgenik’, J. AgroBiogen., vol. 5, no. 1, hlm. 25-31. 4. Ambarwati, AD 2010, ‘Pemanfaatan gen RB dalam
pengembangan tanaman kentang tahan penyakit hawar daun (Phytophthora infestans)’, disertasi, Institut Pertanian Bogor, Bogor.
5. Ambarwati, AD, Herman, M, Purwito, A, Sumaraw, SM & Aswidinnoor, H 2011, ‘Resistance evaluation on populations of crosses between transgenic potato Katahdin RB and non-transgenik Atlantic and Granola to late blight (Phytophthora
infestans) in confined field trial’, Indonesian J. Agric. Sci., vol
2, no. 1, pp. 33-9.
6. Basuki, RS, Kusmana & Dimyati, A 2005, ‘Analisis daya hasil, mutu, dan respons pengguna terhadap klon 380584.3, TS-2, FBA-4, I-1085, dan MF-II sebagai bahan baku keripik kentang’, J. Hort, vol. 15, no. 3, hlm. 160-70.
7. Bradeen, JM, Iorizzo, M, Mollov, DS, Raasch, J, Kramer, LC, Millet, BP, Austin-Phillips, S, Jiang, J & Carputo, D 2009, ‘Higher copy numbers of the potato RB transgene correspond to enhanced transcript and late blight resistance levels’, Molecular
Plant-Microbe Interactions, vol. 22, no. 4, pp. 437-46.
8. Colton, LM, Groza, HI, Wielgus, SM & Jiang, J 2006, ‘Marker-assisted selection for the broad-spectrum potato late blight resistance conferred by gene RB derived from a wild potato species’, Crop Sci., vol. 46, pp. 589-94.
9. Halterman, DA, Kramer, LC, Wielgus, S & Jiang, J 2008, ‘Performance of transgenic potato containing the late blight
infestans’, In Tech Information Bull, International Potato
Center, Lima, Peru, PP. 13.
11 Herman, M 1999, ‘Tanaman hasil rekayasa genetik dan pengaturan keamanannya di Indonesia’, Bul. AgroBio., vol. 3, no. 1, hlm. 8-26.
12. Kuhl, JC, Zarka, K, Coombs, J, Kirk, WW & Douches, DS 2007, ‘Late blight resistance of RB transgenic potato lines’, J.
Amer. Soc. Hort. Sci., vol. 132, no. 6, pp. 783-89.
13. Kusmana 2003, ‘Evaluasi beberapa klon kentang asal stek batang untuk uji ketahanan terhadap Phytophthora infestans’,
J. Hort., vol. 13, no. 4, hlm. 220-28.
14. Kusmana 2004, ‘Evaluasi resistensi 26 genotip kentang terhadap penyakit busuk daun di Cibodas, Lembang’, J. Hort., vol. 14, no. 1, hlm. 15-24.
15. Kramer, LC, Choudoir, MJ, Wielgus, SM, Bhaskar, PB & Jiang, J 2009, Correlation between transcript abundance of the RB gene and the level of the RB-mediated late blight resistance in potato, MPMI, vol. 22, no. 4, pp. 447-55.
16. Landeo, JA 1999, Data processing and interpretation of
resistance parameters, CIP, PERU.
17. Leonards-Schippers C, Gieffers, W, Schafer-Pregl, R, Ritter, E, Knapp, SJ, Salamini, F & Gebhardt, C 1994, ‘Quantitative resistance to Phytophthora infestans in potato: a case study for QTL mapping in an allogamous plant species’, Genetics, vol. 137, pp. 67-77.
specific PCR and RT-PCR assays for clustered resistance genes using a potato late blight resistance as a model’, Theor. Appl.
Genet., vol. 114, pp. 501-13.
19. Millett, BP, Mollov, DS, Iorizzo, M, Carputo, D & Bradeen, JM 2009, ‘Changes in disease resistance phenotypes associated with plant physiological age are not caused by variation in R gene transcript abundance’, MPMI, vol. 22, no. 3, pp. 362-68.
20. Naess, SK, Bradeen, JM, Wielgus, SM, Haberlach, GT, Mc Grath, JM & Helgeson, JP 2000,’Resistance to late blight in
Solanum bulbocastanum is mapped to chromosome 8’, Theor. Appl. Genet., vol. 101, pp. 697-704.
21. Ojiambo, PS, Nyanapah, JO, Lung’aho, C, Karinga, JK & Kidanemariam, HM 2000, ‘Comparing different epidemiological models in field evaluations of selected genotypes from Solanum tuberosum CIP population A for resistance to Phytophthora infestans (Mont. De Bary) in Kenya’, Euphytica, vol. 111, pp. 211-18.
22. Song, J, Bradeen, JM, Naess, SK, Raasch, JA, Wielgus, SW, Haberlach, GT, Liu, J, Kuang, H, Austin-Phillips, S, Buell,CR, Helgeson, JP & Jiang, J 2003, ‘Gene RB cloned from Solanum
bulbocastanum confers broad spectrum resistance to potato late