Secara molekular pengendalian sifat-sifat organisme oleh gen dilakukan melalui
pengendalian reaksi pada rantai metabolisme. Gen akan mengendalikan pembentukan enzim
yang berperan sebagai katalisator pada setiap tahap reaksi metabolisme, dan hasil akhir
metabolisme itu muncul sebagai sifat-sifat organisme. Gen mengendalikan pembentukan
seluruh jenis protein, dan proses pengendalikan ini disebut ekspresi gen.
Hubungan antara gen dengan protein mula-mula ditunjukkan oleh Archibald Garrod
(1902), yang mengamati penyakit alkaptonuria, suatu penyakit yang ditunjukan adanya
akumulasi senyawa asam.homogentisik pada urin penderita. Penyakit ini bersifat genetik atau
diwariskan pada generasi berikutnya, dan ternyata berhubungan dengan kerusakan enzim
penderita. Pada tahun 1941 Beadle dengan Tatum melaporkan hasil percobaan mutasi pada
Neurospora dan diperoleh informasi bahwa mutasi pada gen menyebabkan terjadinya
perubahan enzim pada mutan. Berarti satu gen berhubungan dengan satu enzim, yang
kemudian dikenal sebagai hipotesis satu gen satu enzim.
Pada tahap awal perkembangan genetik orang menduga bahwa DNA gen pada
kromosom yang menjadi model cetakan bagi pembentukan protein, namun kemudian
dibuktikan bahwa RNA yang menjadi model protein. Kemudian diketahui bahwa ekspresi
gen terbagi menjadi dua tahapan, yaitu tahapan transfer informasi genetik dari DNA ke RNA
(transkripsi), dan selanjutnya tahapan penterjemahan informasi genetik yang terdapat pada RNA ke dalam rantai polipeptida (translasi). Pada bab ini akan dijelaskan mengenai transkripsi, yang merupakan proses penyalinan runtunan basa DNA (gen) ke dalam runtunan
basa molekul RNA.
A. Utas DNA Sebagai Model untuk Sintesis RNA
Terdapat dua perangkat penting dalam transkripsi yaitu pertama utasan model cetakan, dan yang kedua enzim pengkatalisis polimerisasi RNA, yang disebut transkriptase. Pengetahuan yang didapat dari proses replikasi semikonservatif DNA, mendorong ke arah
5
BAB
TRANSKRIPSI INFORMASI
dari GEN ke RNA
pemikiran bahwa sintesis semua rantai polinukleotida, termasuk sintesis RNA, akan
memerlukan keberadaan satu rantai polinukleotida lama sebagai model cetakannya. Dalam
hal transkripsi pemikiran yang paling masuk akal ialah DNA yang ada pada kromosom
menjadi model cetakan, dan hal ini terbukti benar.
Pembuktian pertama mengenai hubungan DNA kromosom dengan RNA dilaporkan
oleh Hall dan Speigelman pada tahun 1961. Melalui percobaan hibridisasi antara DNA dan
RNA virus T2 terlihat adanya komplementasi antara molekul RNA dengan utasan DNA.
Secara garis besar percobaan dimulai dengan membiakan virus pada bakteri E.coli. Kemudian
DNA virus didenaturasi dengan suhu 1000C sehingga dihasilkan DNA utas tunggal.
Selanjutnya ke dalam tabung DNA tersebut dimasukan RNA dan dihasilkan adanya
perpasangan antara utasan DNA dengan utasan RNA. Dalam molekul hibrid DNA-RNA
virus tersebut terlihat adanya perpasangan basa yang sempurna antara kedua rantai DNA
dengan RNA tersebut. Hal ini dijadikan bukti bahwa RNA virus dibentuk oleh DNAnya.
Dalam proses transkripsi dari dua utasan DNA pada suatu gen hanya satu yang
digunakan untuk membentuk RNA; dan RNA akan mempunyai runtunan basa yang
antiparalel terhadap runtunan basa DNA yang menjadi model cetakannya (Gambar 5.1).
Pembuktian hal ini dilakukan melalui percobaan hibridisasi DNA-RNA virus SP8. Bila DNA
virus utas ganda didenaturasi akan terbentuk DNA utas tunggal, dan bila kemudian
direnaturasi secara pelahan utasan-utasan tersebut dapat kembali membentuk heliks ganda.
Dalam percobaan, saat proses renaturasi ke dalam tabung dimasukan RNA yang diproduksi
virus SP8, dan hasilnya terjadi perpasangan antara salah satu ruas DNA dengan RNA,
sedangkan utasan lain tetap dalam utas tunggal. Hal ini menunjukkan bahwa hanya satu dari
dua utasan DNA utas ganda yang digunakan sebagai model cetakan, menghasilkan RNA.
sedangkan utasan yang lainnya merupakan utas pendamping (Gambar 5.1). Dengan sistem
transkripsi seperti ini maka dari satu gen hanya ada satu RNA yang dihasilkan, dan berarti
--AAG TCA TGT TCA CTA CGA TGC GTC
---TTC AGT ACA AGT GAT GCT ACG CAG
---UUC AGU ACA AGU GAU GCU ACG CAG
Gambar 5.1. RNA merupakan hasil transkripsi dari utas cetakan. RNA antiparalel terhadap utas cetakan
DNA
RNA
selanjutnya hanya satu protein yang akan dihasilkan melalui translasi. Hal ini menunjukan
kekhasan produk gen.
Proses pemilihan utas model dan utas cetakan dilakukan oleh enzim transkriptase. Secara kimia sebenarnya kedua ruas tunggal DNA dapat digunakan sebagai model cetakan.
Sintesis RNA secara in vitro, atau dalam tabung percobaan di luar sel inang, dengan
menggunakan dua utasan DNA virus T2 yang telah dipisahkan melalui proses denaturasi,
menunjukan bahwa kedua utasan DNA tersebut dapat menghasilkan RNA. Hal ini ditunjukan
dengan dapat terbentuknya hibrid DNA-RNA antara kedua utasan DNA T2 tersebut dengan
RNA T2 yang dihasilkan secara in vitro. Namun bila sintesis RNA T2 dilakukan secara in
vivo (RNA dihasilkan dalam sel inang setelah DNA T2 diinfeksikan terlebih dahulu kedalam
E. coli), ternyata RNA in vivo tersebut dapat berpasangan hanya dengan salah satu utas DNA
saja. Berarti dalam sintesis secara in vivo enzim transkriptase yang ada dalam sel mempunyai
kemampuan membedakan kedua utasan DNA menjadi utas cetakan dan utas pendamping.
Pembedaan utasan ini dilakukan oleh transkriptase dengan cara mengenali promotor, yaitu titik awal transkripsi.
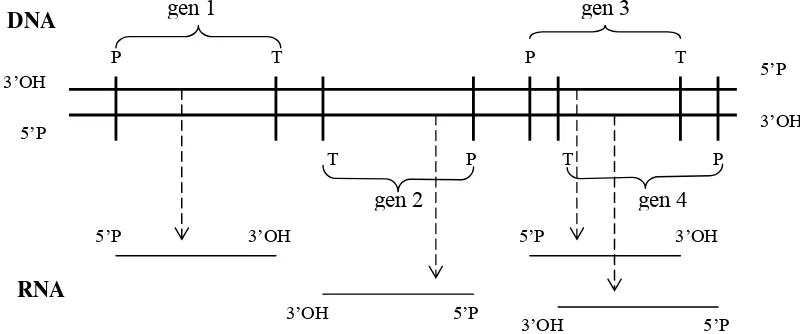
Wilayah DNA yang digunakan sebagai model cetakan dalam transkripsi RNA ialah
ruas yang dibatasi oleh promotor sebagai titik awal transkripsi dan terminator sebagai titik akhir transkripsi. Ruas yang terdapat antara promotor dan terminator disebut ruas penyandi, dan gen terletak pada ruas penyandi ini (Gambar 5.2). Promotor merupakan rangkaian
nukleotida yang tersusun sedemikian rupa sehingga dapat menjadi isyarat bagi faktor sigma
(satu komponen transkriptase) untuk membawa holoenzim transkriptase ke titk awal
DNA
tanskripsi untuk mulai bekerja mensintesis RNA. Terminator juga merupakan rangkaian basa
dengan susunan tertentu, yang berfungsi memberi tanda pada transkriptase untuk
menghentikan sintesis RNA.
Promotor dan terminator akan mempunyai makna seperti di atas seandainya dibaca
dari arah tertentu, dan tidak bermakna bila dibaca dari arah sebaliknya. Sebagai contoh pada
promotor E. coli terdapat tiga wilayah penting, yaitu kotak –35, kotak –10 dan titik awal
(lihat Gambar 5.5, dan penjelasannya). Wilayah tersebut bermakna seandainya dibaca dari
arah ujung 3’OH. Berdasarkan arah pembacaan promotor dan terminator maka transkriptase
akan dapat menentukan utasan mana dari dua utasan DNA yang akan dijadikan sebagai
model cetakan, yaitu dengan urutan promotor akan lebih dekat ke ujung 3 dan terminator
dekat ke ujung 5'. Hal ini merupakan konsekueansi pertumbuhan 5'→3' pada sintesis
polinukleotida sehingga utasan model harus dibaca dari 3' ke arah 5'. Dengan demikian kedua
utasan DNA dapat menjadi utas cetakan atau utas pendamping, pada gen-gen yang berbeda,
tergantung arah promotor dan terminatornya (Gambar 5.2).
B. Enzim Transkriptase
Perangkat yang kedua ialah enzim transkriptase yaitu suatu enzim yang berperan
dalam polimerisasi RNA, atau perangkaian nukleotida-nukleotida menjadi suatu rantai RNA.
Ditemukan pertama kali oleh Grunberg-Manago dan Ochoa pada 1955, dan terbukti bahwa
enzim ini memerlukan utasan DNA sebagai model dalam pembentukan RNA.
Terdapat berbagai struktur transkriptase, yang berbeda-beda antara bakteri dan
eukariot, yang juga berbeda dalam proses kerjanya. Yang paling rinci dipelajari adalah
transkriptase E. coli.. Pada bakteri E. coli ada subunit-subunit protein yang menyusun
holoenzim dan faktor-faktor yang tidak termasuk pada holoenzim. Holoenzim terbagi atas
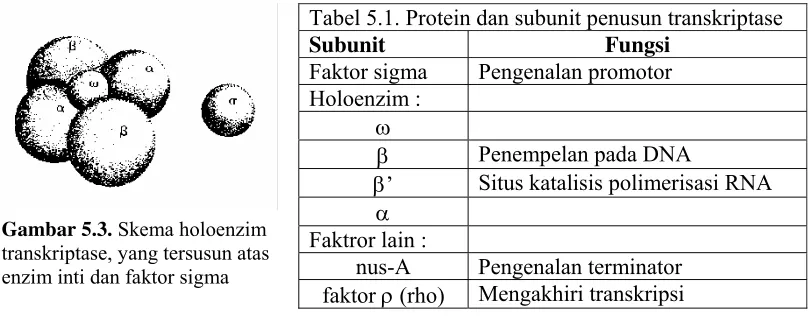
Tabel 5.1. Protein dan subunit penusun transkriptase
Subunit Fungsi Faktor sigma Pengenalan promotor
Holoenzim : ω
β Penempelan pada DNA
β’ Situs katalisis polimerisasi RNA α
Faktror lain :
nus-A Pengenalan terminator faktor ρ (rho) Mengakhiri transkripsi Gambar 5.3. Skema holoenzim
enzim inti dan faktor sigma. Enzim inti disusun oleh 5 subunit yaitu (ω, β, β',2α). Antara
subunit-subunit tersebut tidak terdapat ikatan kovalen, mereka dihubungkan oleh ikatan
sekunder (Tabel 5.1 dan Gambar 5.3). Satu subunit lain, yaitu faktor sigma (σ), akan
bergabung dengan enzim inti membentuk holoenzim, yang mempunyai bobot total sekitar
450000 dalton. Garis tengah holoenzim berukuran sekitar 100 A0 dengan panjang sekitar 30
pasang-basa dupleks DNA; tetapi pada kenyataannya sanggup melingkupi sekitar 60
nukleotida.
Sintesis polimerisasi RNA dilakukan oleh enzim inti, sedangkan faktor sigma
berfungsi mengenali promotor. Polimerisasi RNA dapat dilakukan oleh enzim inti tanpa
faktor sigma. Namun enzim inti ini tidak mampu mengenali dengan tepat promotor, dan
untuk mengenalinya diperlukan faktor sigma. Percobaan sintesis RNA secara in vitro
menunjukkan bahwa bila enzim yang digunakan untuk sintesis adalah enzim inti tanpa sigma
maka RNA yang dihasilkan tidak selalu sama ukurannya, dan bila enzim yang digunakan
adalah holoenzim ternyata RNA yang dihasilkan mempunyai ukuran sama. Hal ini menun
jukan bahwa tanpa faktor sigma, enzim inti dapat melakukan penyalinan basa DNA menjadi
RNAsintesis RNA, tetapi tidak dapat memulai penyalinan tersebut pada situs yang tepat,
karena faktor sigma yang dapat mengenali dengan tepat situs tersebut.
Terdapat dua subunit lain yaitu faktor rho dan nusA, yang ikut dalam proses
transkripsi tetapi bukan penyusun holoenzim transkriptase. Protein nusA akan menempel
pada enzim inti menggantikan faktor sigma, dan kemungkinan berfungsi dalam sintesis
perpanjangan rantai RNA. Faktor rho akan menempel pada enzim inti untuk menghentikan
sintesis RNA, dan membebaskan transkriptase dari DNA dan RNA yang dihasilkan. Tetapi
tidak semua proses sintesis RNA memerlukan faktor rho, hal ini akan dijelaskan kemudian.
Transkripsi merupakan proses sintesis pembentukan rantai RNA, melalui proses
pembacaan runtunan basa DNA model, dan menyalinnya menjadi runtunan basa RNA.
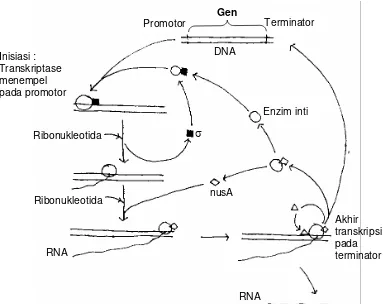
Terdapat tiga peristiwa penting dalam proses transkripsi yaitu inisiasi, sintesis perpanjangan
RNA, dan proses akhir transkripsi (Gambar 5.4).
A. Inisiasi Transkripsi pada Promotor
Proses inisiasi akan menentukan apakah suatu gen akan dapat ditranskripsikan atau
dengan pengenalan promotor oleh faktor sigma, dilanjutkan dengan penempelan enzim inti
pada promotor, dan pengudaran pilinan heliks ganda untuk memulai sintesis RNA (Gambar
5.4 dan 5.5). Terdapat hubungan khusus antara satu jenis faktor sigma dengan satu jenis
promotor, interaksi ini akan menentukan kemampuan faktor sigma mengenali promotor.
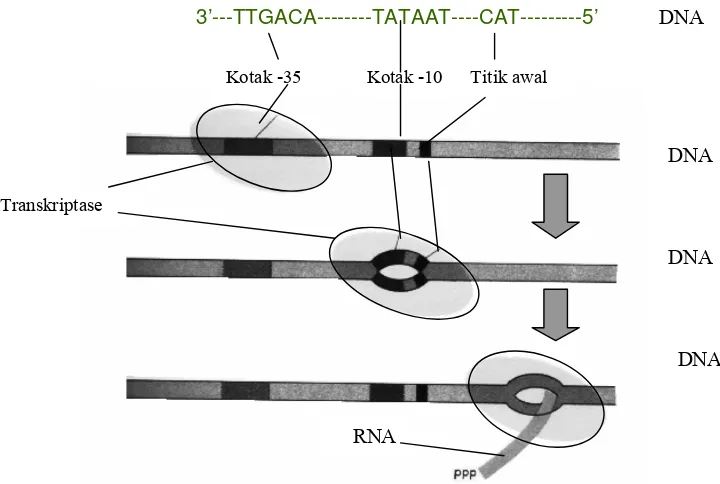
Promotor E. coli merupakan promotor yang paling banyak dipelajari, dan menjadi
model promotor bakteri. Promotor ini mempunyai ukuran sekitar 40 pasang basa, dengan tiga
titik penting yaitu kotak -35, kotak -10 dan titik awal transkripsi (Gambar 5.5). Kotak –35
mempunyai makna bahwa pada sebagian besar organisme titik tengah kotak tersebut terdapat
pada basa yang berjarak 35 pasang basa ke arah hulu dari titik awal transkripsi, hal yang sama
berlaku untuk kotak -10. Antara kotak -35 dengan kotak Pribnow dipisahkan (dalam 90%
kasus) oleh 16 sampai 18 pasang nukleotida.
Kotak -35 dan kotak -10 terdiri dari beberapa pasang basa, dan runtunannya hampir
sama untuk berbagai individu, atau disebut runtunan konsensus, sedangkan bagian lainnya
mempunyai runtunan basa yang bervariasi. Kotak –35, dengan rangkaian konsensus Gambar 5.4. Skema Proses translasi, dimulai dengan insiasi translasi (faktor σ
membawa enzim inti ke promotor, dan diakhiri dengan lepasnya enzim transkriptase pada teminaotor dengan bantuan nusA dan faktor ρ
Promotor Gen Terminator
Inisiasi : Transkriptase menempel pada promotor
σ
Enzim inti Ribonukleotida
Ribonukleotida
RNA
DNA
nusA
Akhir transkripsi pada terminator
5'TGTTGACA3', mempunyai fungsi sebagai isyarat penempelan buat transkriptase pada
DNA. Isyarat ini dapat dikenali oleh faktor sigma, salah satu subunit dari transkriptase, yang
akan menggiring enzim ini agar dapat menempel pada tempat yang tepat
Kotak -10, yang juga disebut kotak Pribnow dengan rangkaian konsensus
5'TATAAT3', merupakan tempat awal transkriptase mengudar pilinan heliks ganda DNA
menjadi dua utas tunggal. Kotak –10 yang disusun oleh rangkaian pasangan basa AT,
pasangan dengan ikatan hidrogen paling lemah, menjadikan wilayah ini utas-gandanya paling
mudah dipisahkan. Terbentuknya utas tunggal merupakan syarat untuk dapat dilakukannya
proses transkripsi.
Yang terakhir yaitu titik awal transkripsi, yang merupakan basa pertama yang
ditranskripsikan menjadi RNA (Gambar 5.5). Sebagian besar dari gen menunjukkan bahwa
titik awal itu merupakan basa purin pertama yang mempunyai jarak sekitar tujuh basa dari
kotak Pribnow. Cukup sering ditemukan bahwa titik awal merupakan titik tengah rangkaian
tiga basa CAT.
B. Proses Sintesis Perpanjangan RNA
Setelah transkriptase mengenali isyarat awal dan beberapa ribonukleotida
dirangkaikan, maka selanjutnya akan berlangsung proses perpanjangan RNA. Dalam proses 3’---TTGACA---TATAAT----CAT---5’
Kotak -35 Kotak -10 Titik awal
Transkriptase
RNA
DNA
DNA
DNA DNA
pemanjangan ini faktor sigma tidak diperlukan lagi dan akan terlepas dari enzim inti, dan
kemungkinan diganti oleh protein lain yaitu nusA (Gambar 5.4). Setelah lepas dari faktor
sigma, yang cara kerjanya sangat teliti dalam memeriksa runtunan basa, enzim inti
transkriptase akan berjalan lebih cepat.
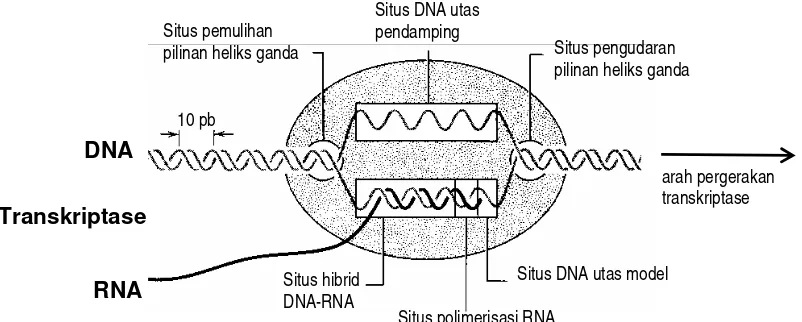
Terdapat tiga pekerjaan yang dilakukan oleh inti transkriptase, yaitu membuka pilinan
heliks DNA, melakukan sintesis RNA, dan memulihkan kembali pilinan heliks DNA
(Gambar 5.6). Ketiga kegiatan ini dilakukan berkat adanya tiga situs aktif pada enzim
tersebut. Situs penguraian heliks DNA terletak setara dengan 12pb ruas DNA dari ujung
muka transkriptase, sedangkan situs pemulihan pilinan terletak sekitar 17pb ke hilir dari situs
pengurai heliks. Pada selang antara kedua situs ini akan terbentuk DNA utas tunggal
setempat, dan pada salah satu utas, yaitu ruas DNA cetakan akan terjadi proses polimerisasi
RNA, setiap basa A, T, C, dan G pada DNA akan disalin menjadi U, A, G, dan C pada RNA.
Proses sintesis ini akan berjalan mulai dari titik awal transkripsi pada promotor, sampai pada
terminator (Gambar 5.4). Protein nusA, diketahui akan bergabung setelah faktor sigma
memisahkan diri dari enzim inti. Kemungkinan selain dalam proses; perpanjangan rantai
RNA nusA berperanan dalarn mengenali isyarat akhir transkripsi atau terminator.
C. Terminator dan Proses Akhir Transkripsi
Terminator merupakan rangkaian nukleotida DNA yang merupakan isyarat bagi
transkriptase untuk mengakhiri proses transkripsi. Terdapat dua jenis terminator, yaitu
terminator yang memerlukan faktor rho dan terminator tanpa faktor rho. Pada terminator jenis
Situs pengudaran pilinan heliks ganda Situs pemulihan
pilinan heliks ganda
Situs DNA utas pendamping
Situs polimerisasi RNA Situs hibrid
DNA-RNA
DNA
RNA
10 pb
Situs DNA utas model
Transkriptase
arah pergerakan transkriptase
pertama transkriptase akan berhenti bekerja dan tetap berada pada DNA sampai datang faktor
rho yang akan memisahkan DNA dari transkriptase serta RNA yang baru dibentuknya.
Sedangkan pada terminator tanpa faktor rho setelah transkriptase mencapai terminator dan
proses transkripsi berhenti, maka kemudian RNA dan enzim transkriptase akan terlepas dari
DNA.
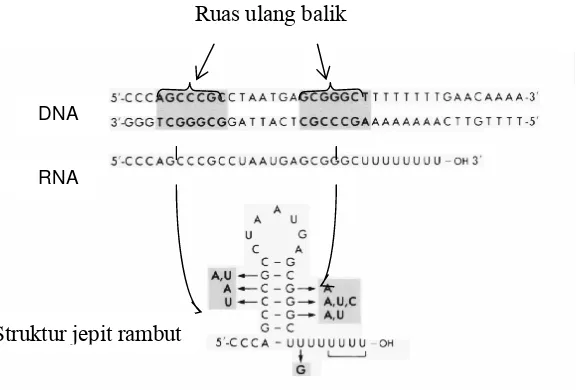
Semua terminator yang dipelajari pada prokariot mengandung dua rangkaian ulang
balik, yaitu rangkaian pasangan nukleotida yang runtunannya merupakan kebalikan dari
runtunan rangkaian yang lain. Dalam satu utasan DNA, satu rangkaian akan merupakan
pasangan antiparalel dari rangkaian yang lain seandainya dibaca dari arah yang berlawanan.
Akibatnya kedua rangkaian tersebut dapat berpasangan satu sama lain. Kedua ruas ulang
balik ini dipisahkan oleh sejumlah basa, misal pada terminator yang terdapat pada ruas
pengawal operon triptofan (trpL) masing-masing ruas ulang baliknya disusun oleh tujuh
pasang basa, dan kedua ruas tersebut dipisahkan oleh 7 pasang basa. (Gambar 5.7).
Basa-basa terminator akan ditranskripsikan ke dalam RNA. Karena adanya dua
rangkaian ulang balik yang dipisahkan oleh sejumiah nukleotida maka pada RNA akan
terdapat dua ruas yang berpasangan, dan bila hal ini terjadi maka akan ditemukan adanya
struktur seperti jepit rambut, yaitu dua batang yang berpasangan yang dihubungkan oleh satu
simpul. Struktur jepit rambut ini memberi isyarat kepada transkriptase untuk mengakhiri
pekerjaannya dalam sintesis RNA. Isyarat tersebut mungkin dapat berupa memperlambat atau
menghentikan pergerakan transkriptase sepanjang utasan DNA.
Pada terminator tanpa faktor rho disamping adanya ruas ulang balik juga terdapat
DNA
RNA
Gambar 5.7. Struktur terminator, yang mengandung ruas ulang balik, sehingga dihasulkan struktur jepit rambut pada RNA
Ruas ulang balik
rangkain pasangan basa poliAT, yang letaknya tepat dihilir ruas ulang balik yang terakhir
(Gambar 5.7). Rangkaian basa A terdapat pada ruas cetakan DNA, sehingga akan
ditranskripsikan menjadi poli-U pada RNA tepat setelah struktur jepit rambut. Jadi setelah
mentranskripsikan poli-AT maka pada situs hibrid DNA-RNA pada transkriptase akan
terdapat pasangan hibrid poliAU. Seperti diketahui bahwa pasangan poliAU merupakan
pasangan yang paling lemah, maka hibrid DNA-RNA ini akan mudah lepas. Jadi dengan
mekanisme ini proses pemisahan antara DNA, RNA, dan transkriptase terjadi pada saat akhir
proses transkripsi.
Terminator dengan faktor rho tidak mengandung ruas poliAT sebagai penutupnya;
jadi pada akhir transkripsi tidak akan ada pasangan poliAU pada pasangan hibrid DNA-RNA.
Setelah terbentuk struktur jepit rambut transkriptase akan mengakhiri proses transkripsi,
tetapi kompleks DNA-RNA- transkriptase belum dapat terpisah. Diperlukan jasa faktor rho,
yaitu suatu protein yang merupakan subunit transkriptase, yang akan berperan memisahkan
DNA, RNA, dan transkriptase dari kompleks yang terbentuk selama transkripsi.
Terdapat empat jenis RNA yang dihasilkan melalui transkripsi, yaitu mRNA, tRNA,
rRNA dan snRNA. Tiga RNA pertama merupakan RNA yang berperanan dalam proses
translasi, sedangkan snRNA terdapat hanya pada inti eukariot dan berperan pada regulasi.
A. RNA duta (mRNA)
RNA duta atau mRNA (messenger RNA) adalah RNA yang menjadi model cetakan
dalam proses penyusunan asam-amino menjadi rantai protein pada saat translasi. Dinamakan
RNA duta karena molekul ini merupakan penghubung DNA dengan protein; seolah-olah
membawa pesan berupa informasi genetik dari DNA kepada protein. Dalam proses
transkripsi rangkaian basa utas DNA menjadi cetakan dalam menyusun basa-basa mRNA,
selanjutnya dalam proses translasi sintesis protein rangkaian basa-basa mRNA ini
diterjemahkan menjadi rangkaian asam-amino rantai polipeptida.
Tabel 5.2. Perbandingan kandungan 3 jenis RNA di dalam sel serta gen penyandinya
Persentase gen penyandinya Jenisi RNA Persentase
dalam sel Bakteri Eukariot
mRNA 2 % 90 % 60 %
tRNA 10 %
rRNA 88 %
10 % 40 %
Rantai mRNA disandikan oleh gen ruas khas, yang merupakan gen paling dominan
dalam genom organisme, sekitar 90% pada bakteri dan 60% dalam eukariot (Tabel 5.2).
Tetapi walaupun gen yang membentuknya terdapat dominan di dalam genom, kuantitas
mRNA merupakan yang paling sedikit di dalam sel, yaitu sekitar 2%. Hal ini terjadi karena
mRNA tidak dibutuhkan untuk berada secara permanen didalam sel. Satu jenis mRNA
diperlukan selama protein yang disandikannya harus diproduksi, dan bila kuantitas protein
tersebut sudah mencukupi maka mRNA tersebut harus diuraikan menjadi
nukleotida-nukleotida bebas. Bentuk mRNA yang linear menjadikannya peka terhadap enzim
ribonuklease dan mRNA akan diuraikan ke dalam mononukleotida.
Informasi genetik yang dibawa oleh mRNA terdapat pada runtunan basa yang
dikandungnya. Setiap jenis kombinasi tiga basa yang berdampingan mengandung sandi
genetik tertentu (disebut kodon), yang dapat diterjemahkan menjadi satu jenis asam-amino
tertentu dalam proses translasi. Oleh karena itu rantai mRNA dapat dipandang sebagai
rangkaian kodon yang dapat diterjemahkan menjadi runtunan asam-amino. Penterjemahan
mRNA menjadi protein dimulai pada kodon awal (AUG) dan diakhiri pada kodon akhir
(UAA, UAG atau UGA). Ruas antara kodon awal dan kodon akhir ini disebut ruas penyandi
protein, dan ruas inilah yang sebenarnya setara dengan gen. Penjelasan lebih rinci mengenai
kodon dan sandi genetik akan ditemukan pada pokok Bahasan berikutnya
Keragaman panjang rantai mRNA berhubungan dengan banyaknya ruas penyandi
polipeptida yang terkandung pada mRNA tersebut serta panjang polipeptida yang akan
dibentuk. mRNA eukariot umumnya hanya mengandung satu ruas penyandi (mRNA
monogen) sedangkan pada mRNA prokariot sering ditemukan lebih dari satu ruas penyandi
(mRNA poligen).
B. RNA transfer (tRNA)
Berbeda dari mRNA yang linear, tRNA mempunyai struktur tiga dimensi (Gambar
5.7), sehingga tRNA lebih stabil dan berumur cukup panjang di dalam sel. Dalam translasi
tRNA mempunyai dua fungsi yaitu pertama menterjemahkan kodon yang terdapat pada
mRNA menjadi satu jenis asam-amino, dan yang kedua sebagai pengangkut asam-amino dari
sitoplasma ke dalam kompleks translasi. Yang dimaksud dengan kompleks translasi ialah
asosiasi mRNA tRNA dan ribosom pada saat berlangsungnya translasi. Struktur tRNA
mendukung kedua fungsi tersebut. Kemampuan menterjemahkan ini dipunyai oleh tRNA
berkat adanya antikodon yang merupakan komplemen dari kodon mRNA, serta berkat
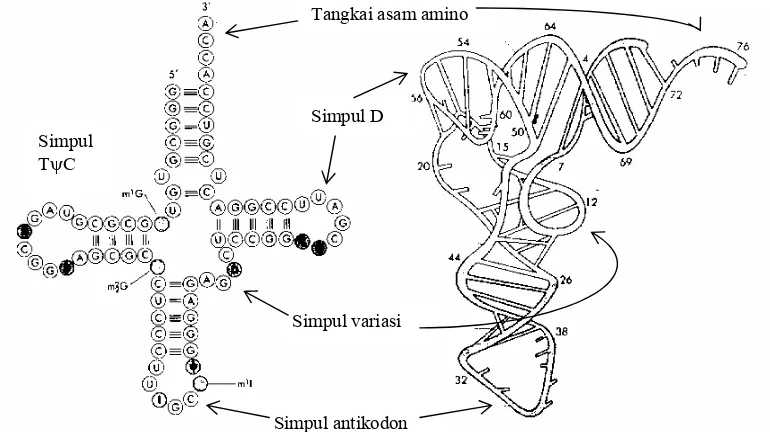
Struktur primer tRNA merupakan rantai polinukleotida linear dengan ukuran panjang
setara dengan 73 sampai 93 nukleotida (dengan berat molekul total antara 25 sampai 30 kdal).
Pada molekul tRNA dikandung nukleotida dengan basa-basa yang tidak umum, atau
termodifikasi pada proses pascatranskripsi. Pada rantai nukleotida tRNA terdapat ruas-ruas
dengan runtunan basa yang antiparalel satu dengan lainnya, sehingga dapat berpasangan
membentuk struktur sekunder.
Semua jenis tRNA, walaupun mempunyai kandungan basa yang berbeda, tetap
mempunyai model struktur sekunder yang sama, yaitu struktur daun semanggi (Gambar 5.8).
Struktur sekunder tersebut mempunyai satu batang dengan tiga cabang berutas ganda, yang
dibentuk oleh perpasangan basa antar ruas. Pada ujung ketiga cabang terdapat simpul yang
dibentuk oleh rangkaian nukleotida yang bebas tidak berpasangan dengan jumlah basa dalam
simpul antara tujuh sampai dua belas. Sesuai dengan basa yang dikandungnya ketiga simpul
tersebut diberi nama sebagai simpul TψC terdiri dari 7 basa, simpul antikodon mengandung 7
basa, dan simpul D mengandung 8 sampai 12 basa. TψC mengandung rangkaian basa
5'TψCG3' yang berfungsi dalam penempelan tRNA pada ribosom. Pada simpul antikodon
terdapat rangkaian tiga basa antikodon yang berfungsi untuk membaca kodon pada mRNA.
Pada bagian tangkai semua basa berpasangan kecuali empat nukleotida yang terdapat
pada ujung 3’. Rangkaian basa keempat nukleotida itu selalu sama untuk berbagai tRNA
yaitu 3’ACCPy5', yang berfungsi sebagai tangkai penerima asam-amino. Pada bagian ujung
3'ACCPy5' akan menempel asam-amino, yang akan dibawa tRNA menempel pada ribosom. Gambar 5.8. Struktur trNA (a) struktur sekunder (a) dan (b) struktur tersier
Simpul antikodon Simpul variasi
Simpul D
Tangkai asam amino
Selain batang dan tiga cabang terdapat satu simpul berukuran kecil yang terletak pada
sudut antara tangkai TψC dengan tangkai antikodon, biasa disebut simpul variasi. Banyaknya
basa yang terletak pada simpul tersebut beragam dari satu jenis tRNA dengan jenis tRNA
yang lain, sehingga memberikan keragaman bentuk akhir tRNA. Struktur tersier tRNA
terbentuk akibat terjadinya perpasangan pada simpul TψC dengan basa pada simpul D.
Struktur tersier tRNA mempunyai bentuk yang menyerupat huruf Γ, dengan sudut huruf Γ
ditempati oleh simpul TψC, dan pada kedua ujung bebas ditempati simpul antikodon dan
tangkai penerima asam-amino (Gambar 5.8).
C. RNA Ribosom (rRNA)
RNA yang ketiga yang ikut terlibat dalam proses sintesis protein yaitu RNA ribosom
(rRNA). RNA ini merupakan RNA terbanyak, sekitar 83%, dari RNA yang dikandung suatu
sel. Besarnya jumlah rRNA dalam sel karena molekul rRNA bersifat stabil, berukuran besar,
dan jumlah ribosom di dalam sel sangat banyak. Ribosom merupakan organel selular yang
sangat penting, terdapat dalam semua sel mahluk hidup, berperan dalam sintesis rantai
protein, yaitu sebagai tempat pertemuan mRNA dengan tRNA pembawa asam-amino.
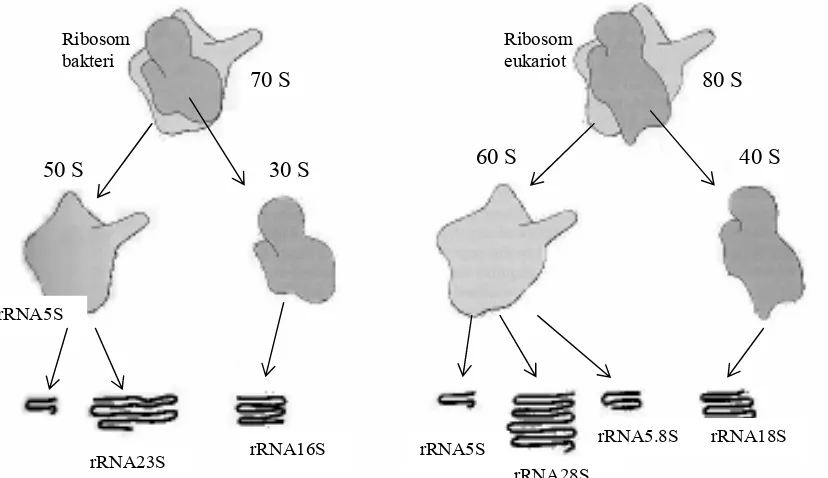
Ribosom disusun oleh dua subunit yaitu ribosom kecil dan ribosom besar, yang
kedua-duanya disusun oleh protein dan rRNA. Kedua subunit tersebut dicirikan oleh
ukurannya yang besarannya ditentukan dengan kecepatan pengendapan. kecepatan Subunit
rRNA5S
Gambar 5.9 Ribosom bakteri dan eukariot beserta komponen-komponen penyusunnya
rRNA23S rRNA16S rRNA5S
rRNA28S
rRNA5.8S rRNA18S Ribosom
bakteri
Ribosom eukariot
70 S
50 S 30 S
80 S
kecil ribosom prokariot berukuran 30 S, dan subunit besar berukuran 50 S, sedangkan
keseluruhannya berukuran sama dengan 70S. Ribosom eukariot mempunyai ukuran lebih
besar, ukuran totalnya 80S dengan subunitnya 60S dan 40S (Gambar 5.9)
Pada E. coli terdapat tiga jenis rRNA, yaitu rRNA16S pada subunit 30S, dengan
panjang rantai setara dengan 1542 nukleotida, kemudian pada subunit 50S diperoleh
rRNA23S yang mengandung 2904 nukleotida, dan rRNA5S dengan 120 nukleotida. Ribosom
eukariot mengandung rRNA yang lebih besar dibandingkan dengan yang dikandung oleh
prokariot. Pada keseluruhan ribosomnya terdapat empat jenis rRNA, satu molekul, yaitu
rRNA 18S (17S pada khamir), terdapat pada subunit ribosom 40S, dan tiga molekul lainnya,
yakni rRNA28S (25S pada khamir), serta dua molekul kecil rRNA 5,8S dan 5S terdapat pada
subunit 60S. Pada ribosom mitokondria terdapat hanya dua molekul rRNA.
Sampai saat ini belum diketahui dengan pasti mengenai fungsi rRNA, mengapa begitu
besar masa rRNA dalarn ribosom. Tetapi kita percaya bahwa banyaknya basa yang tidak
berpasangan pada rRNA berhubungan dengan proses penempelan molekul RNA yang lain
pada ribosom. Beberapa basa dekat ujung 3' pada rRNA 16S dapat membentuk pasangan basa
temporer dengan situs Shine-Dalgarno molekul mRNA; yang merupakan situs pengenalan
mRNA oleh ribosom. Begitu pula ruas rRNA dapat berinteraksi dengan semua tRNA tanpa
kecuali selama penempelan tRNA pada ribosom. Juga basa-basa rRNA mungkin membentuk
pasangan yang berhubungan dengan terbentuknya kompleks RNA- protein yang terlibat
dalam menuntun proses sintesis protein baru didalarn sel.
Gen diekspresikan melalui peranan dalam pengendalian sifat-sifat organisme. Peran ini dijalankan melalui pengendalian proses pembentukan protein dan enzim-enzim yang mengkatalisis reaksi kimia pada berbagai tahapan metabolisme. Gen diekspresikan melalui dua tahapan yaitu transkripsi dan translasi. Transkripsi adalah penyalinan informasi dari gen kedalam molekul RNA, yang dalam pelaksanaannya adalah berupa penyusunan basa-basa pada rantai RNA dengan menggunankan runtunan basa DNA gen sebagai modelnya. Translasi adalah penterjemahan informasi berupa runtunan basa RNA menjadi rangkaian asam-amino pada protein.
Dalam transkripsi terdapat dua perangkat yaitu ruas DNA yang menjadi model, dan enzim transkriptase yang mengkatalisis proses sintesis RNA. Ruas DNA yang dijadikan model atu juga disebut ruas penyandi ialah ruas yang dibatasi oleh promotor dan terminator. Promotor ialah segmen DNA yang berfungsi sebagai tanda bagi enzim transkriptase untuk mengawali proses transkripsi atau penyalinan basa DNA menjadi basa RNA. Terminator ialah segmen DNA yang menjadi tanda untuk berakhirnya proses transkripsi. Panjang ruas antara promotor dan terminator (ruas penyandi) akan sama dengan panjang RNA yang dihasilkan.
Terdapat empat jenis RNA hasil transkripsi yaitu mRNA, tRNA, rRNA, dan snRNA. Tiga RNA selain snRNA berperanan dalam proses translasi atau sintesis protein. mRNA berperan sebagai model untuk menyusun runtunan asam-amino rantai polipeptida atau protein. tRNA berperan sebagai pengangkut asam-amino dan penterjemah rangkaian kodon-kodon yang terdapat pada mRNA menjadi rangkaian asam-amino. rRNA berfungsi sebagai rangka ribosom dan mengenali tRNA dan mRNA.
Pilih jawaban A, B, C, atau D yang benar, atau pilih A bila 1&2 benar, B bila 1&3 benar, C bila 2&3 benar, dan D bila 1&2&5 benar
1. Ekspresi gen merupakan proses molekular pengendalian suatu sifat oleh suatu gen. Hal ini dilakukan melalui
1.Pengendalian proses metabolisme 2.Pengendalian pembentukan enzim 3.Pengendalian pembentukan RNA
A, B, C, D
2. Gen diekspresikan melalui proses 1. Replikasi
2. Transkripsi 3. Translasi
A, B, C, D
3. Yang dibentuk pada proses transkripsi dan berperanan dalam proses translasi adalah
1. mRNA
2. tRNA 3. rRNA
A, B, C, D
4. RNA yang keberadaannya permanen di dalam sel ialah
1. mRNA
2. tRNA 3. rRNA
A, B, C, D
5. mRNA mempunyai sifat dan peran berikut 1. mempunyai struktur linear
2. berperan sebagai model dalam pembentukan protein 3. dalam sel dibentuk saat diperlukan untuk translasi A, B, C, D
6. tRNA berperan sebagai
1. model untuk merangkaikan asam amino
2. penterjemah kodon-kodon yang ada pada mRNA 3. pengangkut asam-amino saat translasi
A, B, C, D 7. rRNA berperan sebagai
1. kerangka ribosom
2. pengenal mRNA saat translasi 3. pengenal tRNA saat translasi A, B, C, D
8. Enzim yang berperan dalam proses ekspresi gen adalah A. Polimerase DNA I
B. Polimerase DNA III C. Transkriptase D. Ligase
9. Pada proses transkripsi akan dilakukan penyalinan basa-basa DNA manjadi basa RNA, yang akan dimulai pada
A. titik ori B. promotor C. kodon awal
D. terminator
Albert B, Bray D, Lewis J, Raff M., Robert K, and Watson J. 1994. Molecular Biology of the
Cell. Fourth Edition. Garland Publishing Inc. New York and London.
Ayala FJ, Keger JA. 1984. Modern Genetics. Second Edition. The Benjamin/Cummings Co.
Inc. Menlo Park, CA
Beadle GW and Tatum EL. 1941. Genetic control of biochemical reactions in Neurospora.
Science 27: 499-506.
Crick FHC, 1966. Codon-anticodon pairing: the wobble hypothesis. J.Mol.Biol. 19: 548-550
Jusuf M. 2001. Genetika I, Struktur dan Ekspresi Gen, Sagung Seto. Jakarta.
Lewin B. 1990. Genes IV. Oxford University Press. Oxford
Niremberg MW and Leader P. 1964. The effect of trinucleotides upon the binding of sRNA to
ribosomes. Sciences 145: 1399-1407
Russel PJ. 1996. Genetics. Fourth Edition. Harper Collins College Publisher. New York.
Stryer L. 1995. Biochemistry. WH Freeman and Company. New York.
Watson JD, Hopkins NH, Robert JW, Steiz JA, Weiner AM. 1987. Molecular Biology of the
Gene. Fourth Edition.Benjamin/Cummung Co. Inc., Menlo Park CA.
Zubay G. 1987. Genetics. The Benjamin Cummings Publishing Company Inc. Menlo Park,
California
Zubay G and Marmur (Eds) 1973. Papers in Biochemical Genetics. Secod Edition. Holt,
Rinehart and Winston. New York.