BIOKIMIA KOMPARATIF
PERBANDINGAN ANABOLISME PROTEIN
DARI HEWAN DENGAN FUNGI
OLEH
MUHAMMAD DAILAMI G85 11 300 21
DEPARTEMEN BIOKIMIA
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
DAFTAR ISI
C. Perbandingan Sintesis Asam Amino pada Hewan & Fungi ... 37
3 PENUTUP ... 39
Kesimpulan ... 39
DAFTAR TABEL
Tabel 1. Perbedaan molekul DNA dan RNA ... 5
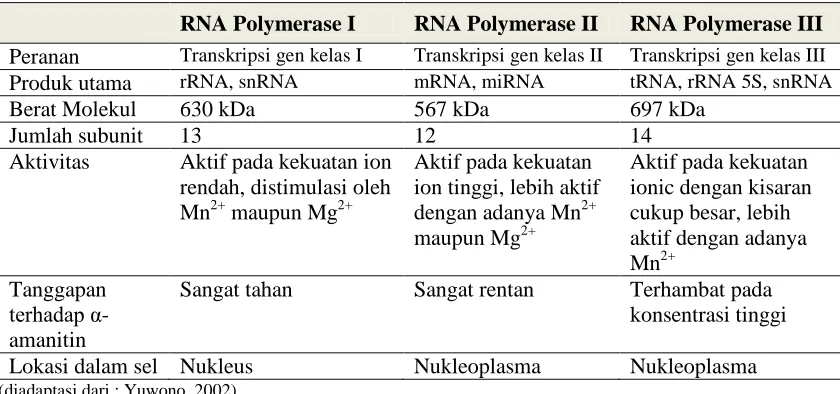
Tabel 2. Perbedaan sifat ketiga RNA polimerase ... 6
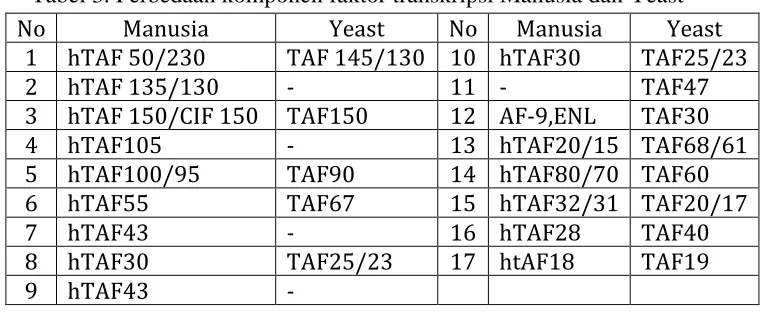
Tabel 3. Perbedaan komponen faktor transkripsi Manusia dan yeast ... 8
Tabel 4. Kode genetik Universal ... 14
Tabel 5. Kebutuhan Asam Amino Manusia ... 23
DAFTAR GAMBAR
Gambar 1. Pohon Filogenetik system tiga domain ... 1
Gambar 2. Struktur sel fungi dan hewan ... 3
Gambar 3. Sentral dogma biologi molekuler ... 4
Gambar 4. Perbedaan DNA dan RNA ... 5
Gambar 5. Pembentukan kompleks pra inisiasi transkripsi ... 7
Gambar 6. Ilustrasi hipotesis transkripsi gen kelas III ... 10
Gambar 7. Bentuk ribosom ... 13
Gambar 8. Struktur dan bagian tRNA ... 15
Gambar 9. Inisiasi Translasi ... 17
Gambar 10. Pemanjangan rantai polipeptida ... 19
Gambar 11. Terminasi Translasi ... 21
Gambar 12. Jalur metabolisme asam amino ... 23
Gambar 13. Kelompok dan Sumber sintesis asam amino pada fungi (yeast) ... 28
I PENDAHULUAN
1.1 Latar Belakang
Metabolisme merupakan salah satu pokok kajian dalam bidang biokimia. Pada dasarnya, metabolisme makromolekul dapat dikelompokkan menjadi dua jalur utama yang berbeda yaitu jalur katabolik dan jalur anabolik. Katabolisme adalah proses degradasi atau penguraian makromolekul besar menjadi monomer-monomernya untuk mendapatkan energy. Sedangkan anabolisme adalah proses pembentukan atau biosintesis molekul yang kompleks dari prekursor molekul yang lebih sederhana yang berlangsung dalam tubuh makhluk hidup.
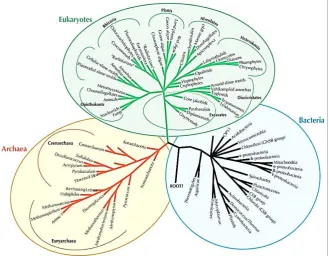
Makhluk hidup yang menghuni bumi memiliki jumlah yang sangat banyak, sehingga penting bagi kita untuk mengetahui klasifikasi atau pengelompokan makhluk hidup itu sendiri. Klasifikasi makhluk hidup dengan sistem tiga domain, menjelaskan bahwa seluruh makhluk hidup dapat dikelompokkan menjadi tiga kelompok utama yaitu: domain bacteria, domain archea dan domain eukariota. Domain eukariot sendiri terbagi lagi menjadi tiga kingdom utama yaitu: kingdom animalia, kingdom plantae dan kingdom fungi. Pohon filogenetik dari klasifikasi ini ditampilkan pada Gambar 1.
Gambar 1. Pohon filogenetik sistem tiga domain (sumber: Eisen, J. 2008)
inti sel seakan-akan menyatu dalam sitoplasma. Berbeda halnya dengan sel eukariot yang telah memiliki membran inti sehingga nukleus (inti selnya) dapat dibedakan dengan jelas. Adanya perbedaan struktur sel dari masing-masing kelompok organisme tentunya akan memberikan penjelasan jalur metabolisme yang berbeda pula. Untuk memahami proses metabolisme secara detail dari setiap kelompok ini perlu adanya pembandingan setiap jalur metabolisme dari satu kelompok dengan kelompok lainnya.
Pembahasan pada paper ini akan memfokuskan pada jalur anabolisme protein yang terjadi pada hewan dan fungi. Hewan dan fungi merupakan dua kingdom makhluk hidup yang termasuk dalam domain eukariot, hal ini akan memudahkan kita dalam mempelajari jalur anabolisme protein pada kedua kelompok ini. Dari dua kelompok yang berada dalam satu domain eukariot ditemukan sedikit perbedaan jalur anabolisme proteinnya dan sebagian besar memiliki tahapan dan proses yang relatif sama. Perbedaan yang signifikan terlihat pada jalur anabolisme antara domain prokariot (bacteria dan archea) jika dibandingkan dengan domain eukariot (animalia, plantae maupun fungi).
Klasifikasi dari kingdom animalia dapat dikelompokan menjadi invertebrate (hewan tidak bertulang belakang) dan vertebrata (hewan bertulang belakang). Pada dasarnya, system metabolisme baik pada vertebrata maupun invertebrate adalah sama, oleh karenanya dalam pembahasan untuk kingdom animalia akan dibahas secara umum. Sedangkan untuk kingdom fungi dapak dikelompokkan lagi menjadi: Zygomycota, Ascomycota, Basidiomycota, Deuteromycota, Mikoriza, Lumut Kerak. Salah satu contoh fungi yang paling banya dipelajari proses metabolismenya adalah yeast. Oleh karena itu, sebagai perwakilan dari fungi akan dibahas anabolisme protein pada yeast.
Untuk memudahkan kita dalam memahami Anabolisme protein pada hewan dan fungi, maka pembahasan dalam paper ini akan dimulai dengan struktur sel dari kedua kelompok organisme ini, proses sintesis protein yang melibatkan proses trasnkripsi dan translasi. Kemudian akan dijelaskan lebih detail mengenai mekanisme sintesis asam amino yang tidak dapat diproduksi sendiri oleh hewan maupun fungi.
1.2 Rumusan Masalah
Rumusan masalah dalam paper ini yaitu bagaimana perbandingan jalur anabolisme protein yang terjadi pada hewan dan yang terjadi pada fungi. Perbandingan struktur sel dari kedua organisme ini dan hal-hal apa yang menjadi perbedaan maupun persamaan dari jalur anabolisme protein pada kedua kelompok organisme ini.
1.3 Tujuan
II PEMBAHASAN
2.1 Struktur Sel Hewan dan Fungi
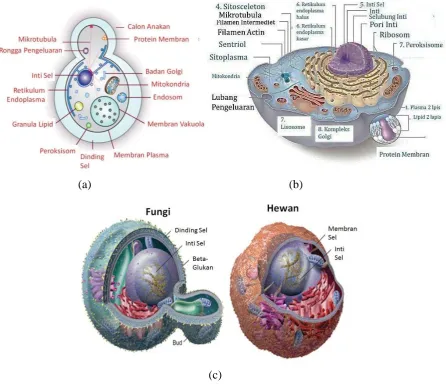
Sel sebagai unit terkecil dari kehidupan memiliki komponen-komponen yang lebih kecil didalamnya. Secara umum, organisme eukariot memiliki bagian-bagian sel yang terdiri dari inti sel, membran inti, sitoplasma, ribosom, mitokondria, retikulum endoplasma, badan golgi. Sel hewan dan fungi memiliki kemiripan struktur, karena keduanya merupakan sel eukariot. Struktur sel secara detail seperti terlihat pada Gambar 2. Perbedaan antara sel hewan dan sel fungi yaitu pada dinding selnya. Sel hewan tidak memiliki dinding sel, hanya memiliki membran sel. Sedangkan fungi memiliki membran sel dan dinding sel. Dinding sel dari fungi terususun atas senyawa khitin, berbeda halnya dengan dinding sel pada tumbuhan. Selain itu, pada fungi terdapat vakuola dengan ukuran yang lebih besar di bandingkan pada hewan.
(a) (b)
(c)
2.2 Sintesis Protein
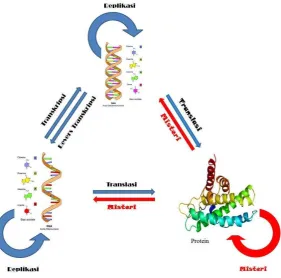
Sentral dogma dalam biologi molekuler menyatakan bahwa proses sintesis protein pada makhluk hidup, dimulai dengan tahap trasnkripsi DNA menjadi RNA yang selanjutnya akan ditranslasikan menjadi urutan asam amino yang spesifik yang menyandi suatu protein. Urutan asam amino ini merupakan ciri yang khas dari setiap protein. Selain mengalami transkripsi menjadi RNA, DNA sebagai cetak biru kehidupan (blue prin of life) juga akan mengalami replikasi guna memperbanyak molekul DNA itu sendiri.
Gambar 3. Sentral Dogma Biologi Molekuler
2.2.1 Transkripsi Hewan dan Fungi
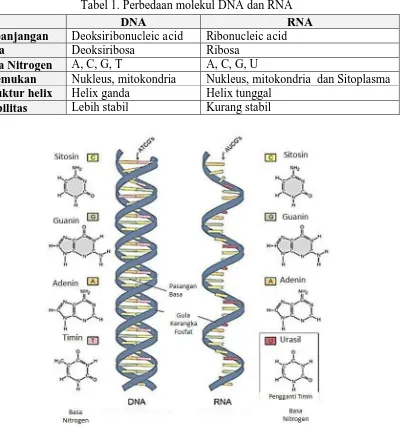
Sebagaimana telah disinggung pada paragraf sebelumnya, transkripsi merupakan proses sintesis RNA dengan cetakan dari molekul DNA, maka perlu kita pahami terlebih dahulu perbedaan antara molekul DNA dan RNA. Gula ribosa pada molekul DNA kehilangan satu atom oksigen pada atom C nomor 2, sehingga disebut sebagai deoksi, sedangkan pada RNA, atom C nomor 3 tetap mengikat gugus OH. DNA hanya dapat ditemukan dalam inti dan mitokondria. Hal ini dikarenakan DNA merupakan materi genetik yang sangat penting sehingga keberadaanya dalam sel harus dijaga dengan baik didalam inti. Struktur DNA merupakan Heliks ganda, sedangkan RNA hanya berupa untaian heliks tunggal, sehingga molekul DNA lebih stabil dari RNA. Kestabilan DNA sangat dipengaruhi karena hilangnya satu atom Oksigen pada atom C nomor 2 sehingga DNA kurang reaktif.
Tabel 1. Perbedaan molekul DNA dan RNA
DNA RNA
Kepanjangan Deoksiribonucleic acid Ribonucleic acid
Gula Deoksiribosa Ribosa
Basa Nitrogen A, C, G, T A, C, G, U
Ditemukan Nukleus, mitokondria Nukleus, mitokondria dan Sitoplasma Struktur helix Helix ganda Helix tunggal
Stabilitas Lebih stabil Kurang stabil
Proses Transkripsi pada eukariot (baik hewan maupun fungi) melalui proses yang cukup kompleks dibandingkan dengan prokariot. Pada eukariot terdapat pembagian gen menjadi tiga kelompok. Gen adalah untaian DNA yang menyandi satu molekul protein atau molekukl RNA. Tiga kelas gen eukariot adalah: gen kelas I, gen kelas II dan gen kelas III. Dilakukan pengelompokkan gen-gen ini, karena proses transkripsi pada masing-masing kelas gen tersebut melibatkan faktor transkripsi dan enzim yang berbeda-beda. Selain itu, kelas-kelas gen ini juga memiliki implikasi pada struktur dari masing-masing kelas gen.
Gen kelas I adalah gen-gen yang menyandi beberapa macam ribosomal RNA (rRNA) yang terdiri dari 18s rRNA, 28s rRNA dan 5,8s rRNA.
Gen Kelas II adalah gen yang mengkode beberapa jenis RNA nukleus dan keseluruhan jenis protein.
Gen Kelas III adalah gen yang menyandi beberapa jenis RNA nukleus dan molekul 5S rRNA serta tRNA.
Proses transkripsi pada masing-masing kelas gen, melibatkan enzim RNA polymerase yang berbeda-beda. Gen kelas I ditranskripsikan oleh enzim RNA polymerase I, gen kelas II oleh RNA polymerase II dan gen kelas III oleh RNA polymerase III (Anonim, 2013). Setiap jenis RNA polymerase memiliki karakter dan sifat yang berbeda-beda. Berikut ini adalah table perbedaan dari setiap RNA polymerase.
Tabel 2. Perbedaan sifat ketiga RNA polymerase
RNA Polymerase I RNA Polymerase II RNA Polymerase III
Peranan Transkripsi gen kelas I Transkripsi gen kelas II Transkripsi gen kelas III
Produk utama rRNA, snRNA mRNA, miRNA tRNA, rRNA 5S, snRNA
Berat Molekul 630 kDa 567 kDa 697 kDa
Sangat tahan Sangat rentan Terhambat pada konsentrasi tinggi
Lokasi dalam sel Nukleus Nukleoplasma Nukleoplasma
(diadaptasi dari : Yuwono, 2002)
A. Mekanisme transkripsi gen kelas II
TFIIH, TFIIJ. Pada dasarnya, transkripsi terjadi melalui tiga tahap utama, yaitu: inisiasi, elongasi dan terminasi (Azhar, 2008).
Inisiasi Transkripsi
Tahap awal dari proses inisiasi ini adalah pembentukan kompleks faktor transkripsi (kompleks pra-inisiasi). Adanya ikatan dengan kompleks pra inisiasi akan menyebabkan terbukanya rantai untai ganda DNA. Urutan penempelan kompleks pra inisiasi adalah sebagai berikut:
1. TFIID akan menempel pada bagian kotak TATA pada promoter, yang dibantu oleh faktor transkripsi TFIIA, kompleks ini disebut kompleks DA.
2. Kemudian diikuti dengan penempelan TFIIB, TFIIF dan RNA polymerase secara berurutan.
3. Akhirnya TFIIE akan menempel dan diikuti oleh TFIIH, kompleks pra inisiasi ini disebut kompleks DABPoIFEH.
Ilustrasi proses penempelan faktor transkripsi dalam membentuk kompleks pra inisiasi disajikan pada Gambar 5. Setelah kompleks pra inisiasi terbentuk, RNA polymerase II bersama dengan TFIIH menutupi daerah promoter mulai dari posisi -34 sampai dengan +17. Tahap selanjutnya yaitu proses pemanjangan rantai atau dikenal dengan elongasi.
Terdapat perbedaan komponen penyusun faktor transkripsi pada yeast dan hewan, yang disajikan pada tabel 3.
Tabel 3. Perbedaan komponen faktor transkripsi Manusia dan Yeast
Elongasi Transkripsi
Pemanjangan rantai RNA terjadi pada bagian gelembung transkripsi, basa-basa RNA akan membentuk hibrid dengan DNA cetakan pada panjang sekitar 12 nukleotida. Ikatan antara RNA dan DNA ini bersifat sementera, dan akan terlepas kembali setelah molekul RNA polymerase berjalan. Sementara itu, molekul heliks ganda yang terbuka akan tertutup kembali. Urutan RNA yang dihasilkan merupakan komplementer dari molekul DNA cetakan, tetapi identik dengan urutan DNA pengkode, sesuai dengan contoh berikut ini:
5’-ATG GTC CTT TAC TTG TCT GTA TTT-3’ DNA pengkode 3’-TAC CAG GAA ATG AAC AGA CAT AAA-5’ DNA cetakan
5’-AUG GUC CUU UAC UUG UCU GUA UUU-3’ RNA hasil transkripsi
Laju pemanjangan transkrip secara umum berkisar antar 30-60 nukleotida perdetik. Dalam proses pemanjangan, molekul nukleotida RNA ditambahkan secara kovalen pada ujung 3’ RNA yang baru. Terdapat dua hipotesis mengenai mekanisme pemanjangan transkrip ini. Hipotesis 1: menyatakan bahwa, selama pemanjangan rantai, RNA polymerase akan bergerak melingkari untaian, sehingga molekul DNA tidak mengalami pelintiran, sedangkan molekul RNA yang baru yang mengalami pelintiran. Hipotesis 2: menyatakan bahwa, selama pemanjangan rantai, RNA polymerase akan berjalan lurus sehingga, molekul DNA akan terpelintir dan ini yang menyebabkan nukleotida didepannya akan membuka dan molekul nukleotida dibelakang akan memuntir menutup kembali. Selain itu, molekul RNA yang terbentuk tidak akan terpelintir. Tahapan berikutnya setelah pemanjangan adalah terminasi.
Terminasi Transkripsi
Pengakhiran proses transkripsi akan terjadi ketika RNA polymerase mengalami bentuk yang tidak terfosforilasi, yang disebabkan oleh adanya aktivitas fosfatase yang spesifik
untuk CTD. Proses terminasi transkripsi pada eukariot baik hewan maupun fungi belum banyak diketahui. Berbeda halnya dengan bakteri atau prokariot.
B. Mekanisme transkripsi gen kelas I
RNA polymerase I merupakan gen yang bertanggung jawab pada proses transkripsi gen kelas I. Tahap awal dimulai dengan pembentukan kompleks pra inisiasi yang dilakukan oleh RNA polimerase I dan dua faktor transkripsi yaitu SL1 dan UBF. Tahapan detail dari mekanisme inisiasi transkripsi gen kelas I belum banyak diketahui. SL1 merupakan faktor transkripsi yang spesifik untuk setiap spesies tertentu. Faktor SL1 diketahui berperan dalam penyusunan kompleks pra inisiasi RNA polimerase 1. Spesifisitas SL1 terhadap suatu promotor dibantu oleh elemen promotor utama (core promoter elemen). Pada manusia, faktor SL1 manusia tidak berikatan pada daerah promotor gen rRNA secara langsung. Sedangkan pada mencit, faktor SL1 mencit menempel langsung pada promotor gen rRNA mencit. Sebagai akibatnya, faktor SL1 manusia hanya aktif terhadap promotor manusia sedangkan SL1 mencit hanya aktif pada promotor mencit. Akan tetapi, faktor UBF manusia dapat menggantikan fungsi UBF dari mencit dan sebaliknya.
RNA Pol I bertanggung jawab dalam sintesis rRNA secara terus-menerus selama interfase. Sel manusia mengandung lima rumpun (cluster) gen penyandi rRNA yang terdiri atas sekitar 40 salinan dan terletak pada kromosom-kromosom yang berbeda. Masing-masing gen rRNA menghasilkan transkrip 45S rRNA yang panjangnya lebih kurang 13.000 nukleotida (nt). Transkrip ini akan terbagi menjadi sebuah 28S (5.000 nt), 18S (2.000 nt), dan 5,8S (160 nt) rRNA. Transkripsi salinan gen-gen rRNA secara berkesinambungan diperlukan untuk mencukupi produksi rRNA yang selanjutnya akan dikemas ke dalam ribosom.
C. Mekanisme transkripsi gen kelas III
RNA polymerase III memiliki tugas dalam transkripsi gen kelas III yang terdiri dari gen tRNA dan 5S rRNA. Enzim ini menyintesis prekursor tRNA, 5S rRNA, serta berbagai snRNA dan RNA sitosolik. Proses transkripsi gen kelas III dibantu oleh sekelompok protein yang dikenal sebagai faktor transkripsi TFIII yang meliputi; TFIIIA, TFIIIB, dan TFIIIC serta protein TBP.
invitro kompleks ikatan TFIIIA, TFIIIB, TFIIIC, dan RNA polimerase III tersebut dapat mendukung proses transkripsi sampai kurang lebih 40 kali. Selama rangkaian proses tersebut, RNA polimerase III akan berdisosiasi dan berasosiasi kembali ke dalam kompleks protein setiap kali terjadi proses transkripsi. Sejauh ini diketahui bahwa terminasi transkripsi gen kelas III terjadi pada suatu daerah tertentu dan tidak melibatkan protein khusus. Ilustrasi dari hipotesis ini disajikan pada Gambar 6.
Dua faktor pengikatan DNA yang kompleks telah diketahui memegang peranan penting dalam inisiasi transkripsi tRNA oleh RNA Pol III TFIIIC mengikat baik kotak A maupun kotak B di dalam promoter tRNA. Sementara itu, TFIIIB mengikat daerah sejauh 50 pb ke arah hulu dari kotak A. TFIIIB terdiri atas tiga subunit, yang salah satu di antaranya adalah TBP, suatu faktor inisiasi umum yang diperlukan oleh ketiga RNA polimerase. Subunit yang kedua dan ketiga masing-masing dinamakan BRF dan B’’.Faktor TFIIIB tidak memiliki spesifisitas urutan sehingga tempat pengikatannya bergantung kepada posisi pengikatan TFIIIC pada DNA. TFIIIB memungkinkan RNA Pol III untuk melakukan inisiasi transkripsi.Begitu TFIIIB terikat, TFIIIC dapat dikeluarkan tanpa mempengaruhi transkripsi.Oleh karena itu, TFIIIC dapat dilihat sebagai faktor perakitan untuk penempatan faktor inisiasi TFIIIB.
Gambar 6. Ilustrasi hipotesis transkripsi gen kelas III
C. Letaknya sekitar 81 hingga 99 pb ke arah hilir dari tapak inisiasi transkripsi. Selain itu, terdapat juga kotak A yang berada pada posisi sekitar +50 hingga +65. Kotak C pada promoter 5S rRNA berperan sebagai tempat pengikatan protein spesifik, yaitu TFIIIA.TFIIIA bekerja sebagai faktor perakitan yang memungkinkan TFIIIC berinteraksi dengan promoter 5S rRNA. Sementara itu, kotak A akan menstabilkan pengikatan TFIIIC sehingga faktor ini berikatan dengan DNA pada posisi yang relatif sama dengan posisi pengikatan pada promoter tRNA. Begitu TFIIIC terikat pada DNA, TFIIIB dapat berinteraksi dengan kompleks pengikatan tersebut dan memungkinkan RNA Pol III untuk melakukan inisiasi transkripsi.
D. Proses Pasca Transkripsi
Pada eukariot, proses translasi tidak terjadi secara bersamaan seperti pada prokariot. Proses translasi hanya bisa berlangsung setelah proses transkripsi selesai dikerjakan. Setelah transkripsi, molekul RNA hasil transkripsi (disebut RNA transkrip) akan mengalami beberapa proses tambahan sebelum dilanjutkan untuk proses translasi. Proses lanjutan ini disebut sebagai proses pasca transkripsi. Terdapat empat proses pasca transkripsi pada eukariot, yaitu :
1. RNA Splicing : pemotongan dan penyambungan RNA
2. Poliadenilasi: penambahan gugus poli-A pada ujung 3’ mRNA
3. Penambahan tudung (cap) pada ujung 5’ mRNA
RNA splicing adalah proses pemotongan dan penyambungan molekul RNA dengan tujuan untuk menghilangkan intron (bagian yang tidak mengkode protein) dari suatu gen. Molekul mRNA hasil transkripsi yang masih mengandung intron, disebut dengan pre-mRNA, setelah proses splicing, maka semua intron akan hilang sehingga mRNA hanya memiliki bagian ekson-ekson. Molekul mRNA yang hanya terdiri dari ekson-ekson disebut dengan mature RNA atau dikenal dengan RNA “matang”. Selain mRNA, splicing juga terjadi pada molekul rRNA dan tRNA (Yuwono, 2002).
Poliadenilasi mRNA adalah proses penambahan molekul poliA (rantai AMP)
pada bagian ujung 3’. Jumlah molekul AMP yang ditambahkan bisa mencapai 200-250
nukleotida. RNA yang mengalami penambahan poliA hanya molekul mRNA, sedangkan tRNA dan rRNA tidak mengalami penambahan poliA. Penambahan poliA dilakukan oleh enzim poli(A)polymerase yang ada dalam nukleus. Fungsi dari molekul poliA bagi mRNA adalah : a) meningkatkan stabilitas mRNA sehingga memiliki umur lebih panjang, b) meningkatkan efisiensi translasi mRNA, c) meningkatkan kemungkinan molekul mRNA untuk mengikat ribosom sehingga proses translasi lebih efisien.
pada sekuens AATAAA namun belum diketahui dengan pasti. Dikatakan berbeda karena pada fungi sangat jarang sekali ditemukan motif AATAAA.
Penambahan tudung (cap) mRNA adalah proses penambahan gugus metil pada ujung 5’ mRNA yang dikenal dengan sebutan tudung mRNA (mRNA cap). Gugus metil ini merupakan molekul 7-metilguanosin (m7G). Fungsi dari tudung mRNA ini adalah: a) melindungi mRNA dari degradasi, b) meningkatkan efisiensi translasi, c) meningkatkan pengangkutan mRNA dari nukleus ke sitoplasma, c) meningkatkan efisiensi proses splicing mRNA.
Pemrosesan rRNA dan tRNA dilakukan untuk menghasilkan molekul rRNA dan tRNA yang siap digunakan oleh sel. Molekul rRNA hasil transkripsi merupakan satu kesatuan molekul panjang rRNA yang belum siap dipakai. Molekul ini harus dipotong-potong sesuai dengan ukuran unit fungsionalnya. Misalnya pada rRNA hewan hasil transkrip merupakan molekul panjang dengan ukuran mencapai 45S. Molekul tersebut merupakan gabungan dari unit fungsional rRNA yang terdiri dari 28S, 18S dan 5,8S. Pada proses pemotongan rRNA ini tidak mengalami penyambungan kembali, karena molekul yang akan digunakan memang dengan ukuran-ukuran kecil tersebut.
Untuk molekul tRNA, juga disintesis dalam bentuk prekusor yang memiliki ukuran lebih panjang. Pemotongan prekusor tRNA masih akan menyisakan beberapa nukleotida pada ujung 5’ dan 3’. Sisa nukleotida pada ujung 5’ dipotong dengan enzim RNAse P, sedangkan pada ujung 3’ dipotong dengan beberapa enzim yakni: RNase D, RNase BN, RNase T, RNase PH, RNase II dan PNPase.
Produk akhir dari keseluruhan proses transkripsi adalah tiga molekul RNA yang berwujud mRNA (messenger RNA), rRNA (ribosomal RNA) dan tRNA (transfer RNA). Ketiga molekul tersebut dibentuk dalam nukleus (inti sel) dan selanjutnya akan di pindahkan ke sitoplasma sel.
2.2.2 Translasi Pada Hewan dan Fungi
a) Ribosom
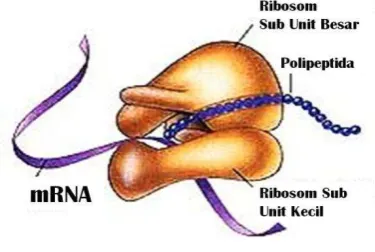
Ribosom merupakan organel sel yang menjadi tempat terjadinya sintesis protein. Struktur utama dari ribosom terdiri dari sub unit besar dan sub unit kecil. Sub unit besar ribosom terdiri dari 28S rRNA, 5.8S rRNA dan 5S rRNA dan 33 macam protein yang dikenal dengan nama L1-L33. Sub unit kecil ribosom terdiri dari 18S rRNA dan 21 macam protein yang dikenal dengan nama S1-S21. Ilustrasi bentuk ribosom ditampilkan pada Gambar 7.
Gambar 7. Bentuk Ribosom
b) Ribosomal RNA (rRNA)
Ribosomal RNA (rRNA) adalah salah satu produk transkripsi yang menjadi penyusun dari ribosom. Pada dasarnya, rRNA pada hewan terdiri dari 28S rRNA, 5,8S rRNA dan 5S rRNA untuk sub unit besar dan 18S rRNA untuk sub unit kecil. Sedangkan untuk fungi, terdapat 25S rRNA, 5,8S rRNA dan 5S rRNA untuk sub unit besar dan 18S rRNA untuk sub unit kecil.
c) Messenger RNA (mRNA)
Messenger RNA (mRNA) adalah duplikat dari molekul DNA yang berfungsi sebagai penyandi urutan asam amino spesifik dari suatu protein. Molekul ini yang dijadikan patokan dalam proses translasi menjadi protein. Sebagaimana telah dijelaskan pada bab sebelumnya mengenai perbedaan antara molekul RNA dan DNA, mRNA pada dasarnya terdiri dari urutan nukleotida RNA yang dapat diterjemahkan menjadi urutan asam amino berdasarkan kode genetik. Penyusunan kode genetik didasarkan pada urutan tiga nukleotida yang menyandi satu asam amino. Tiga nukleotida ini dikenal dengan nama kodon. Pada dasarnya, kode genetik yang digunakan pada hewan dan fungi adalah sama. Adapun detail kode gentik beserta dengan asam amino yang disandinya disajikan pada Tabel 4.
pada urutan pertama tingkat bahayanya bersifat moderat atau berada ditengah-tengah antara mutasi pada posisi ketiga dan kedua (Rodwell, 2003).
Tabel 4. Kode Genetik Universal
Perbedaan kode genetik yang ditemukan pada Candida albicans (sejenis fungi) adalah pada kodon CTG pada DNA inti dimana pada fungi ini diterjemahkan sebagai tryptophan sedangkan pada kode universal diterjemahkan sebagai leusin.
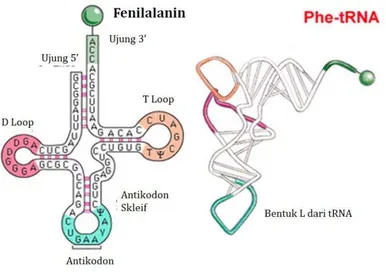
d) Transfer RNA (tRNA)
Gambar 8. Struktur dan bagian tRNA
Bagian tRNA yang berfungsi mengenali urutan nukleotida pada molekul mRNA adalah bagian antikodon loop. Sedangkan ujung 3’ merupakan tempat pengikatan asam amino yang akan dipolimerisasi menjadi protein. Molekul asam amino, akan diikat pada ujung Adenosin (3’) melalui ikatan ester antara gugus karboksil asam amino dengan gugus hidroksil pada 2’ atau 3’ dari molekul adenosine paling ujung. Setiap tRNA memiliki asam amino yang spesifik.
Pada fungi, terdapat kurang lebih 275 gen yang menyandi tRNA dalam genomnya, sedangkan pada manusia terdapat sekitar 20.848 gen yang menyandi tRNA baik pada DNA inti maupun DNA mitokondria.
e) Asam Amino
Asam amino adalam monomer dari protein yang terdapat dua puluh jenis. Asam amino ini yang digunakan dalam proses sintesis protein sebagai bahan baku utama. Asam amino yang digunakan pada proses sintesis protein adalah asam amino yang telah berada dalam tubuh. Asam amino bisa diperoleh dari proses degradasi protein yang kita makan atau hasil sintesis yang dilakukan oleh tubuh. Proses sintesis asam amino akan dijelaskan pada sub bab berikutnya.
1). Aktivasi Asam Amino
Proses pengaktifan asam amino yang dimaksud disini adalah proses perubahan asam amino menjadi aminoasil-tRNA dengan bantuan ATP. Pada tahap ini, asam amino akan berikatan dengan molekul tRNA pembawa yang spesifik untuk setiap asam amino. Proses aktivasi ini terjadi dalam dua tahap, yaitu:
Enzim Mg2+
Asam amino pertama yang akan ditambahkan saat inisiasi translasi pada eukariot adalah metionin. tRNA yang membawa asam amino awal ini berbeda dengan tRNA yang membawa metionin pada bagian tengah dari rantai polipetida. Molekul tRNA yang membawa awalan asam amino ini disebut sebagai tRNA inisiator (tRNAimet). Ribosom
bersama-sama dengan tRNAimet dapat menemukan posisi start kodon dengan cara
Berikut ini adalah ilustrasi proses inisiasi translasi :
Gambar 9. Inisiasi translasi
Pada translasi, terdapat beberapa macam faktor translasi, yaitu: 1, 2, eIF-3, eIF-5, dan eIF-6. Pada awal inisiasi translasi, molekul 18S rRNA akan berikatan dengan molekul mRNA. Proses ini memerlukan faktor translasi eIF-3. Amino asil tRNAimet berineteraksi dengan GTP dan eIF-2. Kompleks yang terbentuk akan berikatan
dan terbentuk kompleks 80S ribosom yang sempurna yang memiliki 2 sub unit utama. Faktor eIF-6 adalah faktor antiasosiasi yang mencegah terjadinya asosiasi antara sub unit 60S dengan 40S sebelum tebentuk kompleks inisiasi. Faktor eIF-4F merupakan faktor yang melekat pada struktur tudung pada ujung 5’.
3). Elongasi Translasi
Proses pemanjangan rantai polipeptida terjadi dalam tiga tahap utama: (1) pengikatan amino asil tRNA pada posisi A ribosom, (2) pemindahan rantai polipeptida yang tumbuh dari tRNA yang ada pada sisi P ke arah sisi A dengan membentuk ikatan peptida, (3) translokasi ribosom sepanjang mRNA ke posisi kodon selanjutnya yang ada di sisi A.
Di dalam kompleks ribosom, molekul tRNAimet menempati sisi P (peptidil), sisi
yang lain pada ribosom, yaitu sisi A (aminoasil), masih kosong pada saat awal sintesis protein. Berpasangannya triplet kodon inisiasi (AUG) pada mRNA dengan antikodon pada tRNAimet di tapak P menentukan urutan triplet kodon dan tRNAimet berikutnya
yang akan masuk ke tapak A. Pengikatan aminoasil-tRNA berikutnya, misalnya alanil- tRNAala, ke tapak A memerlukan protein-protein elongasi EF-Ts dan EF-Tu.
Pembentukan ikatan peptida antara gugus karboksil pada tRNAimet di tapak P dan
gugus amino pada alanil-tRNAala di tapak A dikatalisis oleh enzim peptidil transferase, suatu enzim yang terikat pada subunit ribosom 50S. Reaksi ini menghasilkan dipeptida yang terdiri atas metionin dan alanin yang terikat pada tRNAala di tapak A. Langkah berikutnya adalah translokasi, yang melibatkan (1) perpindahan met-ala tRNAala dari tapak A ke tapak P dan (2) pergeseran posisi mRNA pada ribosom sepanjang tiga basa sehingga triplet kodon yang semula berada di tapak A masuk ke tapak P. Dalam contoh ini triplet kodon yang bergeser dari tapak A ke P tersebut adalah triplet kodon untuk alanin. Triplet kodon berikutnya, misalnya penyandi serin, akan masuk ke tapak A dan proses seperti di atas hingga translokasi akan terulang kembali.
Berikut adalah ilustrasi dari proses pemanjangan rantai polipeptida:
4). Terminasi Translasi
Translasi akan berakhir pada waktu salah satu dari ketiga kodon terminasi (UAA,UGA,UAG) yang ada pada mRNA mencapai posisi A pada ribosom. Dimana RF1 yang mengenali kodon UAA atau UAG sehingga rantai kodon tersebut akan terlepas, kemudian RF2 akan mengenali kodon UAA atau UGA sehingga rantai kodon tersebut terlepas. Proses terminasi ditandai oleh terlepasnya mRNA, tRNA di tapak P, dan rantai polipeptida dari ribosom. Selain itu kedua subunit ribosom juga memisah, pada terminasi diperlukan aktivitas dua protein yang berperan sebagai faktor pelepas atau releasing factors, yaitu RF-1 dan RF-2 yang bekerja sama dengan RF-3. Berikut ini adalah ilustrasi proses terminasi translasi.
Gambar 11. Terminasi Translasi
5). Pengolahan Pasca Translasi
dari daerah yang terlindung di dalam ribosom ini lalu mengalami pelipatan membentuk konformasi tiga-dimensi polipeptida.
Protein berikatan dengan polipeptida nascent(yaitu polipeptida yang sedang dalam proses sintesis) dan memperantarai proses pelipatan tersebut. Mediator ini disebut chaperone karena mencegah terjadinya interaksi yang tidak sesuai. Pembentukan ikatan disulfida antara residu sistein juga berperan membentuk struktur tiga dimensi polipeptida (Toha, 2001).
Residu asam amino dapat mengalami modifikasi dengan penambahan berbagai jenis gugus fungsional. Asam amino terminal-N kadang-kadang mengalami asetilasi. Ke residu lisin dapat ditambahkan gugus metil. Residu prolin dan lisin dapat mengalami modifikasi melalui hidroksilasi, terutama pada kolagen. Karboksilasi merupakan modifikasi yang penting terutama untuk fungsi protein yang terlibat dalam pembekuan darah. Dapat dilakukan penambahan asam lemak yang membentuk regio hidrofobik untuk merekatkan protein ke membran . Dapat dilakukan pemindahan sebuah gugus ADPR dan NAD ke protein tertentu. Penambahan dan pengeluaran gugus fosfat (yang berikatan secara kovalen dengan residu serin, treonin,dan triosin) berfungsi untuk mengubah aktivitas banyak protein (misal enzim pada sintesis dan penguraian glikogen). Glikosilasi, penambahan gugus karbohidrat, merupakan modifikasi yang terutama terjadi pada protein yang akan disekresikan atau digabungkan ke membran (Lehninger, 1982).
2.2.3 Sintesis Asam Amino pada hewan dan fungi A. Sintesis Asam Amino pada Hewan
Asam amino merupakan komponen utama penyusun protein. Mengingat begitu banyaknya molekul tubuh ini yang tersusun oleh protein, maka proses regulasi (baik pendegradasian maupun sintesis protein) harus terus berjalan dalam tubuh guna menyeimbangkan proses-prose kimiawi dalam tubuh. Oleh karenanya, dibutuhkannya supplay asam amino dalam tubuh, baik yang disintesis sendiri oleh tubuh, maupun yang diperoleh tubuh melalui asupan makanan. Sebelum kita membahas mengenai proses sintesis asam amino dalam tubuh, perlu kita tinjau kembali bagaimana hubungan antara proses katabolisme dan anabolisme protein dalam tubuh. Berikut ini ilustrasi skema lengkap proses degradasi dan sintesis protein dalam tubuh hewan, serta hubungan diantara keduanya.
Gambar 12. Jalur metabolisme asam amino
Pada hewan (manusia), terdapat dua belas asam amino yang dapat dibentuk oleh tubuh, seperti ditampilkan pada table 4. Kedua belas asam amino yang non esensial ini, 9 diantaranya dapat dibentuk dari zat antara amfibolik dan 3 asam amino (sistein, tirosin dan hidroksilisin) dibuat dari asam amino yang esensial. Zat antara amfibolik pembentuk asam amino ini adalah : α-ketoglutarat (Glu, Gln, Pro,Hyp), oksaloasetat (Asp, Asn), 3-Fosfoglierat (Ser, Gly). Serin dan homosistein merupakan bahan baku pembentukan sistein. Fenil alanin merupakan precursor pembentukan tirosin. Mekanisme pembentukannya akan dibahas pada bagian selanjutnya.
Dalam proses sintesis asam amino ini, terdapat tiga enzim yang berperan sangat penting, yaitu : glutamat dehydrogenase, glutamin sintetase, amino transferase.
Kombinasi dari ketiga enzim ini dapat mengubah ion ammonium menjadi nitrogen α
-amino dari berbagai asam -amino.
Table 5. kebutuhan Asam amino manusia
No Asam amino Esensial Asam amino non esensial
Glutamat dan Glutamin. Asam amino glutamat dibentuk oleh reaksi aminasi reduktif α-ketoglutarat dikatalisis oleh glutamat dehydrogenase. Sesuai persamaan reaksi berikut ini:
Sedangkan asam amino glutamin dibentuk dari reaksi aminasi glutamat menjadi glutamin dikatalisis oleh glutamin sintetase. Sesuai persamaan reaksi berikut ini:
Alanin. Asam amino alanin dibentuk dari reaksi transaminasi piruvat menggunakan donor amina dari glutamat atau aspartat dan akan menghasilkan α -ketoglutarat atau oksaloasetat. Persamaan reaksinya adalah sebagai berikut:
Aspartat dan Asparagin. Transaminasi oksaloasetat membentuk aspartat.
Serin. Pembentukan serin terjadi melalui oksidasi gugus α-hidroksil dari 3-fosfogliserat menjadi asam okso, dan dilanjutkan dengan trans aminasi dan defosforilasi sehingga menghasilkan serin.
Glisin. Asam amino glisin dapat dibentuk dari reaksi yang dikatalisis oleh glisin aminotransferase dengan bahan baku glioksilat dan glutamat atau alanin. Selain itu, dapat juga dibentuk dari serin dan kolin.
Prolin. Dibentuk dari pembalikan reaksi katabolisme prolin dengan bahan baku glutamat.
Sistein. Sistein dibentuk dari metionin yang diubah menjadi homosistein, selanjutnya homosistein bersama dengan serin membentuk sistationin. Hidrolisis dari sistationin akan menghasilkan sistein dan homoserin.
Hidroksiprolin dan hidroksilisin. Kedua asam amino ini terdapat dalam kolagen. Asam amino ini tidak menjadi penyusun suatu protein sehingga keduanya mengalami tidak ditemukan dalam makanan dan akan dihidrolisis sempurna. Hidroksi prolin dibentuk dari prolin dan hidroksi lisin dibentuk dari lisin, pembentukan keduanya terjadi setelah asam amino prolin dan lisin tergabung dalam peptida.
Struktur selenosistein dan pembentukannya
B. Sintesis Asam amino pada Fungi
Berbeda halnya dengan hewan, yang hanya mampu membentuk 12 asam amino dalam tubuhnya. Fungi memiliki kemampuan yang luar biasa dimana dia bisa membentuk hampir semua asam amino secara mandiri. Mekanisme pembentukan asam amino pada fungi melibatkan proses enzimatis yang sangat rumit.
Pada dasarnya, sintesis asam amino pada yeast menggunakan beberapa senyawa intermediet dari glikolisis sebagai donor kerangka karbonnya. Sedangkan kerangka amin diperoleh dari glutamat. Glutamat dan glutamin merupakan asam amino yang sangat berperan penting pada proses sintesis asam amino pada yeast. Senyawa intermediet yang berperan dalam biosintesis asam amino pada yeast yaitu : intermediet glikolisis terdiri dari piruvat, 3-fosfogliserat, fosfoenol piruvat. Intermediet siklus asam sitrat : α-ketoglutarat, oxaloasetat. Intermediet siklus pentosa fosfat ribosa 5-fosfat dan eritrosa 4-fosfat.
Pengelompokkan dan sumber biosynthesis asam amino pada fungi disajikan pada Gambar 13.
Gambar 13. Kelompok dan sumber sintesis asam amino pada fungi (Lanjutan)
SINTESIS ASAM AMINO FAMILI α-KETOGLUTARAT
Glutamat. Pembentukan asam amino glutamat dikatalisis oleh reaksi enzim NADP glutamat dehydrogenase.
Sintesis Glutamin, Prolin dan Ornitin dari α-ketoglutarat
Arginin merupakan asam amino yang disintesis dari family α-ketolgutarat dengan jalur sintesis yang sangat panjang. Seperti terlihat pada reaksi pembentukan prolin, pada jalur kekanan dengan adanya glutamat akan membentuk ornitin. Ornitin inilah yang digunakan dalam sintesis arginine.
SINTESIS ASAM AMINO FAMILI PIRUVAT
Asam amino alanin disintesis dengan menggunakan asam amino glutamat sebagai penyumbang kerangka amin dan glutamat sebagai donor kerangka karbon.
Pada dasarnya Lisin adalah asam amino yang dibentuk dari oksaloasetat, akan tetapi, yeast memiliki jalur untuk membentuk lisin dari piruvat dengan sepuluh tahap reaksi.
Biosintesis leusin pada fungi terjadi dengan jalur piruvat, dengan menggunakan 8 tahapan reaksi sebagai berikut:
SINTESIS ASAM AMINO FAMILI OKSALOASETAT
Aspartat. Pembentukan aspartat melalui reaksi yang melibatkan senyawa oksaloasetat dan glutamat.
Asparagin dibentuk secara langsung dari asam aspartate, secara struktur, perbedaan antara aspartate dan asparagine hanya terletak pada gugus NH2. Sintesis
Treonin dibentuk dari oksaloasetat yang diubah menjadi L-Aspartat dan selanjunya menjadi L-Treonin.
Metionin dapat disintesis dari homoserin, serin, dan homo sistein. Berikut ini adalah jalur sintesis metionin dari ketiga senyawa asal tersebut.
Isoleusin dibentuk dengan senyawa awal adalah treonin yang mengalami deaminasi.
SINTESIS ASAM AMINO FAMILI SERIN-GLISIN
Serin. Asam amino serin dibentuk dari senyawa 3-fosfogliserat.
Glisin adalah asam amino yang paling sederhana dengan rantai samping hanya berupa satu atom hidrogen. Asam amino ini disintesis dari serin.
SINTESIS ASAM AMINO FAMILI FENILALANIN, TIROSIN, TRIPTOFAN
SINTESIS ASAM AMINO FAMILI HISTIDIN
Berikut ini adalah ringkasan dari proses sintesis asam amino pada yeast.
Gambar 14. Jalur sintesis asam amino pada yeast.
C. Ringkasan Perbandingan Sintesis asam amino pada hewan dan fungi.
Table 6. Perbandingan prekusor sisntesis asam amino pada hewan dan fungi
No Asam Amino Prekursor
Human Yeast
1 Glutamat α-ketoglutarat α-ketoglutarat
2 Glutamin Glutamat Glutamat
3 Alanin Piruvat Piruvat
4 Aspartat Oksaloasetat Oksaloasetat
5 Asparagin Aspartat Aspartat
6 Serin 3-fosfogliserat 3-fosfogliserat
7 Glisin
Glioksilat, glutamat/alanin, kholin, serin
3-fosfogliserat
8 Prolin Glutamat α-ketoglutarat
9 Sistein Metionin 3-fosfogliserat
10 Tirosin Fenil alanin Fosfoenol-piruvat,
eritrose 4-fosfat
11 Hidroksiprolin Prolin Tidak ada
12 Hidroksisistein Lisin Tidak ada
13 Valin Esensial Piruvat
14 Leusin Esensial Piruvat
15 Isoleusin Esensial Okasloasetat
16 Selenosistein Sistein Tidak ada
17 Arginin Esensial α-ketoglutarat
18 Histidin Esensial Pentose 5-fosfat
19 Fenilalanin Esensial Fosfoenol-piruvat,
eritrose 4-fosfat
20 Tryptophan Esensial Fosfoenol-piruvat,
eritrose 4-fosfat
21 Threonin Esensial Oksaloasetat
22 Metionin Esensial Oksaloasetat
23 Lysin Esensial Oksaloasetat
3 PENUTUP
3.1KESIMPULAN
Secara umum, proses sintesis protein baik pada hewan maupun fungi, terjadi melalui mekanisme yang sama. Tahapan dari proses ini yaitu transkripsi dan translasi. Akan tetapi terdapat beberapa perbedaan antara biosintesis protein pada hewan dan fungi, diantaranya yaitu:
1. Struktur sel fungi memiliki dinding sel, sedangkan hewan hanya memiliki membran sel.
2. Komponen Faktor Transkripsi TFIID pada hewan berbeda dengan fungi. Pada hewan terdapat 16 komponen penyusun faktor transkripsi TFIID sedangkan pada yeast terdapat 13 komponen penyusunnya.
3. Pada saat pasca transkripsi: molekul sinyal untuk proses poliadenilasi pada mamalia adalah rangkaian nukleotida AATAAA sedangkan pada fungi ada variasi pada sekuens AATAAA namun belum diketahui secara pasti. Dikatakan berbeda karena pada fungi sangat jarang sekali ditemukannya motif AATAAA. 4. Transkripsi gen kelas 1: faktor transkripsi SL1 merupakan faktor spesifik untuk
spesies tertentu. Jadi setiap spesies memiliki faktor SL1 yang berbeda-beda. 5. Perbedaan kode genetik yang ditemukan pada Candida albicans (sejenis fungi)
adalah pada kodon CTG pada DNA inti dimana pada fungi ini diterjemahkan sebagai tryptophan sedangkan pada kode universal diterjemahkan sebagai leusin. 6. Molekul rRNA Hewan memilik subunit 28S sedangkan pada fungi memiliki
25S.
7. Pada hewan terdapat 20.848 gen yang menyandi tRNA sedangkan pada fungi hanya terdapat 275 gen penyandi tRNA.
8. Hewan (manusia) hanya bisa mensintesis sekitar 12 jenis dari 20 asam amino, sedangkan fungi (yeast) bisa mensintesis 20 jenis asam amino.
DAFTAR PUSTAKA
Anonim. 2013. Transkription (Genetics). http://en.wikipedia.org/wiki/ Transcription_ (genetics). Akses 26-Sep-13 08:06
Azhar, T.N. 2008. Dasa-Dasar Biologi Molekuler. ISBN: 978-602-8323-22-2. Penerbit Widya Padjadjaran. Bandung
Murray, R.K., Graner,D.K, Rodwell, V.W, 2006. BiokimiaHarper (Harper Illustrated Biochemistry), terjemahan: Pendit,B.U. Penerbit EGC, Jakarta. Eisen, J. 2008. Twisted Tree of Life Award #2: Science Friday on the Five
Kingdoms. http://phylogenomics.blogspot.com/2008/10/twisted-tree-of-life-award-2-science.html. Akses 21-Sep-13 14:55
Rodwell V.W. 2003. Nucleotide. Harper’s Illustrated Biochemistry. Medical Publishing Division. New York, 296-330.
Lehninger. A.L. 1982. Dasar-Dasar Biokimia. Terjemahan: Meggy Thenawijaya. Erlangga. Jakarta.