II. TINJAUAN PUSTAKA
2.1. MIKROBIOTA DALAM SALURAN PENCERNAAN
Saluran pencernaan dimulai dari rongga mulut yang terdiri atas mulut, hidung, dan kerongkongan. Saluran pencernaan selanjutnya adalah lambung, usus kecil, usus besar dan berakhir di anus. Pada setiap bagian saluran pencernaan terdapat bakteri yang berbeda–beda. Terdapat perbedaan yang besar pada jumlah dan jenis populasi bakteri yang terdapat pada lambung, usus kecil, dan usus besar. Perbedaan ini terjadi karena adanya perbedaan waktu transit, sekresi, dan kesediaan nutrisi (Lambert dan Hull 1996; Guilliams 1999 diacu dalam Tamime 2005).
Usus merupakan sebuah ekosistem kompleks yang terdiri atas tiga komponen yang saling berhubungan yaitu sel inang, nutrisi, dan mikrobiota. Fungsi usus antara lain untuk proses pencernaan makanan, penyerapan zat gizi, dan pertahanan terhadap serangan dari luar. Komponen pertahanan usus terdiri dari atas tiga jenis yaitu mikrobiota, pertahananan mukosa, dan sistem imun internal (Bourlioux et al. 2002).
Usus halus merupakan habitat dari enterococci, enterobacteria, lactobacilli, bacteroides, dan clostridia. Jumlah mikroba meningkat dari 104-106 cfu/ml pada usus halus menjadi 1011-1012 cfu/ml pada usus besar (Salminen et al. 1998 diacu dalam Tamime 2005). Diperkirakan 95% dari semua sel hidup dalam tubuh manusia adalah bakteri usus besar (Gibson et al 2004). Sedangkan jumlah bakteri akhir di dalam feses didominasi oleh Bacteroides spp sebanyak 109 cfu/ml (Salminen et al. 2004).
Sebelum dilahirkan, saluran pencernaan bayi masih dalam keadaan steril. Pada saat bayi dikeluarkan melalui vagina, kolonisasi mikrobiotia natural pada bayi pertama kali berasal dari vagina maupun saluran pencernaan ibunya. Dalam perkembangannya, asupan makanan bayi berperan penting terhadap mikrobiota saluran pencernaannya. Selama masa menyusui, komposisi mikrobiota saluran pencernaan berkembang dalam jangka waktu yang singkat, dan didominasi oleh Bifidobacteria (Boehm dan Moro 2008).
Flora vagina dan saluran pencernaan ibu merupakan sumber bakteri bagi bayi yang akan berkolonisasi dalam saluran usus bayi yang didominasi oleh strain anaerob fakultatif seperti enterobacteria, koliform dan Lactobacillus. Setelah tidak menyusu, komposisi mikroflora secara berangsur-angsur berubah menyerupai mikroflora dewasa. Diperkirakan terdapat 500 spesies mikroba yang berbeda pada saluran pencernaan, akan tetapi dari segi jumlahnya, saluran pencernaan didominasi oleh 20 jenis mikroba. Mikrobiota tersebut antara lain Bacteroides, Lactobacillus, Clostridium, Fusobacterium, Bifidobacterium, Eubacterium, Peptococcus, Peptostreptococcus, Escherichia, dan Veillonella termasuk bakteri yang dapat memberikan efek menguntungkan yaitu Bifidobacteria and Lactobacilli (Harish dan Varghese 2006).
Lactobacillus dan Bifidobacterium merupakan bakteri penghuni normal ekosistem kompleks saluran pencernaan. Lactobacillus memiliki kemampuan untuk menempel pada sel inang, untuk mengeluarkan atau mengurangi bakteri patogen, dan menghasilkan asam, hidrogen peroksida, dan bakteriosin yang mampu menghambat pertumbuhan bakteri patogen. Keberadaan Lactobacillus dalam saluran pencernaan berpotensi memberikan efek kesehatan bagi inang seperti mencegah terjadinya infeksi saluran pencernaan dan meningkatkan respon imun (Sharma et al. 2005).
Profil saluran pencernaan pada manusia dewasa memperlihatkan terjadinya perkembangan populasi mikroba yang dapat mengatasi tekanan psikologis dan mikrobiologis. Kondisi yang stabil memberikan ketahanan bagi inang, atau yang sering disebut dengan barrier effect, kemampuan melawan invasi mikroba, baik yang patogen maupun yang tidak berbahaya. Mikrobiota dalam saluran pencernaan harus mampu beradaptasi untuk berkompetisi mendapatkan nutrisi dan tempat dengan
mikroba pendatang, yang mungkin dapat menghambat dengan menghasilkan komponen penghambat (Alderbeth et al. 2000).
Keberadaan mikroorganisme dalam saluran pencernaan ditentukan oleh interaksi dengan lingkungan sekitarnya dan pengaruh dari sekitarnya yang mampu mendukung mikroorganisme tersebut melawan kompetitornya. Hal ini dapat dicapai setelah melalui mekanisme yang sangat banyak, seperti meningkatnya suasana anaerobik atau melalui komponen yang mengganggu, contohnya asam dan substansi antimikroba. Komponen-komponen tersebut memberikan efek menguntungkan bagi inang (Fooks dan Gibson 2002).
Fungsi utama dari mikrobiota usus yaitu aktivitas metabolik yang menyebabkan penyimpanan energi dan nutrisi, efek nutrisi terhadap epitel usus, dan perlindungan terhadap serangan bakteri yang merugikan (Harish dan Varghese 2006). Fungsi penting lainnya dari mikrobiota dalam saluran pencernaan adalah produksi vitamin B dan K, yang dibuktikan dalam studi dimana hewan percobaan yang disuplementasi dengan vitamin B dan K untuk memenuhi kebutuhannya setara dengan hewan percobaan dengan mikrobiota (Hooper et al. 2002).
Mikroflora saluran pencernaan merupakan bagian penting dalam dinding pertahanan saluran usus. Komposisi awal dari mikroflora saluran pencernaan yang berkembang merupakan faktor penentu perkembangan fungsi pertahanan saluran pencernaan normal. Penyimpangan spesifik pada mikroflora dalam saluran pencernaan dapat menyebabkan penyakit pada inang. Mekanisme pertahanan mukosa saluran usus yaitu peran lumen dan mukosa yang membatasi kolonisasi bakteri patogen pada permukaan mukosa. Mikrobiota saluran pencernaan normal dapat mencegah pertumbuhan yang berlebihan dari bakteri patogen dalam saluran pencernaan (Harish dan Varghese 2006).
2.2. BAKTERI ASAM LAKTAT SEBAGAI PROBIOTIK
Probiotik didefinisikan sebagai sediaan sel mikroba hidup yang memiliki pengaruh menguntungkan terhadap kesehatan dan kehidupan inangnya (Schmid et al. 2006). Efek positif dari aktivitas probiotik terbagi menjadi tiga aspek yaitu nutrisi, fisiologi, dan antimikroba. Aspek nutrisi berasal dari penyediaan enzim yang membantu metabolisme penyerapan laktosa (laktase), sintesis beberapa jenis vitamin (vitamin K, asam folat, piridoksin, asam pantotenat, biotin, dan riboflavin), dan dapat menghilangkan racun hasil metabolit komponen makanan di usus (Naidu dan Clemens 2000).
Aspek fisiologis meliputi kemampuan untuk menjaga keseimbangan komposisi mikrobiota usus sehingga menekan resiko infeksi penyakit dan menstimulasi sistem kekebalan tubuh. Aspek kemampuan antimikroba dinyatakan melalui kemampuan memperbaiki ketahanan terhadap patogen (Naidu dan Clemens 2000). Probiotik dapat meningkatkan pertahanan usus terhadap patogen karena probiotik berperan sebagai adjuvant sistem imun dan memperkuat lapisan mukosa (Walker 2008).
Mekanisme probiotik dalam memberikan efek positif pada saluran pencernaan memang kurang dimengerti. Akan tetapi secara garis besar, menurut Sartor (2004), mekanismenya antara lain : (1) adherence/ menempel dan membentuk koloni di dalam saluran pencernaan, (2) menekan pertumbuhan atau mengikat/ menyerang bakteri patogen dan memproduksi substansi antimikroba, (3) meningkatkan fungsi pertahanan saluran pencernaan, (4) mengontrol transfer antigen, dan (5) menstimulasi sistem imun mukosa dan sistemik pada inang.
FAO/WHO (2001) menyatakan probiotik adalah mikroorganisme hidup yang masuk dalam jumlah yang cukup sehingga dapat memberikan manfaat kesehatan bagi inang. Jumlah yang cukup menurut FAO/WHO adalah 106-108 cfu/g dan diharapkan dapat berkembang menjadi 1012 cfu/g di dalam kolon. International Dairy Federation (IDF) memberikan standar acuan untuk minuman probiotik hidup sebanyak 106 cfu/ml pada produk akhir (Indratingsih et al. 2004).
Produk probiotik dapat diklaim memberi keuntungan kesehatan harus mengandung bakteri probiotik dengan konsentrasi minimal 106 cfu/ml atau gram, sebagai dosis minimum terapi per harinya disarankan mengonsumsi 108-109 cfu/ml (Shah 2000). Akan tetapi, beberapa studi menunjukkan viabilitas probiotik yang berbeda-beda dalam fermentasi susu, khususnya yogurt (La Torre et al. 2003). Beberapa faktor diklaim mempengaruhi viabilitas kultur probiotik dalam fermentasi susu, seperti pH produk akhir, kesediaan nutrisi, oksigen terlarut dan penyerapan oksigen melalui kemasan produk (Tamime 2005).
Jenis bakteri yang banyak digunakan sebagai probiotik adalah bakteri asam laktat (BAL), seperti dari genus Lactobacillus dan Bifidobacteria (Winarno 2003). Menurut Salminen et al. (2004), beberapa syarat yang harus dipenuhi oleh BAL yang berfungsi sebagai mikroba probiotik antara lain:
1) Probiotik harus non patogenik yang mewakili mikrobiota normal usus dari inang tertentu, dan masih aktif pada kondisi asam lambung serta konsentrasi garam empedu yang tinggi dalam usus
2) Probiotik yang baik mampu tumbuh dengan cepat dalam jumlah yang tinggi pada usus 3) Probiotik yang ideal dapat mendiami beberapa bagian dari saluran usus untuk sementara 4) Probiotik dapat memproduksi asam-asam organik secara efisien dan memiliki sifat
antimikroba terhadap bakteri merugikan
5) Mudah diproduksi, mampu tumbuh dalam sistem produksi skala besar dan hidup selama kondisi penyimpanan.
Ketahanan terhadap asam lambung merupakan syarat penting suatu organisme untuk dapat menjadi probiotik karena pH asam lambung sekitar 2 (Almatsier 2005). Pada BAL terjadi perubahan yang dinamis pada pH intraseluler seiring dengan terjadinya penurunan pH ekstraseluler sehingga tidak terjadi gradien proton yang besar. Gradien proton yang besar dapat menyebabkan akumulasi anion, asam organik dalam sitosol yang bersifat toksik bagi sel tersebut. Selain itu gradien proton membutuhkan energi yang besar untuk translokasi proton sehingga sangat merugikan (Siegumfeldt et al. 2000).
BAL yang telah melewati dua barrier yaitu asam lambung dan empedu akan menuju ke usus. Bakteri tersebut berkembang pada saat menuju ke jejunum dan akhirnya sampai ke kolon. Waktu transit yang lama memungkinkan bakteri untuk berkembang. Selain itu terdapat nutrisi sisa-sisa makanan yang tidak diserap, material dari inang (mukus dan sel mati), dan metabolit bakteri yang dapat digunakan sebagai sumber makanan (Bourlioux et al. 2002).
Selain ketahanannya dalam saluran pencernaan, aktivitas antimikroba terhadap berbagai bakteri patogen juga merupakan syarat yang penting bagi suatu organisme untuk menjadi probiotik. Bakteri seperti Bifidobacteria spp. dan Lactobacillus spp. memiliki aktivitas penghambatan terhadap bakteri pathogen Listeria monocytogenes, Escherichia coli, Salmonella spp. (Jenie 2003).
Probiotik dianggap sukses bila mampu berkolonisasi pada saluran usus, paling tidak untuk sementara, dengan cara menempel pada mukosa usus. Dengan penempelan itu, probiotik juga mampu mencegah penempelan bakteri patogen seperti bakteri koliform dan Clostridia, dan mampu menstimulasi penghilangan patogen tersebut dari jalur infeksi saluran pencernaan (Lee et al. 2000).
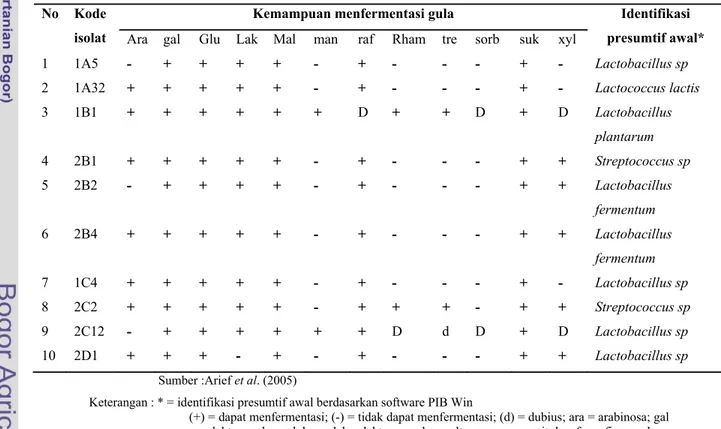
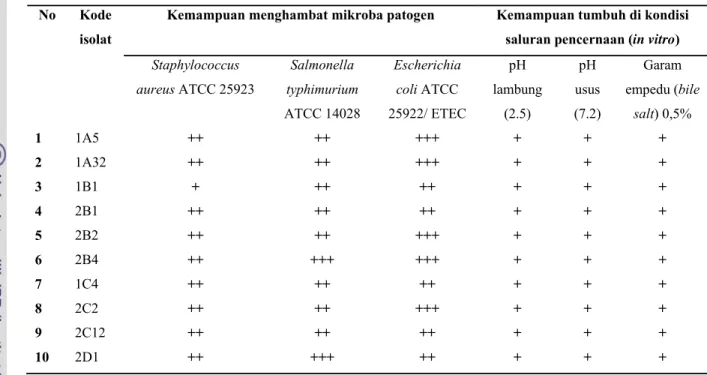
Arief et al. (2008) menyatakan BAL golongan Lactobacillus, Lactococcus, dan Streptococcus dari daging sapi bangsa Peranakan Ongol yang dijual di berbagai pasar tradisional daerah Bogor mampu bertahan pada kondisi saluran pencernaan seperti pH saluran pencernaan dan garam empedu, serta memiliki aktivitas antimikroba terhadap bakteri patogen. Kemampuan bakterisidal terhadap bakteri patogen ini disebabkan bakteri ini mampu menghasilkan senyawa bioaktif asam laktat, asam asetat, dan senyawa bakteriosin. Sifat dasar kesepuluh bakteri asam laktat yang diisolasi dari daging sapi tersebut dan kemampuannya sebagai probiotik dinyatakan pada Tabel 1, 2 dan 3.
Tabel 1. Morfologi isolat indigenus bakteri asam laktat No Kode isolat Bentuk Pertumbuhan di suhu Pertumbuhan di NaCl 6,5% Menghasilkan NH3 dari arginin Menghasilkan gas dari glukosa 15oC 45oC 1 1A5 Batang + + + - - 2 1A32 Kokus + + + + - 3 1B1 Batang + + + - + 4 2B1 Kokus + + + - - 5 2B2 Batang + + + + - 6 2B4 Batang + + + - + 7 1C4 Batang + + + + - 8 2C2 Batang + + + - - 9 2C12 Kokus + + + + - 10 2D1 Batang + + + - - Sumber : Arief et al. (2005)
Tabel 2. Hasil fermentasi terhadap beberapa jenis gula sederhana No Kode
isolat
Kemampuan menfermentasi gula Identifikasi presumtif awal* Ara gal Glu Lak Mal man raf Rham tre sorb suk xyl
1 1A5 - + + + + - + - - - + - Lactobacillus sp 2 1A32 + + + + + - + - - - + - Lactococcus lactis 3 1B1 + + + + + + D + + D + D Lactobacillus plantarum 4 2B1 + + + + + - + - - - + + Streptococcus sp 5 2B2 - + + + + - + - - - + + Lactobacillus fermentum 6 2B4 + + + + + - + - - - + + Lactobacillus fermentum 7 1C4 + + + + + - + - - - + - Lactobacillus sp 8 2C2 + + + + + - + + + - + + Streptococcus sp 9 2C12 - + + + + + + D d D + D Lactobacillus sp 10 2D1 + + + - + - + - - - + + Lactobacillus sp Sumber :Arief et al. (2005)
Keterangan : * = identifikasi presumtif awal berdasarkan software PIB Win
(+) = dapat menfermentasi; (-) = tidak dapat menfermentasi; (d) = dubius; ara = arabinosa; gal = galaktosa; glu = glukosa; lak = laktosa; mal = maltosa; man= manitol; raf = rafinosa; rham = rhamnosa; tre = trehalosa; sorb = sorbitol; suk = sukrosa; xyl = xylosa
Tabel 3. Sifat dasar probiotik isolat indigenus bakteri asam laktat No Kode
isolat
Kemampuan menghambat mikroba patogen Kemampuan tumbuh di kondisi saluran pencernaan (in vitro) Staphylococcus aureus ATCC 25923 Salmonella typhimurium ATCC 14028 Escherichia coli ATCC 25922/ ETEC pH lambung (2.5) pH usus (7.2) Garam empedu (bile salt) 0,5% 1 1A5 ++ ++ +++ + + + 2 1A32 ++ ++ +++ + + + 3 1B1 + ++ ++ + + + 4 2B1 ++ ++ ++ + + + 5 2B2 ++ ++ +++ + + + 6 2B4 ++ +++ +++ + + + 7 1C4 ++ ++ ++ + + + 8 2C2 ++ ++ +++ + + + 9 2C12 ++ ++ ++ + + + 10 2D1 ++ +++ ++ + + + Sumber : Arief et al. (2008)
Hasil penelitian Arief et al. (2008) menunjukan terdapat 10 BAL isolat indigenus yang mempunyai kemampuan bertahan pada pH lambung yaitu pada pH 2.5 dan pH usus yaitu pada pH 7.2, serta pada kondisi garam empedu 0.5% sesuai dengan kondisi saluran pencernaan. Selain itu, BAL tersebut memiliki aktivitas penghambatan yang baik terhadap tiga jenis bakteri enteropatogenik yaitu Salmonella thypimurium ATCC 14028, Escherichia coli ATCC 25922 (ETEC), dan Staphylococcus aureus ATCC 25923.
Berdasarkan kriteria dasar probiotik oleh FAO/WHO (2002) yaitu kemampuannya untuk bertahan pada kondisi lambung (pH rendah) dan adanya garam empedu, serta penghambatannya terhadap bakteri patogen, maka kesepuluh BAL isolat indigenus tersebut dapat dinyatakan sebagai probiotik.
2.3. PROBIOTIK SEBAGAI IMUNOMODULATOR
Probiotik didefinisikan sebagai mikroorganisme hidup yang pada jumlah yang sudah diatur dapat memberikan efek yang menguntungkan bagi kesehatan inang (Reid et al. 2003). Strain yang paling sering digunakan dalam penelitian adalah Lactobacillus dan Bifidobacterium, antara lain Lactobacillus achidophilus, Lactobacillus casei, Lactobacillus fermentum, Lactobacillus plantarum, Lactobacillus reuteri, Bifidobacterium infantis, Bifidobacterium breve, Bifidobacterium animalis, Bifidobacterium adolescentis, dan Bifidobacterium longum. Bakteri–bakteri tersebut termasuk bakteri gram positif, fakultatif yang normal terdapat dalam usus besar manusia dan sebagian besar merupakan mikroflora anaerobik (Liong 2007).
Probiotik merupakan preparasi sel bakteri hidup yang mampu memberikan efek menguntungkan pada kesehatan inang. Banyak BAL sebagai probiotik, digunakan untuk memperbaiki mikroflora usus yang terganggu fungsinya dan permeabilitas saluran pencernaan yang tidak normal (Lee et al. 2000).
Kullisaar et al. (2003) menyatakan bahwa mengonsumsi susu fermentasi yang mengandung L. fermentum ME-3 memperlihatkan efek antioksidatif dan antiaterogenik.
Bakteri probiotik memproduksi berbagai jenis substansi yang mampu menghambat bakteri gram positif dan gram negatif. Komponen tersebut tidak hanya mengurangi jumlah sel hidup, tetapi juga memberikan dampak terhadap hasil metabolisme bakteri atau produksi toksin. Fakta tersebut membuktikan bahwa probiotik dapat memberikan perlindungan terhadap penyakit saluran usus melalui stimulasi terhadap sistem imun spesifik dan non-spesifik. Pemberian oral Lactobacillus sp. selama terjadinya diare rotavirus akut berkaitan dengan peningkatan respon imun terhadap rotavirus. Mekanisme stimulasi sistem imun belum dimengerti, akan tetapi komponen sel permukaan spesifik atau sel lapisan dapat berperan sebagai adjuvant (agen yang dapat menstimulasi sistem imun) dan meningkatkan respon imun humoral. Studi lain menyatakan penghambatan patogen oleh probiotik disebabkan adanya kompetisi terhadap nutrisi dan tempat reseptor, yang akhirnya probiotik menggunakan nutrisi yang ada, sehingga tidak dapat dikonsumsi oleh patogen (Rolfe 2000).
Sistem imun manusia sebagian besar dirancang untuk menghancurkan mikroorganisme asing seperti virus dan bakteri. Manusia sehat memiliki dua mekanisme imun, sistem imun spesifik (acquired), yang memberikan respon pada antigen spesifik dan ditingkatkan dengan adanya paparan yang berulang-ulang; dan sistem imun non-spesifik (innate), yang tidak membutuhkan stimulasi, tidak meningkat dengan adanya paparan berulang-ulang, dan memiliki mekanisme yang menyertakan pertahanan fisik (Meydani 2000). Kedua sistem imun ini mampu mempengaruhi respon imun sistemik dan mukosa (Galdeano dan Perdigon 2006).
Sistem imun innate dan adaptive merupakan dua bagian yang saling tergantung dari satu kesatuan sistem imun. Pada tingkat mukosa saluran pencernaan, respon imun innate tidak hanya menjadi garis pertama pertahanan melawan mikroorganisme patogen, akan tetapi juga memberikan sinyal biologi untuk menginstruksikan sistem imun adaptive agar mendapatkan respon tersebut. Pada sistem imun, bakteri probiotik berperan merangsang respon imun mukosa saluran pencernaan (Galdeano dan Perdigon 2006).
Pemberian Lactobacillus dan Bifidobacterium secara oral memperlihatkan terjadinya peningkatan respon antibodi pada ovalbumin dan respon imunoglobulin A (IgA) terhadap toksin kolera pada tikus percobaan, meningkatkan sel yang mensekresi antibodi spesifik terhadap rotavirus pada anak dengan diare rotavirus akut, dan meningkatnya jumlah sel yang mensekresi antibodi terhadap β-laktoglobulin pada tikus yang masih menyusu (Isolauri et al. 2001). Liong (2007) menyatakan peningkatan jumlah sel penghasil IgA di mesenteric node kelenjar getah bening pada tikus percobaan, mengawali adanya kombinasi vaksin laktokokal yang dapat merangsang respon sistemik dan respon imun mukosa.
Sumber : Saulnier et al. (2009)
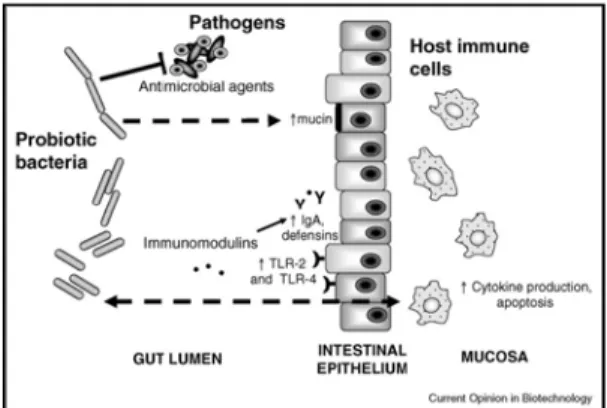
Probiotik dapat menstimulasi respon imun dengan meningkatkan produksi antibodi mukosa, mendorong ekspresi sitokin pro-inflamatori, dan meningkatkan produksi defensin inang (Gambar 1). Efek supresif menunjukan penurunan ekspresi sitokin, inflamasi sistemik, proliferasi sel, dan peningkatan apoptosis. Protein permukaan merupakan faktor kunci dalam imunomodulasi. Sebagai contoh, agregasi-kompeten L.crispatus merangsang ekspresi reseptor Toll-like receptor-2 (TLR-2)dan Toll-like receptor-4 (TLR-4) di permukaan sel epitelium mukosa kolon tikus percobaan. Probiotik juga mensekresikan faktor yang bertanggung jawab untuk memodulasi respon imun. Sebagai contoh, faktor yang disekresikan oleh L.reuteri menurunkan ekspresi inti gen factor-kB-dependent, yang menghasilkan berkurangnya proliferasi sel dan meningkatnya aktivitas protein kinase yang mengaktivasi mitogen yang mampu merangsang apoptosis (Saulnier et al. 2009).
Beberapa penelitian menyatakan pengaruh probiotik dalam sistem imun manusia. Dalam penelitian tersebut dinyatakan probiotik terdiri dari komponen dinding sel seperti peptidoglikan (30-70% dari total dinding sel), polisakarida, teichoic acid, yang merupakan komponen imunostimulator. Probiotik juga mengandung peptidoglikan yang dilepaskan dari dinding sel bakteri pada usus, merangsang aktivitas adjuvant di permukaan mukosa dan mengakibatkan adanya respon imun (Meydani 2000). Adanya hubungan antara probiotik dan materi molekuler seperti peptidoglikan, lipotechoic acid, dan DNA bakteri, membuktikan adanya materi reseptor (TLR) yang ada di permukaan sel imunokompeten yang dapat memicu dilepasnya sejumlah sitokin yang dapat meningkatkan respon imun (Gill dan Guarner 2004).
Efek modulasi dari sitokin pada sel imun termasuk merangsang produksi interferon-γ oleh sel T, meningkatkan jarak penghubung fagosit mikroba, menambah vaksin imunogenisitas, meningkatkan aktivitas mikrobisidal makrofag, dan menggunakan efek sitotoksin melawan sel tumor (Gill 2003).
2.4. PREBIOTIK
Prebiotik merupakan karbohidrat yang tidak dapat dicerna oleh enzim-enzim pencernaan usus manusia, tetapi menguntungkan terhadap penghuni bakteri kolon, dengan cara meningkatkan pertumbuhan dan keaktifan satu atau lebih jenis bakteri baik yang berada dalam kolon (Winarno 2003). Menurut Zakaria (2003), prebiotik merupakan pangan yang dapat memacu pertumbuhan bakteri probiotik, agar dapat diperoleh kondisi fisiologis dan metabolik yang dapat memberikan perlindungan pada saluran pencernaan, khususnya usus halus dan kolon.
Prebiotik adalah komponen pangan yang tidak dapat dicerna dan dapat memberikan efek yang menguntungkan bagi inang karena prebiotik merangsang proliferasi atau aktivitas populasi bakteri yang diinginkan di dalam kolon secara selektif (Mattila-Sandholm et al. 2002).
FAO (2007) menyatakan bahwa prebiotik adalah komponen pangan tak hidup yang memberi keuntungan kesehatan inang yang berasosiasi dengan memodulasi mikrobiota. Peraturan FAO (2007) juga menegaskan bahwa prebiotik bukan merupakan organisme atau obat, dapat dikarakterisasi secara kimia, dan aman sebagai pangan.
Bahan pangan akan dikelompokkan sebagai prebiotik bila memenuhi persyaratan berikut yaitu tidak terhidrolisa atau terserap pada saluran pencernaan bagian atas sehingga dapat mencapai kolon tanpa perubahan struktur atau diekskresikan dalam feses (Gibson et al. 2004), dan berperan sebagai substrat yang secara selektif dapat menstimulir pertumbuhan bakteri yang menguntungkan pada kolon (Gmeiner et al. 2000). Selain itu menurut Lisal (2005), bahan pangan dapat digolongkan sebagai prebiotik jika mampu mengubah keseimbangan flora usus besar ke arah komposisi yang menguntungkan kesehatan dan merangsang timbulnya efek-efek luminal (lokal) dan sistemik yang menguntungkan inang.
Prebiotik dapat berperan sebagai alternatif untuk probiotik atau sebagai kofaktor probiotik. Karbohidrat kompleks yang melalui usus kecil menuju kolon tersedia untuk beberapa bakteri kolon namun sebagian besar bakteri yang terdapat di kolon tidak memanfaatkannya. Laktosa, galaktooligosakarida, fruktooligosakarida, inulin dan hasil hidrolisanya, maltooligosakarida dan pati resisten merupakan prebiotik yang umumnya digunakan dalam nutrisi manusia. Produk akhir utama pada metabolisme karbohidrat adalah asam lemak rantai pendek (ALRP), yang disebut asetat, butirat, dan propionat, yang selanjutnya akan digunakan organisme dalam inang sebagai sumber energi (Harish dan Varghese 2006).
Analisis secara in vitro dan in vivo menunjukkan bahwa prebiotik tidak dicerna oleh enzim, tetapi difermentasi oleh bakteri anaerob dalam usus besar. Prebiotik yang telah difermentasi dalam usus besar menghasilkan ALRP, menstimulasi pertumbuhan berbagai bakteri termasuk Lactobacilli dan Bifidobacteria, dan dapat menghasilkan gas. Fortifikasi menggunakan Bifidobacteria/Lactobacilli dengan prebiotik dapat memperbaiki efek perlindungan usus besar terhadap berbagai mikroorganisme patogen dalam usus (Molis et al. 1996 diacu dalam Wang 2009).
Pada awalnya yang disebut sebagai bahan pangan prebiotik adalah serat pangan yang memacu pertumbuhan Bifidobacteria yaitu fruktooligosakarida (FOS), baik yang sintetik maupun yang diisolasi dari bahan pangan atau tanaman. Dengan berkembangnya penelitian mengenai fungsi FOS, diketahui bahwa FOS sebagai substrat Bifidobacteria, mampu mempercepat pertumbuhan bakteri ini sehingga menghasilkan metabolit-metabolit yang berfungsi sebagai penjaga kesehatan usus halus dan kolon, terutama melalui mekanisme antagonisme dengan bakteri patogen, metabolit ALRP, dan peningkatan respon imun pada usus halus (Zakaria 2003).
Prebiotik yang umum digunakan adalah inulin dan FOS, yang merupakan serat yang dapat larut dan dapat difermentasi (Gibson et al. 2004). Inulin dan FOS sering digunakan dalam studi secara in vivo karena resisten terhadap pencernaan asam lambung dan enzim pankreas (Cummings et al. 2001). FOS biasanya digunakan untuk formula produk bayi, yogurt dan produk pangan dan suplemen lainnya (Kaplan dan Hutkins 2000).
FOS secara kimiawi adalah senyawa β-D-fruktans rantai pendek atau sedang, yang terikat dengan ikatan β-2-1 glikosidik, yang tidak dapat diuraikan oleh enzim pencernaan mamalia. Dibandingkan dengan karbohidrat simpleks maupun kompleks lainnya, FOS difermentasikan secara selektif oleh hampir semua strain Bifidobacteria. Bila FOS dikonsumsi dalam jumlah yang cukup banyak maka FOS secara dramatik dan konsisten merangsang proliferasi Bifidobacteria menjadi mikroflora yang predominan dalam kolon (Lisal 2005) .
FOS memiliki nilai DP (derajat polimerisasi) lebih rendah daripada inulin, yaitu berkisar antara 2-8 (Franck dan De Leenheer 2005). FOS menunjukkan efek bifidogenik yang berbeda pada kondisi yang berbeda. Pada studi secara in vitro menunjukkan bahwa FOS dan inulin menghasilkan efek bifidogenik yang optimum pada pH 6.8 dan 1 g/100 ml karbohidrat, yang setara dengan 4 g/hari (Tungland 2003 diacu dalam Tamime 2005). Selain efek bifidogenik, FOS juga menambah nutrisi yang dapat mempengaruhi parameter fisiologis pencernaan seperti pH kolon dan stool bulking, yang dapat menggolongkan prebiotik sebagai serat (dietary fiber) (Roberfroid 1997 diacu dalam Tamime 2005).
Penggabungan prebiotik dan probiotik terbukti dapat meningkatkan keuntungan bagi kesehatan inang. Misalnya gabungan inulin (FOS) dengan Bifidobacterium longum terbukti dapat menurunkan risiko kelainan pre-neoplastik kolon lebih efektif dibandingkan hanya dengan perlakuan probiotik atau prebiotik saja pada tikus percobaan. Penambahan pati jagung yang kaya amilosa (Resistant Starch 2) ke dalam suatu preparat probiotik akan mempertahankan densitas probiotik yang hidup dibandingkan tanpa penambahan RS2 (Lisal 2005).
Meskipun FOS dapat ditemukan pada bermacam-macam pangan yang menjadi makanan kita sehari-hari, seperti asparagus, pisang, gandum, bawang putih, artichoke (tumbuhan yang bunganya dimakan sebagai sayuran) dan chicory (tanaman yang akarnya dibakar sebagai pengganti kopi), namun jumlah FOS dalam pangan tersebut sangat kecil. Jumlah rata-rata asupan harian tiap individu dari sumber pangan tersebut berkisar antara 2-10 g di Eropa, dan antara 1-4 g di Amerika serikat (van Loo et al. 1995 diacu dalam Tamime 2005).
Peraturan mengenai standar jumlah prebiotik yang dikonsumsi belum ada karena umumnya konsumsi prebiotik tergantung pada kebiasaan penduduk suatu negara (FAO 2007). Dosis konsumsi harian 5-8 g FOS/GOS dapat memberikan efek prebiotik pada orang dewasa. Surono (2004) menyarankan jumlah prebiotik yang efektif adalah 1-3 g per hari untuk anak-anak dan 5-15 g per hari untuk orang dewasa. Indonesia memiliki regulasi tentang prebiotik dalam Peraturan Pangan Fungsional yang dikeluarkan oleh BPOM tahun 2005, namun regulasi jumlah asupan prebiotik tidak dijelaskan.
2.5. YOGURT SINBIOTIK
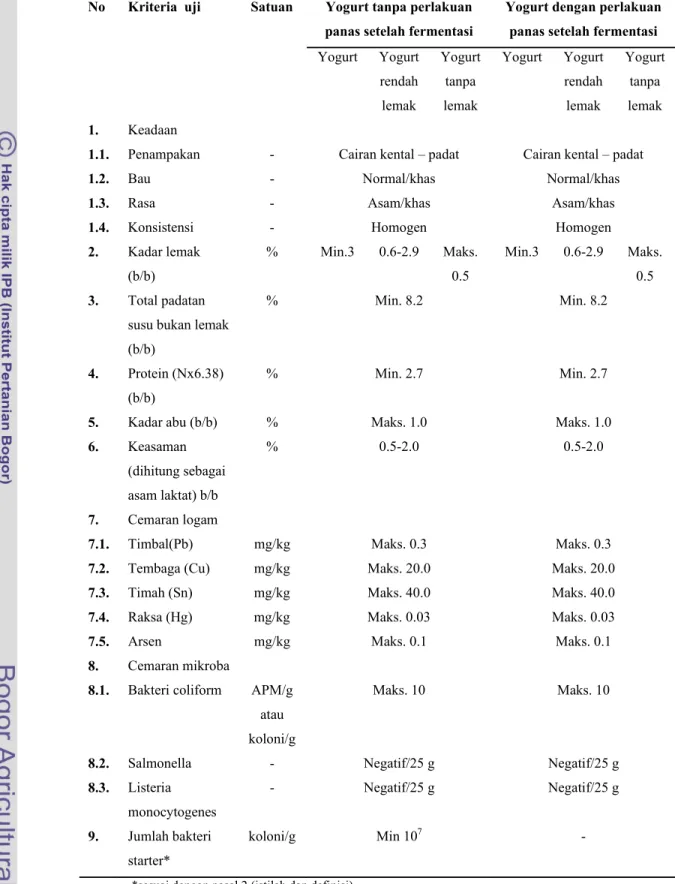
Menurut SNI (2009), definisi yogurt adalah produk yang diperoleh dari fermentasi susu dan atau susu rekonstitusi dengan menggunakan bakteri Lactobacillus bulgaricus dan Streptococcus thermophillus dan atau bakteri asam laktat lain yang sesuai, dengan/tanpa penambahan bahan pangan lain dan bahan tambahan pangan yang diizinkan. Syarat mutu yogurt menurut SNI (2009) dapat dilihat pada Tabel 4.
Proses fermentasi dapat terjadi karena pada susu sapi terdapat protein susu (kasein) dan gula susu (laktosa). Laktosa digunakan oleh kedua starter bakteri di atas sebagai sumber karbon dan energi utama untuk pertumbuhanya. Proses fermentasi tersebut menyebabkan laktosa berubah menjadi asam piruvat, yang selanjutnya diubah menjadi asam laktat. Asam laktat menyebabkan penurunan pH susu, atau meningkatkan keasaman. Akibatnya kasein menjadi tidak stabil dan terkoagulasi (menggumpal), membentuk gel yogurt, berbentuk setengah padat (semi padat), dan menentukan tekstur yogurt. Selain itu asam laktat juga berfungsi memberikan ketajaman rasa asam, dan menimbulkan aroma khas pada yogurt (Santoso 2009).
Mikroflora yang terdapat dalam yogurt merupakan kelompok BAL. Bakteri yang umum digunakan adalah Lactobacillus bulgaricus dan Streptococcus thermophilus (Silvia 2002). Streptococcus thermophilus merupakan satu-satunya spesies dalam genusnya yang digunakan sebagai kultur starter dalam produk olahan susu. Organisme ini termasuk dalam termotoleran dan digunakan dalam fermentasi produk olahan susu yang membutuhkan suhu lebih tinggi dalam inkubasi yaitu pada suhu 35-43oC. Pada industri produk olahan susu, organisme ini sering disebut dengan kokus. Biasanya kultur S.thermophilus menghasilkan koagulat yang lemah dalam susu karena produksi asam yang rendah. Penggunaan S.thermophilus biasanya digabungkan dengan Lb. delbrueckii subsp. bulgaricus. Lactobacillus subspesies ini biasanya disebut dengan rod (bakteri berbentuk batang) dalam industri produk olahan susu, dan kombinasi kedua bakteri ini disebut rod-kokus (Chandan 2006).
Tabel 4. Syarat mutu yogurt menurut SNI (SNI 2981, 2009) No Kriteria uji Satuan Yogurt tanpa perlakuan
panas setelah fermentasi
Yogurt dengan perlakuan panas setelah fermentasi Yogurt Yogurt rendah lemak Yogurt tanpa lemak Yogurt Yogurt rendah lemak Yogurt tanpa lemak 1. Keadaan
1.1. Penampakan - Cairan kental – padat Cairan kental – padat
1.2. Bau - Normal/khas Normal/khas
1.3. Rasa - Asam/khas Asam/khas
1.4. Konsistensi - Homogen Homogen
2. Kadar lemak (b/b) % Min.3 0.6-2.9 Maks. 0.5 Min.3 0.6-2.9 Maks. 0.5 3. Total padatan
susu bukan lemak (b/b)
% Min. 8.2 Min. 8.2
4. Protein (Nx6.38) (b/b)
% Min. 2.7 Min. 2.7 5. Kadar abu (b/b) % Maks. 1.0 Maks. 1.0
6. Keasaman (dihitung sebagai asam laktat) b/b
% 0.5-2.0 0.5-2.0
7. Cemaran logam
7.1. Timbal(Pb) mg/kg Maks. 0.3 Maks. 0.3 7.2. Tembaga (Cu) mg/kg Maks. 20.0 Maks. 20.0 7.3. Timah (Sn) mg/kg Maks. 40.0 Maks. 40.0 7.4. Raksa (Hg) mg/kg Maks. 0.03 Maks. 0.03 7.5. Arsen mg/kg Maks. 0.1 Maks. 0.1 8. Cemaran mikroba
8.1. Bakteri coliform APM/g atau koloni/g
Maks. 10 Maks. 10
8.2. Salmonella - Negatif/25 g Negatif/25 g 8.3. Listeria monocytogenes - Negatif/25 g Negatif/25 g 9. Jumlah bakteri starter* koloni/g Min 107 -
a S r t k a p F f m a f m e c L s p m i k p P d i i Kombina antara kokus S.thermophilus relatif kurang tetapi memilik kedua bakteri agresif, karena proteolitiknya Fermentasi ole format sebaga menstimulasi p asam amino ol fermentasi sus menggantikan eksopolisakarid Yogurt si campuran beb Lactobacillus seperti FOS. probiotik oleh manfaat yang l Konsep d imunomodulat kondisi inflam pencernaan, ya Pada kenyataa dampak keseha Mikrobio infeksi akut, t inflamasi. Norm
asi rod-kokus m dan rod in s lebih aeroto bagus kemam ki aktivitas pep tersebut tumb a lebih aerotol lebih baik, me eh S.thermoph ai hasil sampi pertumbuhan L eh peptida akti su ketika pH S.thermophilu da untuk mend inbiotik merup berapa kultur achidophilus, Kombinasi pr karena substra lebih sempurna dari penggabun tor probiotik m masi. Penggun ang menjadika annya, keseim atan secara imu
ota saluran pe etapi juga dap malisasi mikro menunjukkan r ni berasal dar oleran dibandin mpuan proteolit ptidase lebih ba buh bersama d leran. Pada tah enyediakan pep hilus menekan ingan metabol Lb. delbrueckii if yang disekre mendekati 5 us pada keselur dapatkan tekstu akan salah satu BAL seperti dan Bifidoba robiotik (BAL at yang spesifik a dari kombina ngan probiotik memperkenalka naan probiotik an probiotik s mbangan mikro unofisiologis (R Sum Gambar encernaan yan pat menyebabk oflora indigenu respon pertumb ri karakteristi ngkan Lb. del tiknya dibandi aik dibandingk dalam susu, p hap ini rod tu ptida yang cuk
potensial oks lisme. Tegang subsp. bulgari esikan oleh S.th 5.0. Selebihny ruhan fermenta ur yang lembut u produk susu i Lactobacillu acterium bifidu L) dan prebiot k telah tersedi asi ini (Zhang d k dan prebiotik an potensi strat k bertujuan un ebagai mediat obiota indigen Rautava et al.
mber: Zhang dan
2. Konsep pro g menyimpan kan penyakit k us yang tidak se
buhan yang sin ik fisiologis lbrueckii subsp ing Lb. delbru kan Lb. delbrue ada awalnya S umbuh lebih la
kup untuk men sidasi-reduksi
an oksigen ya icus, yang lebih hermophilus. D a, Lb. bulgar asi. Strain S.th t tanpa pemisah fermentasi yan us bulgaricus, um, yang diko tik dapat men
a untuk fermen dan Ghosh 200 k dapat dilihat
tegi terapi untu ntuk mengura tor anti-inflam nus dalam salu
2004).
Ghosh (2001) obiotik dan preb ng tidak hany
kronis dari ale eimbang denga nergis dalam s dari masing-sp. bulgaricus. ueckii subsp. b eckii subsp. bu S.thermophilus ambat, namun nstimulasi pert sistem, dan m ang rendah da h lanjut dibant Dominasi S.ther ricus secara b hermophilus jug
han whey (Cha ng dibuat denga Streptococcu ombinasikan d ningkatkan day ntasi sehingga 01). pada Gambar uk melawan al angi inflamasi masi (Zhang da uran pencerna biotik a dapat mend ergi autoimun an menggunaka susu. Sinergism masing bakte . S.thermophil bulgaricus, ak ulgaricus. Keti s tumbuh leb karena aktivit tumbuhan koku melepaskan asa an asam form tu oleh pelepas rmophilus dala berangsur-angs ga memproduk andan 2006). an menggunak us thermophilu dengan prebiot ya tahan bakte tubuh mendap r 2. Kemampu lergi, infeksi d i dalam salur an Ghosh 200 aan memberik dasari terjadin hingga penyak an strain spesif me eri. lus kan ika bih tas us. am mat san am sur ksi kan us, tik eri pat uan dan ran 1). kan nya kit fik
mikroflora yang baik untuk saluran pencernaan merupakan alasan yang mendasar dari terapi probiotik (Harish dan Varghese 2006).
Yogurt dalam industri biasanya menggunakan Streptococcus thermophillus dan Lactobacillus delbrueckii subsp. bulgaricus sebagai kultur starternya. Mikroorganisme probiotik pada umumnya memiliki pertumbuhan yang lambat pada medium susu, oleh karena itu, ditambahkan kultur starter yogurt untuk meningkatkan proses fermentasi (Tamime 2005). Bakteri yang berpotensi sebagai probiotik yang dapat digunakan dalam produk olahan susu dapat dilihat pada Tabel 5.
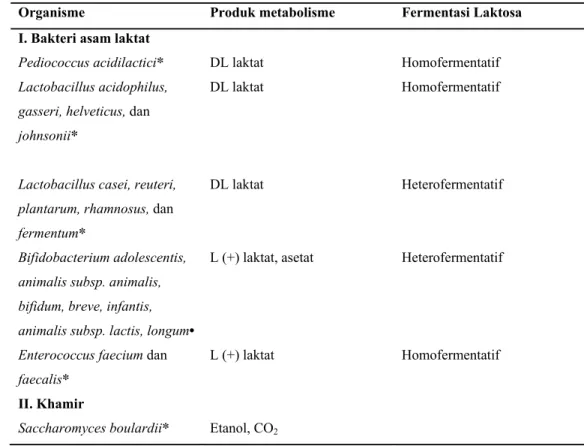
Tabel 5. Probiotik yang dapat digunakan dalam produk olahan susu. Organisme Produk metabolisme Fermentasi Laktosa I. Bakteri asam laktat
Pediococcus acidilactici* DL laktat Homofermentatif
Lactobacillus acidophilus, gasseri, helveticus, dan johnsonii*
DL laktat Homofermentatif
Lactobacillus casei, reuteri, plantarum, rhamnosus, dan fermentum*
DL laktat Heterofermentatif
Bifidobacterium adolescentis, animalis subsp. animalis, bifidum, breve, infantis, animalis subsp. lactis, longum•
L (+) laktat, asetat Heterofermentatif
Enterococcus faecium dan faecalis*
L (+) laktat Homofermentatif II. Khamir
Saccharomyces boulardii* Etanol, CO2
Sumber : •Masco et al. (2004) dan *Tamime et al. (2005)
Untuk mendapatkan efek kesehatan yang diinginkan, bakteri probiotik yang digunakan harus dapat tumbuh dalam media susu dan dapat tumbuh pada kondisi yang terbatas (Tamime 2005). Pada umumnya bakteri probiotik dapat tumbuh baik pada media sintetik seperti tryptose peptone yeast (TPY) dan de Man Rogosa and Sharpe (MRS) broths daripada pada media susu (Shah 2000).
Pada produk pangan, probiotik yang terkandung di dalamnya minimum 106 cfu/g atau asupan harian sebesar 109 cfu/g. Tingginya asupan probiotik untuk mengimbangi kemungkinan hilangnya jumlah mikroorganisme probiotik pada saat melalui perut dan saluran usus. Di jepang, Asosiasi Minuman Fermentasi Susu dan Bakteri Asam Laktat meningkatkan standar yaitu dibutuhkan sedikitnya 107 cfu/ml sel hidup yang ada dalam produk olahan susu.
Viabilitas dan aktivitas bakteri menjadi sangat penting, karena bakteri harus mampu bertahan dalam produk selama masa simpan, selama melalui kondisi asam dalam perut, dan tahan terhadap degradasi oleh enzim hidrolitik dan garam empedu di usus kecil (Tamime 2005). Viabilitas bakteri probiotik dalam yogurt tergantung pada strain yang digunakan, interaksi antar spesies yang ada, produksi hidrogen peroksida yang merupakan hasil metabolisme bakteri, dan keasaman produk akhir.
Viabilitas probiotik juga tergantung pada ketersediaan nutrisi, pendukung dan penghambat pertumbuhan, konsentrasi gula, oksigen terlarut dan perembesan oksigen melalui kemasan (khususnya Bifidobacterium spp.), tingkat inokulasi, dan waktu fermentasi (Oliveira dan Damin 2003 diacu dalam Tamime 2005). Pertumbuhan bakteri probiotik yang lambat pada media susu akan menyebabkan terjadinya pertumbuhan bakteri yang tidak diinginkan dan strain cenderung tidak tumbuh dengan baik dan memproduksi flavor yang tidak diharapkan (Tamime 2005).
2.6. DIARE
Diare atau mencret didefinisikan sebagai buang air besar dengan feses yang tidak berbentuk (unformed stools) atau cair dengan frekuensi lebih dari tiga kali dalam 24 jam (Zein 2004). Diare merupakan perubahan dari kebiasaan normal usus yang dicirikan dengan meningkatnya kandungan air, volume, atau frekuensi buang air besar.
Diare dapat juga didefinisikan ketika terjadi penurunan konsistensi feses (menjadi lembek atau bahkan cair) dan terjadi peningkatan frekuensi buang air besar lebih dari tiga kali dalam sehari (Guerrant et al. 2001). Bila diare berlangsung kurang dari dua minggu, disebut sebagai diare akut. Apabila diare berlangsung dua minggu atau lebih, maka digolongkan pada diare kronik (Zein 2004).
Secara etiologi, diare akut dapat disebabkan oleh infeksi, intoksikasi (poisoning), alergi, reaksi obat-obatan, dan juga faktor psikis (Schiller 2000 diacu dalam Zein 2004). Diare akut infeksi diklasifikasikan secara klinis dan patofisiologis menjadi diare non-inflamasi dan diare inflamasi. Diare inflamasi disebabkan invasi bakteri dan sitotoksin di kolon yang disertai lendir dan darah. Gejala klinis yang sering terjadi adalah adanya keluhan abdomen seperti mulas sampai nyeri seperti kolik, mual, muntah, demam, tenesmus, serta gejala dehidrasi. Pada pemeriksaan tinja rutin secara makroskopis ditemukan lendir dan/atau darah, dan secara mikroskopis ditemukan sel leukosit polimorfonuklear (Zein 2004).
Pada diare non-inflamasi, diare disebabkan oleh enterotoksin yang mengakibatkan diare cair dengan volume yang besar tanpa lendir dan darah. Gejala dehidrasi cepat timbul, terutama pada kasus yang tidak mendapat cairan pengganti. Pada pemeriksaan tinja secara rutin tidak ditemukan leukosit (Zein 2004).
Menurut Bowen (2006), mekanisme terjadinya diare terbagi menjadi empat jenis yaitu diare osmotik, diare sekretorik, diare eksudatif dan infeksi, serta diare yang dikaitkan dengan pengacauan motilitas. Diare osmotik terjadi bila ada bahan yang tidak dapat diserap meningkatkan osmolaritas dalam lumen yang menarik air dari plasma sehingga terjadi diare. Contohnya adalah malabsorbsi karbohidrat akibat defisiensi laktase atau akibat garam magnesium.
Diare sekretorik terjadi bila ada gangguan transport elektrolit, baik absorbsi yang berkurang ataupun sekresi yang meningkat. Hal ini dapat terjadi akibat toksin yang dikeluarkan bakteri, misalnya toksin kolera atau pengaruh garam empedu, ALRP, atau laksatif non-osmotik. Beberapa hormon intestinal seperti gastrin vasoactive intestinal polypeptide (VIP) juga dapat menyebabkan diare sekretorik. Diare eksudatif dan inflamasi akan mengakibatkan kerusakan mukosa, baik usus halus maupun usus besar. Inflamasi dan eksudasi dapat terjadi akibat infeksi bakteri atau bersifat non-infeksi seperti gluten sensitive enteropathy, inflamatory bowel disease (IBD) atau akibat radiasi (Bowen 2006).
Diare sampai saat ini masih menjadi masalah kesehatan, tidak saja di negara berkembang, tetapi juga di negara maju. Penyakit diare masih sering menimbulkan KLB (Kejadian Luar Biasa) dengan penderita yang banyak dalam waktu yang singkat (Manatsathit et al. 2002). WHO memperkirakan 4 milyar kasus terjadi di dunia pada tahun 2000 dan 2.2 juta di antaranya meninggal, sebagian besar anak-anak di bawah umur 5 tahun (Adisasmito 2007).
Data dari profil kesehatan Indonesia 2003, penyakit diare menempati urutan kelima dari 10 penyakit utama pada pasien rawat jalan di rumah sakit dan menempati urutan pertama pada pasien rawat inap di rumah sakit. Berdasarkan data tahun 2003 terlihat bahwa frekuensi kejadian luar biasa (KLB) penyakit diare sebanyak 92 kasus dengan 3865 orang penderita dan 113 orang meninggal (Adisasmito 2007).
Salah satu faktor risiko yang sering diteliti adalah faktor lingkungan yang meliputi sarana air bersih (SAB), sanitasi, jamban, saluran pembuangan air limbah (SPAL), kualitas bakterologis air, dan kondisi rumah. Menurut Sharma et al. (2005), penyebab terjadinya diare pada negara berkembang adalah buruknya sanitasi dan nutrisi. Data terakhir menunjukkan bahwa kualitas air minum yang buruk menyebabkan 300 kasus diare per 1000 penduduk. Sanitasi yang buruk dianggap sebagai penyebab banyaknya kontaminasi bakteri E.coli dalam air bersih yang dikonsumsi masyarakat. Bakteri E.coli mengindikasikan adanya pencemaran tinja manusia. Kontaminasi bakteri E.coli terjadi pada air tanah yang banyak disedot penduduk di perkotaan, dan sungai yang menjadi sumber air baku di PDAM pun tercemar bakteri ini (Adisasmito 2007).
2.7.
ENTEROPHATOGENIC Escherichia coli
(EPEC)
Escherichia coli penyebab diare dapat diklasifikasikan menjadi enam kategori: Enteropatogenik E.coli (EPEC), Enterotoksin E.coli (ETEC), Enterohemoragik E.coli (EHEC), Enteroinvasif E.coli (EIEC), diffusely adderent E.coli (DAEC), dan Enteroagregatif E.coli (EAggEC) (Miyazaki et al. 2010). EPEC merupakan salah satu penyebab utama penyakit diare dan kematian akibat diare pada anak-anak di negara berkembang (Clarke et al. 2002).
EPEC adalah salah satu patogen yang dapat menyebabkan lesi attaching dan effacing (A/E) pada sel usus. Ciri dari patogen A/E adalah terletak pada tumpuannya di permukaan sel epitel inang dan menyebabkan kerusakan pada mikrofili usus. EPEC melekat dan berkolonisasi pada epitel mukosa deodenum dan proksimal jejunum. EPEC menimbulkan kerusakan pada epitel jejunum melalui pembentukan mikrokoloni yang ditunjukkan dengan pelekatan yang terlokalisasi (Moat et al. 2002). Bakteri ini juga melekat dan berkolonisasi pada kolon atau usus besar bagian ascending (naik) dan transverse (melintang)(Jay 2000).
Sumber : Lu dan Walker (2001)
Gambar 3. Infeksi EPEC pada epitel usus inang
EPEC menempel pada sel epitel dan membentuk lesi A/E (Gambar 3). Tahap awal penempelan EPEC pada sel epitel diperantarai oleh bundle-forming pilus (BFP). Setelah pelekatan awal, mikrovili
usus diganggu dan EPEC mensekresikan beberapa faktor virulen melalui sekresi tipe III dan mensekresikan reseptor Tir ke dalam sel inang. EPEC kemudian mengikat Tir melalui protein membran luar, intimin. Sinyal transduksi terjadi dalam sel inang, termasuk aktivasi protein kinase C (PKC), inositol triphosphate (IP3), dan pelepasan Ca2+. Beberapa protein sitoskeletal termasuk aktin, menjadi tempat melekatnya EPEC. Dan pada akhirnya, terjadi penyusunan kembali sitoskeletal setelah Tir-intimin berikatan, dan menghasilkan formasi pedestal-like structure (Lu dan Walker 2001).
EPEC biasanya memiliki locus of enterocyte effacement (LEE), yang membantu perkembangan lesi A/E. LEE terdiri dari gen eae yang mengkode intimin, protein membran terluar yang berikatan dengan protein (dengan tirosin terfosforilasi 90 kDa) di dalam membran inang, sehingga dapat membentuk lesi A/E (Gomes et al. 2004). Tirosin dipindahkan dari sel bakteri ke membran inang sehingga terfosforilasi pada satu atau lebih residu tirosin, berfungsi sebagai reseptor untuk pengikatan intimin. Kemudian sel epitel kehilangan mikrovili dan membentuk cup and pedestal pada tempat melekatnya koloni EPEC. Hal ini menunjukkan bahwa EPEC mampu menginduksi perubahan transport elektrolit ke sel inang. Pada studi yang lain menyatakan bahwa EPEC dapat menyebabkan penurunan transepithelial electrical resistance (TEER) dengan mengganggu tight junction intraseluler (Michail dan Abernathy 2002).
Mekanisme utama dari patogenesis EPEC (Gambar 4) adalah lesi A/E yang dicirikan dengan melekatnya bakteri pada epitelium saluran usus (Nougayrède et al. 2003). Gen eae terletak di area patogenitas LEE dan gen bfpA terletak di plasmid yang disebut EPEC adherence factor (EAF), yang digunakan untuk mengklasifikasikan kelompok bakteri ini menjadi strain tipikal dan atipikal (Kaper 1996 diacu dalam Afset et al. 2004).
Sumber : Nougayrède et al. (2003)
Gambar 4. Fase patogenesis EPEC
Strain E.coli dengan genotipe A/E (eae-) yang mendarat pada plasmid EAF (bfpA-) digolongkan sebagai EPEC tipikal. Kebanyakan dari strain ini termasuk dalam serotipe O:H (Trabulsi et al. 2002). Strain dengan genotipe A/E yang tidak berpengaruh dengan plasmid EAF (bfpA-) diklasifikasikan sebagai EPEC atipikal. Gen eae positif pada strain E.coli ini mendarat pada gen Shiga toksin (stx1 dan/atau stx2) yang sering diklasifikasikan sebagai Enterohaemorrhagic E.coli (Afset et al. 2004).
Menurut Oyetayo (2004), dosis E.coli 105 cfu/ml telah dapat menimbulkan diare pada tikus percobaan. EPEC dapat menyebabkan diare yang durasinya kurang lebih lima hari (Janda dan Abbot 2006).
2.8. USUS HALUS
Usus halus merupakan tempat penyerapan sari-sari makanan. Untuk itu, usus halus memiliki struktur khusus yang dapat meningkatkan pencernaan dan penyerapan sari-sari makanan, seperti adanya mukosa plicae dan vili. Modifikasi dan peningkatan mikrovili di sepanjang permukaan apikal sel luminal didesain untuk memperluas luas permukaan penyerapan (Samuelson 2007).
Panjang usus halus tergantung pada ukuran seluruh tubuh hewan. Pada anjing, usus halus mencapai 3-3.5 kali panjang tubuhnya. Pada kuda mencapai lima kali atau lebih panjang tubuhnya. Lipatan mukosa atau plicae (juga disebut plicae circulares) merupakan perpanjangan semisirkular yang mencapai lumen dan menghasilkan lebih dari dua kali permukaan dari lapisan epitel. Di sepanjang mukosa dalam, perpanjangan yang lebih kecil dari plicae yang mencapai lumen, membentuk vili. Setiap vili memiliki inti lamina propria, yang terdiri dari loops kapiler, saluran limfatik, jaringan yang terhubung lepas dengan sel plasma dan limfosit, corresponding extracelluler matrix, dan serabut otot halus yang memanjang secara vertikal. Vili meningkatkan luas permukaan hingga 10 kali atau lebih. Selain plicae dan vili, kebanyakan sel yang membentuk epitelium mukosa memiliki mikrovili yang seperti pasak, modifikasi sel yang dapat meningkatkan luas permukaan hingga 20-40 kali (Samuelson 2007).
Meskipun sel penyerapan mampu menyediakan material pencernaan (seperti enzim) yang dapat meningkatkan penyerapan, sumber enzim pencernaan lain terdapat di sepanjang usus halus. Sumber tersebut antara lain, kelenjar submukosa yang berada di antara plicae dan vili, dan pankreas. Usus halus terbagi menjadi 3 region yaitu duodenum, jejunum, dan ileum. Duodenum memiliki vili paling banyak dan paling besar dibandingkan jejunum dan ileum. Crypts pada duodenum paling berkembang dan paling banyak (Samuelson 2007).
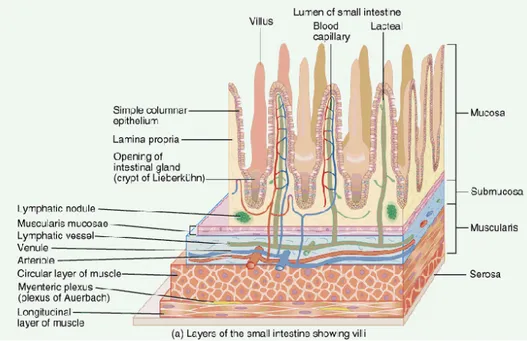
Mukosa usus halus berperan dalam penyerapan nutrisi dan dengan demikian mukosa memiliki permukaan yang lembut dan mudah diserang. Mukosa terdiri dari epitelium sendiri dan didukung oleh jaringan penghubung yang longgar, yang disebut lamina propia, yang berada di bawah epitelium (Gambar 5). Jaringan penghubung yang lebih dalam yang mendukung mukosa disebut submukosa. Dalam saluran pencernaan (GI tract), terdapat lapisan tipis otot halus, mukosa muskularis, yang berada di perbatasan antara mukosa dan submukosa. Epitelium terdiri dari sel penyerapan (enterosit) dengan sel goblet (mensekresi mukus untuk pelumasan) yang tidak beraturan. Lapisan epitel ditingkatkan melalui pembentukan vili. Pelekukan crypts yang terdiri dari sel batang akan mengganti sel epitelium secara terus menerus. Lamina propria menempati inti dari vili, menyelubungi crypts, dan memiliki banyak sel imun. Muskularis mukosa berperan sebagai pendukung lokal dalam menggerakan permukaan mukosa untuk meningkatkan sekresi dan penyerapan nutrisi (King 2009).
Konsentrasi bakteri di dalam usus halus mencapai 1 juta per ml. Pada bagian duodenum jumlah bakteri mencapai 101-103 cfu/ml dan yang tumbuh dengan baik adalah bakteri kokus dan batang gram positif. Pada bagian jejunum, jumlah bakterinya 104-107 cfu /ml dan terdiri dari berbagai macam bakteri seperti Enterococcus faecalis, Lactobacilli, diphtheroids, dan khamir Candida albicans. jumlah bakteri pada ileum sama dengan pada jejunum karena kondisinya yang hampir sama, namun jenis bakteri yang terdapat di ileum sama dengan bakteri yang terdapat pada kolon yang mayoritas adalah bakteri anaerob (Slonczewski dan Foster 2010).
Sumber : DOTE Anatomy Topics,University of Debrecen (2008).
Gambar 5. Histologi usus halus yang menunjukkan vili dan lapisan mukosa.
Fungsi mikroflora dalam usus halus antara lain proteksi, struktur, dan metabolik. Fungsi proteksi antara lain (1) mikroflora dalam usus halus melindungi inang dengan mencegah patogen menempel, (2) bakteri menstimulasi pertumbuhan lapisan usus dan sistem imun pada usus, (3) bakteri berkompetisi dengan patogen untuk mendapatkan nutrisi, sehingga menyulitkan patogen untuk tumbuh, (4) mikroflora menghasilkan antibakteri untuk membunuh kompetitor seperti patogen (Slonczewski dan Foster 2010).
Fungsi strukturnya antara lain (1) flora dalam usus halus menyusun bagian dari barrier usus, (2) flora dalam usus merupakan faktor penting dalam perkembangan sistem imun. Fungsi pada metabolik antara lain (1) mikroflora melindungi inang dari metabolik karsinogenik, (2) mikroflora menyediakan vitamin sintesis seperti biotin dan folat, (3) membantu pembuatan vitamin K, yang diserap dan digunakan oleh inang, dan (4) bakteri sangat penting dalam aktivitas otot usus halus (Slonczewski dan Foster 2010).