TINJAUAN PUSTAKA
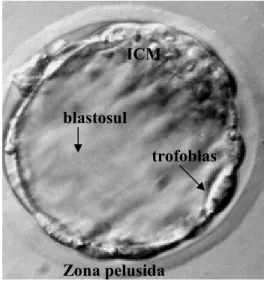
Embrio PraimplantasiPerkembangan embrio praimplantasi dimulai setelah fertilisasi yakni dari mulai tahap zigot yang selanjutnya mengalami pembelahan mitosis sampai membentuk blastosis. Pada mencit blastosis terbentuk empat hari setelah fertilisasi. Struktur blastosis terdiri dari dua tipe sel yaitu inner cell mass (ICM) dan sel trofoblas. Inner cell mass (ICM) akan membentuk embrionic stem cell (ESC) pluripotent yang dapat berdiferensiasi menjadi berbagai tipe sel dari tubuh fetus. Sementara itu sel-sel trofoblas akan berperan dalam proses implantasi dan menjadi selaput ekstraembrionik (Gambar 1) (Kimber & Spanswick 2000; Hardy & Spanos 2000; Goetz 2002). Di bagian dalam dari lapisan sel-sel trofoblas terdapat suatu rongga berisi cairan yang disebut blastosul. Di bagian paling luar, blastosis dibungkus oleh suatu selaput glikoprotein yang disebut dengan zona pelusida. Seiring dengan pertumbuhan embrio praimplantasi, blastosul akan mengalami pembesaran akibat terjadinya akumulasi cairan sehingga ukuran blastosis bertambah besar dan tahap tersebut disebut dengan blastosis yang meluas (blastosis ekspan). Selanjutnya blastosis eskpan akan mengalami nidasi (hatching).
Gambar 1 Blastosis ekspan mencit. Zona pelusida
blastosul ICM
Nidasi Blastosis
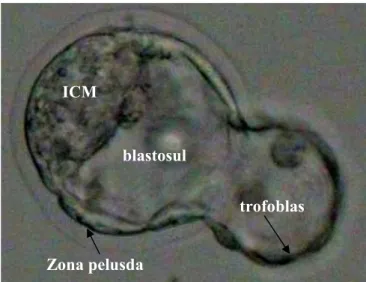
Nidasi merupakan proses keluarnya blastosis dari zona pelucida. Nidasi dimulai dengan adanya akumulasi cairan secara bertahap dalam blastosul sehingga terjadi peningkatan ukuran atau volume blastosis dari ukuran awalnya. Akumulasi cairan dalam blastosul mengandung Na+/K+ dan ATPase yang menyebabkan terjadinya peningkatan tekanan hidrostatik internal pada trofoblas dan zona pelusida. Akibatnya zona pelusida menjadi tipis dan elastis (Gambar 2) (Balaban et al.2002; Gonzales et al. 1996; Jones et al. 2000).
Gambar 2 Proses nidasi blastosis dari zona pelusida
Blastosis keluar dari zona pelusida pada kutub yang berlawanan dengan inner cell mass (ICM) dan proses nidasi akan selesai apabila seluruh blastosis telah keluar dari zona pelusida. Selanjutnya fimbria halus dan kecil muncul pada sel trofoblas dan bergerak secara amuboid ke dinding endometrium untuk melakukan proses implantasi yang diawali dengan terjadinya perlekatan dengan epitel endometrium (Horse et al. 2004; Osagie & Biggers 2003).
Pada blastosis faktor-faktor yang dapat mempengaruhi proses nidasi diantaranya, adalah: (1) ketersediaan energi (ATP) dalam blastosis yang diperlukan untuk menekan zona pelusida, (2) ketebalan zona pelusida, yakni semakin tebal zona pelusida maka energi dan tekanan yang dibutuhkan akan semakin besar untuk dapat menembus zona pelusida, serta (3) aktivitas enzim proteolitik yang berperan penting dalam mencerna zona pelusida sehingga zona
ICM
blastosul
Zona pelusda
pelusida menjadi tipis dan blastosis dapat dengan mudah keluar dari zona pelusida (Osagie et al. 2003; Fong et al.2001).
Implantasi Embrio
Proses implantasi dimulai setelah blastosis keluar dari zona pelusida yang diikuti dengan kontak secara fisik dan fisiologis antara trofoblas dengan dinding endometrium uterus (Horse et al. 2004). Proses ini melibatkan serangkaian interaksi intraseluler serta interaksi antara sel dan matriks (Dey et al. 2004).
Ada tiga tahapan dalam proses implantasi, yaitu (1) aposisi, (2) perlekatan (adhesi) dan (3) invasi. Aposisi merupakan adhesi yang tidak stabil dan terjadi setelah blastosis berkontak dengan dinding uterus; sedangkan perlekatan (adhesi) merupakan perlekatan yang stabil antara trofoblas dengan sel epitel uterus. Selanjutnya sel trofoblas melakukan infiltrasi ke dalam endometrium yang diikuti dengan invasi di dalam endometrium membentuk anchoring vili agar fetus memperoleh makanan dari induk (Gambar 3) (Dey et al. 2004; Dominguez et al. 2005).
Pada manusia dan mencit endometrium mengalami desidualisasi, yaitu sel- sel stroma mengalami perubahan morfologi, perubahan komposisi matriks interseluler serta peningkatan pertumbuhan kapiler-kapiler pembuluh darah. Adanya desidualisasi menyebabkan dikeluarkannya bahan-bahan metabolit, seperti karbohidrat, protein, lemak dan asam nukleat. Bahan-bahan tersebut penting untuk nutrisi embrio selama implantasi (Dey et al. 2004; Kliman 2000).
INVASION
Gambar 3 Proses implantasi embrio ( Dominguez et al. 2005).
Berdasarkan perbedaan cara interaksi antara blastosis dan sel-sel uterus, implantasi dapat dikelompokkan menjadi tiga jenis, yaitu: (1) sentral (superficial), (2) eksentrik dan (3) interstisial (profundal). Pada implantasi superficial, implantasi hanya berupa perlekatan pada permukaan uterus dan tidak terjadi infiltrasi pada epitel endometrium. Tipe ini terdapat pada mamalia seperti kuda, sapi, domba dan kambing. Implantasi eksentrik terjadi pada hewan anjing, kucing, tikus dan mencit; pada tipe ini implantasi terjadi pada epitel uterus yang mengalami invaginasi dan terjadi sedikit kerusakan pada stroma uterus. Pada tipe interstitial terjadi pada babi, simpanse dan manusia. Pada tipe ini, implantasi blastosis terjadi dalam subepitel stroma.
Diferensiasi Sel Trofoblas
Setelah blastosis keluar dari zona pelusida, sel-sel trofoblas akan melekat pada dinding endometrium uterus. Sel trofoblas merupakan sel-sel yang berperan penting dalam proses implantasi dan membentuk hubungan antara induk dan fetus (Horse et al.2004; Nadra et al. 2006). Pada masa proses implantasi sel-sel trofoblas mengalami proliferasi dan diferensiasi. Secara morfologi, diferensiasi sel-sel trofoblas dibagi kedalam empat tipe, yaitu: 1) giant trophoblast cell, 2) sel spongiotrofoblas (spongiotrophoblast), 3) sel glikogen, dan 4) sel sinsitiotrofoblas (syncytiotrophoblast) (Kliman 2000; Kimber & Spanswick 2000). Keempat tipe sel tersebut merupakan hasil penggabungan (fusi) atau diferensiasi sitotrofoblas (cytotrophoblast). Sebagian sitotrofoblas bergabung bersama membentuk sinsitiotrofoblas. Selain itu sitotrofoblas juga berdiferensiasi menjadi giant trophoblast cell, spongiotrofoblas dan sel glikogen trofoblas (Lunghi et al. 2007; Kliman 2000).
Giant trophoblast cells terdapat pada koriovitelin dan korioalantois plasenta. Sel ini berukuran besar dengan satu nukleus. Giant trophoblast cells menghasilkan beberapa growth factor (baik secara autokrin maupun parakrin) dan hormon-hormon yang mendukung pertumbuhan dan perkembangan fetus serta interaksi fisiologis antara fetus dan induk (Dey et al. 2004). Sel ini melakukan proses endoreduplikasi, seperti melanjutkan sintesis DNA tanpa pembelahan sel. Diferensiasi giant trophoblast cells didukung oleh dua gen faktor transkripsi basic helix-loop helix (bHLH) yaitu Hand 1 dan Stra13.
Sel spongiotrofoblas secara morfologi berbeda dengan giant trophoblast cells dan turut berperan dalam aktivitas endokrin. Sel glikogen sesuai dengan namanya, sel ini banyak mengandung glikogen yang berpotensi dalam penyimpanan energi dan muncul pada akhir kebuntingan (Kliman 2000).
Sel sinsitiotrofoblas merupakan sel dengan dua atau lebih nukleus (multinukleus) berasal dari hasil penggabungan sitotrofoblas. Sel ini bertanggung jawab dalam pengangkutan nutrien dan pembuangan sisa metabolisme, mensekresikan hormon human chorionic gonadotrophin (hCG) yang penting untuk mempertahankan korpus luteum untuk menghasilkan progesteron yang diperlukan pada awal kebuntingan. Sinsitiotrofoblas menghasilkan laktogen
plasenta yang berperan sebagai regulator metabolisme lipid dan karbohidrat pada induk. Selain itu sinsitiotrofoblas juga menghasilkan ß1-glycoprotein spesifik, plasminogen activator inhibitor type 2, growth hormone, collagenase, thrombomodulin dan reseptor-reseptor growth factor ( Haig 1996; Frendo et al 2003).
Secara in vitro, diferensiasi sel sitotrofoblas diketahui dipengaruhi oleh beberapa faktor seperti growth factor (epidermal growth factor, kelompok granulosit-makrofag) dan hormon-hormon (hCG, estradiol) (Lunghi et al. 2007; Kliman 2000). Menurut Frendo et al (2003) terdapat keterlibatan cyclic AMP (cAMP), protein kinase serta stres oksidatif dengan diferensiasi dan fusi sel trofoblas. Selain itu, kandungan oksigen berhubungan dengan respirasi dan pembentukan ATP serta cAMP. Rendahnya kandungan oksigen atau sampai dalam kondisi hipoxia dapat menghambat proliferasi sitotrofoblas dan diferensiasi sinsitiotrofoblas (Aplin & Kimber 2004; Cartwright et al.2002; Lunghi et al. 2007).
Implantasi dan Perkembangan Sel Trofoblas dalam Sistem Kultur In Vitro Upaya untuk mempelajari proses implantasi secara in vivo mengalami kendala. Untuk dapat mengamati proses implantasi khususnya pertumbuhan dan perkembangan sel-sel trofoblas diperlukan sistem in vitro yang memadai. Berbagai model implantasi blastosis telah dilakukan, diantaranya menggunakan feeder layer seperti sel epitel endometrium, sel stroma, sel desidua serta matriks ekstraseluler seperti laminin dan fibronectin yang berperan sebagai tempat perlekatan dan invasi menyerupai sel-sel uterus maternal (Tayade et al. 2005; Harun 2006).
Dalam sistem kultur in vitro, telah dilaporkan bahwa sel-sel trofoblas mampu tumbuh sampai mengalami perlekatan (attachment) dan pertumbuhan (outgrowth) yang mirip dengan proses adesi dan invasi secara in vivo. Sel-sel trofoblas yang dikultur secara in vitro dapat tumbuh dengan baik apabila dikondisikan sesuai dengan suasana in vivo. Penggunaan sistem kultur yang baik mampu mendukung pertumbuhan dan perkembangan sel-sel trofoblas, terutama penggunaan medium kultur yang tepat, penambahan serum, substrat energi (piruvat dan glukosa), metabolit esensial (karbohidrat, asam amino, vitamin, protein dan peptida), ion-ion anorganik, matriks ekstraseluler seperti laminin, serta sistem inkubasi yang meliputi suhu, pH, fase gas serta osmolalitas medium (Gardner et al.2002; Summers 2003).
Pada lingkungan in vitro, medium merupakan hal yang penting dalam perkembangan sel-sel trofoblas. Medium yang digunakan harus mengandung glukosa konsentrasi tinggi sebagai sumber energi. Sedangkan piruvat dan laktat dalam konsentrasi rendah. Hal ini disebabkan pada perkembangan embrio pasca implantasi terjadi peningkatan aktivitas metabolisme sehingga memerlukan energi yang banyak. Selain itu, penambahan asam amino dan growth factor penting dalam meningkatkan viabilitas dan perkembangan embrio secara in vitro (Gardner et al. 2002).
Sel-sel trofoblas di dalam medium kultur akan melakukan perlekatan pada dasar cawan petri atau pada feeder layer setelah blastosis mengalami nidasi. Selanjutnya sel-sel trofoblas akan berproliferasi dan tumbuh ke arah luar dari inner cell mass (ICM) (outgrowth). Semakin lama kultur maka pertumbuhan sel
trofoblas akan semakin luas sampai mencapai batas maksimum (Gambar 4) (Tayade et al. 2005).
Hari Tingkat Gambaran embrio Potongan melintang
0 Blastosis 1 Nidasi 2 Perlekatan 3-5 Awal pertumbuhan 6-7 maksimum pertumbuhan 8-9 akhir pertumbuhan
Mitokondria
Mitokondria berasal dari bahasa Yunani yaitu mito yang berarti benang dan chondrion yang berarti granul atau butiran-butiran, sehingga dapat diartikan organel yang mempunyai rangkaian granul-granul atau butiran-butiran yang tersusun seperti benang. Mitokondria merupakan organel yang unik karena memiliki materi genetik (DNA) sendiri berbentuk lingkaran yang turut berperan dalam pewarisan sifat keturunan. Materi genetik ini berasal dari maternal, sehingga setiap oosit mengandung banyak mitokondria pada sitoplasmanya (Tamassia et al. 2004).
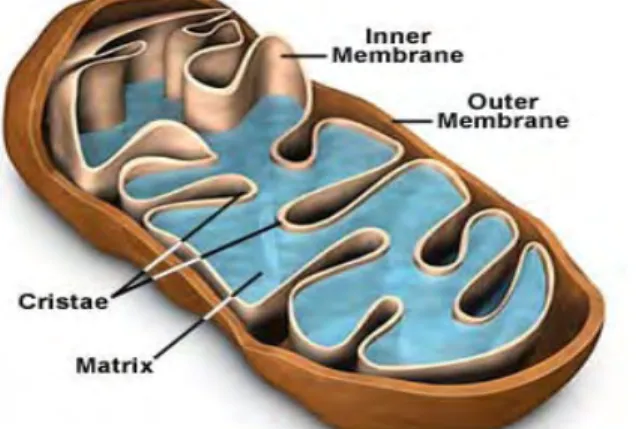
Struktur mitokondria berbentuk oval dengan panjang 1-2 mm dan lebar 0,5-1 mm, berada dalam sitoplasma yang strukturnya dikelilingi oleh dua membran, yaitu membran luar dan membran dalam. Keduanya merupakan fosfolipid lapis ganda (bilayer) yang terdiri atas kumpulan protein yang unik. Pada membran luar terlihat halus tetapi mengandung protein transpor yang disebut porin, sehingga membran luar berfungsi sebagai tempat keluar masuknya ion atau molekul kecil, termasuk protein berukuran kecil kedalam ruang antar membran, namun tidak semuanya melewati membran dalam yang bersifat impermeabel seperti di ruang matriks. Pada membran dalam terdapat bagian yang berlipat-lipat, disebut krista. Membran dalam membagi mitokondria menjadi dua ruang. Ruang pertama berupa ruang antar membran, daerah sempit antara membran dalam dan membran luar. Ruang kedua, yaitu matriks mitokondria yang dilingkupi oleh membran dalam (Gambar 4) (Brokes et al. 2004; Cummins 2001; Dimauro & Schon 2003).
Membran dalam dan matriks merupakan tempat utama aktivitas mitokondria. seperti siklus asam sitrat, oksidasi asam lemak, sintesis urea serta pembentukkan energi. Hal ini disebabkan di dalam matriks banyak terkandung enzim yang mencapai sekitar 67%. Sedangkan krista membuat membran dalam mitokondria mempunyai pemukaan yang luas sehingga produktivitas respirasi seluler meningkat (Dimauro & Schon 2003; Davidson 2006).
Produksi ATP dalam Mitokondria
Mitokondria pada awal perkembangan mempunyai struktur yang lebih kecil dibanding individu dewasa dengan diameter < 0,5 µm. Selain itu mitokondria pada stadium ini terdiri dari krista pendek dengan jumlah sedikit, sehingga aktivitas respirasi rendah. Respirasi ini akan meningkat seiring dengan perkembangan embrio. Struktur mitokondria juga akan mengalami perubahan sesuai aktivitasnya, karena harus menghasilkan ATP (Adenosin Triphosphat) dalam jumlah banyak. Dengan demikian konsumsi oksigen pada embrio semakin lama akan meningkat sesuai perkembangannya (Trimarchi et al. 2000; Cummins 2001; Harvey et al. 2002; Osagie et al. 2003).
Pada awal perkembangannya embrio sangat membutuhkan energi untuk aktivitasnya dan hampir 85% energi embrio dihasilkan dalam mitokondria melalui metabolisme fosporilasi oksidasi dan siklus Kreb dalam bentuk ATP (Blerkom 2004; Harvey et al. 2002; Gardner et al. 2002). Energi atau ATP sangat dibutuhkan untuk aktivitas biosintesis seluler, produksi membran plasma dan proses perkembangan morfodinamika yang kritikal seperti pembentukan blastosul pada proses blastulasi dan ekspansi blastosis. Selain itu ATP juga diperlukan pada waktu nidasi agar embrio dapat implantasi (Gardner et al. 2002; Ludwig et al. 2001; Fleming 2004).
Aktivitas mitokondria dalam membentuk energi membutuhkan oksigen dan nutrien atau substrat seperti glukosa, piruvat dan laktat yang akan diubah menjadi ATP, proses ini disebut respirasi aerobik. Oleh sebab itu, mitokondria juga dikenal sebagai power house cell. Produksi energi dalam mitokondria melibatkan dua proses metabolisme, yaitu (1) siklus asam sitrat dan (2) rantai
transpor elektron. Siklus asam sitrat merubah karbohidrat dan asam lemak menjadi ATP dan hidrogen dalam bentuk nicotinamide adenine dinucleotida dehydrogenase (NADH) dan flavin adenin dinucleotida dehidrogenase (FADH2) yang merupakan molekul tinggi energi karena masing-masing molekul tersebut mengandung sepasang elektron yang mempunyai potensial transfer tinggi.
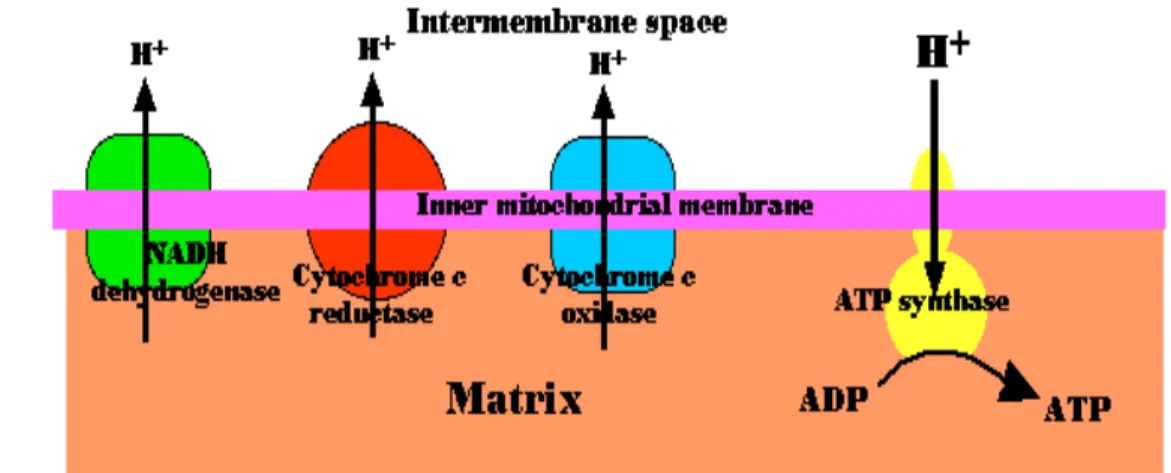
Rantai tranpor elektron merupakan kumpulan molekul yang tertanam pada membran dalam mitokondria. Pelipatan membran dalam untuk membentuk krista meningkatkan luas permukaannya untuk dapat melakukan transpor elektron. Elektron yang masuk ke rantai transpor elektron berasal dari hasil reaksi glikolisis dan siklus krebs yang disimpan dalam bentuk NADH. Bila masuk kedalam sistem rantai transpor elektron dan terjadi penggabungan hidrogen dan oksigen maka dapat membentuk ATP, sehingga disebut juga fosforilasi oksidatif. Proses ini menghasilkan ATP lebih banyak dibanding siklus asam sitrat dan glikolisis, yaitu 30 ATP yang terbentuk, 26 ATP berasal dari proses fosforilasi oksidatif. Sebagian besar komponen rantai transpor elektron merupakan protein.
Reaksi fosforilasi oksidatif dalam mitokondria terdiri atas lima kompleks protein, yaitu (1) kompleks I mengkatalis NADH dari siklus asam sitrat dengan NADH-coenzym Q reductase sebagai reseptor menjadi bentuk tereduksi. NADH merupakan kunci utama dari reaksi fosforilasi oksidatif yang berperan sebagai perantara antara siklus krebs dengan rantai transpor elektron pada komplek I. Langkah awal adalah pengikatan NADH dan transfer dua elektronnya ke flavin mononukleotida (FMN), gugus prostetik komplek ini, menjadi bentuk tereduksi, FMNH2. Elektron kemudian ditransfer dari FMNH2 keserangkaian rumpun belerang besi (4Fe-4S), jenis kedua gugus prostetik dalam NADH-Q reduktase. Elektron dalam rumpun belerang-besi kemudian diangkut ke ko-enzym Q, dikenal juga sebagai ubiquinon. Ubiquinon mengalami reduksi menjadi radikal bebas anion semiquinon dan reduksi kedua terjadi dengan pengambilan elektron kedua membentuk ubiquinol (QH2) yang terikat enzim. Pasangan elektron pada QH2 dipindahkan ke rumpun belerang besi (2Fe-2S) kedua yang ada pada NADH-Q reduktase, dan akhirnya ke Q yang bersifat aktif dalam inti hidrofobik membran dalam mitokondria. Aliran dua elektron ini menyebabkan terpompanya empat H+ dari matriks kesisi sitosol membran dalam mitokondria. Gangguan pada rantai
transpor elektron terutama pada enzim kompleks I dapat mengurangi produksi ATP, meningkatkan kebocoran elektron dan meningkatkan produksi superoksida (Grivennikova 2001). (2) Kompleks II menerima elektron dari FADH2 dan juga melewati kompleks III melalui coenzyme Q, (3) kompleks respirasi III dengan cytochrome c tereduksi dan cytochrome c oxidoreductase sebagai akseptor dan (4) kompleks respirasi IV (cytochrome c oxidase) mengkatalis oksidasi cytochrome c. (5) Kompleks V merubah adenosin diphosphate (ADP) menjadi ATP dengan bantuan enzim ATP synthase. Adapun enzim yang terlibat dalam reaksi ini yaitu NADH-coQ reductase, coQH2-cytochrome c reductase, cytochrome c reductase dan ATP synthase (Dimauro dan Schon 2003;Klobuear dan Gorup 2004).
Gambar 6 Rantai respirasi elektron dalam mitokondria.
Secara garis besar reaksi pembentukan ATP yang berlangsung di mitokondria dapat dibagi menjadi tiga tahap: (1) Reaksi oksidasi piruvat (atau asam lemak) menjadi CO2. Reaksi ini terkait dengan reduksi NAD+ dan FAD menjadi NADH dan FADH2. Reaksi-reaksi ini berlangsung dalam ruang matriks mitokondria, (2) transfer elektron dari NADH dan FADH2 ke O2. Rangkaian reaksi ini berlangsung pada membran dalam mitokondria dan (3) pemanfaatan energi yang tersimpan dalam bentuk gradien elektrokimia untuk memproduksi ATP. Reaksi ini dikatalisis oleh enzim ATP sintetase yang berlokasi pada membran dalam mitokondria (Campbell et al.2002).
Gangguan pada sistem rantai transpor elektron dapat meningkatnya produksi radikal bebas. Radikal bebas adalah molekul reaktif yang dapat merusak DNA dan membran sel melalui jalur oksidasi. Normalnya, rantai respirasi mitokondria membuat radikal bebas dalam jumlah yang rendah selama proses pembuatan ATP. Dengan meningkatnya radikal bebas dapat menyebabkan kerusakan lebih lanjut DNA mitokondria (mtDNA) yang akan mengakibatkan vicious cycle timbulnya kerusakan dan produksi radikal bebas, salah satunya apoptosis dan mempercepat penuaan (aging) (Turrens 2003; Tamassia et al.2002; Cummin 2001 ).
Mitokondria dan Apoptosis
Mitokondria selain berfungsi sebagai tempat penghasil energi (ATP) juga berperan penting sebagai pengontrol reaksi reduksi oksidasi, homeostasis dan apoptosis serta aging. Gangguan fungsi mitokondria dalam memproduksi energi dapat mengakibatkan gangguan pada fungsi lain dari mitokondria, seperti terjadinya peningkatan apoptosis dari aktivitas normalnya (Vogel 2005; Watson et al. 1998).
Pada proses implantasi, dinding endometrium mengalami apoptosis pada waktu sel trofoblas melakukan invasi dan infiltrasi ke dalam endometrium. Selain itu, sel-sel trofoblas juga mengalami apoptosis pada waktu invasi. Namun jumlah apoptosis yang meningkat dari normalnya dapat mengakibatkan terjadi kelainan implantasi atau kebuntingan, seperti preeclamsia (Zhang & Paria 2006; Jauniaux et al. 2003; Joswig et al. 2003; Thouas et al. 2005).
Apoptosis merupakan suatu proses kematian sel terencana (programmed cell death) yang dicirikan dengan terjadinya pengerutan sel dan nukleus. Kromatin nukleus pada proses apoptosis mengalami kondensasi, yang kemudian pecah. Pecahan ini kemudian dibungkus dalam suatu vesikel, disebut sebagai apoptotic bodies, yang mengandung pula sisa-sisa organel sel dan fragmen DNA lain, untuk selanjutnya difagositosis baik oleh sel-sel tetangganya maupun mengalami degradasi otomatis (Wang 2001; Vogel 2005).
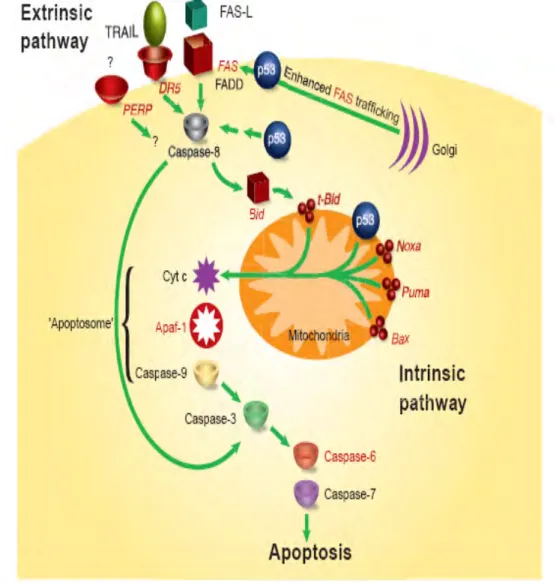
Apoptosis terjadi melalui 2 jalur utama yakni (1) jalur ekstrinsik atau death receptor (DR) pathway, dan (2) jalur intrinsik atau jalur mitokondria. Pada jalur ekstrinsik, apoptosis dimulai setelah DR pada membran plasma berikatan dengan protein Fas, suatu glycocylated cell-surface protein dengan berat molekul 42-52 kDa. Reaksi ini akan diikuti oleh apoptotic pathway yang terdiri dari seperangkat enzim (FAAD, TRADD, caspase 8 dan 10), berfungsi sebagai penggerak efektor apoptosis (Haupt et al. 2003; Wang 2001).
Pada jalur intrinsik yang terjadi di intraseluler akan muncul inisiasi apoptosis oleh produksi biokimia yang berasal dari intraseluler stres, seperti oksidatif stres, perubahan redoks, ikatan kovalen, peroksidasi lipid. Jalur apoptosis distimulus dari luar yang mengaktifkan caspase untuk menginduksi lepasnya sitokrom c. Lepasnya sitokrom c akan berikatan dengan Apaf-1, yaitu protein yang terdapat dalam sitosol yang normalnya merupakan monomer inaktif. Selanjutnya akan membentuk apoptosom yang melepaskan procaspase 9 dan menginduksi dilepasnya caspase 3, sehingga sel mengalami apoptosis (Gambar 5) (Wang 2001; Vogel 2005; Zess 2005; Haupt et al.2003).
Gambar 7. Dua jalur apoptosis (jalur ekstrinsik dan intrinsik) (Haupt et al. 2003).
Proses apoptosis tidak menyebabkan terjadinya reaksi inflamasi, namun prosesnya diinduksi oleh reaksi inflamasi sebelumnya. Apoptosis membutuhkan energi, tetapi proses ini dapat menghabiskan energi akibat banyaknya pori dari membran mitokondria yang terbuka, akibatnya sel yang mengalami apoptosis mempunyai energi (ATP) yang rendah atau menurun (Straszewski-Chavez et al. 2005; Zess 2005).
Tinggi atau rendahnya aktivitas mitokondria berhubungan erat dengan banyak atau sedikitnya jumlah dan distribusi mitokondria dalam sitoplasma. Semakin banyak jumlah mitokondria dan pola distribusinya merata didalam sitoplasma menunjukkan aktivitas mitokondria bekerja dengan baik. Sebaliknya
jumlah mitokondria yang sedikit dan pola distribusi yang tidak merata atau mengelompok menunjukkan aktivitas mitokondria yang terganggu atau terhambat akibatnya energi atau ATP yang dihasilkan dalam jumlah sedikit. Akibatnya sel dapat berhenti bekerja dan mengalami apoptosis (Cummins 2001; Tarin et al. 2000).