TINJAUAN PUSTAKA Klasifikasi dan Biologi Rayap
Rayap di daerah subtropik disebut dengan ’semut putih’ (white ants) karena memiliki morfologi yang mirip dengan semut. Berdasarkan hubungan evolusi (filogeni), tidak ada hubungan antara rayap dengan semut. Hubungan lebih dekat terjadi antara rayap dengan kecoa (Blattodea) (Lo et al. 2000; Inward et al. 2007).
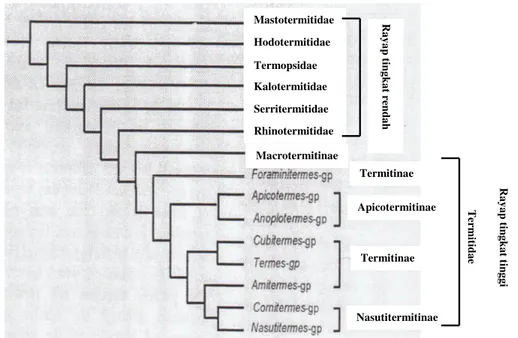
Rayap merupakan salah satu kelompok serangga dengan jumlah keragaman yang besar. Rayap (Ordo Isoptera) terdiri atas tujuh famili, yaitu Mastotermitidae, Kalotermitidae, Termopsidae, Hodotermitidae, Rhinotermitidae, Serritermitidae, dan Termitidae. Sampai sekarang sudah tercatat 14 subfamili, 281 genus dan lebih dari 2600 spesies termasuk dalam kelompok ini (Kambhampati & Eggleton 2000). Famili Termitidae merupakan famili terbesar dalam Ordo Isoptera dan mencakup tiga perempat spesies yang diketahui dan merupakan kelompok rayap yang paling maju. Spesies rayap yang termasuk famili Termitidae dikelompokkan ke dalam rayap tingkat tinggi, sedangkan enam famili yang lain dikelompokkan ke dalam rayap tingkat rendah (Gambar 1). Pembagian kelompok ini didasarkan atas morfologi dan DNA mitokondria (16S rRNA dan NADH 5 dehydrogenease). Karakter yang digunakan sebanyak 197 karakter morfologi dan biologi dari pekerja dan prajurit dengan menekankan pada struktur usus pekerja (Kambhampati & Eggleton 2000). Gen pada mitokondria seperti 16S rRNA, NADH 5 dehydrogenease, dan cytochrome oxidase II (COII) dapat digunakan sebagai kronometer evolusi karena bukan merupakan hasil rekombinasi, berubah sesuai dengan waktu tetapi tidak terlalu cepat, ada pada semua organisme, dan memiliki fungsi yang sama (Sogin et al. 1986; Hoy 1994; Gupta 1998; Philippe et al. 2000; Freeland 2005)
Rayap merupakan serangga sosial yang memiliki pembagian tugas yang jelas yang dinyatakan dalam pembagian kasta. Berdasarkan kemampuan bereproduksi rayap dibagi menjadi dua kasta yaitu kasta reproduktif dan kasta steril (Krishna 1969; Lee & Wood 1971).
Kasta reproduktif terdiri atas reproduktif primer dan reproduktif sekunder. Kasta reproduktif primer (pendiri koloni) biasa disebut laron terdiri atas jantan (raja) and betina (ratu). Ciri khas kasta reproduktif primer adalah adanya sepasang
sayap pada bagian toraks. Sedangkan kasta reproduktif sekunder berfungsi menggantikan kasta reproduktif apabila raja atau ratu mati atau untuk menambah jumlah telur apabila telur yang dihasilkan oleh ratu tidak mencukupi kebutuhan koloni (Krishna 1969).
Kasta steril terdiri atas pekerja dan prajurit. Ciri dari kasta ini adalah tidak adanya sayap dan perkembangan organ seksual ditekan atau tidak berkembang. Pekerja bertanggungjawab untuk mencari makan dan memelihara telur, larva dan ratu. Larva, prajurit dan ratu tidak mampu untuk memberi makan dirinya sendiri sehingga bergantung pada makanan yang diberikan pekerja. Jumlah pekerja mencapai 90% dari seluruh anggota koloni. Rayap prajurit bertugas menjaga koloni dari serangan musuh dan juga menjaga pekerja yang mencari makan di sekitar sarang. Prajurit dibedakan dengan pekerja berdasarkan modifikasi bagian mulut dan kepala yang mengalami kitinasi yang kuat, biasanya terpigmentasi dan seringkali lebih besar daripada ukuran kepala kasta yang lain (Krishna 1969).
Secara umum makanan rayap adalah semua bahan yang mengandung selulosa. Bignell dan Eggleton (2000), membagi rayap menjadi beberapa kelompok berdasarkan jenis makanannya. Pertama, rayap pemakan tanah
(soil-Gambar 1 Filogeni rayap berdasarkan morfologi dan DNA mitokondria (16S rRNA dan NADH 5 dehydrogenease) (Kambhampati & Eggleton 2000). Termitinae Apicotermitinae Termitinae Nasutitermitinae T er m it id a e Macrotermitinae Rhinotermitidae Serritermitidae Kalotermitidae Termopsidae Hodotermitidae Mastotermitidae R a y a p t in g k a t r en d a h R a y a p t in g k a t t in g g i
feeder) yang mendapatkan makanan dari mineral tanah. Material yang dicerna sangat heterogen, mengandung banyak bahan organik tanah dan silika. Rayap jenis ini ditemukan pada Apicotermitinae, Termitinae, Nasutitermitinae, dan Indotermitinae. Kedua, rayap pemakan kayu (wood-feeder) yang mendapatkan makanan dengan memakan kayu dan sampah berkayu, termasuk cabang mati yang masih menempel di pohon. Hampir semua rayap tingkat rendah adalah pemakan kayu, semua subfamili dari Termitinae kecuali Apicotermitinae. Ketiga, rayap pemakan serasah (litter-feeder). Rayap jenis ini mendapatkan makanan dari daun atau kayu-kayu kecil. Rayap ini terdapat pada Macrotermitinae, Apicotermitinae, Termitinae, dan Nasutitermitinae.
Beberapa jenis rayap yang ada di Indonesia antara lain Neotermes tectonae, Cryptotermes cyanocephalus, C. curvignathus, Schedorhinotermes javanicus, Procapritermes setiger, Pericapritermes semarangi, Macrotermes gilvus, Microtermes insperatus, Microtermes jacobseni, Nasutitermes javanicus, dan Nasutitermes matangensis (Lee & Wood 1971; Harris 1971; Teguh 2009).
Taksonomi dan Biologi C. curvignathus
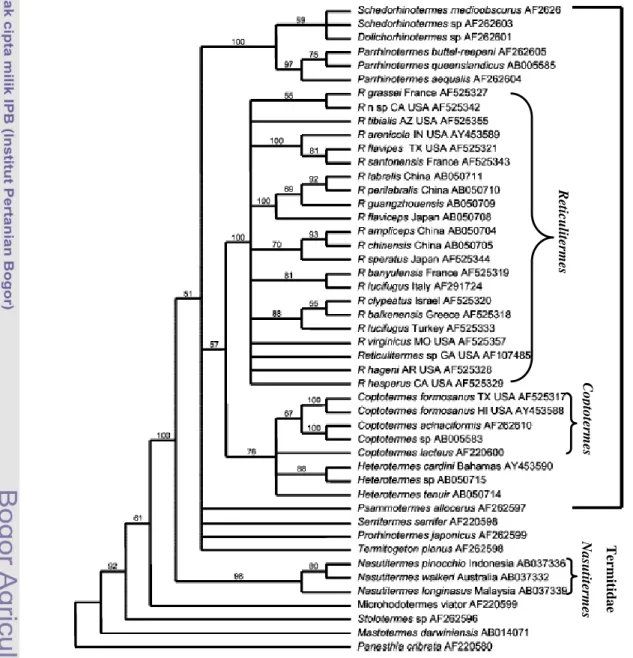
Coptotermes curvignathus merupakan salah satu rayap subteran. Secara taksonomi C. curvignathus termasuk dalam Klas Insekta, Ordo Isoptera, Famili Rhinotermitidae, Subfamili Coptotermitinae, Genus Coptotermes (Ahmad 1958; Krishna 1969). Berdasarkan analisis gen cytochrome oxidase II (COII) genus Coptotermes terletak diantara genus Reticulitermes dan Nasutitermes (Gambar 2) (Austin et al. 2004).
Rayap C. curvignathus berwarna pucat (Gambar 3a). Prajurit memiliki kepala berbentuk oval, melebar, dan datar dengan fontanel menonjol yang dapat mengeluarkan cairan berwarna putih seperti susu apabila diserang. Eksudat ini berguna untuk melumpuhkan musuhnya. Mandibula berwarna merah kecoklatan dan melengkung di bagian ujungnya. Ukuran prajurit relatif besar (±4.5 mm); lebar kepala 1.34-1.52 mm; antena 14-15 ruas, ruas kedua dua kali panjangnya atau sedikit lebih panjang daripada ruas ketiga (Gambar 3b) (Ahmad 1958).
Perbedaan rayap prajurit C. curvignathus dengan beberapa rayap prajurit spesies Coptotermes yang lain adalah pada Coptotermes gestroi lebar pronotum 0.65 mm, kepala kuning dengan rambut yang jarang, antenna 15 ruas, ruas ketiga
Gambar 2 Filogeni genus Reticulitermes, Coptotermes, dan Nasutitermes berdasarkan gen cytochrome oxidase II (COII) (Austin et al. 2004).
N a su tit er m es C o p to te rm es R et ic u lit er m es R h in o te rm it id a e T er m it id a e
sedikit lebih pendek dari ruas keempat. Sedangkan pada Coptotermes formosanus kepala menyempit sebanding dengan panjang, perbedaan antara panjang dan lebar kepala 0.26-0.38, tebal kepala 0.86, antenna 13-15 ruas, ruas kedua sedikit lebih panjang dari ruas ketiga. Ciri Coptotermes havilandi adalah pronotum menyempit dan tepi anterior sedikit cembung, labrum lebih panjang dari lebarnya dan lancip, kepala oval memanjang, dan antena 15-18 ruas, ruas kedua satu setengah kali lebih panjang daripada ruas ketiga (Ahmad 1958).
Coptotermes curvignathus terdapat di Asia Tenggara seperti Malaysia, Singapura, Indonesia (Jawa, Sumatra, Kalimantan), Thailand, Vietnam, Kamboja, Filipina, dan Cina bagian selatan. Rayap ini termasuk hama yang serius menyerang pohon karet hidup di Asia Tenggara dan menyerang kulit kayu dan bagian lain termasuk bagian dalam kayu, bahkan sering membunuh tanaman. Untuk mencapai makanannya rayap ini membuat saluran bawah tanah dan jika diperlukan mampu membentuk saluran-saluran yang ditutupi tanah yang dibuat di atas tanah (Roonwal 1970).
Simbiosis Pada Saluran Pencernaan Rayap dan Metabolisme Selulosa
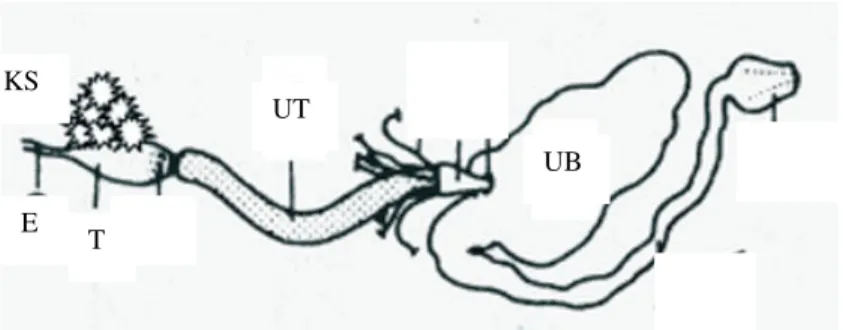
Saluran pencernaan rayap terdiri atas usus depan, usus tengah, dan usus belakang (Gambar 4). Saluran pencernaan ini menempati sebagian besar dari abdomen. Usus depan terdiri atas esofagus dan tembolok yang dilengkapi dengan kelenjar saliva. Esofagus dan tembolok memanjang pada bagian posterior atau bagian tengah dari thorak. Kelenjar saliva mensekresikan endoglukanase dan enzim lain ke dalam saluran pencernaan. Usus tengah merupakan bagian yang berbentuk tubular yang mensekresikan suatu membrane peritrofik di sekeliling material makanan. Usus tengah pada rayap tingkat juga diketahui mensekresikan endoglukanase. Usus belakang merupakan tempat bagi sebagian besar simbion (Noirot & Noirot-Timothée 1969; Scharf & Tartar 2008).
Gambar 3 Rayap C. curvignathus (a) pekerja dan (b) prajurit. (a)
1 mm
1 mm
Gambar 4 Morfologi saluran pencernaan rayap (Scharf & Tartar 2008). KS=kelenjar saliva, E=esofagus, T=tembolok, UT=usus tengah, UB=usus belakang.
Rayap bersimbiosis dengan bakteri dan protozoa pada saluran pencernaannnya. Pada rayap tingkat rendah lebih banyak bersimbiosis dengan protozoa dibandingkan dengan bakteri, sebaliknya pada rayap tingkat tinggi lebih banyak bersimbiosis dengan bakteri dibandingkan dengan protozoa (Krishna 1969; Bignell 2000; Breznak 2000).
Protozoa yang bersimbiosis dengan rayap tingkat rendah berbeda pada tiap spesies. Zootermopsis angusticollis bersimbiosis dengan Tricercomitis, Hexamastix, dan Trichomitopsis. Mastotermes darwiniensis bersimbiosis dengan Mixotricha paradoxa (Breznak 2000). Coptotermes formosanus bersimbiosis dengan Pseudotrichonympha grassii, Spirotrichonympha leidy, Holomastigoides mirabile (Inoue et al. 2005; Nakashima et al. 2002b), dan Holomastigoides hartmanni (Tanaka et al. 2006). Coptotermes lacteus bersimbiosis dengan Holomastigoides mirabile (Watanabe et al. 2002). Reticulitermes speratus bersimbiosis dengan Teranympha mirabilis, Triconympha agilis (Ohtoko et al. 2000), Dinenympha exilis dan Pyrsonympha grandis (Todaka et al. 2007).
Rayap dapat bertahan hidup dengan memanfaatkan lignoselulosa yang mengandung sedikit nutrisi (Ohkuma 2003). Rayap dapat mencerna lignoselulosa dengan menggunakan enzim selulase yang dihasilkan oleh rayap itu sendiri dan simbion (Watanabe et al. 1998; Ohkuma 2003; Scharf & Tartar 2008).
Lignoselulosa adalah suatu campuran dari tiga polimer yang dihasilkan tanaman yaitu selulosa, hemiselulosa, dan lignin. Selulosa adalah polimer glukosa yang terikat oleh ikatan β-1,4 dan terikat bersama dengan hemiselulosa (Scharf & Tartar 2008). UB UT KS T E
Selulase merupakan enzim ekstraseluler yang terdiri atas kompleks endo-ß-1,4-glukanase (endoglukanase), kompleks ekso-ß-endo-ß-1,4-glukanase (eksoglukanase) yang terdiri dari cellobiohydrolase (EC 3.2.1.91) atau cellodextranase (EC 3.2.1.74), dan ß-glukosidase (EC 3.2.1.21). Endoglukanase aktif dalam menghancurkan kristal atau selulosa murni. Eksoglukanase menghidrolisis selulosa non-kristal atau turunan selulosa terlarut. ß-1,4-glukosidase menghidrolisis selobiosa menjadi glukosa (Wood & Garcia-Campayo 1990).
Endoglukanase rayap termasuk ke dalam GHF 9. Anggota GHF 9 rayap terdiri dari endoglukanase dari rayap tingkat tinggi (Tokuda et al. 1999, 2004; Watanabe & Tokuda 2001) dan rendah (Nakashima et al. 2002b). Anggota GHF 9 famili selulase yang secara taksonomi tersebar luas, termasuk pada tanaman, bakteri, slime mold, dan rayap. Selulase simbion diklasifikasikan menjadi beberapa famili GHF yaitu 3, 5, 7, 8, 10, 11, 26, 43, 45, 62. Selulase simbion dari C. formosanus termasuk anggota GHF 5 (Inoue et al. 2005) dan GHF 7 (Nakashima et al. 2002a). Selulase simbion dari C. lacteus (Watanabe et al. 2002) dan R. flavipes (Zhou et al. 2007) termasuk anggota GHF 7. Selulase simbion dari R. speratus termasuk anggota GHF 3, 5, 7, 8, 10, 11, 26, 43, 45, dan 62 (Ohtoko et al. 2000; Todaka et al. 2007). Selulase simbion dari M. darwiniensis termasuk anggota GHF 45 (Li et al. 2003) dan selulase bakteri simbion Nasutitermes termasuk anggota GHF 11 (Brennan et al. 2004).
Rayap tingkat rendah secara utama memakan kayu yang kaya selulosa kristalin dan amorf, hemiselulosa dan lignin. Tingkat hidrolisis hemiselulosa dan lignin yang tinggi dapat membatasi aktivitas selulolitik melalui penghambatan produk akhir. Hal ini yang menyebabkan rayap mengekspresikan berbagai macam gen selulase. Pada R. speratus dan R. flavipes dihasilkan selulase yang termasuk GHF 7, 9, dan 45 (Ohtoko et al. 2000; Zhou et al. 2007).
Struktur Ekson-Intron dan Ekspresi Gen Endoglukanase Rayap
Struktur ekson-intron gen endoglukanase rayap belum banyak diketahui. Sebagian besar data yang ada merupakan data sebagian (partial) cDNA dan cDNA lengkap. Data genomik DNA lengkap pada N. takasagoensis (Tokuda et al. 1999) dan data partial genomik gen endoglukanase terdapat pada R. speratus (Itakura et al. 2006). Genom DNA NtEG memiliki panjang 13005 pb terdiri atas
sepuluh ekson dengan sembilan intron. Semua intron NtEG diawali dengan GT dan diakhiri dengan AG. Pada partial genomik RsEG diketahui ukuran dari tiga intron. Dua intron pada RsEG memiliki posisi yang identik dengan NtEG yaitu intron tujuh dan delapan (Tokuda et al. 1999). Itakura et al. (2006) mengkarakterisasi intron sembilan RsEG sebesar 529 pb. Situs pemotongan intron 10 berbeda dengan situs pemotongan eukariot pada umumnya (GT/AG), yaitu diawali dengan AG dan diakhiri dengan GC.
Data cDNA lengkap terdiri atas R. flavipes (Scharf et al. 2005), R. speratus (Tokuda et al. 1999), C. formosanus (Nakashima et al. 2002b), C. acinaciformis, N. walkeri (Tokuda et al. 1999). Data partial cDNA terdiri atas M. darwiniensis (Tokuda et al. 1999; Li et al. 2003), H. sjoestedti (Tokuda et al. 2004), H. japonica (Lo et al. 2000), Glyptotermes sp. (Tokuda et al. 1999), N. koshunensis (Lo et al. 2000; Tokuda et al. 2004), R. speratus (Watanabe et al. 1998; Itakura et al. 2006), S. mushae, dan O. formosanus (Tokuda et al. 2004).
Endoglukanase rayap diekspresikan pada saluran pencernaan rayap. Terdapat perbedaan ekspresi endoglukanase pada rayap tingkat rendah dan rayap tingkat tinggi. Ekspresi NtEG dan NwEG yang termasuk ke dalam rayap tingkat tinggi terjadi di usus tengah (Tokuda et al. 1999). Sedangkan pada rayap tingkat rendah terjadi perbedaan ekspresi dari tiap spesies. Ekspresi RsEG terjadi di kelenjar saliva/usus depan (Tokuda et al. 1999), CfEG diekspresikan di kelenjar saliva, usus depan, dan usus tengah, dan RfEG terdapat pada bagian kelenjar saliva, usus depan, usus tengah, dan usus belakang (Zhou et al. 2007). RfEG diekspresikan paling tinggi pada pekerja kemudian nimfa. Ekspresi EGase menurun pada laron tetapi meningkat 50% pada kasta reproduktif sekunder (Scharf et al. 2005).
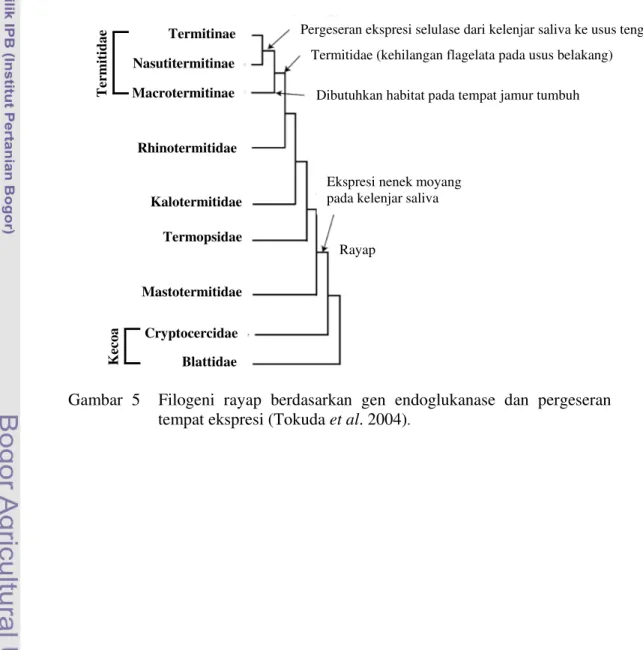
Secara filogeni, taksa rayap yang berada pada bagian basal secara konsisten mengekspresikan selulase secara spesifik pada kelenjar saliva (Mastotermitidae, Termopsidae, Kalotermitidae, Rhinotermitidae). Sedangkan taksa yang lebih maju mengekspresikan selulase terutama pada bagian usus tengah (Termitidae) (Gambar 5). Hal ini menunjukkan adanya pergeseran tempat ekspresi selulase dari kelenjar saliva ke usus tengah selama proses evolusi (Tokuda et al. 2004).
Aktivitas selulase rayap tingkat rendah dan rayap tingkat tinggi berbeda. Pada rayap tingkat rendah M. darwiniensis (Mastotermitidae), H. sjoestedti (Termopsidae), N. koshunensis (Kalotermitidae), R. speratus (Rhinotermitidae), dan C. formosanus (Rhinotermitidae), aktivitas endoglukanase tertinggi ditemukan di kelenjar saliva (45-86%). Secara keseluruhan pada rayap tingkat rendah aktivitas endoglukanase tertinggi pada Kalotermitidae dan Rhinotermitidae. Sedangkan Termitidae yang termasuk rayap tingkat tinggi yaitu O. formosanus (Macrotermitinae), N. takasagoensis (Nasutitermitinae), dan S. mushae (Termitinae) aktivitas endoglukanase tertinggi ditemukan di usus tengah (96-99%) (Tokuda et al. 2004). Termitinae Nasutitermitinae Macrotermitinae Rhinotermitidae Kalotermitidae Termopsidae Mastotermitidae Cryptocercidae Blattidae
Pergeseran ekspresi selulase dari kelenjar saliva ke usus tengah Termitidae (kehilangan flagelata pada usus belakang)
Dibutuhkan habitat pada tempat jamur tumbuh
Ekspresi nenek moyang pada kelenjar saliva
Rayap
Gambar 5 Filogeni rayap berdasarkan gen endoglukanase dan pergeseran tempat ekspresi (Tokuda et al. 2004).
T er m it id a e K ec o a