Uji In-vitro Pengendalian Hayati oleh Trichoderma spp. terhadap
Ganoderma yang Menyerang Sengon
In-vitro Test of Biological Control by Trichoderma spp. Toward Ganoderma that attacked
Sengon
Elis Nina Herliyana
1, Ratna Jamilah
1, Darmono Taniwiryono
2dan Muhammad Alam Firmansyah
1Departemen Silvikultur Fakultas Kehutanan, IPB 2 Balai Penelitian Bioteknologi Perkebunan Indonesia
* Corresponding author:
E-mail : [email protected]; [email protected]
ABSTRACT
Ganoderma infection, red root-rot fungi or basal stem rot disease is becoming more prevalent and causing significant loss in sengon (Falcataria moluccana) in Indonesia. Trichoderma spp. is the alternative choice in the biological control of Ganoderma sp The objectives of the research were to study the potential of biological agents antagonism Trichoderma harzianum and Trichoderma pseudoconingii against Ganoderma that attacked Sengon with in vitro method. Antagonism test of Trichoderma spp with co-culture method modification to five Ganoderma isolates on PDA medium was conducted in Forest Pathology Laboratory, IPB, Bogor. The potential biological agents for protecting Ganoderma spp. on sengon were T. harzianum strain DT38 and T. pseudoconingii DT39. The research results showed that the Trichoderma spp. inhibit growth of five Ganoderma isolates that were attacked sengon tree from Lampung (Ganoderma L12, L6, L3) and Kalimantan Selatan (K2, dan K1) between 11,7 – 48,8%.
Keywords: Biological Control, Falcataria moluccana, Ganoderma sp., Trichoderma harzianum, Trichoderma pseudokoningii,
PENDAHULUAN
Peran Hutan Tanaman Industri (HTI) sebagai penyediaan bahan baku kayu di Indonesia kini semakin meningkat. Bagian terbesar kayu dari HTI luar Jawa adalah untuk kayu pulp yang didominasi oleh Acacia mangium, Acacia crassicarpa, Eucalyptus pellita, Eucalyptus urophylla dan Falcataria moluccana (sengon) (Rimbawanto 2008). Saat ini, sengon merupakan salah pohon yang banyak dikembangkan pada program pembangunan hutan, baik hutan tanaman maupun hutan rakyat di Indonesia. Pohon sengon mempunyai pertumbuhan yang cepat dan kegunaan kayunya yang beragam diantaranya untuk bahan konstruksi di bawah atap, mebel sederhana, peti kemas, dan papan sambung. Sengon juga mempunyai manfaat yang cukup besar dalam upaya rehabilitasi lahan kritis, maka pemerintah, melalui Departemen Kehutanan, telah lama mencanangkan program ‘sengonisasi’ (Atmosuseno 1998). HTI tersebut umumnya dilakukan secara monokultur dan seringkali dihubungkan dengan peningkatan resiko serangan hama penyakit, terutama bila benihnya berasal dari induk pohon yang secara genetik keragamannya rendah atau berkerabat (Rimbawanto 2008).
Serangan penyakit busuk akar yang disebabkan oleh jamur Ganoderma spp. pada tanaman sengon telah banyak dilaporkan (Solomon et al. 1993; Basset dan Peters 2003; Widyastuti 2007; Herliyana et al. 2012). Serangan Ganoderma sp. di lapangan sulit di deteksi
karena gejalanya mirip dengan gejala kekeringan. Meskipun tanaman sudah menunjukkan gejala sakit, namun terkadang tubuh buah Ganoderma spp. belum terbentuk. Dan di lain pihak, pada tanaman yang tampak sehat ditemukan tubuh buah Ganoderma spp. di pangkal batangnya (Bassett dan peters 2003).
Jamur pelapuk kayu dan penyebab penyakit pada pohon hutan sebagian besar berasal dari beberapa spesies Ganoderma sp.. Ganoderma sp. merupakan jamur tingkat tinggi yang tergolong dalam kelas Basidiomycetes ordo Polyporanceae family Ganodermataceae. Ganoderma sp. mempunyai daerah penyebaran tempat tumbuh yang cukup luas dan dikenal sebagai penyebab penyakit akar pada banyak jenis tanaman berkayu. Di hutan alam, jamur ini cenderung menyerang pohon-pohon tua atau yang telah mengalami penurunan pertumbuhan, dan juga dapat menyebabkan pembusukan kayu yang sudah mati. Pada HTI dan perkebunan jamur ini telah dilaporkan menjadi patogen akar yang potensial dan telah banyak menyerang beberapa jenis tanaman (Semangun 2000). Pada tahun 2009 sampai 2011, tubuh buah Ganoderma ditemukan pada beberapa pohon sengon di daerah Lampung dan Kalimantan Selatan. Di Lampung, selain itu juga ditemukan pada jengkol, nangka, alpukat dan kihiang. Di Kalimantan juga ditemukan pada pohon lainnya (Herliyana et al. 2009, tidak dipublikasikan).
Infeksi patogen pada tanaman umumnya lebih mudah terjadi melalui luka dan lentisel, yang sering ditemukan pada bagian leher akar yang pecah, dan ini Vol. 04 No. 3 Desember 2013, Hal. 190 – 195
merupakan tempat yang baik bagi infeksi jamur. Patogen kemudian menyebar ke bagian yang lebih dalam dari akar. Serangan akan lebih tinggi ditemukan pada tanaman hasil okulasi dibandingkan dengan tanaman hasil biji. Hal ini disebabkan pada tanaman hasil okulasi ada bagian-bagian luka, sehingga memudahkan Ganoderma sp. untuk mengadakan infeksi (Sinulingga 1989). Infeksi atau penularan penyakit ini terjadi melalui kontak akar tanaman sehat dengan sumber infeksi di dalam tanah seperti potongan akar dan batang yang mengandung koloni patogen (Widyastuti 2007; Herliyana et al. 2012, Herliyana 2012).
Pengendalian hayati menggunakan agen antagonis dengan satu kali pemakaian dapat menekan pertumbuhan dan perkembangan patogen untuk jangka waktu yang relatif panjang tanpa menimbulkan pencemaran lingkungan (Baker dan Cook 1974; Achmad et al. 2009). Pengendalian hayati merupakan pengendalian penyakit yang ramah lingkungan karena bersifat tidak membahayakan kehidupan makhluk hidup dan lingkungan. Pengendalian Ganoderma sp. secara hayati dapat dilakukan dengan cara pemanfaatan mikroorganisme atau agens antagonis Trichodema sp. merupakan pilihan alernatif yang dapat meminimalkan gangguan terhadap keseimbangan biologis disamping menurunkan biaya pengendalian. Oleh karena itu perlu dilakukan penelitian untuk mengetahui potensi Trichoderma spp. sebagai agen pengendalian Ganoderma sp. secara hayati.
Tujuan penelitian ini adalah untuk mengetahui potensi atau efektifitas agens antagonis Trichoderma spp. dalam menghambat pertumbuhan Ganoderma sp. yang menyerang sengon yang berasal dari Kalimantan Selatan dan Lampung. Penelitian ini diharapkan dapat memberikan informasi yang berkaitan dengan kemampuan isolat Trichoderma sp. sebagai agen antagonis terhadap Ganoderma sp.. Hasil penelitian diharapkan dapat diaplikasikan dalam budidaya sengon, sehngga untuk perkembangan selanjutnya dapat meningkatkan kualitas dari tanaman sengon.
BAHAN DAN METODE Waltu dan Tempat
Kegiatan uji antagonis Trichoderma spp. terhadap Ganoderma sp. dilaksanakan di Laboratorium Pathologi Hutan IPB dan Laboratorium Balai Penelitian Bioteknologi Perkebunan Indonesia, Bogor.
Bahan dan Alat
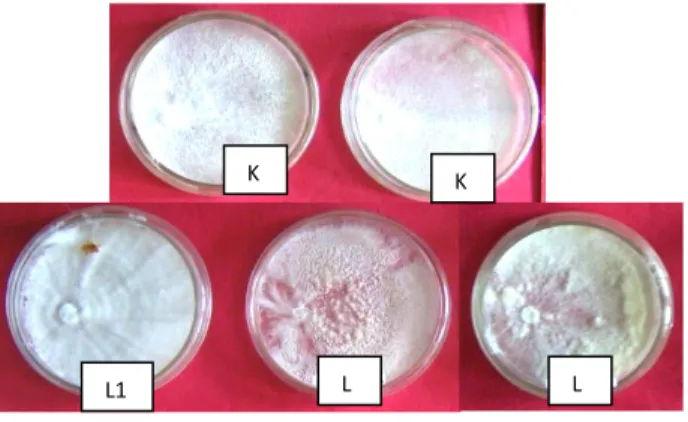
Bahan dan alat yang digunakan dalam penelitian ini antara lain, media PDA Steril, isolat patogen Ganoderma sp. yang menyerang sengon di Kalimantan Selatan (K1 dan K2), isolat Ganoderma sp. yang menyerang sengon di, di Lampung (L12, L3, dan L6)
(Gambar 1)
, isolat agens antagonis Trichodermaharzianum (T38) dan Trichoderma pseudokoningii
(T39) koleksi Dr Ir Darmono Taniwiryono
(Gambar 2)
, cawan Petri, autoclave, dan laminar air flow.Gambar 1 Isolat Ganoderma sp. yang menyerang sengon di Kalimantan Selatan (K1 dan K2), isolat Ganoderma sp. yang menyerang sengon di, di Lampung (L12, L3, dan L6).
Gambar 2 Isolat Trichoderma
harzianum (T38) dan
Trichoderma pseudokoningii (T39).
Metode PenelitianMetode Uji Antagonis
Uji antagonisme dalam penelitian ini dilakukan dengan cara metode biakan ganda dengan perbandingan 1 : 1 secara in-vitro dalam satu cawan konfrontasi atau modifikasi co-culture method (Johnson 1957 dalam Widyastuti 2007). Koloni patogen Ganoderma sp. diinokulasikan dalam cawan konfrontasi terlebih dahulu sebelum memasukan koloni cendawan antagonis dengan masa inkubasi selama 3 sampai 5 hari sehingga isolat patogen Ganoderma sp. berukuran cukup besar. Kemudian isolat antagonis Trichoderma sp. ditumbuhkan pada cawan konfrontasi pada sisi yang berlawanan dengan jarak 4 cm dari koloni fungi patogen (Gambar 3). Jari-jari koloni dari kedua isolat diukur panjangnya setiap 24 jam sampai hari kelima semenjak kedua isolat disatukan. Kemudian setelah hari kelima dilakukan pengamatan zona penghambatan dan persen penghambatan.
Zona penghambatan adalah panjang wilayah dalam cawan konfrontasi yang tidak ditumbuhi oleh kedua isolat yang saling antagonis. Pengukuran dilakukan dengan cara mengukur panjang dari zona kosong tersebut. Persentase Penghambatan dihitung dengan rumus yang dipakai Rohana (1998) :
K 1 L1 2 L 3 L 6 K 2 T38 T39
Gambar 3 Pola Penempatan isolat Ganoderma sp. dan Trichoderma sp. pada cawan konfrontasi. Kelima isolat Ganoderma sp. (L12, L6, L3, K2, dan K1) ditanamkan dalam cawan konfrontasi bersama dengan isolat Trichoderma sp. (T38 dan T39). Masing-masing perlakuan memiliki tiga ulangan. Perlakuan yang digunakan dalam penelitian ini dijelaskan pada Tabel 1.
Tabel 1 Jenis perlakuan uji antagonis
Perlakuan Keterangan
L12 x T38 Isolat Ganoderma Sp L12 ditanamkan bersama Isolat Trichoderma sp. T38 L12 x T39 Isolat Ganoderma Sp L12 ditanamkan
bersama Isolat Trichoderma sp. T39 L6 x T38 Isolat Ganoderma Sp L6 ditanamkan
bersama Isolat Trichoderma sp. T38 L6 x T39 Isolat Ganoderma Sp L6 ditanamkan
bersama Isolat Trichoderma sp. T39 L3 x T38 Isolat Ganoderma Sp L3 ditanamkan
bersama Isolat Trichoderma sp. T38 L3 x T39 Isolat Ganoderma Sp L3 ditanamkan
bersama Isolat Trichoderma sp. T39 K1 x T38 Isolat Ganoderma Sp K1 ditanamkan
bersama Isolat Trichoderma sp. T38 K1 x T39 Isolat Ganoderma Sp K1 ditanamkan
bersama Isolat Trichoderma sp. T39 K2 x T38 Isolat Ganoderma Sp K2 ditanamkan
bersama Isolat Trichoderma sp. T38 K2 x T39 Isolat Ganoderma Sp K2 ditanamkan
bersama Isolat Trichoderma sp. T39
Analisis Statistik
Variabel yang diamati dalam penelitian ini adalah persen penghambatan. Penelitian ini menggunakan analisis statistik dengan model RAL (Rancangan Acak Lengkap) dua faktor. Faktor yang digunakan dalam penelitian ini adalah faktor jenis isolat Ganoderma sp. dan faktor jenis isolat Trichoderma sp.. Faktor jenis isolat Ganoderma sp. terdiri dari isolat L12, L6, L3, K2, dan K1. Faktor jenis isolat Trichoderma sp. terdiri dari isolat T38 dan T39. Pengolahan data menggunakan software SAS 9.
Pengolahan data dimulai dengan analisis sidik ragam dengan tingkat kepercayaan 95%, apabila blok atau
faktor berbeda nyata, pengolahan data dilanjutkan dengan uji lanjut. Uji lanjut digunakan untuk membandingkan perlakuan mana yang paling baik dalam percobaan. Pengujian lanjut ini menggunakan uji Duncan karena uji inilah yang paling sering digunakan dalam pengujian lanjut analisis sidik ragam. Analisis Statistik bertujuan untuk melihat pengaruh dari masing masing isolat Ganoderma sp. dan Trichoderma sp. terhadap variabel persen penghambatan.
HASIL DAN PEMBAHASAN
Kedua cendawan agens antagonis isolat
Trichoderma
T38
dan
T39
mempunyai
pertumbuhan diameter koloni rata-rata lebih cepat
dibanding kelima isolat patogen Ganoderma pada
media PDA. Trichoderma T38 dan T39 dapat
memenuhi permukaan cawan Petri berukuran
diameter 9 cm dalam waktu lebih kurang 4 (empat)
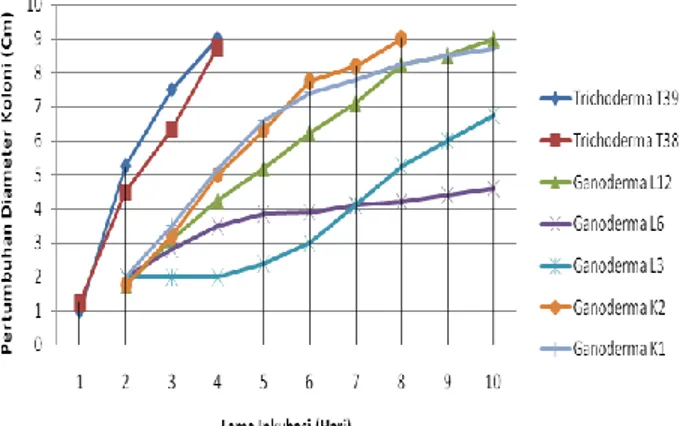
hari inkubasi. Pertumbuhan diameter koloni
isolat-isolat Ganoderma dari yang paling cepat ke yang
paling lambat berturut-turut adalah Ganoderma
K2, L12, K1, L3 and L6. Isolat-isolat Ganoderma
dapat memenuhi permukaan cawan Petri berukuran
diameter 9 cm dalam waktu lebih dari 8 (delapan)
hari inkubasi (Gambar 4).
Gambar 4 Pertumbuhan diameter koloni dua isolat cendawan antagonis Trichoderma dibanding dengan lima isolat patogen Ganoderma pada media PDA.
Perlakuan kontrol koloni Ganoderma sp. menunjukan pertumbuhan jari-jari koloni patogen yang lebih lambat dibandingkan pertumbuhan jari-jari koloni Trichoderma sp.. Setelah dilakukan pengujian pada satu cawan konfrontasi, hasil pengamatan visual uji antagonisme Trichoderma sp. dengan Ganoderma sp. memperlihatkan bahwa pertumbuhan jari-jari koloni Ganoderma sp. ke arah titik tengah cawan konfrontasi lebih lambat dibanding pertumbuhan jari-jari koloni Trichoderma sp.. Purwantisari dan Hastuti (2009) menyatakan bahwa Trichoderma sp. merupakan jenis yang potensial untuk pengendalian penyakit secara hayati. Hasil penelitian yang dilakukan mendukung pendapat tersebut, dimana Trichoderma sp. T38 dan
r1-r2
PP = --- x 100%
r1
Keterangan: PP = Persentase Penghambatan r1 = Jari-jari 1 isolat r2 = Jari-jari 2 isolat Keterangan P = isolat Patogen A = isolat antagonis t = titik tengah cawan Petri r1 = radius 1 pertumbuhan isolat P r2 = radius 2 pertumbuhan isolat Ar
1
r
2
r
1
r
2
r1
r2
P
A
T39 mampu menghambat pertumbuhan koloni patogen Ganoderma spp. berdasarkan hasil uji in-vitro pada PDA (Gambar 5). Cendawan yang tumbuh cepat mampu berkompetisi mengungguli dalam penguasaan ruang dan nutrisi pada akhirnya bisa menekan pertumbuhan cendawan lawannya. Kompetisi tersebut dalam memperoleh nitrogen dan karbon (Harman et al. 2008).
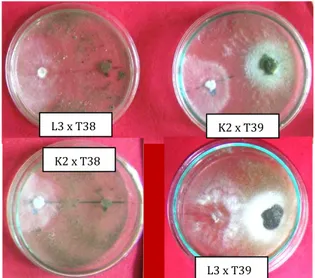
Gambar 5 Pengamatan visual uji antagonisme hari ke 5 inkubasi. Gambar atas kiri dan kanan: Patogen L3 x antagonis T38 dan patogen K2 x antagonis T39. Gambar bawah kiri dan kanan: Patogen K2 x antagonis T38 dan patogen L3 x antagonis T39.
Dari kedua jenis jamur antagonis yang diuji, Trichoderma sp. T39 memiliki potensi antagonistik yang kuat dibandingkan dengan Trichoderma sp. T38. Isolat Ganoderma sp. L12 dan L6 merupakan isolat yang pertumbuhannya paling terhambat dibanding isolat yang lain (Gambar 6) (Tabel 2).
Gambar 6 Persen penghambatan secara in-vitro pertumbuhan ke-lima isolat Ganoderma oleh Trichoderma T38 and T39 pada media PDA.
Tabel 2 Hasil uji Duncan pengaruh jenis isolat Trichoderma sp. terhadap persen penghambatan Jenis isolat Trichoderma sp. Rata-rata persen penghambatan (%) T39 32.6 a T38 27.4 a
Keterangan: Huruf yang sama dibelakang angka menunjukkan pengaruh yang tidak berbeda nyata pada tingkat kepercayaan 95%.
Berdasarkan hasil penelitian yang didapatkan, faktor yang lebih mempengaruhi daya penghambatan adalah faktor jenis isolat Ganoderma sp. (Tabel 3 dan 4). Perbedaan daya hambat menggambarkan perbedaan kemampuan dari masing-masing isolat untuk menghambat pertumbuhan mikrooganisme pesaing. Perbedaan ini diduga dipengaruhi oleh jenis, jumlah, dan kualitas dari antibiotik atau zat lain yang dihasilkan Trichoderma sp. yang dapat menghambat pertumbuhan Ganoderma sp.. Adanya penghambatan terhadap pertumbuhan diameter koloni patogen Ganoderma sp. diduga karena adanya enzim dan senyawa metabolit yang terdapat pada fungi antagonis Trichoderma sp. yang mampu merusak dinding sel patogen Ganoderma sp. sehingga menyebabkan pertumbuhan diameter koloni patogen menjadi lambat. Menurut Harman et al. (2008), Trichoderma spp. mempunyai mekanisme pengendalian utama sebagai mikoparasit atau memarasit miselium cendawan lain dengan menembus dinding sel dan masuk ke dalam sel untuk mengambil zat makanan dari dalam sel sehingga cendawan akan mati.
Tabel 3 Rekapitulasi hasil sidik ragam terhadap variabel yang diamati
Variabel Isolat Ganoderma sp. (G) Isolat Trichoderma sp. (T) G*T Persen peng-hambatan * tn tn
Keterangan : ***= Berbeda nyata pada taraf uji 0,01; **= Berbeda nyata pada taraf uji 0,05; *= Berbeda nyata pada taraf uji 0,1; tn= Tidak berbeda nyata
Tabel 4 Hasil uji Duncan pengaruh jenis isolat Ganoderma sp. terhadap persen penghambatan
Jenis isolat Ganoderma sp. Rata-rata persen penghambatan (%) L6 39.6 a L12 38.1 a L3 30.4 ab K2 29.4 ab K1 12.4 b
Keterangan: Huruf yang sama dibelakang angka menunjukkan pengaruh yang tidak berbeda nyata pada tingkat kepercayaan 95%.
L3 x T38 K2 x T39
K2 x T38
Terbentuknya zona penghambatan antar organisme pada media padat merupakan indikasi bekerjanya mekanisme antibiosis. Bekerjanya mekanisme antibiosis dikuatkan oleh tertekannya pertumbuhan cendawan patogen pada media padat. Harman et al. (2008) melaporkan juga bahwa Trichoderma spp. dapat menghasilkan antibiotik seperti alametichin, paracelsin, trichotoksin yang dapat menghancurkan sel cendawan melalui pengrusakan terhadap permeabilitas membran sel, serta menghasilkan enzim khitinase yang dapat menyebabkan lisis dinding sel serta dapat melakukan interfensi hifa. Pada penelitian ini tidak ditemukannya zona penghambatan, hal ini dapat disebabkan karena media yang digunakan adalah PDA. PDA dapat menetralisir pengaruh metabolit penghambat pertumbuhan patogen (Achmad 1991). Tetapi dapat juga disebabkan tidak terjadinya mekanisme antibiosis, dan Trichoderma sp. diduga tidak menghambat pertumbuhan koloni Ganoderma sp. setelah terjadi kontak hifa.
Menurut Smith dan Moss (1985), beberapa anggota genus Trichoderma sp. menghasilkan toksin (mycotoxin) yaitu trichodermin. Toksin ini dihasilkan oleh cendawan, bila berada atau hidup pada tanaman hidup, bahan yang mengurai, dan produk-produk yang disimpan di gudang. Selain itu, adanya aktifitas metabolik hifa yang tinggi pada bahan organik dapat juga menyerang dan menghancurkan propagul patogen yang ada disekitarnya (Lewis dan Papavizas 1984).
Lambatnya pertumbuhan diameter koloni patogen Ganoderma sp. pada perlakuan pemberian fungi antagonis Trichoderma sp. diduga karena telah terjadi reaksi antara senyawa toksik dari fungi antagonis Trichoderma sp terhadap patogen Ganoderma sp. Trichoderma sp. adalah suatu jenis yang baik sebagai pengendali hayati karena terdapat di mana-mana, mudah diisolasi dan dibiakkan, tumbuh dengan cepat pada beberapa macam substrat, mempengaruhi patogen tanaman, jarang bersifat patogenik pada tanaman tingkat tinggi, bereaksi sebagai mikroparasit, bersaing dengan baik dalam hal makanan, tempat dan menghasilkan antibiotik (Wells 1988). Anggraeni (2004) menyatakan bahwa Trichoderma sp. dapat digunakan sebagai agen biokontrol melawan beberapa cendawan petogenik tular tanah. Selama Trichoderma sp. tumbuh aktif menghasilkan sejumlah besar enzim ekstra selular ß (1.3) glukonase, dan kitinase, yang dapat melarutkan dinding sel patogen (Lewis dan Papavizas 1984).
KESIMPULAN DAN SARAN Kesimpulan
Dari kedua jenis jamur antagonis yang diuji Trichoderma sp. T39 memiliki potensi antagonistik yang lebih kuat dibanding Trichoderma sp. T38 dengan kemampuan menghambat pertumbuhan jamur patogen dengan persentase penghambatan lebih tinggi pada isolat Ganoderma sp. L12 dan L6. Uji antagonis pada media PDA ini tidak diperoleh zona penghambatan.
Saran
Pengamatan mikroskopis disarankan dilakukan juga. Penelitian ini bisa dilanjutkan dengan penelitian lanjutan yaitu berupa pembuatan formula dan aplikasinya di lapangan.
UCAPAN TERIMAKASIH
Penelitian ini merupakan bagian dari penelitian kami yang didanai KKP3T Badan Litbang Pertanian tahun 2009. Penulis ingin mengucapkan terimakasih kepada semua pihak yang tidak dapat disebut satu persatu, yang telah membantu dalam penelitian ini.
DAFTAR PUSTAKA
Achmad S. 1991. Kemampuan Rhizopogon sp. untuk Perlindungan Hayati terhadap Penyebab Penyakit Lodoh pada Pinus merkusii [Tesis]. Bogor: Program Pascasarjana, IPB.
Achmad, S. Hadi, S. Harran, E. Gumbira Sa’id, B. Satiawihardja, M. Kosim Kardin. 2009. Pengendalian Hayati Penyakit Lodoh Pada Semai Pinus Merkusii : Potensi Antagonistik In-vitro Trichoderma harzianum DAN Trichoderma pseudokoningii. Jurnal Litbang Tanaman.
Anggraeni I. 2004. Identifikasi dan Patogenitas Penyakit Akar pada Acacia mangium Willd. Buletin Penelitian Hutan. 645: 61-73.
Atmosuseno BS. 1998. Budi Daya, Kegunaan, dan Prospek Sengon. Jakarta : Penebar Swadaya. Baker KF, Cook RJ. 1974. Biology Control of Plant
Pathogens. San Fransisco: W.H. Freeman and Co. Basset K, Peters. 2003. Ganoderma: A Significant Root
Pathogen. Arborilogical Services Inc. Publication. http://www.arborilogical.com/articles/ganoderma.ht m. [6 Februari 2010].
Harman GE, Björkman T, Ondik K, Shoresh M. 2008. Trichoderma spp. for Biocontrol. Changing paradigms on the mode of action and uses of Trichoderma spp. for biocontrol. Research Information. Cornell University, USA. DOI:10.1564/19feb00.
Herliyana EN, Taniwiryono D, Minarsih H, Firmansyah MA, Sukamto S. 2009. Pengembangan Teknik Perlindungan Tanaman Sengon Sebagai Pelindung Tanaman Kopi dan Kakao dari Serangan Ganoderma spp.. Laporan Akhir KKP3T. IPB dan Badan Litbang Pertanian. 73 halaman.
Herliyana EN, Taniwiryono D, Minarsih H. 2012. Root diseases Ganoderma sp. on the Sengon in West and East Java. Journal of Tropical Forest Management 18 (2):94-99. DOI:10.7226/jtfm.18.2.94.
Herliyana EN. 2012. Early Report of Red Root Rot of Ganoderma sp. on Agathis sp. (Damar) in Mount Walat Education Forest, Sukabumi, West Java. Jurnal Silvikultur Tropika 3(2): 102-107.
Lewis JA, Papavizas GC. 1983. Production of Clamidospores and Conidia by Trichoderma sp. In Liquid and Solid Growth Media. J. Soil Biology and Biochemistry, 15 (4): 351-357.
Purwantisari S, Hastuti RB. 2009. Uji Antagonisme Jamur Patogen Phytophthora infestans Penyebab Penyakit Busuk Daun dan Umbi Tanaman Kentang Dengan Menggunakan Trichoderma spp. Isolat Lokal. J. BIOMA 11(1): 24-32.
Rimbawanto A. 2008. Pemuliaan tanaman dan ketahanan penyakit pada sengon. Yogyakarta: Balai Besar Penelitian Bioteknologi dan Pemuliaan Tanaman Hutan.
Rohana I. 1998. Efektifitas Penggunaan Trichoderma harzianum dan Fungisida Mankozeb untuk Pengendalian Rhizoctonia solani Penyebab Penyakit Lodoh pada Acacia mangium [Skripsi]. Fakultas Kehutanan IPB, Bogor.
Semangun H. 2000. Penyakit-Penyakit Tanaman Perkebunan Indonesia. Yogyakarta: Gajah Mada Univ Press.
Sinulingga, W. 1989. Pengendalian Biologi Penyakit Cendawan Akar Putih pada Tanaman Karet. Pusat Penelitian Perkebunan Sei. Putih, Galang. Hal. 1-7. Smith JE, Moss MO. 1985. Mycotoxin, Formation
Analysis and Significant. John Willey and Sons, inc. New York.
Solomon, JD, Leininger TD, Wilson AD, Anderson RL, Thompson LC, McCracken FI. 1993. Ash Pests: A Guide to Major Insect, Diseases, Air Pollution Injury and Chemical Injury. Gen. Tech. Rep. SO-96. New Orleans, LA: U.S. Department of Agriculture, Forest Service , Southern Forest Experiment Station. 45p.
Widyastuti SM. 2007. Peran Trichoderma spp. Dalam Revitalisasi Kehutanan di Indonesia. Yogyakarta: Gajah Mada University Press, 255p.
Wells HD. 1998. Trichoderma as A Biocontrol Agent. dalam Biocontrol of Plant Disease, Vol 1. Mukerji KG, Garg KL (ed.). CRC Press, Inc. Boca Raton, Florida. 72-79.