SEMINAR KIMIA
Judul : Pengaruh Malation terhadap gangguan fungsi mitokondria
Bidang : Biokimia
Penyaji : Anis Ro’iyatunisa
Pembimbing : Dr. Heli Siti Halimatul Munawaroh, S.Pd., M.Si. Tempat : Laboratorium Kimia Instrumen (LKI)
ABSTRAK
Disfungsi mitokondria pada keracunan akut akibat pestisida organofosfat dapat menyebabkan kematian pada manusia. Malation merupakan salah satu jenis insektisida organofosfat yang digunakan untuk membunuh serangga. Pada makalah ini dibahas mengenai pengaruh malation terhadap gangguan fungsi mitokondria. Dari hasil penelitian diperoleh bahwa paparan malation sebanyak 400mg/kg berat tikus, dapat menghambat aktivitas enzim asetilkolinesterase (AChE) sebanyak 70%. Asetilkolin (ACh) yang tidak terdegredasi menumpuk pada sinapsis saraf dan neuromuscular junction (NMJ) yang dapat mengakibatkan kelemahan otot atau yang disebut miopati mitokondria. Malation dapat mengakibatkan penurunan aktivitas beberapa enzim yang terlibat dalam rantai respirasi mitokondria (Mitochondria Respiratory Chain) serta jalur metabolisme. Malation juga dapat menghambat sinyal apoptosis pada mitokondria (kematian sel yang terprogram untuk regenerasi sel). Toksisitas malation sangat cepat, karena memiliki daya absorbsi yang baik dan afinitas yang tinggi terhadap enzim AChE.
Kata kunci : disfungsi mitokondria , malation, asetilkolinesterase (AChE), asetilkolin (Ach), neuromuscular junction (NMJ), Mitochondria Respiratory Chain (MRC).
DAFTAR PUSTAKA
Mohajeri, S.K. et al. (2014). “Mechanisms of muscular electrophysiological and mitochondrial dysfunction following exposure to malation, an organophosphorus pesticide”. Journal of Hum Exp Toxicol. 33, (3), 251–263.
Mohajeri, S.K, dan Abdollahi, M. (2013). “Mitochondrial dysfunction
and organophosphorus compounds”. Journal of Toxicology and
Applied Pharmacology, 270,39–44.I. PENDAHULUAN
Jumlah penduduk dari tahun ketahun terus mengalami peningkatan. Menurut proyeksi data statistik Indonesia tahun 2013 jumlah penduduk Indonesia mencapai 242 juta jiwa, sedangkan pada tahun 2014 diperkirakan jumlah penduduk Indonesia meningkat menjadi 244,8 juta jiwa (Data Statistik Indonesia, 2014). Pertambahan penduduk ini tentunya berbanding lurus dengan kebutuhan pangan. Dalam rangka mencukupi kebutuhan tersebut, pemerintah Indonesia mencanangkan beberapa program dibidang pertanian. Salah satunya adalah program intensifikasi tanaman pangan. Program ini tentu ditunjang dengan perbaikan teknologi pertanian, penggunaan varietas, perbaikan teknik budidaya yang meliputi pengairan, pemupukan dan pengendalian hama penyakit. Awalnya program ini berjalan dengan baik, tetapi lambat laun muncul berbagai masalah seperti munculnya hama pada tanaman pangan. Untuk memecahkan masalah tersebut, dilakukanlah upaya untuk mematikan hama dengan pemakaian pestisida (Rini, 2007).
II. PENGARUH PESTISIDA MALATION TERHADAP GANGGUAN FUNGSI MITOKONDRIA
A. Pestisida malation 1. Pengertian pestisida
Secara harfiah pestisida berasal dari kata pest: hama dan cide: membunuh, jadi pestisida diartikan sebagai pembunuh hama. Berdasarkan SK Menteri Pertanian RI No. 434.1/Kpts/TP.270/7/2001 tentang syarat dan tata cara pendaftaran pestisida, yang dimaksud dengan pestisida adalah semua zat kimia atau bahan lain serta jasad renik dan virus yang digunakan untuk beberapa tujuan berikut :
a) memberantas atau mencegah hama dan penyakit yang merusak tanaman, bagian tanaman, atau hasil-hasil pertanian;
b) memberantas rerumputan;
c) mematikan daun dan mencegah pertumbuhan yang tidak diinginkan;
d) mengatur atau merangsang pertumbuhan tanaman atau bagian-bagian tanaman (tetapi tidak termasuk golongan pupuk).
Toksisitas atau daya racun pestisida merupakan sifat bawaan pestisida yang menggambarkan potensi pestisida untuk menimbulkan kematian langsung (atau bahaya lainnya) pada hewan tingkat tinggi, termasuk manusia. Toksisitas dibedakan menjadi toksisitas akut dan toksisitas kronik. Toksisitas akut merupakan pengaruh merugikan yang timbul segera setelah pemaparan dengan dosis tunggal suatu bahan kimia atau pemberian dosis ganda dalam waktu kurang lebih 24 jam. Toksisitas akut dinyatakan dalam angka LD50 (lethal dose), yaitu dosis yang bisa mematikan 50% dari binatang uji (umumnya tikus) yang dihitung dalam mg/kg berat badan. Toksisitas kronik adalah pengaruh merugikan yang timbul akibat pemberian takaran harian berulang dari pestisida atau pemaparan pestisida yang berlangsung cukup lama, biasanya lebih dari 50% rentang hidup (Runia, 2008).
yang dipergunakan untuk mematikan atau memberantas serangga. Berdasarkan
Pestisida organofosfat ditemukan melalui sebuah riset di Jerman, selama Perang Dunia II dalam usaha menemukan senjata kimia untuk tujuan perang. Pada tahun 1937, G. Schrader menyusun struktur dasar organofosfat (Runia, 2006). Organofosfat adalah derivat dari asam fosfat atau asam tiosulfat. Jenis pestisida ini mengandung unsur-unsur fosfat, karbon, oksigen serta hidrogen. Pestisida organofosfat terdiri dari satu atau lebih gugus fosfor yang terikat pada molekul organik. Organofosfat dibuat dari suatu molekul organik yang direaksikan dengan fosforilat (Sinulingga, 2006). Beberapa jenis pestisida organofosfat antara lain: diazinon, dibrom, di-syston, dylox, etion, gution, malation, paration, fosdrin, systox, TEPP, forate, trition, dan DIPF.
pirosulfat, metasulfat dan bentuk senyawa organik yang larut dalam air misalnya asam nukleat dan phitin. Fosfat mudah bergerak antar jaringan tanaman.
3. Malation
Malation Salah satu jenis pestisida organofosfat yang sering digunakan, karena relatif lebih aman dibandingkan dengan jenis organofosfat yang lain. Malation diperkenalkan pada tahun 1952. Malation merupakan pro-insektisida yang dalam proses metabolisme serangga akan diubah menjadi senyawa lain yang beracun bagi serangga. Insektisida ini bertindak sebagai racun kontak dan racun lambung, serta memiliki efek sebagai racun inhalasi. Malation juga digunakan dalam bidang kesehatan masyarakat untuk mengendalikan vektor penyakit. LD50 dari malation berdasarkan Mohajeri (2014) adalah 1200 mg/Kg. Stuktur dari malation ditunjukkan pada Gambar 1.
Gambar 1. Struktur malation
(O,O-dimethyl-S-(1,2-dicarbethoxyethyl)phosphorodithioate)
B. Enzim asetilkolinesterase (AChE)
Enzim asetilkolinesterase (AChE) merupakan enzim golongan hidrolase yang berperan untuk memecah atau mendegradasi asetilkolin (Ach) menjadi kolin dan asam asetat. Enzim ini dijumpai terutama dalam serum, hati, dan pankreas. Reaksi hidrolisis asetilkolin oleh enzim AChE terjadi di daerah sinapsis dan jaringan saraf pada vertebrata dan invertebrata. Enzim ini sangat diperlukan untuk menjamin kelangsungn fungsi sistem saraf manusia.
Asetilkolin adalah suatu neurotransmiter yang berfungsi menghantarkan impuls saraf pada saraf otonom (parasimpatik) dan dan saraf somatik (otot rangka). Reseptor dari asetilkolin adalah nikotinik dan muskarinik. Kelebihan asetilkolin akan menyebabkan perangsangan parasimpatik (perangsangan reseptor nikotinik dan muskarinik), sedangkan jika kekurangan asetilkolin akan menyebabkan depresi parasimpatik. Jadi kelebihan atau kekurangan Ach akan berbahaya (Zuraida, 2012).
Reaksi hidrolisis asetilkolin oleh enzim asetilkolinesterase (AChE) adalah sebagai berikut:
C. Fungsi mitokondria
mitokondria adalah untuk menghasilkan energi, mereka juga memainkan peran penting dalam metabolisme dan sintesis zat lainnya dalam tubuh (Sridianti, 2014).
Fungsi mitokondria bervariasi sesuai dengan jenis sel di mana mereka berada. Fungsi mitokondria antara lain:
1) menghasilkan energi melalui fosforilasi oksidatif;
2) menjaga konsentrasi ion kalsium yang tepat dalam berbagai kompartemen sel; 3) membantu dalam membangun bagian-bagian tertentu dari darah dan hormon
seperti testosteron dan estrogen;
4) mitokondria dalam sel-sel hati memiliki enzim yang dapat mendetoksifikasi amonia;
5) mitokondria juga memainkan peran penting dalam proses apoptosis (kematian sel yang terprogram).
Berdasarkan fungsi-fungsi mitokondria yang telah disebutkan, maka akan dibahas mengenai fungsi mitokondria sebagai penghasil energi dan peran mitokondria dalam apoptosis sel.
1. Proses fosforilasi oksidatif pada mitokondria atau mitochondria
respiratory chain (MRC)
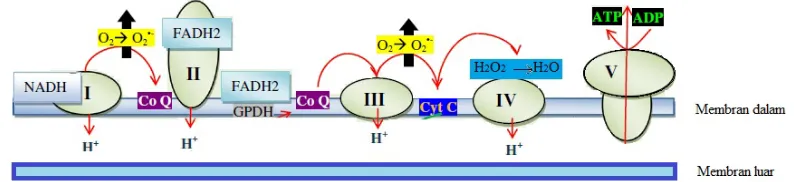
Mitokondria memiliki fungsi sebagai penghasil energi melalui fosforilasi oksidatif. Sumber energi yang dimetabolisme di sitoplasma ditransfer ke dalam mitokondria, kemudian mitokondria melanjutkan proses metabolisme dengan menggunakan jalur metabolik meliputi siklus Krebs, oksidasi asam lemak dan oksidasi asam amino. Hasil akhir dari jalur-jalur tersebut adalah produksi dua donor elektron yang kaya energi, yaitu: NADH dan FADH2. Elektron dari donor tersebut akan dilewatkan melalui rantai transport elektron menuju oksigen (O2), yang kemudian direduksi menjadi air. Hal ini merupakan proses redoks panjang atau mitochondria respiratory chain (MRC) yang terjadi di membran dalam mitokondria.
dari substrat ke oksigen. Protein tersebut merupakan enzim yang terbagi kedalam beberapa kompleks. Kompleks I (NADH-koenzim Q oksidoreduktase), kompleks II (suksinat-koenzim Q oksidoreduktase), kompleks III (koenzim Q sitokrom c reduktase), komplek IV (sitokrom c oksidase), dan kompleks V (ATP sintase). Transfer elektron dari NADH menuju O2 melibatkan membran dalam multi-subunit kompleks I, III, dan IV, koenzim Q (kompleks II) serta sitokrom c. Elektron melintasi pada setiap kompleks secara berurutan melalui beberapa elektron pembawa. Perpindahan elektron secara keseluruhan adalah sebagai berikut:
NADH →Kompleks I→Q→ Kompleks III→cytochrome c→ Kompleks IV→ O2 ↑ Kompleks II
Gambar 2. Proses pembentukan ATP berdasarkan teori chemiosmotic coupling pada matriks mitokondria.
2. Proses apoptosis sel
Apaptosis merupakan kematian sel yang terprogram, melalui proses kerusakan kromatin pada nukleus / inti sel. sel menyusut dengan pembentukan badan-badan apoptosom (apoptotic body) dan sel mengepak dirinya sendiri untuk dimakan makrofag. Apoptosis terjadi setiap hari dalam tubuh kita, sel dalam tubuh ada yang berproliferasi (lahir) dan ada yang mati. Untuk terjadi apoptosis ada berbagai macam stimulus, stimulusnya sangat terkontrol (bukan sesuatu yang asal lalu mati). Sentral pada proses apoptosis yaitu caspase, memiliki peran sebagai protein eksekutor yang memutuskan sel untuk apoptosis (Wane, 2011).
molekul caspase dapat mengaktifkan molekul caspase yang lainnya (snowball effect). Apoptosis dibagi 2 jalur utama yaitu : intrinsik (mitochondrial pathway) dan ekstrinsik (death receptor). Namun, pada pembahasan ini hanya ditekankan pada jalur intrinsik (Wane, 2011).
Jalur intrinsik (mitochondrial pathway apoptosis) intinya adalah pelepasan sitokrom c, karena begitu sitokrom c keluar maka akan diikat oleh Apaf-1 (apoptosis activating factor) yang akan membentuk apoptoseome. Apoptosome akan mengaktivasi caspase 9 (caspase awal yang diaktifkan oleh sitokrom c), sehingga caspase 9 akan mengaktivasi caspase selanjutnya (3, 6, dan 7) dan mengaktifkan sistem yang lain. Protein yang berperan dalam apoptosis yaitu BH1, BH2, BH3, dan BH4, artinya satu gen yang terdiri dari beberapa lobus. Bila yang diaktifkan adalah protein BH1, BH2, BH3, BH4 maka akan menjadi Bcl2/Bcl-XL yang merupakan anti-apoptosis (Bcl-2, Bcl-XL, Bcl-w, Mcl-1, A1), tetapi jika yang diaktifkan adalah protein BH1, BH2, BH3 (atau hanya BH3) maka akan menjadi pro-apoptosis (Bim, Puma, tBid, Bad, Noxa, Bax). Contoh dari apoptosis sel jalur intrinsik adalah selular stress yang diakibatkan oleh radiasi, infeksi virus, faktor pertumbuhan, dan stress oksidasif. Intinya, sinyal apoptosis melalui pintu mitokondria yang melibatkan protein bcl-2 dan bugs (p53), walaupun rangsangannya dari luar, tetapi hancurnya sel terjadi di mitokondria (Wane, 2011).
D. Pengaruh malation terhadap gangguan fungsi mitokondria
malation sebanyak 50, 100, 150, 200, 250, 300, 400, 500 dan 600 mg/Kg, sedangkan kelompok kontrol hanya diberi makan minyak jagung. Setelah 24 jam paparan malation, tikus tersebut dibius menggunakan katamine-xylazine dan diambil sampel darahnya, kemudian dipreparasi dan dilakukan analisis enzim. Hasil analisis ditunjukkan pada Gambar 3.
Gambar 3. Grafik hubungan dosis malation dengan penghambatan aktivitas enzim asetilkolinesterase (AChE).
Penghambatan malation terhadap enzim AChE bersifat irreversible (tidak dapat diperbaharui) kecuali dengan sintesis enzim asetilkolinesterase baru. Toksisitas malation sangat cepat, karena memiliki daya absorbsi yang baik serta afinitas yang tinggi terhadap AChE.
Asetilkolin (ACh) yang tidak dapat didegradasi oleh asetilkolinesterase (AChE) akan menumpuk pada sinapsis saraf, karena asetilkolin merupakan suatu senyawa neurotransmiter yang berfungsi di dalam bagian sinaps yang dihasilkan oleh ujung syaraf (akson) yang telah menerima impuls. Asetilkolin yang dihasilkan diteruskan ke sel syaraf lainnya atau ke efektor (misalnya otot) untuk meneruskan impuls syaraf (Suhara, 2009). Akan tetapi, sebelum impuls kedua dapat dipancarkan melalui sinaps, asetilkolin yang dihasilkan setelah impuls pertama harus dihidrolisis oleh asetilkolisnesterase (AChE) pada sambungan sel syaraf atau neuromuscular junction (NJM). Namun, karena AChE tidak dapat menghidrolisis asetilkolin, maka impuls kedua dan seterusnya tidak akan sampai ke efektor. Akumulsi asetilkolin pada neuromuscular junction (NJM) akan mengakibatkan kontraksi otot, yang diikuti dengan kelemahan, hilangnya reflex dan paralisis.
waktu antara stimulus dengan respon), dan tes kecepatan konduktivitas saraf atau NVC (nerve conduction velocity) untuk mendeteksi tanda-tanda cidera saraf.
Kelompok tikus yang digunakan untuk uji EMG adalah kelompok tikus yang mendapatkan paparan malthion sebanyak 400mg/Kg. Setelah 24 jam tikus dibius menggunakan ketamin-xylazin. Bagian yang diuji adalah otot kaki dan otot gastrosel (otot lambung). Hasil pengujian elektromiokgrafi (EMG) dan NVC ditunjukan pada Tabel 1.
Tabel 1. Perbandingan statistik dari amplitudo CMAP, durasi CMAP, latensi,dan NVC terhadap stimulasi respon dari saraf sciatik dari otot gastrosel tikus pada kelompok kontrol dan kelompok perlakuan.
amplitudo CMAP (mV) 20,040 ± 1,293 12,260 ± 1,570
Durasi CMAP (ms) 13,330 ± 0,500 9,905 ± 0,650
Eek akut dari malation menyebabkan kelemahan otot, akibatnya otot membutuhkan ATP yang sangat banyak, karena ATP merupakan sumber energi yang digunakan untuk menggerakkan otot. Mitokondria merupakan organel utama penghasil ATP, oleh karena itu untuk mengetahui pengaruh malation terhadap fungsi mitokondria dapat dilihat melalui pengukuran terhadap enzim-enzim yang terlibat dalam proses transfer elektron di mitokondria atau mitochondria respiratory chain (MRC).
2. Interaksi malation dengan enzim-enzim yang terlibat dalam mitochondria
respiratory chain (MRC)
Penelitian yang dilakukan terhadap aktivitas enzim-enzim yang terdapat pada komplek mitokondria, yaitu: kompleks I (NADH-koenzim Q oksidoreduktase), kompleks II (suksinat-koenzim Q oksidoreduktase), kompleks IV (sitokrom c oksidase), dan kompleks V (ATP i). Sampel pengujian diambil dari otot gastrosel tikus yang mendapatkan paparan malation sebanyak 400mg/Kg. Otot gastrosel dipilih karena berdasarkan penelitian yang dilakukan oleh Thompson, dkk (1998), konsentrasi malation paling banyak ditemukan di otot gastosel. Hasil pengujian aktivitas enzim tersebut, ditunjukan pada Tabel 2.
Tabel 2. Efek akut paparan malation terhadap aktivitas enzim pada kompleks I, II, IV dan V yang diukur dari otot gastrosel tikus pada kelompok kontrol dan kelompok perlakuan.
Aktivitas enzim Kelompok kontrol Kelompok perlakuan NADH-oksidoreduktase (mM/
min/mg) 0,010 ± 0,001 0,006 ± 0,0006
Suksinat-oksidoreduktase (mM/
min/mg) 0,170 ± 0,067 0,090 ± 0,010
Sitokrom c oksidase (K/min/mg) 3,136 ± 0,287 1,657 ± 0,275 ATP sintase (mM/min/mg) 0,103 ± 0,015 0,064 ± 0,009
penurunan dibandingkan dengan kelompok kontrol. Hal tersebut membuktikan bahwa malation berpengaruh pada enzim-enzim yang berkaitan dengan mitochondria respiratory chain (MRC).
Metabolisme aerobik glukosa digabungkan dengan fosforilasi oksidatif (transfer elektron) oleh FADH2 pada kompleks II melalui coupling reaksi suksinat menjadi fumarat dalam siklus asam sitrat. Oleh karena itu, ketika terjadi penurunan terhadap aktivitas enzim-enzim MRC dapat mengakibatkan penurunan produksi ATP. Pengukuran rasio ATP dilakukan melalui teknik high-performance liquid chromatography (HPLC). Hasil pengukuran ditunjukkan melalui data rasio ADP/ATP pada Gambar 4.
Gambar 4. Grafik perubahan rasio ADP/ATP pada otot gastrosel tikus dari kelompok kontrol dan perlakuan.
Gambar 5. Grafik perubahan kadar glikogen pada otot gastrosel tikus dari kelompok kontrol dan kelompok perlakuan.
Selain mengurangi kadar glikogen dalam otot, dalam kondisi hipoksia (kekurangan oksigen), memicu terjadinya metabolisme anaerob dari glukosa. Hasil akhir dari metabolisme anaerob glukosa adalah asam laktat sehingga konsentrasi rata-rata asam laktat dalam otot tikus yang mendapatkan paparan malation selama 24 jam, secara signifikan lebih tinggi dibandingkan dengan kelompok kontrol, ditunjukkan pada Gambar 6.
Gambar 6. Grafik perubahan konsentrasi asam laktat pada otot gastrosel tikus dari kelompok kontrol dan kelompok perlakuan.
asetil KoA. Jalur metabolisme karbohidrat secara ringkas ditunjukkan pada Gambar 7.
Gambar 7. Jalur perubahan glukosa.
Kondisi hipoksia (kekurangan oksigen) memicu terjadinya iskemia. Iskemia adalah suatu keadaan kekurangan oksigen yang bersifat sementara dan reversibel. Penurunan suplai oksigen akan meningkatkan mekanisme metabolisme anaerobik. Metabolisme anaerobik sangat tidak efektif selain energi yang dihasilkan tidak cukup besar, juga meningkatkan pembentukan asam laktat. Iskemia dapat menyebabkan nyeri sebagai akibat penimbunan asam laktat yang berlebihan. Iskemia yang lama dapat menyebabkan kematian otot atau nekrosis. Keadaan nekrosis yang berlanjut dapat menyebabkan kematian otot jantung (infark miokard) (Word health, 2012).
mengganggu metabolisme karbohidrat, lipid dan protein. Menurut Wijaya (dalam Maslachah, 2008) ROS merupakan oksidan yang sangat reaktif dan mempunyai aktivitas yang berbeda. Dampak negatif senyawa tersebut timbul karena aktivitasnya, sehingga dapat merusak komponen sel yang sangat penting untuk mempertahankan integritas sel. Setiap ROS yang terbentuk dapat memulai suatu reaksi berantai yang terus berlanjut sampai ROS itu dihilangkan oleh ROS yang lain atau sistem antioksidannya.
Mitokondria adalah konsumen utama molekul oksigen di dalam sel. Selama metabolism oksidasi di mitokondria termasuk MRC, sebagian besar oksigen dirubah menjadi air, tetapi ada 3% dari seluruh oksigen yang dikonsumsi dirubah dalam bentuk superoksida. Reduksi oksigen oleh satu elektron menghasilkan ROS (contohnya O2-•, H
2O2 , •OH). Produksi ROS pada mitokondria diukur melalui emisi H2O2, karena O2-• akan dirubah oleh enzim mangan-superoxide dismutase (MnSOD) menjadi H2O2.Hasil penelitian menunjukan bahwa pembentukan H2O2 pada otot gastrosel intak mitokondria tikus dari kelompok perlakuan mengalami penurunan dibandingkan dengan kelompok kontrol, ditunjukkan pada Gambar 8.
Gambar 8. Grafik pembentukan H2O2 pada otot gastrosel intak mitokondria dari tikus kelompok kontrol dan kelompok perlakuan.
glutation peroksidase superoksida dismutase
Mohajeri.S.K dan Abdollahi.M (2013) tingginya oksisidan di dalam mitokondria harus diimbangi dengan pertahanan (antioksidan) agar dapat mengurangi kerusakan oksidatif. Berdasarkan hal tersebut, penurunan kadar H2O2 dari tikus kelompok perlakuan menunjukakkan bahwa adanya malation dapat meningkatkan level dari glutation.
Mekanisme pembtukan ROS pada kompleks mitokondria ditunjukkan pada Gambar 9.
Gambar 9. Mekanisme pembentukan ROS (O2.) pada kompleks I dan III mitokondria
Setelah terbentuk superoksida (O2.) pada kompleks I dan III, maka superoksida tersebut akan dirubah menjadi H2O2 oleh enzim superoksida dismutase (SOD) dan kemudian H2O2 didegradasi menjadi O2 dan H2O oleh enzim glutation peroksidase. Reaksi yang terjadi adalah sebagai berikut:
O2. H2O2 O2 + H2O
3. Pengaruh malation terhadap apoptosis sel
ATGTCA-3’. Primer untuk gen bcl-2 yaitu: 5’-CTGCATCTCATGCCAAGGGG-3’ dan ‘5-ACCATAGCACTTTTTCGC GTCC-5’-CTGCATCTCATGCCAAGGGG-3’. Tingkat ekspresi gen bax dan bcl-2 untuk tiga sampel di setiap kelompok (kontrol dan perlakuan) dinormalisasi dengan membandingkan ambang siklus/ cycle treshold (CT) dari β-aktin sebagai housekeeping gen. β-aktin merupakan protein yang selalu diekspresikan dalam semua keadaan, sehingga bisa dijadikan standar untuk melihat perubahan terhadap ekspresi gen tertentu. Berikut ini adalah primer untuk β-aktin: ‘5-TCCTGTGGCATCCACGAAACT-3’ dan ‘5-ACTTGCGCTCAGGAGGAGCA A-3’. Hasil analisi terhadap ekspresi gen bax dan bcl-2 ditunjukkan pada Tabel 3.
Tabel 3. Perbedaan lipatan eksprsi mRNA dari gen bax dan genbcl-2 dengan β-aktin sebagai housekeeping pada otot gastosel tikus dari kelompok
kontrol dan kelompok perlakuan.
Kelompok kontrol Kelompok perlakuan
Gen ΔCT 2β-ΔCT ΔCT 2-ΔCT
Bax 2,357 ± 0,072 0,195 ± 0,010 5,093 ± 0,072 0,029 ± 0,001
Bcl-2 -5,740 ± 0,017 53,450 ± 0,642 -5,357 ± 0,037 41,000 ± 1,066
Dari hasil analisis, dapat dilihat bahwa terjadi penurunan terhadap ekspresi gen bax dari kelompok kontrol (0,195 ± 0,010), sedangkan kelompok perlakuan (0,029 ± 0,001). Begitupun dengan ekspresi dari gen bcl-2, untuk kelompok kontrol (53,450 ± 0,642) dan kelompok perlakuan (41,000 ± 1,066). Hasil tersebut menunjukkan bahwa ada pengaruh malation terhadap ekspresi dari gen bax dan bcl-2. Selain menganalisis ekspresi dari kedua gen tersebut, dilakukan juga analisi terhadap pelepasan sitokrom c dari mitokondria.
yang berada di sitosol membantuk kompleks dengan Apaf-1, ATP, dan caspase 9 dinamakan apoptosom (suatu holoenzim, gabungan beberapa protein). Kompleks ini adalah suatu protease yang bertugas memotong/degradasi protein lain. (CCRC, 2012). Hasil ditunjukan pada Gambar 10.
Gambar 10. Grafik pelepasan sitrokrom c dari mitokondria pada tikus kelompok kontrol dan kelompok perlakuan.
Berdasarkan hasil penelitian, ditunjukkan bahwa pelepasan sitokrom c dari mitokondria pada tikus kelompok perlakuan secara signifikan lebih rendah dibandingkan dengan kelompok kontrol. Keluarnya sitokrom c dari mitokondria akan diikat oleh Apaf-1 (apoptosis activating factor) yang akan membentuk apoptoseome. Apoptosome akan mengaktivasi caspase 9 (caspase awal yang diaktifkan). Caspase 9 yang aktif disebut (cleaved caspase 9), sedangkan yang tidak aktif disebut (non cleaved caspase 9).
Gambar 11. Analisis western blot pada otot gastrosel dari kelompok kontrol dan kelompok perlakuan.
(a) Visualisasi protein β-aktin, serta protein cleaved dan non cleaved caspase-9 melalui analisis SDS-PAGE.
(b) Perbandingan antara protein cleaved dan non cleaved caspase-9, dibandingkan dengan protein β-aktin.
Hasil yang diperoleh, tidak ada perubahan yang signifikan dari ekspresi protein non cleaved dan cleaved caspase-9 mitokondria. Berarti, pengaruh malation tidak sampai mempengaruhi ekspresi dari caspase 9. Namun, adanya malation mengakibatkan penumpukan asetil kolin pada reseptor nikotinik dan muskarinik memberikan survival signaling pada gen bax yang mengakibatkan gangguan terhadap keluarnya sitokrom c dari mitokondria. Jika sitokrom c tidak dapat keluar dan tidak diikat oleh Apaf 1, maka tidak akan terjadi proses apoptosis. Artinya tidak akan terjadi regenerasi sel, sel yang sudah mati tidak dapat dirilis.
elektrofilik yang dapat bereaksi dengan atom O dan N dari basa nitrogen adenine, sitosin dan guanine dari DNA. Genotoksisitas dan kerusakan DNA pada petani yang terekspos OP pernah diberitakan (Mphajeri, 2014). Salah satu indikator kerusakan DNA akibat ionisasi radiasi, mutagen atau karsinogen adalah pembentukan senyawa 8-hidroksi-2deoksiguanosin (8-OHdG) sebagai hasil oksidasi terhadap nukleosida DNA. Struktur dari senyawa 8-OHdG ditunjukkan pada Gambar 12.
Gambar 12. Struktur 8-hidroksi deoksiguanosin (8-OHdG)
Untuk mengetahui pengaruh genotoksisitas tersebut, maka diuji juga kadar senyawa 8-OHdG pada tikus. Hasil yang diperoleh (Gambar 13), ditunjukkan bahwa pada tikus yang mendapat paparan malation, memiliki kadar senyawa 8-OHdG yang lebih banyak dibandingkan dengan kelompok kontol.
Pengaruh malation terhadap gangguan fungsi mitokondria dapat dijelaskan melalui Gambar 14.
Gambar 14. Model sederhana mekanisme potensial seluler berkaitan dengan efek Malthion terhadap aktivitas enzim AChE dan gangguan fungsi
mitokondria.
Pada tahap awal keracunan, malation yang berikatan dengan enzim asetilkolinesterasi (AChE) menyebabkan penumpukan asetilkolin (Ach) pada neuromuscular junction, tepatnya pada reseptor nikotinik dan muskarinik, akibatknya transmisi impuls saraf terhambat sehingga timbul gejala berupa efek nikotinik dan muskarinik. Akumulsi pada neuromuscular junction (NJM) akan mengakibatkan kontraksi otot, yang diikuti dengan kelemahan, hilangnya reflex dan paralisis.
Paparan malation menyebabkan penurunan terhadap aktivitas enzim-enzim yang berhubungan dengan rantai transport elektron (ETC electron transfer chain) yang berkaitan dengan aktivitas kompleks I dan II berhubungan dengan stres oksidatif, sedangkan aktivitas kompleks IV dan V menyebabkan runtuhnya potensial membran mitokondria dan deplesi ATP. Oleh karena itu, penurunan ROS mitokondria serta kompleks IV dan V dapat menurunan produksi ATP dan sinyal apoptosis.
III. RANGKUMAN
Malation merupakan salah satu jenis insektisida organofosfat yang digunakan untuk membunuh serangga. Dalam proses metabolisme serangga, malation akan diubah menjadi senyawa lain yang beracun bagi serangga. Dosis toksik dari malation dalam kasus keracunan pada manusia adalah 350-1300 mg/kg. AChE merupakan enzim yang berfungsi untuk menghidrolisis Asetilkolin (ACh) menjadi asam asetat dan kolin. Malation menyebabkan penurunan jumlah AChE, karena AChE akan berikatan dngan malation sehingga asetilkolin (Ach) tidak dapat dihidrolisis. Adanya penumukan asetilkolin (AChE) pada sistem saraf dan neuromuscular junction. Akumulsi pada neuromuscular junction akan mengakibatkan kontraksi otot yang diikuti dengan kelemahan, hilangnya reflex dan paralisis.
DAFTAR PUSTAKA
Badan Penelitian dan Pengembangan Kesehatan Kementrian Kesehatan RI. (2013). Riset Kesehatan Dasar (RIKESDAS) 2013. Jakarta: Kementrian Kesehatan RI. Bellinda, R. R. (2008). Mutasi DNA Mitokondria Manusia yang Berkaitan dengan
Diabetes Militus. (Skripsi). Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Teknologi, Bandung.
CCRC. (2012). Pertanan Mtokondria dalam Apoptosis. [Onine]. Diakses dari
http://ccrc.farmasi.ugm.ac.id/?p=2551.
Data Statistik Indonesia. (2014). Proyeksi Penduduk 2000-2025. [Onine]. Diakses
dari
http://www.datastatistik-indonesia.com/proyeksi/index.php?option=com-proyeksi&task= show&Itemid=941.
Maslachah, L. et al.(2008).“Hambatan Produksi Reactive Oxygen Species Radikal Superoksida (O2.-) oleh Antioksidan Vitamin E (α- tocopherol ) pada Tikus Putih (Rattus norvegicus) yang Menerima Stressor Renjatan Listrik”. Jurnal: Media Kedokteran Hewan, 1(24), hlm. 21-26.
Mohajeri. et al. (2014). “Mechanisms of muscular electrophysiological and mitochondrial dysfunction following exposure to malation, an organophosphorus pesticide”. Journal: Journal of Hum Exp Toxicol, 3(33), hlm. 251–263.
Mohajeri, S.K, and Abdollahi, M. (2011). “Toxic influence of organophosphate, carbamate, and organochlorine pesticides on cellular metabolism of lipids, proteins, and carbohydrates: A systematic review”. Journal: Journal of Hum Exp Toxicol, 9(30), hlm. 1119–1140.
Mohajeri, S.K, and Abdollahi, M. (2013). “Mitochondrial adysfunction
and organophosphorus compounds”. Journal: Journal of
Toxicology and Applied Pharmacology, (270). Hlm. 39–44.Kesehatan Lingkungan, Program Pasca Sarjana, Universitas Diponogoro, Semarang.
Russell, R.W, and Overstreet, D.H. (1987). Mechanism Underlying Sensitivity to Organophosphorus Anticholinesterase Compound. Journal: Journal of Progress in Neurobiology, (28), hlm. 97-129.
Poedjiadi, A, dan Titin, F. M. (2007). Dasar-dasar Biokimia. Jakarta: UI Press.
SK Menteri Pertanian RI No. 434.1/Kpts/TP.270/7/2001.
Sinulingga, D. (2006). Pengaruh Tempe Terhadap Peningkatan Aktivitas Enzim Asetilkolinesterase pada Petani Penyemprot Terpapar Pestisida Organofosfat. (Disertasi). Sekolah Pasca Sarjana, Universitas Sumatra Utara, Medan.
Siwiendrayanti, A. et al. (2012). “Hubungan Riwayat Pajanan Pestisida Dengan Kejadian Gangguan Fungsi Hati (Studi pada Wanita Usia Subur di Kecamatan Kersana Kabupaten Brebes)”. Jurnal: Jurnal Kesehatan Lingkungan Indonesia,
1(11), hml. 9-14.
Sridianti. (2014). Pengertian Mitokondria dan Bagian-bagian Mitokondria. [Onine]. Diakases dari http://www.sridianti.com/struktur-fungsi-mitokondria.html .
Suhara. (2009). Dasar dasar biokimia. Bandung: Prisma Press.
Sukmawati, A dan Maharani, A. (2004). “Hubungan Antara Perilaku Dalam Pengolahan Pestisida Dengan Aktivitas Enzim Cholinesterase Darah Pada Petani Cabe Di Desa Santan Mekar Kecamatan Cisayong Kabupaten Tasikmalaya”. Jurnal: Jurnal Ekologi Kesehatan, 2 (3), hlm. 80-89.
Thompson. et al. (1998). Case study: Fatal Poisoning by Malathion. Journal: Journal of Forensic Science International, (95), hlm. 89–98.
Voet, D. V. J. (1995). Biochemistry. New York: 2nd Ed.John Wiley & Sons Inc.
Wane. (2011). Pengertian Apoptosis dan Proses Apoptosis. [Onine]. Diakses dari