UNIVERSIDAD DE COSTA RICA
SISTEMA DE ESTUDIOS DE POSGRADO
VARIACIÓN GENÉTICA ASOCIADA CON
INTOLERANCIA A LA LACTOSA EN GRUPOS
INDÍGENAS DE BAJA CENTROAMÉRICA Y SU
POTENCIAL COMO PROBLEMA DE SALUD
Tesis sometida a la consideración de la Comisión del
Programa de Estudios de Posgrado en
Antropología para optar al grado y título de Maestría
Académica en Antropología
ANA YANCY MORALES ARCE
DEDICATORIA
A mis seres queridos.
AGRADECIMIENTOS
A los pueblos indígenas de Costa Rica y Panamá.
Al Dr. Jorge Azofeifa por su invaluable guía y sus constantes consejos.
Al Dr. Ramiro Barrantes y a la Dra. Silvia Salgado por su interés en el tema y su colaboración.
A la Universidad de Costa Rica, por el apoyo logístico, financiero e institucional por medio del proyecto “Variación genética y evolución de la región del gen de la lactasa LCT en la población de Costa Rica” No 111-A8-069 y el programa “Ecología, diversidad genética y salud de los amerindios de CR” No 742-93-903.
A aquellas personas que estuvieron en algún momento durante el proceso de laboratorio y mostraron interés y colaboración con el trabajo.
A mis amigos y familiares por su paciencia.
HOJA DE APROBACIÓN
“Esta tesis fue aceptada por la Comisión del Programa de Estudios de Posgrado en Antropología de la Universidad de Costa Rica, como requisito parcial para optar al grado y título Maestría
Académica en Antropología”
____________________________________
Eric Fuchs Castillo, Ph.D Representante de la Decana Sistema de Estudios de Posgrado
______________________________________ Jorge Azofeifa Navas, Dr.sc.hum.
Director de Tesis
______________________________________ Ramiro Barrantes Mesén, D.Sc.
Asesor
_______________________________________ Silvia Salgado González, Ph.D.
Asesora
_______________________________________ Dra. María del Carmen Araya Jiménez
Directora
Programa de Posgrado en Antropología
_______________________________________
Ana Yancy Morales Arce
TABLA DE CONTENIDOS
1.1 Características de la lactosa y su relación con la lactasa ... 1
1.2 Lactasa e intolerancia a la lactosa ... 2
1.3 Desarrollo histórico de la dilucidación de las bases genético-moleculares de la intolerancia a la lactosa ... 5
1.4 Distribución de la capacidad para digerir la lactosa en diferentes poblaciones del mundo ... 7
2. Objetivos ... 10
2.1 Objetivo General ... 10
2.2 Objetivos específicos ... 10
3. Marco Teórico ... 11
3.1 Desarrollo histórico de las hipótesis que explican el fenómeno de intolerancia a la lactosa. ... 11
4. Metodología ... 17
4.1 Poblaciones de estudio ... 17
4.1.1 Huetar ... 17
4.2 Encuesta sobre la percepción de los efectos del consumo de leche en dos comunidades indígenas. ... 19
7.1 Referencias de internet ... 57
RESUMEN
La documentación más antigua sobre el consumo de leche por adultos data desde hace unos 7000 años en la región de Libia. La práctica del consumo fue posible entre las poblaciones que domesticaron rumiantes entre ellas las de medio oriente, algunas africanas y europeas; que siglos más tarde se incorporó en otras regiones del mundo. Los cambios industriales globales relacionados con la comercialización de la leche y sus derivados se intensificaron desde 1960 dando pie a un incremento en su producción y consumo global. La disponibilidad y diversidad de productos lácteos, y en particular de leche fresca han puesto en evidencia que existen diferencias entre poblaciones e individuos para digerir dichos alimentos, debido principalmente a uno de los componentes de la leche: la lactosa, y se ha estimado que afecta a la mayoría de la población mundial. La ingestión de leche o productos lácteos no deslactosados, puede provocar intolerancia a lactosa que produce síntomas como distensión abdominal, cólicos, flatulencia, diarrea, nauseas y vómito. A nivel de ADN se han reportado marcadores genéticos (SNPs) corriente arriba del gen de la lactasa que se asocian con los fenotipos tolerancia/intolerancia a lactosa en determinadas poblaciones, entre ellas, europeas (T -13910 y A -22018) y africanas (C -14010). El objetivo de este trabajo fue estimar la variación genética asociada con intolerancia a la lactosa en grupos amerindios de Baja Centroamérica. Se utilizaron 214 muestras de ADN de las poblaciones amerindias repartidas entre los grupos Maleku, Ngöbe, Huetar, Bribri, Cabécar, Chorotega y Rama. Se amplificaron las regiones que contienen los SNPs y se obtuvieron los genotipos por medio de electroforesis capilar y enzimas de restricción. Se obtuvo una variación entre 0.01 y 0.99 en los diferentes SNPs a excepción de los SNPs 4 (C -14010) y 7 (A -22018) que mostraron fijación. Las mayores frecuencias de alelos europeos se encontraron en la población huetar mientras que la mayor frecuencia del alelo africano se encontró en las poblaciones de Rama Cay (rama) y Limoncito (ngöbe). Se estimaron 55, de los cuales el 2 y el 4 alcanzaron las mayores frecuencias. Los haplotipos asociados con tolerancia en Europa alcanzaron solamente un 3% en la población maleku y un 10% en la población huetar de Quitirrisí. El valor de Fst (9.7%)
ABSTRACT
LISTA DE CUADROS
Cuadro 1. SNP´s analizados en las muestras de ADN para todas las poblaciones. ... 21 Cuadro 2. Frecuencias alélicas de 9 loci (SNP) analizados en 10 poblaciones indígenas. Los loci que se encuentran en desequilibrio Hardy-Weinberg se muestran sombreados..25 Cuadro 3. Diversidad génica (Ĥ) por locus y por población y heterocigosis promedio (desviación estándar) para 10 poblaciones indígenas.. ... 26
Cuadro 4. Frecuencias haplotípicas absolutas (relativas) en 10 poblaciones indígenas. 27 Cuadro 5. Diversidad haplotípica (Ĥh) y Fst por población y total. ... 29
Cuadro 6. Prueba de neutralidad Ewens-Waterson para 10 poblaciones indígenas. ... 31 Cuadro 7. Frecuencias genotípicas conformados por la combinación de los SNP`s 3 (C/T-13910) y 7 (G/A-22018) asociados en los grupos europeos con el polimorfismo tolerancia/intolerancia ………..……….……….32
LISTA DE FIGURAS
1. Introducción
La documentación más antigua sobre el consumo de leche por adultos data desde hace unos 7000 años en la región de Libia (Davidson, 1999); dicho consumo fue posible solamente entre las poblaciones que domesticaron rumiantes. Esta práctica permaneció en poblaciones de medio oriente, algunas africanas y europeas hasta que fue incorporada en diferentes regiones del mundo como Asia alrededor de 3 200 años atrás y en el nuevo mundo por los conquistadores quinientos años atrás (Simoons, 1971;Wiley, 2008). Siglos más tarde la lechería se industrializó y su consumo se oficializó en políticas de ayuda alimentaria primero en los Estados Unidos y posteriormente al resto de países americanos (USDA, 2002). Los cambios industriales globales relacionados con la comercialización de la leche y sus derivados se intensificaron desde 1960 dando pie a un incremento en su producción y consumo global (IDOE, 2010; Wiley, 2008). Desde entonces la disponibilidad y diversidad de productos lácteos, y en particular de leche fresca han puesto en evidencia que existen diferencias entre poblaciones e individuos para digerir dichos alimentos debido principalmente a uno de los componentes de la leche: la lactosa (Harrison, 1975). Esta complicación asociada con el consumo de leche y sus derivados es conocida como intolerancia a la lactosa, hipolactasia de los adultos, o restricción de la lactasa, (MIM 223100) y se ha estimado que afecta a la mayoría de la población mundial (Johnson et al. 1981; Bloom & Sherman, 2005).
1.1 Características de la lactosa y su relación con la lactasa
La lactosa (4-0-ß-D-galactopyranosyl-D-glucopyranose / 4-β-galactosilglucosa), es un disacárido producto de un enlace β-1,4-glicosídico entre la glucosa y la galactosa que se sintetiza en el aparato de Golgi de las glándulas mamarias a partir de glucosa y UDP-galactosa en una reacción unidireccional catalizada por la enzima lactosa sintasa (E.C. 2.4.1.22 / UDP-galactosa:D-glucosa 4β-D-galactotransferasa) (Keller et al. 1993).
del volumen en la leche humana, 5.1% en bovinos, 4.6% en caprinos hasta porcentajes menores, tales como en canguro (cantidades traza), oso polar (0.5%), conejo (1.8%), ballena (1.8%), venado (2.6%) (Jensen, 1995). La lactosa es el principal osmoregulador en la leche ya que a través de su síntesis se introduce agua en ella, obteniendo así su valor hidratante. Sin embargo, el papel preponderante de la lactosa es ser fuente de energía (Hurley, 2006).
En humanos, la lactosa proporciona al menos la mitad de la energía calórica durante los primeros dos años de vida, a partir de ese momento, entre los 2 y los 20 años la digestión de la lactosa disminuye hasta cantidades muy bajas y hasta nulas en la mayoría de las personas (Holden & Mace, 1997; Aoki, 2001; Swallow, 2003). Este hecho se atribuye al proceso natural (Troelsen et al. 1994, Rings et al. 1992) de disminución gradual de la producción de lactasa después de la infancia, proceso que ocurre en todos los mamíferos, y que puede interpretarse en términos evolutivos como un evento que provoca al infante un rechazo por la leche materna, llevando a la madre a recuperar su fecundidad al dejar de amamantar (Trivers, 1974). Esto a su vez podría explicar la variación en cuanto al contenido de lactosa en la leche de diferentes especies pues optimiza el tiempo y la tasa de destete de cada una de ellas (Liberman, 1978).
Por otra parte la leche contiene, además de la lactosa, nutrientes tales como calcio, vitaminas A y D, riboflavina y fósforo (Inman-Felton, 1998), nutrientes que son necesarios también después de la infancia. En occidente se ha promovido a la leche como una de las principales fuentes de calcio. Es por esto que se recomienda el consumo de leche y sus derivados en la adultez para prevenir enfermedades como la osteoporosis, hipertensión y ciertos tipos de cáncer (Inman-Felton, 1998; Yates et al. 1998; Peña et al. 2002).
1.2 Lactasa e intolerancia a la lactosa
alactasia congénita (MIM 223000)1 y la hipolactasia del tipo adulto (MIM 223100), mientras que las secundarias son, por lo general, transitorias, producidas por causas ambientales, como infecciones intestinales (Enattah, 2005). La principal causa de la intolerancia es la deficiencia de la enzima lactasa o lactasa-florizina hidrolasa (β -d-Galactosidasa- galactohidrolasa /EC 3.2.1. 23) que se clasifica dentro del grupo de las betagalactosidasas.
La digestión de la lactosa se da principalmente en la región yeyunal del intestino delgado (Holden & Mace, 1997). La lactasa presente hidroliza la lactosa en sus monosacáridos constituyentes, glucosa y galactosa, los cuales son absorbidos por las células de la membrana del intestino hacia el torrente sanguíneo (Kretchmer, 1972). En las personas donde está ausente la capacidad de digerir lactosa, la ingestión de una pequeña cantidad de leche (que puede variar entre 12 a 18 gramos o de 8 a 12 onzas de leche) (Swagerty et al. 2002) causa molestias ya que cuando los niveles de lactasa son insuficientes para poder descomponer la cantidad de lactosa consumida. Esta lactosa pasa intacta al colon, donde es parcialmente digerida por coliformes; el resultado es una acumulación de agua (debido la carga osmótica de la lactosa no hidrolizada) y desechos de bacterias (CO2 y ácidos grasos) que originan fermentación, produciéndose así
síntomas como son, distensión abdominal, cólicos, flatulencia, diarrea, nauseas y vomito (Fig 1) (Rosensweig, 1969); incluso pueden presentarse síntomas más severos (Wiley, 2008). La presencia de estos síntomas no sólo conllevan las molestias obvias sino que contribuyen a una mala digestión de otros nutrientes, especialmente de minerales en el tracto gastrointestinal (Phillips, 1981).
Existe una variación individual en la manifestación de los síntomas que depende de la cantidad de productos lácteos consumidos y de la sensibilidad individual a los dolores estomacales (Enattah, 2002). Sin embargo los niveles reducidos de lactasa no siempre causan síntomas incluso cuando se consume lactosa (De Vrese et al. 2001). Es por ello que la intolerancia no debe ser confundida con malabsorción, pues la intolerancia se
1 MIM son las iniciales de Mendelian Inheritance in Man, un catálogo de todas las características humanas
presenta a pesar del consumo continuado de productos lácteos, y la malabsorción sólo se presenta después de mucho tiempo de no consumir productos lácteos; lo que al reanudarse puede provocar síntomas de intolerancia a la lactosa pero que disminuyen con el tiempo, al producirse nuevamente una adaptación bacteriana al consumo, lo que es independiente de la actividad de la lactasa (Ransome-Kuti, 1977; Johnson et al 1981).
Figura 1. Representación de la absorción (panel A) y malabsorción (panel B) de la lactosa en el yeyuno.En el panel A se encuentra lactasa para romper el enlace (hidrólisis) entre glucosa y galactosa los cuales conforman la lactosa. En el panel B los bajos niveles de actividad de la lactasa permite el paso de lactosa hacia el colon donde es fermentada por bacterias (Zihlman, 1982 modificado por Durham, 1991).
1.2.1 Diagnóstico de intolerancia a la lactosa
Si bien es cierto la determinación a nivel clínico de la intolerancia a la lactosa es difícil, se han tratado de desarrollar algunos métodos. El método más directo involucra la toma de una biopsia del intestino y se ha realizado en poblaciones hospitalizadas (Dunphy et
al. 1965; Enattah et al. 2002; 2005). Otra de las pruebas requiere de la medición de
niveles de glucosa en la sangre luego de una a dos horas (en intervalos de 15 a 20 minutos) de la ingestión de 50 gramos de lactosa en una solución al 12. 5%, lo que equivale a un cuarto de litro de leche. Así, un aumento de menos de 20 mg de glucosa por cada 100 ml de sangre es considerado un indicador de intolerancia a la lactosa (Hoffman, 1937; Dubowsky, 1962). Algunas modificaciones se han hecho a los métodos de mediciones sanguíneas, por ejemplo, administrando previamente etanol (Jussila, 1969) o por medio de lactosa marcada con isótopos radiactivos (Laws et al. 1967). Sin embargo, han surgido controversias sobre la confiabilidad de estos métodos para el diagnóstico (Cuatrecasas et al. 1965; McGill & Newcomer, 1966; McMichael et al. 1966) además de ser poco prácticos (Harrison, 1975). Es por esto que se trataron de encontrar técnicas clínicas indirectas, que causaran los inconvenientes mínimos; una de ellas está basada en pruebas de orina que revelen una reducción de azúcares (Cook & Howells, 1968) y la otra en un análisis de carbono o hidrógeno expirado a intervalos regulares (Calloway & Murphy, 1968). Aún así, los métodos de medición directa ofrecen mejores resultados y son utilizados actualmente por presentar mayores índices de confiabilidad (Pimentel et al. 2004). Lo más importante y que es subyacente a todos estos métodos de diagnóstico es que requieren de facilidades clínicas, personal médico y pacientes concientes de las molestias que podrían tener durante las pruebas (Harrison, 1975).
1.3 Desarrollo histórico de la dilucidación de las bases genético-moleculares de la
intolerancia a la lactosa
ser digerida por 30 de 41 (73 %) adolescentes y adultos negros en Baltimore (Maryland) en contraste con una alta frecuencia de digestión “normal” en un grupos de blancos (16 de 19, 84%). Al profundizar las investigaciones Bayless y Rosensweig (1967) determinaron que la hipolactasia en los adultos es un rasgo determinado genéticamente y Kretchmer (1971, 1972) mostró que la hipolactasia es recesiva.
En 1988 Kruse et al. encontraron evidencia de que el gene de la lactasa (LPH) se encuentra en el brazo corto del cromosoma 2 (2q) y en 1993 Harvey et al. refinaron su ubicación a la banda 2q21, mientras que Boll et al. (1991) no encontraron diferencias en la estructura del gen, a nivel de ADN, ni en la de la proteína que producen los individuos con niveles altos o bajos de lactasa. Esto es válido también para la alactasia congénita (Järvelä et al. 1998). En ambos tipos de intolerancias primarias se ha encontrado que la causa genética de los problemas se encuentra en regiones vecinas del gen de la LPH pero separadas de él (Järvelä et al. 1998; Enattah et al. 2002).
Estudios de la biología molecular del gen (Ennatah et al. 2002) que codifica a la enzima lactasa han mostrado que el problema se origina en la reducción de la expresión del gen a proporciones que varían entre el 2% y el 10% con respecto a aquellos de la edad lactante y que se conoce como no persistencia de la lactasa, mientras que en las personas que no sufren el padecimiento, en quienes los niveles de expresión génica casi no cambian durante el desarrollo, se le denomina persistencia de la lactasa en adultos. Estos correlacionan directamente con los fenotipos de intolerancia/ tolerancia a la lactosa (Enattah, 2005).
A nivel de ADN Enattah et al. (2002) reportaron para poblaciones europeas dos SNPs (el término SNPs es usado universalmente, en inglés, single nucleotide polimorphisms y en español se entiende como mutación de punto) corriente arriba del codón iniciador del gen de la LPH que correlacionan con los dos fenotipos, uno, C/T a –13910 pb, y el otro, G/A a –22018 pb.
G/A-22018 muestran persistencia de la lactasa (Troelsen, 2005). Recientemente se han asociado otros marcadores en grupos que cuentan con historia pastoril; para africanos estos son los SNPs G/C-14010, T/G-13915 y C/G-13907 (Tishkoff et al. 2006) y para grupos pastoriles árabes T/C -3712 y T/G -13915 (Enattah et al. 2008). De esta manera, el conocimiento de los genotipos en cada uno de estos sitios y de los haplotipos que conforman la combinación de genotipos entre estos marcadores además del aspecto ancestral podría servir como indicadores de susceptibilidad a padecer de problemas asociados a la digestión de la lactosa.
1.4 Distribución de la capacidad para digerir la lactosa en diferentes poblaciones
del mundo
La distribución de los polimorfismos asociados con la capacidad de digerir la lactosa se ha correlacionado con las prácticas culturales de las regiones en que se domesticó el ganado, y en consecuencia surgieron las condiciones para el consumo de leche fresca (Durham, 1991; Hollox et al 2001).
En las regiones en que se adoptó la práctica pastoril, norte de Europa y noreste de África, el porcentaje de personas con persistencia de la lactasa es muy alta (> 70%), mientras que ésta disminuye (30-70%) en las regiones Mediterránea, del Oriente medio, sur de África, así como en Asia Central y Sur (Holden & Mace, 1997). Esta correlación ha sugerido que en los grupos que histórica y culturalmente han consumido leche fresca y sus derivados, se dió una selección natural a favor de la persistencia de la lactasa y por ello se observan los diferentes patrones de tolerancia.
55% (Peña et al. 2002), Guinea 83% (Gibney et al. 1981), Estados Unidos 25% (en este caso se da el porcentaje por etnia dentro de la población estadounidense con una intolerancia de 80% para afroamericanos, 62% a 100% indígenas Pima y 90% asiáticos) (Inman-Felton, 1998). Para poblaciones mongoloides incluyendo los amerindios, la actividad desaparece después de los primeros años de vida (Huang & Bayless, 1968; Newcomer et al, 1977; Jobling, 2004).
Debido al desarrollo de la industria lechera estas diferencias se hicieron evidentes conforme se promovió el consumo de lácteos por parte de las empresas productoras y programas de ayuda nutricional (USDA, 2010). Algunos investigadores señalaron la existencia de “etnocentrismo nutricional” (McCraken, 1971), “falacia de la normalidad médica” (Williams, 2000) o “bioetnocentrismo” (Wiley, 2008) como crítica a las políticas nutricionales que promueven el consumo de leche en todas las poblaciones debido a la tendencia de considerar como “normales” características que son comunes sólo en ciertos grupos, principalmente europeos, considerando así la hipolactasia como una desviación y no como una condición ancestral (Liberman, 1978; Wiley, 2008).
Es de particular importancia el conocimiento sobre la susceptibilidad a la intolerancia a la lactosa en estos grupos debido al potencial impacto de los cambios dietéticos sufridos como consecuencia de la creciente aculturación (Bozzoli & Guevara, 2002). Se espera que estos cambios sean mayores en poblaciones donde se facilite el acceso al consumo de los productos lácteos que se da como resultado de los procesos de expansión comercial de empresas productoras o cooperativas locales (Mata et al. 1985) o que llegan a ellas a través del estado con programas de alimentación que actualmente se hacen efectivos a través de asociaciones de desarrollo locales llamadas Pro-CEN-CINAI, las cuáles distribuyen mensualmente leche íntegra a sectores con mayores índices de pobreza. Actualmente los beneficiados de esta distribución son 72 300 niños a nivel nacional entre los que se toma en cuenta a los grupos indígenas (MIDEPLAN, 2006). Es un número considerable de hogares que disponen de este alimento mensualmente agregándose a los productos de uso común, dentro de la canasta básica (INEC, 2002).
El interés en el estudio de las poblaciones amerindias obedece, además de su aislamiento durante miles de años de poblaciones con cultura del consumo de lácteos, a su importancia demográfica (altas tasas de reproducción) repercutiendo y teniendo su impacto sobre los grupos mestizados con las cuales ellos comparten su caudal genético (Arias, 1991; Barrantes & Morera, 1999).
2. Objetivos
2.1 Objetivo General
Estudiar la variación molecular asociada con intolerancia a la lactosa en grupos amerindios de Baja Centroamérica.
2.2 Objetivos específicos
1. Caracterizar y cuantificar la variación genética de una región que comprende al gen
LCT en distintos grupos amerindios.
2. Estimar si la prevalencia de variantes europeas y africanas concuerda con los datos preexistentes de mezcla para cada población.
3. Evaluar la neutralidad de las variantes encontradas en la región genómica que incluye al gen LCT para comprobar la hipótesis de la influencia de selección natural sobre este locus.
3. Marco Teórico
Con los cambios que se dieron alrededor del neolítico en algunas sociedades humanas, cuando surge el pastoralismo, y por consiguiente se inicia el consumo de leche en edades posteriores a la ablactación, se origina un cambio ambiental que ha puesto en evidencia la diferencia genética entre individuos en su capacidad de digerir la lactosa en esas edades. Por lo tanto las principales hipótesis para explicar el fenómeno de tolerancia se ubica en dos categorías, la primera se centra en la selección natural a favor de la mutación que confiere tolerancia asociada a un cambio dietético por disponibilidad de nutrientes y prácticas culturales (Feldman & Cavalli- Sforza, 1976; Simoons, 1978; Durham, 1991; Bersaglieri et al. 2004; Enattah, 2002 et al; 2007; Tishkoff et al. 2006; Burger et al. 2007) y, la segunda, que por el contrario se basa en la distribución de frecuencias alélicas dependientes de procesos evolutivos no determinísticos como son la deriva genética, el efecto fundador y la migración, todos ellos relacionados entre sí (Nei & Saitou, 1986; Swallow, 2003). Esta hipótesis es llamada de la causa inversa (McCraken, 1971), y se fundamenta en la presencia significativa de la mutación en las poblaciones antes de la domesticación por lo que su propagación no fue impulsada inicialmente por la selección (Peña et al. 2002; Brines, 2004). Los detalles de las hipótesis mencionadas además de otras que se han planteado, se reseñan a continuación con mayor detalle.
3.1 Desarrollo histórico de las hipótesis que explican el fenómeno de intolerancia a
la lactosa.
a) Hipótesis de la enfermedad. Según ésta, algunos grupos eran más propensos a sufrir intolerancia debido a su alta prevalencia de enfermedades infecciosas, principalmente aquellas de tipo gastrointestinal (Keush, 1972; Ransome-Kuti, 1977; Anderson & Vullo, 1994), sin embargo esto no se sostuvo ya que podía ser aplicable a casos individuales o grupos aislados pero no explicaba las diferencias observadas entre los grupos de distinta ancestría que compartían el mismo ambiente (Harrison, 1975).
b) Hipótesis de la inhibición dietética. Se refería a que algunos grupos consumen en su alimentación o en sus tratamientos médicos sustancias que pueden inhibir la actividad de la lactasa (Simoons, 1969). Sin embargo ésta no se ha comprobado para ningún alimento o grupo de ellos, pues ninguno ha provocado una mala absorción de forma tan profunda en tantas dietas como existen sistemas de subsistencia y ambientes diferentes.
c) Hipótesis de la inducción. Propuesta por Bolin et al (1969), señalaba que si después del destete en la infancia se continuaba con el consumo en pequeñas o grandes cantidades, se mantendría inducida la lactasa y persistiría en la adultez. Por lo demás, esto sería congruente con el hecho de que el operón de la lactasa en bacterias es el ejemplo clásico de la inducción enzimática (Jacob & Monod, 1961; Watson et al. 2008). Un número de experimentos trataron de comprobarlo sin embargo tales intentos fallaron (Cuatrecasas et al. 1965; Johnson et al. 1981; Hawkins, 1996); pues aunque se lograba una mejor absorción y se disminuían los síntomas, la actividad de lactasa se mantenía en bajos niveles, y el cambio se debía a la presencia de ciertas bacterias intestinales capaces de digerir la lactosa, concluyendo así, que la inducción enzimática no aplicaba en este caso (Johnson et al. 1981; De Vrese et al. 2001).
ocasionaba esta distribución no estaba claro (Harrison, 1975). Se plantearon varias hipótesis, una de ellas propuesta por Flatz y Rotthauwe (1973) que sugería que en las latitudes altas donde la luz solar es limitada los humanos carecen de vitamina D y consecuentemente adolecen de raquitismo, por lo que la ingesta de leche resultaría favorable para quienes toleracen la lactosa y con la ingestión de leche se promoviera la absorción de calcio, como lo hacía la vitamina D (Holden & Mace, 1997). Otra hipótesis fue la de Cook y Al-Torki (1975) en la que se establecía que en lugares altamente áridos donde la disponibilidad de agua era limitada el consumo de leche fue importante para llevar a cabo la hidratación y por lo tanto se produjo un diferencial selectivo.
Estas diferencias muestran también una distribución particular entre grupos étnicos con localizaciones geográficas asociadas al desarrollo pastoril. A partir de esta distribución se plantea la hipótesis de coevolución gen-cultura, también conocida como hipótesis histórico-cultural, la cual sigue la línea de la hipótesis genética, proponiendo que la persistencia de la lactasa en la adultez es una adaptación milenaria al pastoralismo y consumo de leche. En este caso se propone que la capacidad para digerir la lactosa tuvo una ventaja selectiva en adultos de poblaciones pastoriles al tener disposición de beneficios nutricionales a través de la leche (Holden & Mace, 1997; Bloom & Sherman, 2005; Enatthah, 2002; Tishkoff et al. 2006; Burger et al. 2007). Recientemente Cochran y Harpending (2009) propusieron que las poblaciones con el rasgo de tolerancia (indoeuropeas) se encontraron en ventaja frente a otros grupos debido a que en procesos bélicos que demandaban movilidad contaban con el alimento y por lo tanto se encontraban mejor nutridas al poder consumir leche y carne de los rumiantes.
reproducción de genotipos. Este enfoque, se enmarca en la propuesta de coevolución genético-cultural de Feldman y Cavalli–Sforza (1976), que señala una coacción entre la transmisión vertical (genes) y la transmisión horizontal (cultura), lo que ha sido considerado específicamente para el caso de la persistencia de la lactasa (Feldman & Cavalli-Sforza, 1976; 1989; Aoki, 2001). Siempre que hubiera una historia cultural sin la práctica lechera o que la preferencia cultural se inclinara hacia el procesamiento de la leche se encontraron bajos niveles de absorción en adultos. Cabe destacar que los estudios realizados en este momento se basan en esta propuesta y últimamente se le ha llamado selección natural mediada por la cultura (Aoki, 1986; Bersaglieri et al. 2004; Tishkoff et al. 2006, Burger et al. 2007; Enattah et al. 2007, Gerbault et al. 2009). Cuando se menciona selección positiva se refiere a un proceso en el cual los individuos de regiones con cultura pastoralista tenían ventaja en su ambiente, sea porque estuviesen en algún momento bajo estrés dietético o por una ventaja a nivel social que se reflejara en una mayor reproducción (Gerbault et al. 2009). Concordante con esto son los resultados de las estimaciones de antigüedad para las altas frecuencias de las mutaciones africanas y europeas están alrededor de 7000 años A.C. (Tishkof et al. 2006).
e) En este contexto surge la hipótesis de la causa inversa, una hipótesis contraria a la mediación cultural. Esta hipótesis no descarta que haya existido un papel selectivo a favor del rasgo de tolerancia, pero contempla su papel dentro de un marco de variación favorable preexistente. Es decir, la variación del rasgo tolerancia como resultado de un efecto fundador y la deriva genética a nivel mundial (características propias de las poblaciones naturales (Kimura, 1968) dado como resultado de las migraciones que se dieron en la dispersión de los seres humanos desde África hacia los diferentes continentes alrededor de 100 000 A.C. (Nei & Saitou, 1986; Swallow, 2003; Brines, 2004).
que se ignora la pregunta sustancial sobre el origen de la tolerancia a lactosa en adultos y no se explica por qué sólo algunos grupos de humanos adultos son los únicos mamíferos equipados bioquímicamente para metabolizar la lactosa. Para McCraken, la hipótesis no es comprobable, pues no puede ser validada o confirmada y es meramente tautológica. No es una hipótesis dinámica, ya que reconoce la influencia de las frecuencias génicas en la adopción de un rasgo cultural pero no en el sentido contrario.
Al tener un marco de hipótesis explicativas con respecto al rasgo en estudio se debe tener presente un aspecto substancial en cuanto a la evolución de los amerindios, para lograr una comprensión del potencial problema de salud.
La llegada de los primeros americanos se dio alrededor de 12 500 años antes del presente (Torroni et al. 1994, Meltzer, 2009). Los grupos que pertenecen al grupo lingüístico chibcha se asentaron en Baja Centroamérica hace alrededor de 10 000 años; y desde entonces estuvieron sujetos a procesos evolutivos de las poblaciones naturales tales como deriva genética y cuellos de botella (Neel & Salzano, 1964). Durante este lapso se desarrolló la agricultura y ganadería en Europa y Medio Oriente por lo que los amerindios se mantuvieron ajenos estas las prácticas culturales durante el neolítico. Con la llegada de los europeos 500 años atrás se produjo además de disminución demográfica, dispersión de los grupos y mezcla interétnica (Barrantes, 1990) la inserción de nuevos rasgos culturales tales como la cría de ganado. El ganado fue introducido al continente en 1493 por Cristóbal Colón en su segundo viaje (Primo, 1992).
4. Metodología
4.1 Poblaciones de estudio
Este trabajo se realizó con grupos indígenas de Costa Rica y uno de Nicaragua de filiación lingüística chibcha a excepción del grupo chorotega (grupo otomangue) localizado en el pacífico costarricense (Constenla, 1991). La mayoría de estos grupos habitan en diferentes sectores montañosos de Costa Rica que anteriormente fueron llamadas “reservas indígenas” pero que actualmente son reconocidos como “territorios indígenas” por el estado costarricense (Guevara & Salas, 2000). Solo un 42% de la población indígena vive dentro de los territorios indígenas (Pérez-Brignoli, 2005) y este porcentaje asciende a un total de 37 404 personas (Guevara & Salas, 2000). La elección de estos grupos está basada principalmente en el interés comparativo tanto por los diferentes grados de mezcla como por la distribución geográfica en que se encuentran. Se presenta una breve reseña de cada uno de ellos a continuación.
4.1.1 Huetar
Este grupo habita en dos comunidades, Quitirrisí y Zapatón, que se encuentran al oeste de San José. Se estima que su población oscila entre 1600 y 1800 personas (Guevara & Salas, 2000; Pérez-Brignoli, 2005). Se ha descrito como una de las poblaciones con mayor mezcla interétnica y por lo tanto con el mayor grado de aculturación (Ruiz-Narváez et al. 2005), sin embargo estudios genéticos han comprobado su ancestría indígena e indicado su relación con otros grupos chibchas de la región (Barrantes et al. 1990); la cual es mayor con los grupos talamanqueños que con los Maleku (Azofeifa et
al. 2001).
4.1.2 Maleku
2000). Estudios genéticos y lingüísticos han relacionado mayormente a esta población con grupos no talamanqueños (Teribe y Kuna) situando la separación de estos alrededor de 7000 años atrás (Barrantes et al. 1990b).
4.1.3 Cabécar
Lo forman alrededor de 14 000 habitantes (Guevara & Salas, 2000). Es uno de los grupo menos aculturados y étnicamente menos mezclados (Barrantes, 1993b), probablemente por su aislamiento en las regiones remotas ubicadas en el sector atlántico de la Cordillera de Talamanca (Gabb, 1981). Este aislamiento ha permitido que el grupo mantenga su cultura lo que se manifiesta en un 85% de la población que habla el idioma (Guevara & Salas, 2000).
4.1.4 Bribri
El grupo bribri también es un grupo talamanqueño que presenta gran similitud cultural y genética con los cabécares. Cerca del 70 % de la población habla el idioma bribri (Guevara & Salas, 2000) lo que refleja un grupo que se mantiene en gran medida su riqueza cultural. Cuentan con un aproximado de 12 000 habitantes (Guevara & Salas, 2000) conformando así uno de los grupos más numerosos de Costa Rica.
4.1.5 Ngöbe
4.1.6 Rama
El grupo Rama habita el sureste caribeño de la costa nicaragüense y se estima que su población se encuentra entre 1350 a 3000 personas (Pérez-Brignoli, 2005; Melton, 2008). Se ha sugerido que son un grupo remanente de los Suerre, Huetar, Corobicí, Maleku, Voto, Melchora, Kukra o Caribes, aunque tomando en cuenta marcadores del cromosoma Y se ha comprobado que se encuentran estrechamente relacionados con el grupo chorotega (Melton, 2008).
4.1.7 Chorotega
Se estima que habitan 1436 personas en la península de Nicoya dentro el territorio (Guevara & Salas, 2000). Aunque poseen una filiación lingüística distinta al chibcha al ser una población mesoamericana son considerados el límite con los grupos chibchas (Constenla, 1991). La evidencia ha mostrado una relación genética estrecha con estos, pues marcadores mitocondriales y del cromosoma Y sugieren que existieron uniones entre hombres de este grupo mesoamericano con mujeres del grupo chibcha (Melton, 2008). Gran parte de su cultura autóctona no se conserva hoy en día, sin embargo movimientos de etnogénesis aspiran a mantener la distinción histórico-cultural de este grupo en relación con los otros grupos remanentes chibchas (Guevara & Salas, 2000).
4.2 Encuesta sobre la percepción de los efectos del consumo de leche en dos
comunidades indígenas.
Palenque Tonjibe. Debido a que el número de cuestionarios que se logró aplicar fue muy bajo y los datos obtenidos no son estadísticamente confiables ni concluyentes, los resultados de esta averiguación se muestran en el anexo 2.
4.3 Análisis genético
Se utilizaron muestras de ADN de las siguientes poblaciones amerindias: Maleku (n2=31), Ngöbe de las comunidades de Abrojo (n=42), Dos Brazos (n=14) y Limoncito (Limoncito) (n=10); Huetar de las comunidades de Zapatón (n=35) y Quitirrisí (n=15); Bribri de la comunidad de Mojoncito (n=21); Cabécar de la comunidad de Alto Telire (n=18); Chorotega de la comunidad de Matambú (n=16) y Rama (n=12) de la comunidad de Rama Cay. Todas estas muestras fueron colectadas durante el desarrollo del programa “Ecología, diversidad genética y salud de los amerindios de Costa Rica”, inscrito en la Vicerrectoría de Investigación bajo el No 742-93-903, por lo que cumplió con todos los procedimientos de aprobación requeridos, así como con los consentimientos de los líderes y comunidades, según era requerido entonces. Más detalles sobre las metodologías de colectas de muestras se encuentran en Barrantes (1990). El número total de muestras analizadas ascendió a 214. En la selección de las muestras se evitó tomar aquellas que pertenecieran a una misma familia. Es importante mencionar que dada la estructura de las poblaciones amerindias el tamaño de la muestra puede representar con confiabilidad el grupo en estudio pues son poblaciones pequeñas y muy emparentadas. Estas relaciones cercanas entre familias son propias de estos grupos amerindios según se ha descrito (Barrantes, 1993a).
Para cada una de estas muestras el ADN fue preparado a concentraciones finales de 25 ng/ul para cada una; esta es una concentración apropiada para llevar a cabo la reacción en cadena de la polimerasa (PCR), técnica básica para esta investigación. Se utilizaron 9 marcadores genéticos de tipo SNP (por sus siglas en inglés Single Nucleotide Polimorfism) (Cuadro 1). Ocho de estos marcadores fueron descritos por Enattah et al. (2005, 2007); el otro que fue relacionado con tolerancia a la lactosa en grupos africanos
2
por Tishkoff et al.(2006). Los 9 se encuentran en una región de 19 kb ubicada corriente arriba del gen de la lactasa (Enattah et al. 2007) (Fig 2).
Cuadro 1. SNP´s analizados en las muestras de ADN para todas las poblaciones. SNP Tipo y posición
respecto al gen de la lactasa
dbSNP (GenBank)* Posición en el cromosoma**
1 C/T -8540 rs3754686 136437008
2 G/C -8630 rs3769005 136437098
3 C/T -13910*** rs4988235 136442378
4 G/C -14010 + -14010 lactasa 136442478
5 C/T -15239 rs4954493 136443707
6 G/C -20077 rs3099181 136448545
7 G/A -22018*** rs182549 136450486
8 C/T -27312 rs4988183 136455779
9 A/C -27807 rs3087343 136456274
*Número de acceso en GenBank (online) se encuentra en: http://www.ncbi.nlm.nih.gov/Genbank/ **Posición en el cromosoma según GenBank.
*** Tipos y posiciones descritas y asociadas para poblaciones europeas.
+
Tipo y posición descrita y asociada para poblaciones africanas.
Figura 2. Distancia (Kb) entre los diferentes marcadores utilizados (Modificado de Enattah et al. 2007).
Para cada muestra se requirió amplificar siete regiones diferentes, pues dos de los SNP`s están contenidos dentro de un mismo producto amplificado. Las disoluciones de PCR se realizaron en volúmenes finales de 25 ul compuestos por 2 ul de ADN [50 ng], 2.5 ul de Buffer (NH4)SO4 [15mM], 5 ul de MgCl2 [25 mM], 1 ul de cada primer [5 uM], 0.2 ul
posteriormente cada ciclo se compuso de una desnaturalización de 95º C por 1 minuto, una temperatura de hibridación (annealing) de 60º C durante 1 minuto, una extensión a 72º C durante 1 minuto y una extensión final de 5 minutos. Los fragmentos de PCR fueron separados por medio de electroforesis en geles de poliacrilamida al 10% seguida de una tinción con plata. Los imprimadores rs4988183 y el rs4954493 se integraron en una PCR dúplex y los imprimadores rs3754686, rs3099181 y rs3087343 en una triplex (Fig 3).
Figura 3. Amplificación de fragmentos por PCR-triplex con los marcadores rs309181, rs3754686 y rs3087343 observados en un gel de acrilamida.
Para la detección de los SNP`s se utilizaron dos métodos de genotipeo: corte por enzimas de restricción visualización en geles de poliacrilamida y detección por medio de electroforesis capilar.
Los RFLPs permitieron analizar dos de las variantes, específicamente la variante C/T en el nucleótido -13910 y la variante G/A en el nucleótido -22018. Los imprimadores, condiciones de amplificación por PCR y las enzimas para analizar las variantes C/T-13910
y G/A-22018 fueron las descritas por Coelho y colaboradores (2005). Estos sitios
polimórficos sirven para definir haplotipos que han sido caracterizados para otras poblaciones en el mundo (Hollox et al. 2001).
Se determinaron 7 genotipos con el método de minisecuenciación, utilizando el kit SNaPshot Multiplex de la casa Applied Biosystems. Para este paso, fue necesario amplificar las regiones en las que se encuentran estos marcadores y purificarlas para
Ladder (50 pb)
1000 pb
300 pb
llevar a cabo la reacción de minisecuenciación según lo especificado por el kit. Luego fue llevado al analizador genético ABI Prism 3130. Para la recopilación de los datos se utilizó el software Foundation Data Collection v.3.0 y para su lectura el software Genemapper v. 4.0.
El genotipeo con el SNaPshot multiplex kit fue probado con los diferentes imprimadores individualmente, y los resultados de algunos de ellos no concordaron con los datos esperados, por lo que fue necesario diseñar nuevos imprimadores (ver anexo 2). También se evaluó el desempeño de los imprimadores al ser combinados en una misma reacción, y el tamaño de los productos fue determinado sin ambigüedad. De esta manera se estandarizó la técnica, utilizándose para ello ADN control, con el que se obtuvo finalmente la adecuada combinación de marcadores (ver anexo 3). Para las PCRs que no amplificaron, se variaron las cantidades de reactivos iniciales y el perfil de amplificación (ver anexo 3). Para corroborar los resultados de la estandarización, algunos segmentos de ADN fueron secuenciados, asimismo se llevo a cabo la secuenciación para aquellas muestras en que el genotipeo no era claro.
Una vez obtenidos los genotipos de cada sitio polimórfico se procedió a estimar las frecuencias alélicas, heterocigosis y equilibrio de Hardy-Weinberg por locus con el programa GenAlex versión 6.2 (Peakall R & Smouse P, 2006). Se estimaron los haplotipos utilizando el programa PHASE versión 2.1 (Stephens et al. 2001) utilizando para ello los parámetros sugeridos en el programa. También se estimaron la medidas de diversidad haplotípica (Ĥh) y Fst para conocer la diferenciación genética entre las
5. Resultados
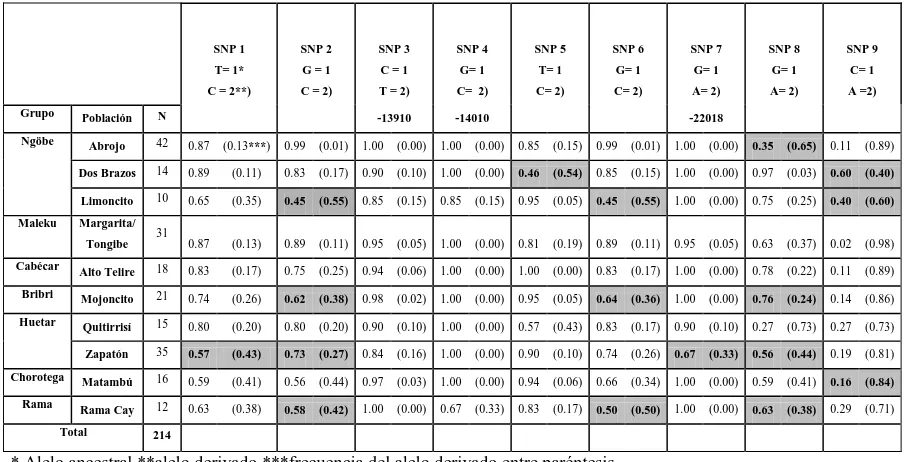
En el cuadro 2 se presentan las frecuencias alélicas para cada uno de los 9 loci analizados. Los alelos codificados como 1 representan el alelo considerado como ancestral, y los alelos codificados como 2 los alelos que cosegregaron con el fenotipo de persistencia de la lactasa según Enattah et al. (2007). Las frecuencias alélicas en los distintos marcadores genéticos (SNPs) mostraron una variación entre 0.5 y 1 en la mayoría de las poblaciones. La excepción se presentó en las frecuencias alélicas de los SNPs 4 y 7 (cuadro 2).Valga señalar que los alelos ancestrales del SNP 4 (G-14010) y los
del SNP 7 (G-22018), que son dos de los asociados con el fenotipo de tolerancia/
intolerancia, están fijados en 8, para el SNP4 y 7 para el SNP 7, de las 10 poblaciones estudiadas. Las frecuencias de los alelos ancestrales en los otros 7 SNPs oscilaron entre 0.11 (SNP 9, Abrojo y Alto Telire) y 1.0 (SNP 3, Abrojo y Rama Cay; SNP 5, Alto Telire). Resalta la alta frecuencia del alelo derivado en el SNP 9 para todas las poblaciones.
Cuadro 2. Frecuencias alélicas de 9 loci (SNP) analizados en 10 poblaciones indígenas. Los loci que se encuentran en desequilibrio Hardy-Weinberg se muestran sombreados.
SNP 1
* Alelo ancestral **alelo derivado ***frecuencia del alelo derivado entre paréntesis.
Cuadro 3. Diversidad génica (Ĥ) por locus y por población y heterocigosis promedio (desviación estándar) para 10 poblaciones indígenas.
Grupo Población SNP 1 SNP 2 SNP 3 SNP 4 SNP 5 SNP 6 SNP 7 SNP 8 SNP 9 Ĥ
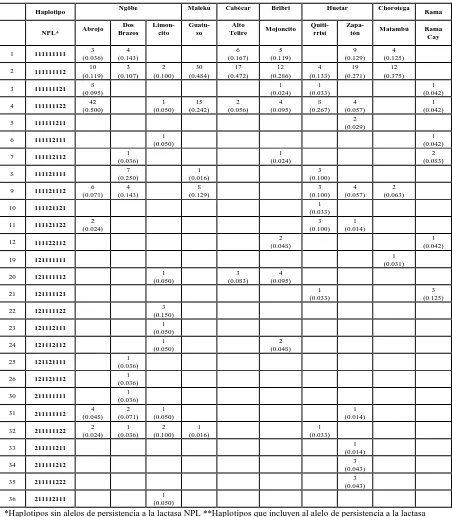
Cuadro 4. Frecuencias haplotípicas absolutas (relativas) en 10 poblaciones indígenas.
Haplotipo Ngöbe Maleku Cabécar Bribri Huetar Chorotega Rama
NPL* Abrojo
*Haplotipos sin alelos de persistencia a la lactasa NPL **Haplotipos que incluyen al alelo de persistencia a la lactasa
Continuación…. Frecuencias haplotípicas absolutas (relativas) en 10 poblaciones
indígenas.
Haplotipo Ngöbe Maleku Cabécar Bribri Huetar Chorotega Rama
NPL* Abrojo
*Haplotipos sin alelos de persistencia a la lactasa NPL **Haplotipos que incluyen al alelo de persistencia a la lactasa
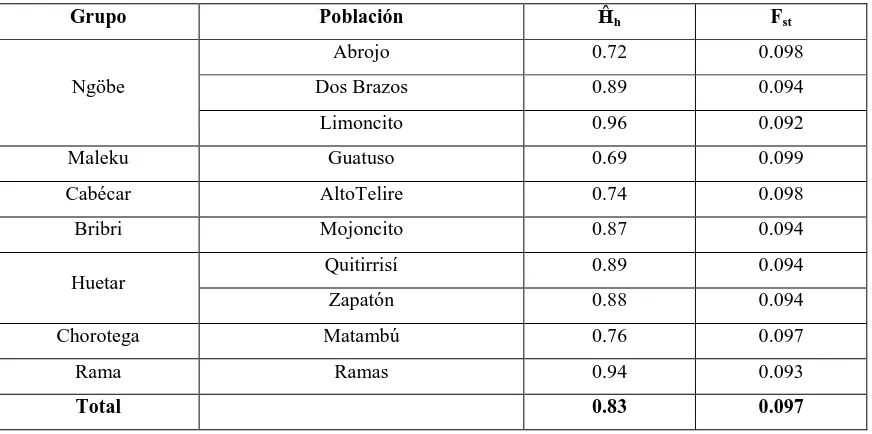
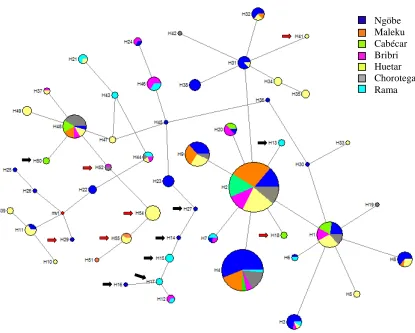
La diversidad haplotípica (Ĥh) para los haplotipos (Cuadro 5) fue mayor en la población
ngöbe de Limoncito (0.96) y el grupo Rama (0.94); contrario a las poblaciones de los grupos ngöbe de Abrojo (0.72), maleku (0.69), cabécar (0.74). El valor de Fst total (de
0.097) refleja poca diferenciación genética entre las poblaciones estudiadas para esta combinación de marcadores; valores similares han sido obtenidos con otros marcadores autosómicos clásicos (Barrantes, 1993b; Ruiz-Narváez et al. 2005). La poca diferenciación de los grupos es reflejada en el diagrama de la figura 4. Los haplotipos más frecuentes en el total (H1, H2, H9 y H48) se distribuyen de manera similar en todas las poblaciones, y son pocos los haplotipos que se encuentran solamente en alguno de los grupos estudiados. Además es evidente que aquellos grupos cuyo número de muestra es mayor presentan más diversidad de haplotipos a bajas frecuencias (ngöbes, representados con el color azul y huetares, con color amarillo) (fig 4).
Cuadro 5. Diversidad haplotípica (Ĥh) y Fst por población y total.
Grupo Población Ĥh Fst
Ngöbe
Abrojo 0.72 0.098
Dos Brazos 0.89 0.094
Limoncito 0.96 0.092
Maleku Guatuso 0.69 0.099
Cabécar AltoTelire 0.74 0.098
Bribri Mojoncito 0.87 0.094
Huetar Quitirrisí 0.89 0.094
Zapatón 0.88 0.094
Chorotega Matambú 0.76 0.097
Rama Ramas 0.94 0.093
Figura 4. Diagrama de haplotipos (Median Joining) para 9 SNPs en la región de 19 kb del LCT. El tamaño del círculo corresponde a la frecuencia haplotípica estimada, y los colores representan los grupos que comparten determinado haplotipo (haplogrupo). Las flechas rojas señalan los haplotipos que contienen la variante europea asociada con tolerancia y flechas negras los haplotipos que contienen la variante africana. La definición de los haplotipos se encuentra en el cuadro 4.
La prueba de neutralidad para haplotipos de Ewens- Waterson no rechazó la hipótesis nula para 9 de las poblaciones (cuadro 6). En el caso de la población de Rama Cay la significancia puede ser debida al tamaño de la muestra (n = 10).
Cuadro 6. Prueba de neutralidad Ewens-Waterson para 10 poblaciones indígenas.
5.3 Estimación de la prevalencia del polimorfismo tolerancia/intolerancia a la
lactosa
Para estimar la prevalencia de este polimorfismo en las poblaciones estudiadas, se asumió que las relaciones observadas entre los polimorfismos del SNP 4 (C/T-13910) y el
SNP 7 (G/A-22018) en poblaciones europeas, y el SNP 4 (G -14010) en poblaciones
africanas, se presenta también en los grupos amerindios.
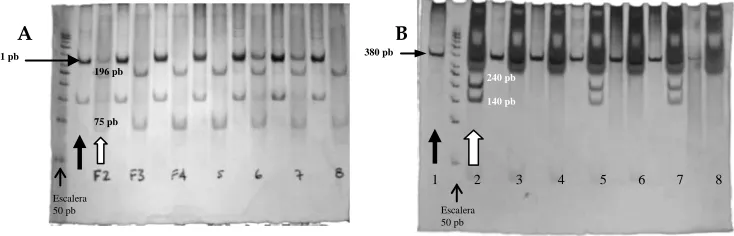
Los alelos de persistencia europeos se pueden visualizar mediante RFLPs dado que el origen de la variación nucleotídica eliminó a la vez los sitios de restricción, esto es, los alelos derivados, correlacionados con la persistencia de la lactasa, no son cortados por las enzimas de restricción (fig. 5).
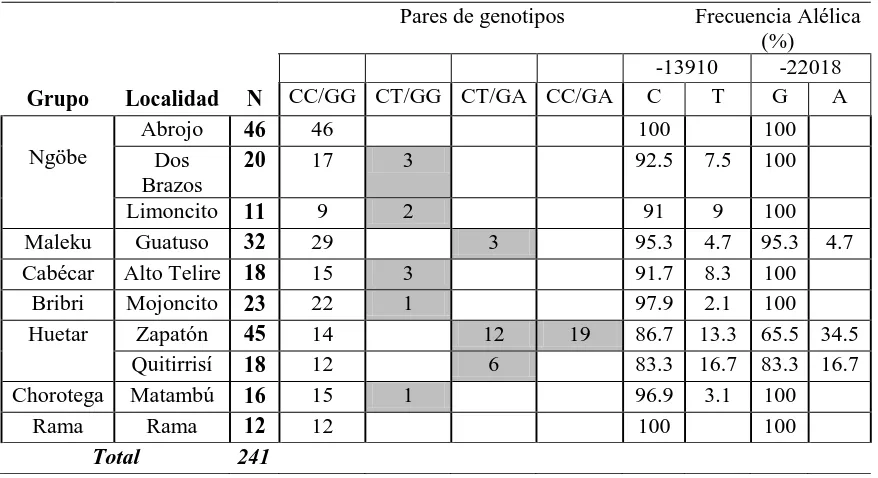
Las mayores frecuencias del alelo A del SNP 7 (gel A de la figura 5) y del alelo T del SNP 3 (gel B en la figura 5) se observaron en el grupo huetar, en el caso particular de la comunidad de Zapatón se observó una mayor frecuencia de individuos con la variante A del SNP 7(Cuadro 7). Considerando solamente la presencia de los alelos europeos asociados con tolerancia/intolerancia la población ngöbe de Abrojo y la población de Rama Cay presentarían un 100% de intolerancia. A su vez las poblaciones huetares, Quitirrisí y Zapatón, son las que presentarían menor grado de intolerancia con un 83.3 % y 86.7% respectivamente.
Cuadro 7. Frecuencias genotípicas conformados por la combinación de los SNP`s 3 (C/T-13910) y 7 (G/A-22018) asociados en los grupos europeos con el polimorfismo tolerancia/ intolerancia (las frecuencias de los genotipos tolerantes se muestran sombreados).
Grupo Localidad N
Pares de genotipos Frecuencia Alélica (%)
Maleku Guatuso 32 29 3 95.3 4.7 95.3 4.7
Cabécar Alto Telire 18 15 3 91.7 8.3 100
Bribri Mojoncito 23 22 1 97.9 2.1 100
Huetar Zapatón 45 14 12 19 86.7 13.3 65.5 34.5
Quitirrisí 18 12 6 83.3 16.7 83.3 16.7
Chorotega Matambú 16 15 1 96.9 3.1 100
Rama Rama 12 12 100 100
Total 241
Como se ha mencionado el SNP 4 (G/C -14010) también ha presentadoasociación con el
según la presencia de ambos, aquellos individuos que porten alguna tendrían menor probabilidad de sufrir intolerancia de acuerdo con los estudios de asociación existentes (Ridefelt & Hakansson, 2005) (Cuadro 8).
Cuadro 8. Frecuencias genotípicas conformados por la combinación de los SNP`s 3 (C/T -13910) y 4 (G/C-14010), el primero asociado en grupos europeos y el segundo en grupos africanos con el polimorfismo tolerancia/intolerancia, y porcentaje esperado* de individuos susceptibles a la intolerancia a la lactosa (las frecuencias de los genotipos tolerantes se muestran sombreados).
Grupo étnico Población N
Combinaciones de frecuencias genotípicas en los SNPs (-13910/-14010)
Susceptibilidad a la intolerancia
a la lactosa (% teórico*)
CC/GG
(susceptibles) CT/GG CC/GC
Ngöbe
*Suponiendo que la asociación encontrada en Europa y Africa es válida también en los amerindios.
6. Discusión
Los valores de frecuencias alélicas muestran la existencia de dos grupos de marcadores, aquellos que se encuentran fijados o a altas frecuencias (SNPs 3, 4 y 7) y los que se encuentran variando entre 0.01y 0.99 (SNPs 1, 2, 5, 6, 8 y9). De manera concordante en los SNPs que alcanzan la fijación se observaron los menores índices de heterocigosis promedio, 0.12, 0.07, 0.07 respectivamente, mientras que el promedio fue de 0.24 (cuadro 3). Las desviaciones estándar de las heterocigosis promedio por locus se mostraron muy bajas, pero con valores similares en todos los SNPs, lo que podría ser una indicación de que tales marcadores han variado de manera aleatoria desde que las poblaciones se separaron de su ancestro común. Por el contrario, los SNPs 4 y 7, cuyos alelos derivados han sido asociados con tolerancia a la lactosa en africanos y europeos respectivamente, muestran una desviación de ese patrón de cinética de las frecuencias alélicas al estar fijados o casi fijados los alelos ancestrales. Tales resultados también han sido observados en grupos asiáticos, como los chinos Han, surcoreanos y en paquistaníes (Ennatah et al. 2007); sin embargo en la mayoría de grupos a nivel mundial las frecuencias de los SNPs 4 y 7 varían entre 0.03 y 0.97 sin fijación. Lo anterior podría sugerir una fijación conservada en estos SNPs durante el proceso evolutivo en algunos grupos humanos, descendientes de Asia. No se descarta que tales resultados sean producto de selección que esté ocurriendo para otro fenotipo (por ejemplo, división celular en el cual está involucrado el MCM6, misma región que contiene las variantes asociadas con tolerancia/intolerancia) y que se está reflejando en estos marcadores. Sin duda este fenómeno no tendría relación con el rasgo de tolerancia a la lactosa, pues la leche no ha sido un factor ambiental para los grupos amerindios ya que la domesticación de animales que producen leche no fue una actividad posible para los primeros habitantes del continente.
población ngöbe de Limoncito, presentó una frecuencia de 0.15 en el SNP 4 (asociado con tolerancia en grupos africanos), una frecuencia aparentemente alta, pero dado el pequeño número muestral (n=10), es producto de 3 alelos. Según estudios genéticos anteriores el grupo ngöbe ha sufrido poca mezcla interétnica, aunque posiblemente esté creciendo en las generaciones recientes. Apoya esta interpretación el hecho de que los tres individuos que presentaron el alelo asociado con poblaciones africanas en condición heterocigota corresponden a individuos entre 20-24 años.
variante africana G (SNP 4) también asociada con tolerancia a la lactosa (cuadro 7) se encontraron resultados aparentemente discordantes, los cuales mostraron que las poblaciones rama y ngöbe de Limoncito son las que presentan las mayores frecuencias de la variante y como consecuencia un mayor grado de tolerancia.
Con respecto a la variante G (SNP 4) es importante aclarar que se trata de un marcador africano asociado con la tolerancia a la lactosa, cuyo origen se traza a África del Este y que alcanza sus mayores frecuencias en Tanzania (50%) y Kenia (30%) (Tishkoff et al. 2006). Al considerar esta variante como posible protector contra la intolerancia a la lactosa los niveles de susceptibilidad cambian los grupos mencionados, y sería para los ramas de un 33.3% y los ngöbes de Limoncito un 40%. La presencia de la variante en la población ngöbe de Limoncito y Ramas podría indicar una introducción por mezcla interétnica de estos dos grupos con pobladores no indígenas y de ascendencia africana (África oriental) sin embargo, es necesario analizar un mayor número de muestras y complementar con otros marcadores genéticos informativos y mediciones directas de tolerancia a lactosa en los individuos para ampliar este análisis.
Para las otras poblaciones indígenas se muestra que en la población de Abrojo, perteneciente al grupo ngöbe, la intolerancia alcanza la totalidad de la población, seguida del grupo bribri, luego chorotega, maleku y las comunidades del grupo huetar. Sin embargo en el caso del grupo ngöbe, se debe considerar que la intolerancia varía dependiendo de la localidad o de la subestructura del grupo y también del grado de mezcla interétnica que presente cada comunidad (Barrantes et al. 1982). Esto a su vez explica que el grupo ngöbe de Abrojo se diferencie de las comunidades de Limoncito y muestren frecuencias opuestas tanto para la variante europea T (SNP 3) como para la variante africana C (SNP 4)(cuadro 7), pues su relación con grupos no indígenas ha sido distinta y ambas poblaciones han mantenido su propia estructura genética. En el caso de Abrojo la dificultad de acceso al territorio en contraste con la población de Limoncito ha afectado el grado de mezcla génica (Barrantes et al. 1982).
los niveles varían aún entre poblaciones y depende también de la ubicación de las localidades, siendo más susceptibles aquellas que se mantienen más alejadas de asentamientos no indígenas que además mantienen niveles menores de mezcla interétnica (Barrantes, 1993b) y bajos niveles de consumo de leche y sus derivados, debido al difícil acceso a los territorios que afecta la comercialización de lácteos. Los niveles de tolerancia estimados genotípicamente en los grupos indígenas concuerdan con los descritos previamente de acuerdo al fenotipo los cuales están entre el 70 al 90% (Montgomery et al. 1991; Swallow, 2003; Bloom & Sherman, 2005).
En suma, el descubrimiento del alelo T (SNP 3) para poblaciones del norte europeo con altos porcentajes del fenotipo tolerancia promovieron la formulación de hipótesis asociadas con selección y una de ellas propuso que la ventaja selectiva radicó en que aquellos individuos portadores del rasgo tolerancia en tales grupos se encontraron ventaja frente a sus enemigos al llevar el alimento consigo y teniéndolo disponible para movilizarse y enfrentar batallas en buen estado nutricional (Cochran & Hapending, 2009), sin embargo el genotipo europeo asociado con el rasgo (variante T, SNP 3) no ha sido útil para predecir tolerancia/intolerancia en otros grupos pastoriles que también presentan altos porcentajes del fenotipo tolerante; tal es el caso de grupos nómadas norasiáticos dentro de los cuales se encuentran los mongoles; estos además de presentar una población mayoritariamente tolerante han sido históricamente grupos guerreros que en gran parte basaron su alimentación en productos pastoriles como los lácteos (Sun et
al. 2007) . Lo anterior sugiere que otros grupos además de los europeos podrían
pasadas se haya señalado al grupo amerindio como un grupo con buenos índices de salud, mejores aún que los esperados para poblaciones pobres (Mata et al. 1985).
Por otra parte se analizaron los haplotipos de los cuales se espera una mayor profundidad del análisis de las variantes asociadas con el rasgo al tomar en cuenta las variantes vecinas genotipeadas. Dada la naturaleza poblacional y no familiar de los datos, los haplotipos sólo se pueden estimar mediante cálculos probabilísticos basados en las distancias físicas entre los marcadores, tasas de recombinación teóricas y el tamaño de la muestra. En este caso, la estimación de haplotipos generó un número de 55 haplotipos, de los cuales, la mayor diversidad haplotípica se presentó en las comunidades de Abrojo (ngöbe) y Rama Cay, seguido de la comunidad huetar de Zapatón (cuadro 5). Los grupos con menor diversidad fueron el grupo maleku y cabécar, que a su vez presentan bajas frecuencias de haplotipos asociados con tolerancia a la lactosa. Los valores de Fst (0.097) mostraron un grado alto de diferenciación entre las poblaciones si lo comparamos con valores de Fst obtenidos con otros marcadores genéticos (entre 0.055 y 0.072) (Barrantes, 1993b; Ruiz-Narváez et al. 2005, Wang et al. 2007), lo que sería esperado entre poblaciones que comparten una ancestría común reciente como es el caso.
aleatoriedad de la cinética de las frecuencias alélicas en diferentes grupos a nivel mundial (ver anexo 6).
La principal distinción de las poblaciones amerindias se presenta en las altas frecuencias de los haplotipos 2 y 4 en comparación con otras poblaciones mundiales (Ver anexo 4). La razón de este fenómeno sería la alta frecuencia de SNP 9 en su forma derivada (cuadro 4) lo cual es, hasta donde los datos de poblaciones publicados muestran, un fenómeno demostrado en grupos amerindios que puede ser explicado por deriva genética (Batista et al. 1988, Ruiz et al. 2005, Wang et al. 2007).
Los datos obtenidos muestran que la mayor variación haplotípica presente en grupos amerindios está asociada con la intolerancia a lactosa, siempre y cuando las asociaciones observadas en Europa y África sean válidas en amerindios. De ser así, existiría una alta probabilidad de que la mayoría de la población muestre síntomas de intolerancia al consumir productos lácteos y que ésta a su vez sea mayor en las poblaciones más alejadas de los centros urbanos de las cuales se esperaría menores grados de mezcla interétnica. La comprobación última de la posible intolerancia sería por medio de los métodos de medición directa o indirecta en las poblaciones indígenas, los cuales requerirían de la necesaria aplicación por personal médico calificado y centros de salud debidamente equipados, además de un genotipeo que incluyera otros marcadores genéticos informativos para definir el grado de mezcla por individuo. Aún así, los resultados aportados por este trabajo son consecuentes con los modelos explicativos existentes en cuanto a la distribución de la tolerancia a la lactosa y su relación con grupos pastoriles (Simoons, 1970; Feldman & Cavalli-Sforza, 1976; 1989, Durham, 1990).
sustituida por otro tipo de hábito como el consumo de leche resulta en perjuicio para la salud infantil (Barrantes, 1984). Aunque las comunidades indígenas se benefician de la política nutricional estatal que distribuye leche en polvo en la comunidades rurales, datos recientes revelan que el 57% de niños indígenas (incluyendo grupos menores de 6 años) y adolescentes sufren anemia debido a insuficiencias nutricionales (Scrimshaw, 2005). Esto indica que las políticas que han sido aplicadas por décadas aún no resuelven el problema de desnutrición infantil y por el contrario se ha agravado si comparamos con datos de 1984 en los cuales los niveles de nutrición infantil eran superiores (Mata et al. 1985). Los cuestionarios aplicados en las comunidades sugieren una percepción positiva sobre el consumo de leche (anexo 2), que podrían deberse a múltiples aspectos, tales como:
• Publicidad en los medios promoviendo el consumo de lácteos, principalmente como parte de las dietas infantiles, incrementando el interés por el consumo de leche y sus derivados.
• Aplicación de políticas estatales de manera sostenida en poblaciones vulnerables durante los últimos 60 años con el fin de mejorar el estado nutricional de los grupos (MIDEPLAN, 2006). El hecho de que algunos indígenas posean ganado vacuno y comercialicen su leche podría indicar que existen costumbres creadas por parte de las políticas estatales y con el paso del tiempo esas costumbres probablemente llegaron a traducirse en modo de subsistencia para los grupos (Anexo 2).
Considerando los procesos de multiculturalidad las políticas públicas deberían tomar en cuenta los diversos aspectos nutricionales antes de aplicar medidas que respectan a la salud y la nutrición (Holden & Mace, 1997). En la comunidad ngöbe de Limoncito se conocía la introducción de productos lácteos a través de cooperativas desde 1979, además de otros productos que señalan una occidentalización de su dieta; la cual podría repercutir en la salud de la comunidad como es el caso de la intolerancia a la lactosa (Mata et al. 1985), ya que al sufrir de los síntomas de la intolerancia se altera la absorción de nutrientes presentes en otros alimentos que se hayan consumido (Wiley, 2008).
7. Referencias Bibliográficas
Adolfsson O, Meydani S and Russel R (2004) Yogurt and gut function. Am J of Clin Nutr 80: 245-256.
Anderson B and Vullo C (1994) Did malaria select for primary adult lactase deficiency? Gut 35: 1487-1489.
Aoki K (1986) A stochastic model of gene-culture coevolution suggested by the "culture historical hypothesis" for the evolution of adult lactose absorption in humans. Proc Natl Acad Sci USA 83: 2929-2933.
Aoki K (2001) Theoretical and empirical aspects of gene-culture coevolution. Theor Popul Biol 59: 253-261.
Arias T (1991) Aproximación antropológica a algunos problemas biomédicos de las poblaciones amerindias. Hombre y Cultura 1: 99-114.
Azofeifa J, Ruiz E and Barrantes R (2001) Blood group, red cell, and serum protein varation in the Cabécar and Huetar, two chibchan amerindian tribes of Costa Rica
Bandelt HJ, Forster P and Röhl A (1999) Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol 16:37-48.
Barrantes R (1984) Variación de un modelo evolutivo poblacional en los amerindios de Costa Rica. Memorias del primer simposio científico sobre pueblos indígenas de Costa Rica: 163-168.
Barrantes R (1993a) Evolución en el trópico: Los amerindios de Costa Rica y Panamá. Editorial de la Universidad de Costa Rica, San José, 221 pp.
Barrantes R (1993b) Diversidad genética y mezcla racial en los amerindios de Costa rica y Panamá. Rev Biol Trop 41: 379-384.
Barrantes R and Mata L (1981) Estudios evolutivos y biomédicos en dos poblaciones indígenas Ngöbees de Costa Rica. Vínculos 7: 1-7.
Barrantes R and Morera B (1999) Contribución del genoma amerindio a la formación de la población costarricense. Vínculos 24: 85-96.
Barrantes R, Smouse P, Neel J, Mohrenweiser H and Gershowitz H (1982) Migration an genetic infraestructure of the central american Ngöbe and their affinitties with other tribal groups
Barrantes R, Smouse P, Mohrenweiser H, Gershowitz H, Azofeifa J, Arias T and Neel J (1990) Microevolution in Lower Central America: Genetic characterization of the chibchan-speaking groups of Costa Rica and Panama, and a consensus taxonomy based on genetic and linguistic affinity. Am J Hum Genet 46: 63-84.
Batista O, Kolman C, Arias T, Guionneau-Sinclair F, Quirós E and Bermingham (1988) Variación en el ADNmt de dos tribus amerindias chibchas, los ngöbe y cuna de Panamá. In: Bozzoli M, Barrantes R, Obando D and Rojas M (eds) Primer Congreso sobre Pueblos Indígenas de Costa Rica y sus Fronteras Memoria 15-35 pp.
Bersaglieri, T.; Sabeti, P.; Patterson, N.; Vanderploeg, T.; Schaffner, S.; Drake, J.; Rhodes, M.; Reich, D.; Hirschhorn, J (2004) Genetic signatures of strong recent positive selection at the lactase gene. Am J Hum Genet 74: 1111-1120.
Bloom G and Sherman P (2005) Dairying barriers affect the distribution of lactose malabsorption. Evol Hum Behav 26: 301.e1–301.e33.
Bodmer W and Cavalli-Sforza L (1976) Genetics, Evolution, and Man. W.H. Freeman and Company, San Francisco, pp 782.
Bolin T, Pirola C and Davis E (1969) Adaptation of intestinal lactase in rat. Gastroenterology 57: 406-409.
Boll W, Wagner P and Mantei N (1991) Structure of the chromosomal gene and cDNA coding for lactase-phlorizin hydrolase in humans with adult-type hypolactasia or persistence of lactase. Am J Hum Genet 48: 889-902.
Bozzoli and M Guevara M (2002) Los indígenas costarricenses en el siglo XXI. EUNED, San Jose, 76 pp.
Brenes W and Barrantes R (1984) Antropogenética dental y salud oral de cinco poblaciones amerindias de Costa Rica. Memorias del primer simposio científico sobre pueblos indígenas de Costa Rica 169-175p.
Brines J (2004) Adult lactose tolerance is not an advantageous evolutionary trait. Pediatrics 114:1372.
Burger J, Kirchner M, Bramanti B, Haak W and Thomas MG (2007) Absence of the lactase-persistence-associated allele in early Neolithic Europeans. Proc Natl Acad Sci USA 104: 3736-3741.
Calloway D and Murphy E (1968) The use of expired air to measure intestinal gas formation. Ann N Y Acad Sc 150: 82-95.
Cochran G and Harpending H (2009) The 10 000 Year Explosion. Basic Books, New York, 288 pp.
Coelho M, Luiselli D, Bertorelle G, Lopes AI, Seixas S, Destro-Bisol G and Rocha J (2005) Microsatellite variation and evolution of human lactase persistence. Hum Genet 117: 329–339
Constenla A (1991) Las lenguas del área intermedia: introducción a su estudio areal. Editorial de la Universidad de Costa Rica, San José, 221 pp.
Cook G and Howells G (1968) Lactosuria in the African with lactase deficiency. Am J Dig Dis 13: 634-637.
Cook GC and Al-Torki MT (1975) High intestinal lactase concentrations in adult Arabs in Saudi Arabia. Br Med J 3: 135-136.
Cooke R (1991) Paleoindian occupation in the Central American tropics. In: Bonnichsen R and Fladmark K (eds) Clovis: Origins and Human Adaptation. Peopling of the Americas. Oregon State University Press, Corvallis, pp 237-253.
Crow J (1986) Basic concepts in population. quantitative and evolutionary genetics. Freeman and Company, New York, 273 pp.
Cuatrecasas P, Lockwood DH and Caldwell JR (1965) Lactase deficiency in the adult: a common occurrence. Lancet I: 14-18.
Davidson A (1999) The oxford companion to food. Oxford University Press, New York, 907 p.
De Vrese M, Stegelmann A, Richter B, Fenselau S, Laue C, Schrezenmeir J (2001) Probiotics-compensation for lactase insufficiency. Am J Clin Nut 73:421S–429S.
Dunphy J, Littman J, Hammond G, Forstner A, Dahlqvist D and Crane R (1965) Intestinal lactase deficit in adults. Gastroenterology 49: 12-21.
Durham W (1991) Coevolution, Genes, Culture and Human Diversity. Stanforf University Press, California, 629 pp.
Dubowski KM (1962) An ortho-toluidine method for body fluid glucose determinations. Clin Chem 8: 215.
Enattah N, Sahi. T, Savilahti E, Terwilliger J, Peltonen L and Järvelä I (2002) Identification of a variant associated with adult-type hypolactasia. Nat Genet 30: 233-237.
Enattah N (2005) Molecular genetics of lactase persistence. PhD Dissertation: Department of Molecular Medicine. National Public Health Institute, Helsinki, Finland and Department of Medical Genetics, Faculty of Medicine, University of Helsinki, Finland, 99 pp.
Verschubskaya G, Comas D, Bulayeva K, Mehdi S, Terwilliger J, Sahi T, Savilahti E, Perola P, Sajantila A, Jarvela I and Peltonen L (2007) Evidence of still-ongoing convergence evolution of the lactase persistence T-13910 Alleles in Humans. Am J Hum Genet 81: 615- 625.
Enattah E, Jensen T, Nielsen M, Lewinski R, Kuokkanen M, Rasinpera H, El-Shanti H, Kee Seo J, Alifrangis M, Khalil I, Natah A, Ali A, Natah S, Comas D, Mehdi Q, Groop L, Vestergaard E, Imtiaz F, Rashed M, Meyer B, Troelsen J and Peltonen L (2008) Independent introduction of two lactase-persistence alleles into human populations reflects different history of adaptation to milk culture. Am J Hum Genet 82: 57–72.
Excoffier L, Laval G and Schneider S (2005) Arlequin ver. 3.1: An integrated software package for population genetics data analysis. Evol Bioinform Online 1:47-50.
Flatz G and Rotthauwe HW (1973) Lactose nutrition and natural selection. Lancet 2: 76-77.
Feldman M and Cavalli-Sforza L (1976) Cultural and biological proceses: selection for a trait under complex transmisión. Theor Popul Biol 9: 238-259.
Feldman M and Cavalli-Sforza L (1989) On the theory of evolution under genetic and cultural trasmission with application to the lactose absortion problem. In: Feldman MW (ed) Mathematical Evolutionary Theory, Princeton University Press, 341 pp.
Gabb W (1981) Talamanca; el espacio y los hombres. 2nd edition. EUNED, San José, Costa Rica, 168 pp.