Group-II phospholipase A
2enhances oxidized low density
lipoprotein-induced macrophage growth through enhancement of
GM-CSF release
Kengo Kaneko, Masakazu Sakai *, Takeshi Matsumura, Takeshi Biwa,
Noboru Furukawa, Tetsuya Shirotani, Shinsuke Kiritoshi, Yoshichika Anami,
Kohji Matsuda, Takayuki Sasahara, Motoaki Shichiri
Department of Metabolic Medicine,Kumamoto Uni6ersity School of Medicine,1-1-1Honjo,Kumamoto860-8556,Japan
Received 26 July 1999; received in revised form 19 November 1999; accepted 19 January 2000
Abstract
Inflammatory process plays an important role in the development and progression of atherosclerotic lesions. Recently, group-II phospholipase A2(PLA2), an inflammatory mediator, was reported to exist in human atherosclerotic lesions and to enhance the
development of murine atherosclerotic lesions. Oxidized low density lipoprotein (Ox-LDL) stimulates the growth of several types of macrophages in vitro. Since proliferation of macrophages occurs in atherosclerotic lesions, it is possible to assume that the Ox-LDL-induced macrophage proliferation might be involved in the progression of atherosclerosis. In this study, the role of group-II PLA2 in the Ox-LDL-induced macrophage growth was investigated using thioglycollate-elicited mouse peritoneal
macrophages. Thioglycollate-elicited macrophages significantly expressed group-II PLA2and released it into the culture medium.
The Ox-LDL-induced thymidine incorporation into thioglycollate-elicited macrophages was three times higher than that into resident macrophages, whereas under the same conditions, granulocyte/macrophage colony-stimulating factor (GM-CSF) equally induced thymidine incorporation into both types of macrophages. Moreover, the Ox-LDL-induced GM-CSF release from thioglycollate-elicited macrophages was significantly higher than that from resident macrophages. In addition, the Ox-LDL-in-duced thymidine incorporation into macrophages obtained from human group-II PLA2transgenic mice and the GM-CSF release
from these cells were significantly higher than those from their negative littermates, and the Ox-LDL-induced thymidine incorporation into human group-II PLA2transgenic macrophages was significantly inhibited by a polyclonal anti-human group-II
PLA2 antibody. These results suggest that the expression of group-II PLA2in thioglycollate-elicited macrophages may play an
enhancing role in the Ox-LDL-induced macrophage growth through the enhancement of the GM-CSF release. © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Oxidized LDL; Macrophage growth; Atherosclerosis; Inflammation; Phospholipase A2; GM-CSF
www.elsevier.com/locate/atherosclerosis
1. Introduction
The presence of a massive cluster of macrophage-derived foam cells in the subendothelial spaces of
arter-ies is one of the characteristic features of the early stages of atherosclerotic lesions [1]. Macrophages take up chemically modified low density lipoproteins (LDL), such as oxidized LDL (Ox-LDL) and acetylated LDL (Ac-LDL), through the scavenger receptor pathways and become foam cells in vitro [2 – 4]. Macrophage-derived foam cells are believed to play an important role in the development and progression of atheroscle-rosis through the production of various cytokines and growth factors [1]. Previous studies have reported that macrophages and macrophage-derived foam cells pro-liferate in atherosclerotic lesions [5 – 7]. Recent studies Abbre6iations:GM-CSF, granulocyte/macrophage
colony-stimulat-ing factor; LDL, low density lipoprotein; Ox-LDL, oxidized LDL; PKC, protein kinase C; PLA2, phospholipase A2; WHHL, Watanabe
heritable hyperlipidemic; ELISA, enzyme-linked immunosorbent as-say.
* Corresponding author. Tel.: +81-96-3735169; fax: + 81-96-3668397.
E-mail address:[email protected] (M. Sakai).
from the laboratory as well as by other investigators [8 – 17] have demonstrated that Ox-LDL could induce macrophage growth in vitro. Based on these findings, it was postulated that Ox-LDL might be involved in macrophage growth in vivo and linked to the progres-sion of atherosclerotic process.
One of the pathological and immunohistochemical features of atherosclerotic lesions is accumulation of inflammatory cells and cytokines [1]. Moreover, dexam-ethasone, an anti-inflammatory agent, is known to ex-hibit an anti-atherogenic effect in animal models of experimental atherosclerosis, such as Watanabe herita-ble hyperlipidemic (WHHL) rabbit [18], cholesterol-fed rabbit [19] and rat balloon angioplasty model [20]. Based on these studies, it seems reasonable to consider atherosclerosis as a chronic inflammatory disease, or that the inflammatory process plays an important role in the development and progression of atherosclerosis. Moreover, recent reports using human group-II phos-pholipase A2 (PLA2) transgenic mouse demonstrated that group-II PLA2, one of the inflammatory media-tors, played an enhancing role in the development of atherosclerotic lesions [21,22]. Thus, to further elucidate the pathophysiological significance of macrophage growth in atherosclerosis, it seems reasonable to investi-gate the growth promoting effect of Ox-LDL on inflam-matory macrophages, and the effect of group-II PLA2 on the Ox-LDL-induced macrophage growth. Here, the growth stimulating effects of Ox-LDL on the thiogly-collate-induced non-infectious inflammatory mouse peritoneal macrophages was examined. The results demonstrated that the responsiveness of thioglycollate-elicited macrophages to the growth-stimulating activity of Ox-LDL was significantly greater than that of resi-dent macrophages, and the expression of group-II PLA2 might play, at least in part, an enhancing role in the growth of thioglycollate-elicited macrophage through the enhancement of granulocyte/macrophage colony-stimulating factor (GM-CSF) release.
2. Methods
2.1. Chemicals
[methyl-3H]Thymidine and [1-14C]oleic acid were pur-chased from NEN Life Science (Boston, MA). A rabbit polyclonal anti-human group-II PLA2 antibody was purchased from Funakoshi (Tokyo, Japan). Other chemicals were of the highest grade available from commercial sources.
2.2. Cell culture
Peritoneal cells were collected from non-stimulated male C3H/He mice (25 – 30 g) (Japan SLC,
Hama-matsu, Japan) and commercially available human group-II PLA2transgenic mice and their negative litter-mates, C57BL/6J strain [23] (Taconic Farms, NY). Serum PLA2activity in transgenic mice was reported to be 8 – 10-fold greater than that in non-transgenic mice [23]. The preliminary experiment using RT-PCR showed the expression of human group-II PLA2 mRNA in PLA2-transgenic macrophages but not in their negative littermates. Kennedy et al. [24] demon-strated that some murine strain had natural null muta-tion for group-II PLA2, such as 129/Sv, B10.RIII and C57BL/6J, but C3H/He produced functional group-II PLA2. Thus, C3H/He mice were used in the present study for determination of the effect of group-II PLA2 on the macrophage growth. Thioglycollate-elicited macrophages were collected from C3H/He mice 5 days after intraperitoneal injection of 1.5 ml thioglycollate [25]. The collected peritoneal cells were suspended in RPMI 1640 medium (Nissui Seiyaku, Tokyo) supple-mented with 10% heat-inactivated fetal bovine serum (Life Technologies), streptomycin (0.1 mg/ml), and penicillin (100 U/ml) (medium A). After 90 min incuba-tion at 37°C, non-adherent cells were removed by tripli-cate washing with prewarmed medium A. After washing, cell numbers of resident and elicited macrophages were decreased to 70%. Macrophages were identified by the following features: (i) adherence to culture plates; (ii) morphological features resembling mononuclear cells after Giemsa staining; (iii) the capac-ity to take up carbon particles; and (iv) positive im-munohistochemistry with antibody for CD68.
2.3. Tritiated thymidine incorporation assay
pore size) and washed three times with 1 ml of 99.5% ethanol. The filters were dried under air and their radioactivity was counted in a liquid scintillation coun-ter [13].
2.4. Cell-counting assay
Peritoneal cells were adjusted to 2×104cells/ml, and 1 ml of cell suspension was dispersed in each well of 24-well tissue culture plates (16 mm in diameter, Fal-con) and incubated for 90 min at 37°C. Non-adherent cells were removed by triplicate washing with 1 ml of prewarmed medium A. The macrophage monolayers thus formed were cultured at 37°C in 1 ml of medium A, with or without the test lipoproteins. After incubation for 7 days, adherent cells in triplicate wells were lysed in 1% (w/v) Triton X-100, and the number of naphthol blue – black-stained nuclei was counted in a hemocytometer, as described previously [8].
2.5. Lipoproteins
Human LDL (d=1.019 – 1.063 g/ml) was isolated by sequential ultracentrifugation of plasma samples ob-tained from consented normolipidemic subjects after overnight fasting [26]. LDL was dialyzed against 0.15 M NaCl and 1 mM EDTA, pH 7.4. Ox-LDL was prepared by incubation of 0.1 mg/ml of LDL in PBS with 5 mM CuSO4 for 20 h at 37°C followed by the addition of 1 mM EDTA and cooling [27]. Acetyl-LDL was prepared by chemical modifica-tion of LDL with acetic anhydride as described previ-ously [28]. The level of endotoxin associated with these lipoproteins was B1 pg/mg protein as measured by a commercially available kit (Toxicolor system, Seika-gaku, Tokyo). Moreover, macrophage growth and vi-ability were not affected by endotoxin at a concentration below 1 ng/ml in the experimental sys-tem.
2.6. RT-PCR analysis for mouse group-II PLA2
Standard molecular biological techniques were used [29]. Macrophages (2×106 cells) were dispersed into each well (3.5 cm in diameter, Nunc) and incubated for 90 min. After washing with medium A, macrophage monolayers thus formed in 2 ml of medium A for 6 h, and then total RNA was extracted with TRIzol (Life Technologies). The first strand cDNA synthesis con-taining 1 mg of total RNA was primed with oligo dT. Primers used for PCR amplification of group-II PLA2 and b-actin were designed on the basis of murine group-II PLA2 cDNA [30] and murine b-actin cDNA [31] sequences as follows: for group-II PLA2, forward primer; TTC TGG CAG TTC CAG AGG ATG G
(nucleotide 241 – 262 of murine group-II PLA2 coding sequence); reverse primer; AAG ACA CTC CCT AGA CAG CAA (nucleotide 678 – 709 of murine group-II PLA2 coding sequence) [30]; for b-actin, forward primer; GTG GGC CGC TCT AGG CAC CAA (nu-cleotide 25 – 45 of murine b-actin coding sequence); reverse primer; CTC TTT GAT GTC ACG CAC GAT TTC (nucleotide 541 – 564 of murine b-actin coding sequence) [31]. The cycling conditions in the GeneAmp 9600 System consisted of a first step of 94°C denaturation for 10 min, followed by 30 cycles of annealing at 54°C for 60 s, extension at 75°C for 90 s, and denaturation at 94°C for 30 s, with a final elongation step at 75°C for 10 min. Amplification products were analyzed by 1.5% agarose gel elec-trophoresis. To verify that amplification products were consistent with the reported sequences of murine group-II PLA2 and b-actin, they were ligated into pGEM-T (Promega, Madison, WI), transfected into Escherichia coli XL1-Blue and sequenced by using 373A DNA sequencer (Applied Biosystems, Foster City, CA).
2.7. Enzyme-linked immunosorbent assay (ELISA) for GM-CSF
Macrophages (5×106
cells) were dispersed into each well (10 cm in diameter, Falcon) and incubated for 90 min. After washing with medium A, macrophage monolayers thus formed were cultured in 15 ml of medium A with or without the lipoproteins to be tested for the indicated times, and then 300ml of the medium were collected. The concentration of GM-CSF protein was determined according to the instructions provided by the manufacturer of mouse GM-CSF-specific ELISA system (Amersham) using recombinant murine GM-CSF as a standard [13].
2.8. PLA2 acti6ity
PLA2 activity was determined using [1-14
C]oleate-la-beled E. coli phospholipid as a substrate, as described previously [32]. Assay mixtures contained 100 mM Tris – HCl (pH 7.4), 1 mM CaCl2, 2×10
8 cells of
[1-14C]oleate-labeledE.coli and enzyme or conditioned serum-free medium. Reaction mixtures were incubated for 1 h at 37°C. The reaction was terminated by the addition of 5 ml propane-2-ol/n-heptane/1 M H2SO4 (40/10/1, by vol.), followed by 2 ml heptane and 3 ml water. After vortex and phase separation, an aliquot (2.5 ml) of the upper phase was passed over a column of silic acid. Free [1-14C]oleate was quantitatively elute with 1 ml ethyl acetate. Radioactivity was determined in a liquid scintillation counter. PLA2 activity was expressed as [1-14
2.9. Miscellaneous
Data were expressed as mean9S.D. Differences be-tween groups were compared for statistical significance using the Student’st-test. A probability value less than 5% was considered significant. The experimental proto-col was approved by the Human Ethics Review Com-mittee and the Ethics Review ComCom-mittee for Animal Experimentation of the institution.
3. Results
3.1. Thioglycollate-elicited macrophages express group-II PLA2
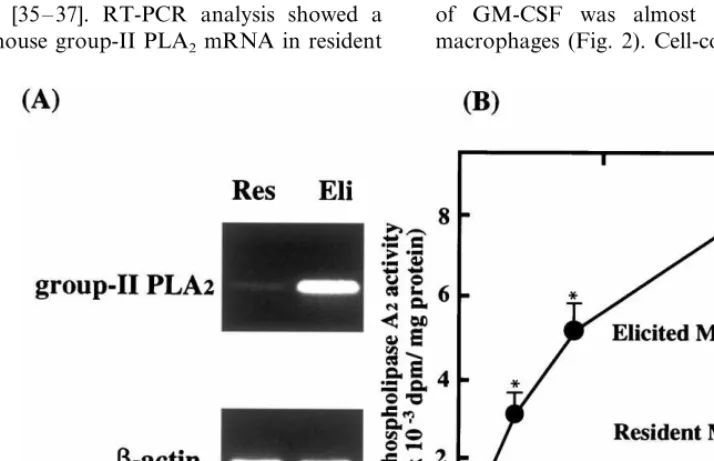
Various inflammatory bioactive molecules are secreted from cells in atherosclerotic lesions [1]. Among them, group-II PLA2 as a candidate for enhancing the Ox-LDL-induced macrophage growth was focused on for the following reasons: (i) previous studies reported the presence of group-II PLA2in human atherosclerotic lesions [33,34], which played an enhancing role in mouse atherosclerotic lesions [21,22]; and (ii) the pres-ence of group-II PLA2in peritoneal exudate induced by various stimuli [35 – 37]. RT-PCR analysis showed a faint band of mouse group-II PLA2 mRNA in resident
macrophages, whereas a clear band of mouse group-II PLA2 mRNA was present in thioglycollate-elicited macrophages (Fig. 1A). Next PLA2 activity in the macrophage conditioned medium was examined. Fig. 1B shows a significantly high PLA2 activity in the conditioned medium containing thioglycollate-elicited macrophages compared to that of resident macrophages. These results suggested that thioglycol-late-elicited macrophages produced a significantly higher amount of group-II PLA2into the medium than resident macrophages.
3.2. Ox-LDL stimulates growth of thioglycollate-elicited macrophages
The effects of Ox-LDL on the growth of mouse resident macrophages and thioglycollate-elicited macrophages were examined. Fig. 2A shows that thymidine incorporation into resident macrophages was induced by Ox-LDL in a dose-dependent manner, but neither by LDL nor acetyl-LDL. Fig. 2B shows that Ox-LDL also induced thymidine incorporation into thioglycollate-elicited macrophages, which was three times higher than that of resident macrophages (Fig. 2). As a control, thymidine incorporation induced by 1 nM of GM-CSF was almost equal in both types of macrophages (Fig. 2). Cell-counting assay also showed
Fig. 1. Polymerase chain reaction (PCR) analysis of mouse group-II phospholipase A2(PLA2) mRNA (A) and PLA2activity in medium (B). (A)
Mouse peritoneal macrophages (2×106cells) were seeded in 3.5 cm dish and incubated for 90 min. Non-adherent cells were removed by washing
with medium A. The cell monolayers thus formed were incubated in 2 ml of medium A alone for 1 h. After incubation, total RNA was extracted from each dish with TRIzol. The expression of mRNA for mouse group-II PLA2(upper panel) orb-actin (lower panel) was evaluated by RT-PCR
as described in Section 2. (B) Mouse peritoneal resident macrophages () or thioglycollate-elicited macrophages ( ) (1×107cells) were seeded
in 10 cm dish and incubated for 90 min. After washing with medium A, the cell monolayers thus formed were incubated at 37°C for 18 h in 10 ml of RPMI 1640 with 3% bovine serum albumin (BSA). After incubation, the conditioned medium was collected and PLA2 activity was
Fig. 2. Oxidized low density lipoprotein (Ox-LDL)-induced growth of resident macrophages (A) and thioglycollate-elicited macrophages (B). Mouse peritoneal macrophages (4×104cells) were dispersed in
each well, and incubated for 90 min. Non-adherent cells were re-moved by washing with medium A. The cell monolayers thus formed were incubated for 6 days in 1 ml of medium A with the indicated concentrations of Ox-LDL (), acetyl-LDL () or LDL (). As a control experiment, cells were incubated with 1 nM of recombinant granulocyte/macrophage colony-stimulating factor (GM-CSF). Eigh-teen hours before the termination of experiments, cells were chased with [3H]thymidine, harvested and radioactivity was determined as
described under Section 2. Data represent the mean9S.D. of four separate experiments. *PB0.01 compared to medium alone, †PB
0.005 compared to medium alone by Student’st-test.
Fig. 3. Effect of oxidized low density lipoprotein (Ox-LDL) on [3H]thymidine incorporation into macrophages derived from human
group-II phospholipase A2 (PLA2) transgenic mice. Mouse resident
peritoneal macrophages from human group-II PLA2transgenic mice
or their negative littermates (4×104cells) were dispersed in 24 well
plates, and incubated for 90 min. After washing, cell monolayers thus formed were incubated at 37°C for 6 days in 1 ml of medium A with the indicated concentrations of Ox-LDL (), acetyl-LDL (), LDL () or 1 nM of recombinant mouse granulocyte/macrophage colony-stimulating factor (GM-CSF) as a control. Thymidine incorporation was determined as described in Section 2. Data represent the mean9
SD of three separate experiments. *PB0.01 compared to medium alone by Student’st-test.
ulating activity of Ox-LDL was significantly greater than that of resident macrophages.
3.3. Group-II PLA2 enhances macrophage growth
To determine the role of group-II PLA2 in the LDL-induced macrophage growth, the effect of Ox-LDL on the growth of macrophages obtained from human group-II PLA2 transgenic mice was examined. Fig. 3 shows that the Ox-LDL-induced thymidine in-that the number of resident macrophages increased 1.8
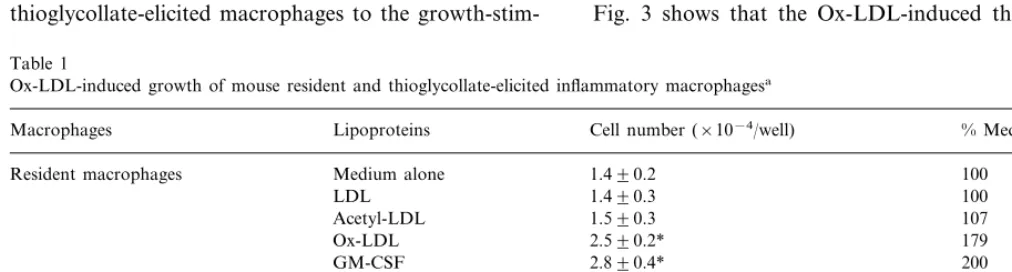
times by Ox-LDL compared to non-loaded resident macrophages, whereas the number of thioglycollate-elicited macrophages was significantly increased 2.3 times by Ox-LDL (Table 1). These results demonstrated that the growth of thioglycollate-elicited macrophages was also induced by Ox-LDL and the responsiveness of thioglycollate-elicited macrophages to the
growth-stim-Table 1
Ox-LDL-induced growth of mouse resident and thioglycollate-elicited inflammatory macrophagesa
Macrophages Lipoproteins Cell number (×10−4/well) % Medium alone
100 Resident macrophages Medium alone 1.490.2
100 1.490.3
LDL
107 1.590.3
Acetyl-LDL
179
Ox-LDL 2.590.2*
GM-CSF 2.890.4* 200
Medium alone
Elicited macrophages 1.590.2 100
107
LDL 1.690.4
Acetyl-LDL 1.890.3 127
227 3.490.2**
Ox-LDL
3.090.3**
GM-CSF 200
aPeritoneal macrophages (2×104cells) were dispersed into culture plates, and incubated for 90 min. After incubation, non-adherent cells were
removed by triplicate washing with medium A. At the start of experiment, cell numbers of resident and elicited macrophages were 1.4 and 1.5×104
cells/well, respectively. Macrophage monolayers thus formed were incubated for 7 days in 1 ml of medium A with 20mg/ml of lipoproteins or 1 nM of GM-CSF as a control. After incubation, counting of solubilized nuclei was performed as described under Section 2. Data are expressed as mean9S.D. of quadriplicate counts. LDL, low density lipoprotein; Ox-LDL, oxidized low density lipoprotein; GM-CSF, granulocyte/ macrophage colony-stimulating factor.
*PB0.01, compared to medium alone.
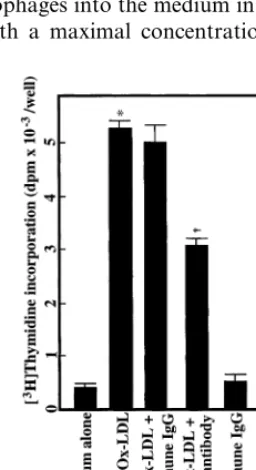
corporation into macrophages from human group-II PLA2 transgenic mice was twice that from their nega-tive littermates. Moreover, the Ox-LDL-induced thymidine incorporation into macrophages from human group-II PLA2 transgenic mice was significantly but partially inhibited by 50% by a polyclonal anti-human group-II PLA2 antibody as compared to that by non-immune IgG (Fig. 4). These results suggested that group-II PLA2 might enhance, at least in part, the Ox-LDL-induced macrophage growth.
3.4. Ox-LDL augments GM-CSF release
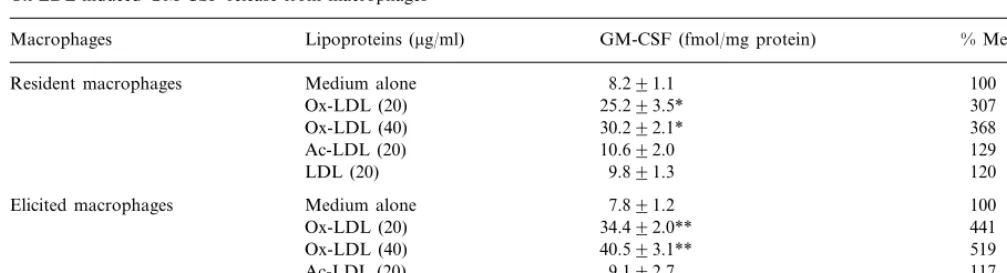
It has recently been reported that GM-CSF played a priming role in the Ox-LDL-induced macrophage growth [13]. Thus, the production of GM-CSF from thioglycollate-elicited macrophages was next examined. When resident macrophages were incubated with medium alone for 6 h, GM-CSF was released at con-centration of 8.2 fM/mg protein (Table 2). Ox-LDL significantly enhanced the GM-CSF release from resi-dent macrophages into the medium in a dose-depenresi-dent manner with a maximal concentration of 30.2 fM/mg
protein at 40mg/ml of Ox-LDL (Table 2). On the other hand, when thioglycollate-elicited macrophages were incubated with medium alone, the concentration of GM-CSF in the medium was 7.8 fM/mg protein (Table 2). However, the GM-CSF release from thioglycollate-elicited macrophages was increased by Ox-LDL with maximal concentration of 40.5 fM/mg protein, which was significantly higher than that from resident macrophages induced by Ox-LDL (Table 2). In con-trast, LDL or acetyl-LDL could not enhance the GM-CSF release from both types of macrophages (Table 2). Combined with the results of macrophage growth (Fig. 2) and PLA2 activity (Fig. 1), these results suggested that PLA2might enhance the effect of Ox-LDL on the GM-CSF release as well as macrophage growth. To confirm this conclusion, the effect of Ox-LDL on the GM-CSF release from human group-II PLA2 trans-genic macrophages was examined. As shown in Table 3, the Ox-LDL-induced GM-CSF release from macrophages obtained from wild type littermates was 2.1 times greater than that from wild cells which were incubated with medium alone. Consistent with macrophage growth (Fig. 4), the Ox-LDL-induced GM-CSF release from human group-II PLA2 trans-genic macrophages was significantly greater than that from wild macrophages (Table 3). These results sug-gested that group-II PLA2 enhances GM-CSF release from thioglycollate-elicited macrophages, thereby en-hancing the Ox-LDL-induced macrophage growth.
4. Discussion
Recent studies from the laboratory as well as by others have demonstrated that Ox-LDL could induce the growth of several types of macrophages in vitro [8 – 17], whereas the importance of in situ macrophage growth during the development of atherosclerotic le-sions is still unknown. It is possible to assume that the Ox-LDL-induced macrophage growth is involved in the development and progression of atherosclerotic lesions because: (i) macrophages and macrophage-derived foam cells are believed to play an essential role in the development and progression of atherosclerosis via pro-duction of various active molecules [1]; and (ii) prolifer-ation of macrophages has been observed in atherosclerotic lesions [2 – 4].
Recent immunohistochemical and pathological stud-ies suggested that the inflammatory process was in-volved in the development and progression of atherosclerosis [1,18 – 20]. Moreover, recent reports us-ing human group-II PLA2 transgenic mouse demon-strated that group-II PLA2, an inflammatory mediator, played an enhancing role in the development of atherosclerotic lesions [21,22]. Thus, it seems reasonable to investigate the growth promoting effects of Ox-LDL Fig. 4. Effect of an anti-human group-II phospholipase A2 (PLA2)
antibody on oxidized low density lipoprotein (Ox-LDL)-induced [3H]thymidine incorporation into macrophages derived from human
group-II PLA2 transgenic mice. Mouse resident peritoneal
macrophages from human group-II PLA2 transgenic mice (4×104
cells) were dispersed in 24 well plates, and incubated for 90 min. After washing with medium A, cell monolayers thus formed were incubated at 37°C for 6 days in 1 ml of medium A with 20mg/ml Ox-LDL in the presence or absence of 10 mg/ml of anti-human group-II PLA2
Table 2
Ox-LDL-induced GM-CSF release from macrophagesa
GM-CSF (fmol/mg protein)
Macrophages Lipoproteins (mg/ml) % Medium alone
Medium alone
Resident macrophages 8.291.1 100
25.293.5*
Ox-LDL (20) 307
30.292.1* 368
Ox-LDL (40)
10.692.0
Ac-LDL (20) 129
LDL (20) 9.891.3 120
7.891.2
Elicited macrophages Medium alone 100
34.492.0** 441
Ox-LDL (20)
40.593.1**
Ox-LDL (40) 519
Ac-LDL (20) 9.192.7 117
7.791.2 99
LDL (20)
aPeritoneal macrophages (2×106cells) were dispersed into culture plates, and incubated for 90 min. After incubation, non-adherent cells were
removed by triplicate washing with medium A. Macrophage monolayers thus formed were incubated for 6 h in 1 ml of medium A with the indicated concentrations of lipoproteins. After incubation, culture medium were taken and the levels of GM-CSF was determined by ELISA as described under Section 2. Contents of cellular proteins were determined using BCA reagent (Pierce). Data are expressed as mean9S.D. of quadriplicate. Ox-LDL, oxidized low density lipoprotein; GM-CSF, granulocyte/macrophage colony-stimulating factor.
*PB0.01, compared to non-loaded resident macrophages.
**PB0.01, compared to non-loaded elicited macrophages by Student’st-test.
on inflammatory macrophages. Jutila and Banks [38] reported that the thymidine incorporation into non-stimulated thioglycollate-elicited inflammatory macrophage was greater than that into non-stimulated resident macrophages, whereas it was unclear whether Ox-LDL could induce the growth of thioglycollate-elic-ited macrophages. The present study demonstrated that Ox-LDL induced the growth of thioglycollate-elicited macrophages, which was significantly higher than the Ox-LDL-induced growth of resident macrophages (Fig. 2 and Table 1).
The results showed that the Ox-LDL-induced growth of thioglycollate-elicited macrophages was significantly greater than that of resident macrophages (Fig. 2 and Table 1), and the expression of group-II PLA2 might play, at least in part, an enhancing role in this phe-nomenon (Fig. 1). This conclusion was supported by the following findings: (i) the Ox-LDL-induced thymidine incorporation into macrophages from human group-II PLA2 transgenic mice was significantly higher than that from their negative littermates (Fig. 3); and (ii) such incorporation was significantly but partially inhibited by a polyclonal anti-human group-II PLA2 antibody (Fig. 4). The presence of group-II PLA2 has been observed in local and systemic inflammatory con-ditions, such as peritonitis, rheumatoid arthritis and septic shock [39,40]. Group-II PLA2plays an enhancing role in inflammation via production of lysophospho-lipids and release of arachidonic acid, a precursor of prostaglandins [39]. Recent immunohistochemical stud-ies demonstrated that group-II PLA2 was also ex-pressed in atherosclerotic lesions [32,33], and its expression was correlated with the degree of atheroscle-rosis [41]. This enzyme is mainly associated with vascu-lar smooth muscle cells in the normal media and intima
of human atherosclerotic arterial wall, which is consis-tent with the results demonstrating the source of group-II PLA2 in other human tissues [42,43]. Moreover, group-II PLA2 in human atherosclerotic lesions is known to be associated with macrophages and lipid core lesions [44]. Interestingly, in addition to atheroscle-rotic lesions, macrophage proliferation has also been observed in tumors and inflamed lung [45 – 48], where the expression of group-II PLA2has been reported [40]. Thus, it is possible that group-II PLA2 may enhance macrophage proliferation by certain stimuli in various tissues. In the present study, the enhancing effect of group-II PLA2 on the Ox-LDL-induced macrophage growth has been focused on. It should be noted, how-ever, that various active molecules are expressed in atherosclerotic lesions apart from group-II PLA2 [1]. Thus, further studies are necessary to examine whether other active molecules enhance the Ox-LDL-induced growth of thioglycollate-elicited macrophages.
group-II PLA2 did not induce macrophage growth di-rectly, and that it exhibited a growth enhancing activity for macrophages via phospholipid metabolites, such as lysophospholipids, arachidonate itself or its metabo-lites. Since it was previously reported that receptor-me-diated endocytosis of lysophosphatidylcholine in Ox-LDL through the scavenger receptors played an important role in the Ox-LDL-induced macrophage growth [8], it was possible that PLA2 might increase in lysophosphatidylcholine content in Ox-LDL, thereby enhancing the Ox-LDL-induced macrophage growth. However, a significant change of lysophosphatidyl-choline content in the conditioned medium could not be detected, in spite of an increase in PLA2 activity. The exact reason of these apparently inconsistent results remains unclear, but it is possible to assume that during incubation with macrophages, lysophosphatidylcholine hydrolyzed from phosphatidylcholine by PLA2 may be further reduced by other enzymes to glycerolphosphate or other metabolites, or may receive re-acylation to phosphatidylcholine in the cell culture system. There-fore, the change of lysophosphatidylcholine contents in the conditioned medium after incubation with macrophages might not be detected. On the other hand, Martens et al. [16] recently demonstrated that oxidized arachidonate might modify apoB and then induce macrophage growth. Moreover, it was reported that free fatty acids, another part of phosphatidylcholine metabolite by PLA2, in LDL were oxidized by cells and enhanced further oxidation of LDL [51]. These findings suggested that arachidonate or its oxidatively modified metabolite(s) might enhance the Ox-LDL-induced growth of thioglycollate-elicited macrophages. How-ever, the precise mechanisms of action are still un-known at present and further studies are needed to elucidate PLA2-mediated enhancement of the Ox-LDL-induced macrophage growth.
In the present study, the Ox-LDL-induced thioglycol-late-elicited macrophage growth, PLA2 activity in con-ditioned medium and GM-CSF release from these cells
were all significantly greater than those of resident macrophages (Figs. 1 and 2, and Table 2). Moreover, the Ox-LDL-induced growth of human group-II PLA2 transgenic macrophages and the GM-CSF release from these cells were both significantly greater than those from wild macrophages (Fig. 3 and Table 3). Further-more, both elicited and resident macrophages were equal in their response to the growth-promoting activ-ity of recombinant GM-CSF (Fig. 2). These results suggested that group-II PLA2 promotes the Ox-LDL-induced macrophage growth through enhancement of the GM-CSF release. A recent study demonstrated that dexamethasone inhibited the Ox-LDL-induced macrophage growth which was explained by the inhibi-tion of the GM-CSF expression [52]. Moreover, dexam-ethasone was reported to inhibit the expression of group-II PLA2 in rat macrophages [53]. These findings also supported the conclusion that group-II PLA2 might enhance the expression of GM-CSF. However, the enhancing mechanism of group-II PLA2 for the GM-CSF release from macrophages remains unclear at present. It was previously demonstrated that a rise in intracellular calcium and a subsequent increase in PKC activity were involved in the Ox-LDL-induced GM-CSF release and the subsequent macrophage growth [14]. Moreover, Martens et al. [15] recently reported that phosphatidylinositol 3-kinase (PI3K) was involved in the Ox-LDL-induced macrophage growth. Further studies are necessary to elucidate the mechanism of the group-II PLA2-mediated enhancement of GM-CSF re-lease and the subsequent macrophage growth.
It was previously demonstrated that PLA2-treated acetyl-LDL significantly induced macrophage growth [8]. However, when elicited-macrophages or human PLA2-transgenic macrophages, which released a signifi-cant amount of group-II PLA2, were incubated with acetyl-LDL, macrophage growth did not observed (Figs. 2 and 3). These results were apparently inconsis-tent. When macrophages were incubated with acetyl-LDL, acetyl-LDL bound to the scavenger receptors and
Table 3
Ox-LDL-induced GM-CSF release from human group-II PLA2 transgenic macrophagesa
Macrophages Ox-LDL (mg/ml) GM-CSF (fmol/mg protein) % Medium alone
0 7.891.4 100
Wild macrophages
40 16.592.8* 212
5.691.3 100
PLA2-transgenic macrophages 0
20.992.2** 373
40
aPeritoneal macrophages (2×106cells) were dispersed into culture plates, and incubated for 90 min. After incubation non-adherent cells were
removed by triplicate washing with medium A. Macrophage monolayers thus formed were incubated for 6 h in 1 ml of medium A with indicated concentrations of lipoproteins. After incubation, culture medium were taken and the levels of GM-CSF was determined by ELISA as described under Section 2. Contents of cellular proteins were determined using BCA reagent (Pierce). Data are expressed as mean9S.D. of triplicate. Ox-LDL, oxidized low density lipoprotein; GM-CSF, granulocyte/macrophage colony-stimulating factor; PLA2, phospholipase A2.
*PB0.01, compared to non-loaded wild macrophages.
was endocytosed by macrophages immediately. More-over, since the medium used in the present study con-tained 10% fetal calf serum (FCS), a large amount of phosphatidylcholine might prevent hydrolysis of phos-phatidylcholine in acetyl-LDL. Thus, it is possible to assume that if a small amount of phosphatidylcholine would be hydrolyzed to lysophosphatidylcholine in ace-tyl-LDL by the action of PLA2, it might not be enough for the induction of macrophage growth.
Based on the finding that Ox-LDL significantly in-duced the growth of peritoneal cells obtained from M-CSF and GM-CSF double knockout mouse, Hamil-ton et al. [17] proposed that the GM-CSF release from macrophages was not required for the Ox-LDL-induced macrophage growth. Since both M-CSF and GM-CSF are well known growth factors regulating the differenti-ation and the proliferdifferenti-ation of monocytes/macrophages, it is possible to assume that peritoneal cells from double knockout mice could represent immature macrophages or poorly differentiated monocytic cells. In fact, M-CSF knockout mice are known to develop osteopetrosis which is caused by immaturation of osteoclast, a resi-dent macrophage [54,55]. Moreover, it is reported that Ox-LDL is able to induce the differentiation of mono-cytes into macrophages [56]. These findings suggested that the effect of Ox-LDL on immature macrophages might differ in quality from that on mature macrophages.
Acknowledgements
This work was supported in part by a grant form Ono Medical Research Foundation.
References
[1] Ross R. The pathogenesis of atherosclerosis: a perspective for the 1990s. Nature 1993;362:801 – 9.
[2] Steinberg D, Parthasarathy S, Carew TE, Khoo JC, Witztum JL. Beyond cholesterol: modification of low-density lipoprotein that increase its atherogenicity. N Engl J Med 1989;320:915 – 24. [3] Palinski W, Rosenfeld ME, Yla¨-Herttuala S, Gurtner GC,
Socher SS, Butler SW, Parthasarathy S, Carew TE, Steinberg D, Witztum JL. Low density lipoprotein undergoes oxidative mod-ification in vivo. Proc Natl Acad Sci USA 1989;86:1372 – 6. [4] Yla¨-Herttuala S, Palinski W, Rosenfeld ME, Parthasarathy S,
Carew TE, Butler S, Witztum JL, Steinberg D. Evidence for the presence of oxidatively modified low density lipoprotein in atherosclerotic lesions of rabbit and man. J Clin Invest 1989;84:1086 – 95.
[5] Gordon D, Reidy MA, Benditt EP, Schwartz SM. Cell prolifera-tion inhuman coronary arteries. Proc Natl Acad Sci USA 1990;87:4600 – 4.
[6] Rosenfeld ME, Ross R. Macrophage and smooth muscle cell proliferation in atherosclerotic lesions of WHHL and compara-bly hypercholesterolemic fat-fed rabbits. Arteriosclerosis 1990;10:680 – 7.
[7] Spagnoli LG, Orlandi A, Santeusanio G. Foam cells of the rabbit atherosclerotic plaque arrested in metaphase by colchicine show a macrophage phenotype. Atherosclerosis 1991;88:87 – 92. [8] Sakai M, Miyazaki A, Hakamata H, Sasaki T, Yui S, Yamazaki
M, Shichiri M, Horiuchi S. Lysophosphatidylcholine plays an essential role in the mitogenic effect of oxidized low density lipoprotein on murine macrophages. J Biol Chem 1994;269:31430 – 5.
[9] Sato Y, Kobori S, Sakai M, Yano T, Higashi T, Matsumura T, Morikawa W, Terano T, Miyazaki A, Horiuchi S, Shichiri M. Lipoprotein(a) induces cell growth in rat peritoneal macrophages through inhibition of transforming growth factor-b activation. Atherosclerosis 1996;125:15 – 26.
[10] Sakai M, Miyazaki A, Hakamata H, Sasaki T, Yui S, Yamazaki Y, Shichiri M, Horiuchi S. Lysophosphatidylcholine potentiates the mitogenic activity of modified LDL for human monocyte-derived macrophages. Arterioscler Thromb Vasc Biol 1996;16:600 – 5.
[11] Sakai M, Miyazaki A, Hakamata H, Kodama T, Suzuki H, Kobori S, Shichiri M, Horiuchi S. The scavenger receptor serves as a route for internalization of lysophosphatidylcholine in oxi-dized low density lipoprotein-induced macrophage proliferation. J Biol Chem 1996;271:27346 – 52.
[12] Sakai M, Shichiri M, Hakamata H, Horiuchi S. Endocytosed lysophosphatidylcholine, through the scavenger receptor, play an essential role in oxidized low density lipoprotein-induced macrophage proliferation. Trends Cardiovasc Med 1998;8:119 – 24.
[13] Biwa T, Hakamata H, Sakai M, Miyazaki A, Suzuki H, Ko-dama T, Shichiri M, Horiuchi S. Granulocyte macrophage colony-stimulating factor plays a priming role in murine macrophage growth induced by oxidized low density lipoprotein. J Biol Chem 1998;273:28305 – 13.
[14] Matsumura T, Sakai M, Kobori S, Biwa T, Takemura T, Matsuda H, Hakamata H, Horiuchi S, Shichiri M. Two intracel-lular signaling pathways for activation of protein kinase C are involved in oxidized low density lipoprotein-induced macrophage growth. Arterioscler Thromb Vasc Biol 1997;17:3013 – 20. [15] Martens JS, Reiner NE, Herrira-Veit P, Steinbrecher UP.
Phos-phatidylinositol 3-kinase is involved in the induction of macrophage growth by oxidized low density lipoprotein. J Biol Chem 1998;273:4915 – 20.
[16] Martens JS, Lougheed M, Gomez-Munoz A, Steinbrecher UP. A modification of apolipoprotein B accounts for most of the induction of macrophage growth by oxidized low density lipo-protein. J Biol Chem 1999;274:10903 – 10.
[17] Hamilton JA, Myers D, Jessup W, Cochrane F, Byrne R, Whitty G, Moss S. Oxidized LDL can induce macrophage suvival, DNA synthesis, and enhanced proliferative response to CSF-1 and GM-CSF. Arterioscler Thromb Vasc Biol 1999;19:98 – 105. [18] Makheja AN, Bloom S, Muesing R, Simon T, Bailey JM.
Anti-inflammatory drugs in experimental atherosclerosis in WHHL rabbits and inhibition by cortisone acetate. Atheroscle-rosis 1989;76:155 – 61.
[19] Naito M, Yasue M, Asai K, Yamada K, Hayashi T, Kuyuya M, Funaki C, Yoshinine N, Kuzuya F. Effects of dexamethasone on experimental atherosclerosis in cholesterol-fed rabbits. J Nutr Sci Vitamin 1992;38:255 – 64.
[20] Villa AE, Guzman LA, Chen W, Golomb G, Levy RJ, Topol EJ. Local delivery of dexamethasone for prevention of neointi-mal proliferation in a rat model of balloon angioplasty. J Clin Invest 1994;93:1243 – 9.
[21] Ivandic B, Castellani LW, Wang X-P, Qiao J-H, Mehrabian M, Navab M, Fogelman AM, Grass DS, Swanson ME, de Beer MC, Lusis A. Role of group II secretory phospholipase A2 in
[22] Leitinger N, Watson AD, Hama SY, Ivandic B, Qiao J-H, Huber J, Faull KF, Grass DS, Navab M, Fogelman AM, de Beer FC, Lusis AJ, Berliner JA. Role of group II secretory phospholipase A2 in atherosclerosis: 2. potential involvement of biologically
active oxidized phospholipids. Arterioscler Thromb Vasc Biol 1999;19:1291 – 8.
[23] Grass DS, Felkner RH, Chiang M-Y, Wallace RE, Nevalainen TJ, Bennett CF, Swanson ME. Expression of human group-II PLA2in transgenic mice results in epidermal hyperplasia in the
absence of inflammatory infiltrate. J Clin Invest 1996;97:2233 – 41.
[24] Kennedy BP, Payette P, Mudgatt J, Vadas P, Pruzanski W, Kwan M, Tang C, Rancourt DE, Cromlish WA. A natural distraption of the secretory group II phospholipase A2gene in
inbred mouse strains. J Biol Chem 1995;270:22378 – 85. [25] Li YM, Baviello G, Vlassara H, Mitsuhashi T. Glycation
prod-ucts in aged thioglycollate medium enhance the elicitation of peritoneal macrophages. J Immunol Method 1997;201:183 – 8. [26] Hakamata H, Miyazaki A, Sakai M, Suginohara Y, Sakamoto Y,
Horiuchi S. Species difference in cholesteryl ester cycle and HDL-induced cholesterol efflux from macrophage foam cells. Arterioscler Thromb 1994;14:1860 – 5.
[27] Sakai M, Miyazaki A, Sakamoto Y, Horiuchi S. Cross-linking of apolipoprotein is involved in a loss of the ligand activity of high density lipoprotein upon Cu2+-mediated oxidation. FEBS Lett 1992;314:199 – 202.
[28] Miyazaki A, Rahim ATMA, Araki S, Morino Y, Horiuchi S. Chemical cross-linking alters high-density lipoprotein to be rec-ognized by a scavenger receptor in rat peritoneal macrophages. Biochim Biophys Acta 1991;1082:143 – 51.
[29] Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Laboratory Manual, 2nd edition. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 1989.
[30] Chen J, Shao C, Lazar V, Srivastava CH, Lee W-H, Tischfield JA. Localization of group IIc low molecular weight phospholi-pase A2 mRNA to meiotic cells in the mouse. J Cell Biol
1997;64:369 – 75.
[31] Alonso S, Minty A, Bourlet Y. Comparison of three actin-coding sequences in the mouse: evolutionary relationships between the actin genes of warm-blooded vertebrates. J Mol Evol 1986;23:11 – 22.
[32] Ma¨rki F, Franson R. Endogenous suppression of neutral-active and calcium-dependent phospholipase A2 in human
polymor-phonuclear leukocytes. Biochim Biophys Acta 1986;879:149 – 56. [33] Menschikowsky M, Kasper M, Lattke P, Schiering A, Schiefer S, Stockinger H, Jaross W. Secretory group-II PLA2 in human
atherosclerotic plaques. Atherosclerosis 1993;118:173 – 81. [34] Hurt-Camejo E, Camejo G. Potential involvement of type II
phospholipase A2in atherosclerosis. Atherosclerosis 1997;132:1 –
8.
[35] Forst S, Weiss J, Elsbach P, Maraganore JM, Reardon I, Heinrikson RL. Structural and functional properties of a phos-pholipase A2purified from an inflammatory exudate.
Biochem-istry 1986;25:8381 – 5.
[36] Chang HW, Kudo I, Tomiyama M, Inoue K. Purification and characterization of extracellular phospholipase A2 from
peri-toneal cavity of caseinate-treated rat. J Biochem (Tokyo) 1987;102:147 – 54.
[37] Kortesuo PT, Nevalainen TJ. Phospholipase A2in human ascitic
fluid. Biochem J 1991;278:263 – 7.
[38] Jutila MA, Banks KL. Locally dividing macrophages in normal and inflamed mammary gland. Clin Exp Immunol 1986;66:615 – 24.
[39] Franson R, Dobrow R, Weiss J, Elsbach P, Weglicki W.
Isola-tion and characterizaIsola-tion of a phospholipase A2from an
inflam-matory exudate. J Lipid Res 1978;19:18 – 23.
[40] Vadas P, Pruzanski W. Biology of disease: role of secretory phospholipase A2 in the pathology of disease. Lab Invest
1986;55:391 – 404.
[41] Schieling A, Menschikowski M, Muller E, Jaross W. Analysis of secretory group II phospholipase A2expression in human aortic
tissue in dependence on the degree of atherosclerosis. Atheroscle-rosis 1999;144:73 – 8.
[42] Nevalainen TJ, Haapanen T. Distribution of pancreatic (group-I) and synovial-type (group-II) phospholipase A2in human tissue.
Inflammation 1993;17:453 – 464.
[43] Andersen S, Sjursen W, Laegreid A, Austgulen R, Johansen B. Immunohistochemical detection of non-pancreatic phospholipase A2(type-II) in human placenta and its possible involvement in
normal parturition at term. Prostaglandins Leucot Essent Fatty Acids 1994;51:19 – 26.
[44] Hurt-Camejo E, Andersen S, Standal R, Rosengren B, Sartipy P, Stadberg E, Johansen B. Localization of nonpancreatic secretory phospholipase A2 in normal and atherosclerotic arteries.
Arte-rioscler Thromb Vasc Biol 1997;17:300 – 9.
[45] Stewart CC. Local proliferation of mononuclear phagocytes in tumor. J Leukoc Biol 1983;34:23 – 7.
[46] Evans R, Cullen RT. In situ proliferation of intratumor macrophages. J Leukoc Biol 1984;35:561 – 72.
[47] Sawter RT. The significance of local resident pulmonary alveolar macrophage proliferation to population renewal. J Leukoc Biol 1986;39:77 – 87.
[48] Bitterman PB, Saltman LE, Adelberg S, Ferrans VJ, Crystal RG. Alveolar macrophage replication: one mechanism for the expan-sion of the mononuclear phagocyte population in the chronically inflamed lung. J Clin Invest 1984;74:460 – 9.
[49] Arita H, Hanasaki K, Nakano T, Oka S, Teraoka H, Matsumoto K. Novel proliferative effect of phospholipase A2in Swiss 3T3
cells via specific binding site. J Biol Chem 1991;266:19139 – 41. [50] Kurizaki T, Egami H, Murata K, Okazaki S, Yoshida N, Ogawa
M. Membrane-associated phospholipase A2stimulates DNA
syn-thesis in two murine fibroblasts. Res Commu Chem Pathol Pharmcol 1992;78:39 – 45.
[51] Upston JM, Neuzil J, Witting PK, Alleve R, Stocker R. Oxida-tion of free fatty acids in low density lipoprotein by 15-lipoxyge-nase stimulates nonenzymic, alpha-tocopherol-mediated peroxidation of cholesteryl esters. J Biol Chem 1997;272:30067 – 74.
[52] Sakai M, Biwa T, Matsumura T, Takemura T, Matsuda H, Anami Y, Sasahara T, Kobori S, Shichiri M. Glucocorticoid inhibits oxidized LDL-induced macrophage growth by suppress-ing the expression of granulocyte/macrophage colony-stimulatsuppress-ing factor. Arterioscler Thromb Vasc Biol 1999;19:1726 – 33. [53] Fan TP, Lewis GP. Mechanism of cyclosporin A-induced
inhibi-tion of prostacyclin synthesis by macrophages. Prostaglandins 1985;30:735 – 47.
[54] Felix R, Hofstetter W, Wetterwald A, Cecchini MG, Fleish H. Role of colony-stimulating factor-1 in bone metabolism. J Cell Biochem 1994;55:340 – 9.
[55] Nilsson SK, Lieschke GJ, Garcia-Wijnen CC, Williams B, Tzelepis D, Hodgson G, Grail D, Dunn AR, Bertoncello I. Granulocyte-macrophage colony-stimulating factor is not re-sponsible for the correction of hematopoietic deficiencies in the maturing op/op mouse. Blood 1995;86:66 – 72.
[56] Frostegard J, Nilsson J, Haegerstrand A, Hamaten A, Wigzell H, Gidlund M. Oxidized low density lipoprotein induces differentia-tion and adhesion of human monocytes and the monocytic cell line U937. Proc Natl Acad Sci USA 1990;87:904 – 8.