Legum Pakan t
Cen trosema pubescens Benth
Cen trosema pubescens Benth disebut j uga centro (Australia), "jetiranan (Argentina, Brazil), "bejuco de chivo*
,
ncompanillan (Columbia) dan "butterfly pean (Ing- gris).
Tanaman tersebut termasuk f amili Leguminoceae(legum) dan sub famili Papilionaceae, berasal dari Amerika Selatan dan telah tersebar luas di daerah tropika maupun subtropika (Skerman, 1977)
.
Disamping sebagai hi jauan pakan dan pengendali erosi, centro telah dimanfaatkan sebagai tanaman penutup tanah ( cover crops) di perkebunan karet di Jawa sejak tahun 1922 (Purwowidodo, 1986). Centro mampu menekan pertumbuhan gulma dan meningkatkan kesuburan tanah (Lukiwati et dl., 1992). Tanaman tersebut dapat dipertimbangkan untuk diterapkan dalam sistem silvopastura(Lukiwati et al., 1995)
.
Centro termasuk tanaman tahunan, tumbuh menj alar dan membelit berlawanan dengan arah jarum jam sepanjang 1-4 meter. Tanaman tersebut berakar pada setiap m a s batang- nya, tetap hijau dan tidak mengayu meskipun sudah b e m m u r 18 bulan (Skerman, 1977)
.
Centro sesuai untuk padang penggembalaan karena tahan terhadap injakan dan disukai ternak serta tidak beracun (Cameron, 1984).
Centro mamputumbuh (persisten) pada lahan marginal (Stiir dan Shelton, 1991) pada ketinggian hingga 250 m di atas penrmkaan laut (Teitzel dan Burt, 1976) serta tidak menghendaki jenis ta- nah tertentu (Nguyen, 1973). Tanaman ini cepat bernodula- si, tetapi rendah produksi bahan keringnya (Chee dan Wong, 1986). Dapat tumbuh baik pada temperatur 25
-
30°c, sedang pada 18-
24'~ pertumbuhan centro akan terhambat(Whiteman et al., 1974). Pendapat tersebut diperkuat oleh Lukiwati et al., (1992) bahwa pertumbuhan centro lebih
baik di dataran rendah pada ketinggian 175 meter diatas permukaan laut (2 27'~) dibanding pada dataran tinggi 1150 m di atas permukaan laut (2 20'~)
.
Bahan tanam centro berupa biji, ditanam sedalam 2,5cm
-
5 cm di bawah permukaan tanah pada jarak 0,5 m x 1 m. Sebelum ditanam, biji centro perlu diretaskan dulu dengan merendam dalam larutan asam sulfat pekat 96 % selama 7 me- nit kemudian dicuci dengan air yang mengalir (Skerman, 1977).
Pertumbuhan awal centro lambat, tetapi setelah berumur 4-
6 bulan akan membentuk hamparan yang cukup le- bat hingga menutup permukaan tanah setebal 45 cm (Yates, 1975).
Tanaman tersebut mampu berkompetisi dengan gulma setelah berumur 4-
6 bulan (Skerman, 1977).Centro mampu berproduksi sebagai hijauan pakan selama 7 tahun di bawah pohon karet (Topark-Ngarm dan Gutteridge, 1986). Persistensi centro cukup baik pada interval potong
90 hari dengan kadar nitrogen sekitar 3 persen (Adjei dan Fianu, 1985)
.
Produksi bahan kering hi jauan centro pada periode pemotongan pertama sekitar 5 ton/ha (Hwang et dl., 1986). Reksohadiprodjo (1985) menyebutkan bahwa produksi bahan kering hijauan centro pada umur 7 minggu hanya sekitar 1 ton/ha. Pada interval potong 60 hari diperoleh produksi bahan kering 20 ton/ha/tahun (Sair et dl., 1991) sedangkan menurut Siregar et al., (1986) hanya sekitar 4 ton/ ha/tahun.Pueraria phaseoloides (Roxb
.
) BenthLegum Pueraria phaseoloides (Roxb
.
) Benth disebut juga P. javanica (Benth) Hook atau puero, tropical kudzu (Australia) dan kacang ruj i (Indonesia).
Tanaman ini termasuk legum tahunan untuk pastura, berasal dari Asia Tenggara yaitu Malaysia dan Indonesia serta telah tersebar di seluruh negara tropika basah (Toledo, 1986). Disamping sebagai hijauan sumber protein yang sangat disukai ternak ruminansia, puero juga berpeqan dalam meningkatkan ke- suburan tanah (Beck dan Vangmai,
1986 ).
Puero tumbuh menjalar dengan sistem perakaran dalam, berperan sebagai pencegah erosi dan menekan gulma (Whiteman et al., 1974) serta mampu menutup tanah setelah berumur 3 bulan (Chee dan Wong, 1986)
.
Pada umur 8-
9 bulan dapat menutup permukaan tanah setebal 60-
75 cm (Skerman, 1977). Apabila puero ditanam sebagai covercrops di bawah pohon karet, dapat dimanfaatkan juga se- bagai hi jauan pakan berkualitas tinggi untuk ternak selama 7 tahun (Topark-Ngarm dan Gutteridge, 1986)
.
Dengan demikian puero dapat pula dipertimbangkan untuk dibudidayakan dalam sistem silvopastura (Lukiwati et al., 1995).Puero toleran terhadap kekeringan serta mampu tumbuh pada berbagai jenis tanah pada ketinggian sampai dengan 1000 meter di atas permukaan laut serta mampu tumbuh pada tanah masam dan kahat unsur hara P (Kerridge et dl., 1986). Bahan tanam centro berupa biji dengan jarak tanam 0,5 m x 1 m sedalam 1,5 cm. Sebelum ditanam, biji puero perlu diretaskan dulu dengan merendam dalam larutan asam sulfat pekat 96 % selama 20 menit kemudian dicuci dengan air yang mengalir (Skerman, 1977)
.
Produksi bahan kering hijauan legum tersebut 17 ton/ha/tahun (Reksohadiprodjo, 1985). Sedang menurut Susetyo et dl., (1969) produksi bahan kering hijauan puero 8-
24 ton/ha setiap kali pemotongan. Apabila ditanam di pot, pada umur 4-
6 bulan diperoleh produksi bahan kering hijauan puero 8-
12 ton/ha/tahun (Whyte et al., 1953). Kadar nitrogen hijauan legum tersebut sekitar 3 persen berdasarkan bahan keringPeranan Pupuk Fosfor terhadap Pertumbuhan Legum
Fosfor (PI mendapat sebutan sebagai kunci kehidupan karena secara tidak langsung terlibat dalam proses-proses kehidupan, yaitu sebagai komponen dari tiap-tiap sel hidup dan cenderung terkonsentrasi dalam bi j i dan titik tumbuh tanaman (Thompson dan Troeh, 1978)
.
Fosfor merupakan unsur mineral yang diperlukan untuk sintesis ATP yaitu suatu senyawa organik yang bersifat sebagai kunci utama dalam reaksi-reaksi energetik pada berbagai proses meta- bolisme (Noggle dan Fritz, 1983).
Konsentrasi P hi jauan pakan sangat berpengaruh terhadap kinerja ternak. Mening- katnya intake P akan meningkatkan pertambahan bobot badan ternak (Little, 1968). Apabila sapi digembalakan pada pastura yang kahat unsur hara P I akan menunjukkan gejala afosforosis antara lain pertumbuhan badan terhambat (ker- dil) dan pertumbuhan tulang abnormal (Winks, 1990).
Tanah-tanah di daerah tropika pada umumnya kahat fosfor, efisiensi pupuk P rendah karena kapasitas fiksasi P tinggi terutama pada tanah masam (Clark dan Brown, 1980)
.
Dengan demikian f osfor adalah faktor pembatas utama terhadap produksi dan nilai nutrisi pastura tropika (Jones, 1990). Nilai nutrisi hijauan pakan antara lain diceminkan dari serapan N dan PI konsentrasi Cu dan Zn serta kecernaan bahan kering hijauan (Norton et al., 1990). Unsur hara P juga penting untuk perbaikan per- tumbuhan, produksi dan persistensi legum di daerah tro- pika maupun sub tropika (Crowder dan Chheda, 1982).Tanaman legum selain mampu mengatasi kekurangan protein dalam pakan ternak juga dapat memperbaiki produktivitas lahan (Coates et al., 1990)
.
Mantapnya (establish) pastu- ra legum seringkali lambat pada tanah masam dan kahat unsur hara P I karena legum menghendaki ketersediaan unsur hara P yang relatif tinggi untuk fiksasi nitrogen (Howeler et dl., 1987).
Oleh karena itu diperlukan sejumlah besar unsur hara P terutama pada tanah dengan kadar fosfor rendah (Whiteman, 1980).
Namun demikian tanaman hanya mampu menyerap 1/3-
1/2 dari fosfor yang ditambahkan, sehingga memerlukan pemupukan dosis tinggi. Tetapi pada pemupukan P yang berlebihan dapat menyebabkan kekurangan unsur Zn dan Cu (Brady, 1974).
Kekurangan Zn pada pakan domba dapat mengakibatkan pertumbuhan ternak terhambat serta mengalami atropi testis dan spermatogenesis tidak sempurna (Underwood dan Somers, 1969). Fertilitas kambing dan domba menurun apabila kekurangan Cu dalam pakamya (Howell dan Hall, 1970; Unanian dan Feliciano-Silva, 1984).
Pemberian pupuk P dapat menurunkan kapasitas fiksasi P tanah (Fox dan Kamprath, 1970) dan meningkatkan produk- si bahan kering hijauan legum siratro (Lowe et dl., 1981). Pemberian pupuk P sebanyak 0, 31, 62, 125, dan 250 kg/ha dapat meningkatkan produksi N Stylosanthes humilis secara linier (Gates, 1974)
.
Di daerah tropika dianjurkan meng- gunakan dosis pupuk P sebesar 100-
400 kg p205/ha dandapat diberikan secara disebar rata pada waktu tanam (Goedert et dl., 1982). Tetapi pada tanah-tanah yang sangat kekurangan fosfor dan curah hujan tinggi maka pemupukan berat sebesar 769
-
1282 kg P205/ha diperlukan untuk merangsang pertumbuhan legum (Yates, 1975). Mening- katnya dosis pemupukan P mampu meningkatkan produksi polong muda kacang panj ang (Lukiwati,
1990) demikian juga kadar P hijauan legum (Coates et al., 1990).Batuan fosfat (BF) termasuk salah satu jenis pupuk P lambat tersedia, mengandung 25-28% P205/ha (Kang, 1968). Batuan fosfat lebih sesuai untuk tanaman tahunan pada tanah masam dibanding TSP karena selain menambah unsur hara P juga unsur Ca (Kerridge, 1978)
.
Kadar P hijauan legum puero meningkat dengan pemupukan BF ( Waidyanatha et al., 1979). Dosis pemupukan BF (0, 100, 200, 300 kg P205/ha) nyata meningkatkan kadar P hi j auan legum masing- masing sebesar 0,lO; 0,15; 0,17; dan 0,20 % (Lukiwati et a1.
,
1 9 9 4 ~ ).
Produksi bahan kering hi jauan legum dengan pemberian pupuk P dalam bentuk TSP maupun BF (200 kg P205/ha) nyata lebih tinggi daripada tanpa pupuk P. Namun antara pemupukan TSP dan BF menghasilkan tingkat produksi bahan kering hi jauan yang tidak berbeda (Lukiwati, 1993).
Produksi ternak juga dipengaruhi oleh intake bahan kering dapat dicerna (Whiteman, 1980).Mikoriza Vesikular-Arbuskular
Mikoriza merupakan suatu struktur yang dibentuk oleh cendawan secara simbiotik dengan sel akar tanaman inang. Cendawan mikoriza terdiri dari 2 tipe utama yaitu endomi- koriza dan ektomikoriza (Peterson dan Bonfante, 1994). Asosiasi ektomikoriza terdapat pada spesies tanaman pohon dari famili Pinaceae, Betulaceae dan Fagaceae dan Diptero-
carpaceae (Smits, 1994). Asosiasi endomikoriza meliputi 3
kelompok yaitu famili Orchidaceae (mikoriza anggrek),
Ericaceae (mikoriza ericoid) dan mikoriza vesikular-arbus-
kular (MVA)
.
Cendawan MVA paling luas penyebaramya antara lain pada famili Gramineae (rumput-rumputan) dan Leguminoceae ( legum) (Gerdemam, 19 6 8 ).
Istilah mikoriza digunakan pertama kali oleh Frank (1885) ketika menguraikan adanya hubungan simbiosis mutua- lis antara cendawan dan akar tanaman. Mikoriza berasal dari 2 kata dalam bahasa Yunani (Greek) yaitu mykes
(cendawan) dan r h i z a (akar tanaman)
.
Mosse dan Bowen (1968) menyebutkan bahwa mikoriza vesikular-arbuskular tennasuk dalam genus Endogone.-
Namun kemudian diketahui bahwa Endogone tidak membentuk arbuskular tetapi membentuk asosiasi ektomikoriza (Gerdemam dan Trappe, 1974).
Pada perkembangan penelitian taksonomi berikutnya diketahui bahwa ordo Endogonales (Zygomycotina) hanya mempunyai satu famili Endogonaceae meliputi 6 genus yaitu AcaulosporaGerdemann & Trappe, Entrophospora Ames & Schneider, Gigas- pora Gerdeman & Trappe, Glomus Tulasne & Tulasne, Sclero- cystis Berkeley & Broome dan Scutellispora Walker & Sand- ers (Benjamin, 1979 dan Morton, 1988)
.
Dari hasil revisi klasifikasi cendawan mikoriza vesikular-arbuskular berda- sarkan morfologi spora ditemukan bahwa 6 genus pembentuk arbuskular tersebut termasuk dalam ordo Glomales (Morton dan Benny, 1990; Schenck dan PBrez, 19.90) sebagai berikut:Ordo : Glomales Morton & Benny Sub Ordo : Glomineae Morton & Benny
Famili : Glomaceae Pirozynski & Dalpe Genus : Glomus Tulasne & Tulasne
Genus : Sclerocystis (Berkeley & Broome) Almeida & Schenck
Famili : Acaulosporaceae Morton & Benny
Genus : Acaulospora (Gerdemann & Trappe) Berch Genus : Entrophospora Ames & Schneider
Sub Ordo : Gigasporineae Morton & Benny Famili : Gigasporaceae Morton & Benny
Genus : Gigaspora (Gerdemann & Trappe) Walker & Sanders
Genus : Scutellispora Walker & Sanders.
Mikoriza vesikular-arbuskular terdapat pada hampir semua jenis tanah dan tidak memerlukan tanaman inang yang
spesifik (Mosse dan Hayman, 1980). Mikoriza vesikular- arbuskular dicirikan dengan adanya struktur vesikel, ar- buskel dan hifa di dalam akar serta spora. Vesikel ber- bentuk kantung, bulat, terminal atau interkalari yang
terdapat di dalam sel atau antar sel (Bonfante-Fasolo, 1984)
.
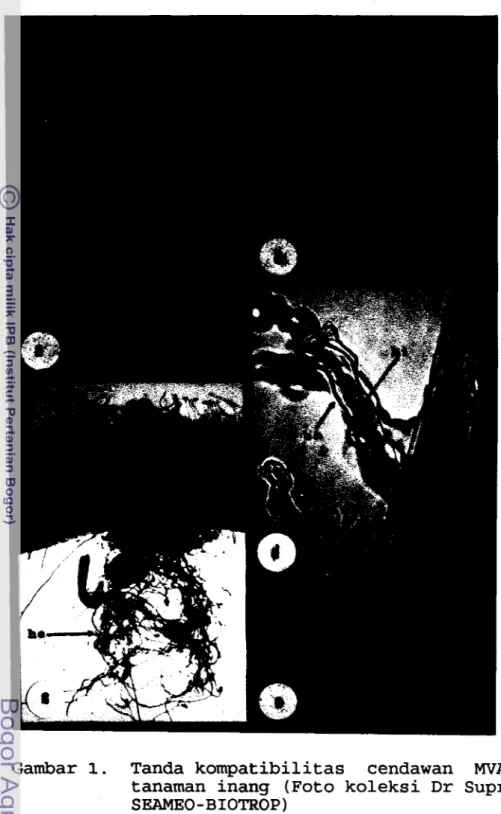
Fungsi vesikel sebagai tempat penyimpanan ca- dangan makanan. Arbuskel merupakan struktur seperti haustorium dan bercabang banyak yang terletak di dalam sel (Gambar la). Fungsi arbuskel adalah sebagai tempat pemindahan nutrisi antara cendawan mikoriza dan tanaman inang. Arbuskel berumur pendek, yaitu antara 1-
3 minggu (Mosse, 1981), sedang menurut Smith dan Smith (1995) hanya sekitar 4 hari. Arbuskel dengan cepat mengalami desinte- grasi atau terjadi lisis/pecah dan membebaskan P ke tanaman inang (Hudson, 1986; Smith dan Smith, 1995). Luas area permukaan arbuskel aktif secara metabolik per meter akar berkurang dengan waktu, sedang hifa mempunyai area permukaan lebihbesar sesudah 63 hari setelah tanam (Smith dan Smith, 1995).
Penelitian-penelitian tentang mikoriza berkembang pesat meliputi identifikasi spesies cendawan MVA mulai dari tahun 1988 yaitu 120 spesies (Schenck dan Perez, 1988)
.
Pada tahun berikutnya meningkat menjadi 126 spesies dapat diidentifikasi (Schenck dan Perez, 1989). Pada penelitian selanjutnya dapat diidentifikasi sebanyak 147 spesies (Schenck dan Perez, 1990) dan 150 spesies(Morton dan Bentivenga, 1994) yang sebagian besar di- dominasi oleh genus Glomus.
Setiap spesies MVA berbeda dalam kemampuannya untuk merangsang pertumbuhan tanaman inang. Hal ini diduga karena ada perbedaan dalam ukuran spora dan dikaitkan dengan pembentukan hifa eksternal yang berperan dalam efisiensi penyerapan unsur hara, antara lain fosfor (Da- niels dan Menge, 1981). Perkembangan kolonisasi MVA dimu- lai dengan pembentukan suatu appresori urn (Gambar lb) pada
permukaan akar oleh hifa eksternal yang berasal dari spora atau akar bermikoriza di dalam tanah. Hifa dari appresorium kemudian menembus sel-sel epidermis dan menja- lar di antara sel (interseluler) ataupun didalam sel (in- traseluler) menembus korteks tetapi tidak meluas ke endo- dermis ataupun stele. Akar bermikoriza dapat membentuk hifa eksternal (Gambar lc) yang merupakan kelanjutan dari hif a internal (Gambar Id) kemudian vesikel terbentuk pada ujung-ujung hifa (Gambar le)
.
Tiap spesies tanaman dapat mengendalikan luasnya kolonisasi dan aktivitas cendawanMVA dalam sistem akarnya segera setelah mikoriza mantap perkembangannya. Meskipun demikian, dalam keadaan ter- tentu pertumbuhan tanaman dapat tertekan sebagai akibat kolonisasi mikoriza. Hal ini diduga karena cendawan MVA
sangat agresif sehingga tanaman tersebut tidak mampu lagi untuk mengendalikan aktivitas mikoriza (Koi.de, 1985).
Gambar 1. Tanda kompatibilitas cendawan MVA dengan tanaman inang (Foto koleksi Dr Supriyanto/ SEAMEO-BIOTROP)
Keterangan :
a. Arbuskel (arb) (pembesaran 400 x)
.
b. Appresorium (ap) (pembesaran 200 x).
c.
Hifa eksternal (he) (pembesaran 40 x).
d. Hifa internal (hi),
bulu akar (ba)(pembesaran 100 x)
Akar bermikoriza dapat diamati dengan mikroskop sesudah di warnai dengan trypanblue atau larutan asam fuchsin- asam laktat (Kormanik dan McGraw, 1982).
Asosiasi simbiosis mutualis mikoriza dikendalikan oleh suatu seri mekanisme dalam tanaman dan diatur oleh keadaan lingkungan, terutama fisiologis dari tanaman inang yang tanggap dalam menentukan kesesuaian terhadap pemben- tukan dan fungsi mikoriza. Spesies tanaman disebut miko- troph fakultatif atau fakultatif mikorizal apabila asosia- si mikoriza tersebut hanya menguntungkan jika media tum- buhnya pada tanah yang tidak subur. Tanaman disebut mikotroph obligat apabila sangat tergantung pada asosiasi mikoriza untuk pertumbuhan, perkembangan dan reproduktif- nya pada semua kondisi lingkungan (Koide dan Li, 1990). Tanaman legum mempunyai sedikit bulu-bulu akar (magnolioid roots) oleh karena itu dalam pertumbuhamya sangat tergan- tung pada asosiasi mikoriza (Mosse, 1981). Ketergantungan suatu jenis tanaman terhadap cendawan mikoriza pada tanah yang disterilisasi dapat diketahui dengan mengguna- kan rumus relative mycorrhizal dependency (RMD) dari Menge et dl., yang dikutip oleh Bagyaraj (1992) sebagai berikut :
BK tanaman bermikoriza
RMD = x 100%
BK tanaman tanpa mikoriza BK = berat kering hijauan
Pada kondisi lapang tanpa sterilisasi, respons per- tumbuhan tanaman terhadap inokulasi mikoriza dapat diketa- hui dengan menggunakan rumus mycorrhi zal inoculation
effect (MIE) dari Bagyaraj
et
al.,yang dikutip oleh Bagya- raj (1992) sebagai berikut :BK dinokulasi MVA
-
BK tanpa inokulasi MVAMIE = x 100 %
BK tanpa inokulasi MVA BK = berat kering hijauan
Rumus tersebut diatas (MIE) sangat berguna untuk mengeta- hui adanya kompetisi antara cendawan MVA yang di introduk- sikan dengan cendawan MVA- indigenous serta pengaruhnya terhadap respons pertumbuhan tanaman.
Morfologi dan geometri sistem perakaran berpengaruh terhadap kemampuan tanaman inang dalam memperoleh unsur hara maupun dalam berasosiasi dengan mikoriza. Faktor- faktor yang mempengaruhi pembentukan asosiasi dengan miko- riza dapat berpengaruh terhadap fungsi simbiotik tersebut. Untuk pertumbuhan dan fungsi mikoriza tergantung pada suplai karbon sebagai derivat fotosintesis dari tanaman inang (Azcon dan Bago, 1994)
.
Hal ini berarti bahwa fungsi normal dari asosiasi simbiotik mikoriza terutama tergantung pada aktivitas fotosintesis. Oleh karena itu cendawan mikoriza yang lebih efektif dalam pengambilan unsur hara, akan lebih tinggi pula kebutuhan karbohidrat yang h a m s diterima dari tanaman inang. Cendawan mikoriza menerima karbohidrat dari tanaman inang sebanyak 4-
14 %dari total karbon yang difotosintesis dialokasikan ke simbiont (Paul dan Clark, 1989).
Di dalam tanah, cendawan MVA pada umumnya berbentuk spora dalam keadaan istirahat (dorman) (Pearson dan Diem, 1982). Spora dapat diisolasi dengan metode tuang saring basah dari contoh tanah rhizosf ir yang diambil pada keda- laman 0
-
15 cm. Metoda tersebut digunakan untuk memisah- kan spora dari material koloidal dan partikel-partikel tanah yang lebih berat (Gerdemam, 1955).
Selanjutnya dengan bantuan mikroskop stereo, spora tersebut dapat diambil dengan menggunakan pipet pasteur atau mikrospatula (Gerdemam dan Nicolson, 1963).
Untuk inokulasi dapat menggunakan inokulum tanah, inokulum akar dan spora. Spora dalam kultur pot dapat disimpan selama 4 tahun (Ferguson dan Woodhead, 1982). Inokulum tanah mampu mempertahankan keefektifamya sampai dengan 3 tahun apa- bila disimpan pada keadaan lembab dalam kamar dingin. Inokulum akar tahan disimpan selama 30 hari pada tempera- tur kamar dan masih seefektif inokulum segar dalam mening- katkan pertumbuhan tanaman pada tanah steril. Namun demi- kian inokulum akar dikhawatirkan dapat membawa masalah timbulnya penyakit tanaman (phytosani tary) (Howeler et a1.,1987). Untuk iriokulasi di lapang, inokulum tanah lebih efektif untuk kolonisasi akar dan produksi spora daripada inokulum lainnya. Hal ini karena inokulum tanah mengandung bermacam-macam bentuk propagule (spora, frag- ment akar bermikoriza dan hifa eksternal) yang mampumeningkatkan persentase kolonisasi (Nadarajah dan Nawawi,
1990). Namun demikian tidak ada korelasi antara persen-
tase kolonisasi MVA dengan jumlah spora yang dihasilkan (Shankar et al., 1990). Pada umumnya untuk inokulasi pada penelitian di pot diperlukan 30
-
50 spora. Apabila kurang dari kisaran tersebut kemungkinan akan lebih rendah tingkat infeksinya (Ross dan Harper, 1970).
Untuk pene- litian di lapang kebutuhan jumlah spora lebih tinggi tetapi besarnya kolonisasi relatif lambat. Hal ini disebabkan karena faktor unsur hara dalam tanah atau genangan air maupun spesies cendawan laimya (Khan, 1974; Tamolang-Castilo, 1993).Metode inokulasi terbaik tergantung pada jenis ta- naman, sistem tanam dan siklus pertumbuhan tanaman. Penggunaan inokulum tanah untuk tanaman legum dapat di- berikan pada jalur di bawah biji atau secara tugal pada waktu tanam. Pemupukan dapat diberikan pada sisi yang lain setelah tanaman tumbuh (Howeler et dl., 1987).
Percobaan MVA pa& Tanah S t e r i l dan Tidak S t e r i l
Para peneliti terdahulu telah banyak melengkapi in- fomasi mengenai peranan MVA dalam meningkatkan produk- tivitas tanaman pada kondisi media tumbuh yang steril. Namun penelitian mengenai perkembangan dan fungsi MVA pada tanah yang tidak disterilisasi jarang dilakukan. Padahal
masih perlu diketahui mengenai respons tanaman bermikoriza pada keadaan aseptik maupun kondisi alami. Hal ini diper- lukan karena hasil percobaan di rumah kaca (lingkungan terkontrol) seringkali tidak konsisten dengan percobaan di lapang. Keadaan tersebut disebabkan karena adanya kompetisi dengan mikroorganisme tanah yang ada (Linderman, 1992). Dengan demikian inokulasi MVA lebih baik pengaruh- nya pada tanah yang sudah disterilisasi dibanding tanpa sterilisasi (Cao et al., 1994)
.
Alasan utama melakukan percobaan pot dengan mengguna- kan tanah steril adalah untuk memudahkan pengendalian mikoriza. Pada tanah steril, MVA lebih efektif dibanding tanpa sterilisasi. Namun demikian, untuk tujuan memheda- kan pertumbuhan dan fungsi mikoriza sebaiknya dilakukan pada kondisi lapang yang secara kualitatif dan kuantitatif adalah realistik (Cao et a1
.
,
1994).
Oleh karena itu cendawan mikoriza ditumbuhkan pada suatu lingkungan yang secara fisik, kimia dan biologik sama dengan yang ada di habitat secara alami. Hal ini berarti bahwa pada tanah yang tidak disterilisasi dapat mewakili kondisi habitat tersebut. Disamping itu, telah diketahui bahwa cendawan MVA di lapang berinteraksi secara luas dengan organisme tanah lainnya di dalam akar dan di rhizosfir. Bentuk- bentuk interaksi dapat menghambat (inhibi tory),
menst imu- lasi (stimulatory),
kompetisi (competitive) dan mutualis- tik (mutualistic).
Pengaruh-pengaruh tersebut dapat'23 terj adi pada semua tahap siklus hidup cendawan mikoriza yai tu mulai dari germinasi spora, kolonisasi akar sampai dengan pertumbuhan hifa eksternal (Fitter dan Garbaye, 1994). Namun proses interaksi berbagai jenis cendawan MVA di lapang selama kolonisasi akar belum diketahui (Abbott dan Gazey, 1994)
.
Lambamya perkembangan pemanfaatan MVA pada tanaman pertanian disebabkan karena respons pertumbuhan tanaman terhadap MVA di lapang sangat bervariasi (Bagyaraj, 1984; Powell, 19841, atau tidak ada respons (Lin dan Hao, 1991). Pengaruh inokulasi MVA bervariasi pada set iap j enis tanah tergantung pada efektivitas spora indigenous maupun inoku- lum yang diintroduksikan (Mitiku-Habte dan Fox, 1993)
.
Namun ada pula respons pertumbuhan tanaman terhadap inoku- lasi MVA pada tanah tidak steril sama dengan pada tanah steril, ha1 ini karena MVA indigenous tidak efektif (Mosse, 1973). Beberapa hasil penelitian mengenai res- spons tanaman terhadap MVA pada tanah steril dan tidak steril antara lain produksi bi j i kedelai bermikoriza nyata lebih tinggi dibanding tanpa mikoriza pada tanah steril, sedang pada tanah tidak steril hasilnya tidak berbeda nyata (Powell, 1984). Produksi biji dan jerami kacang pan j ang (cowpea) (Ramara j dan Shanmugam, 1990),
produksi bahan kering, kadar N dan kolonisasi MVA pada Leucaena esculenta (Valdes et a1.
,
1993 ) dengan inokulasirhizobium tidak berbeda dengan apabila diinokulasi ganda MVA dan rhizobium pada tanah tidak steril. Pada peneli- tian lapang, tidak terdapat perbedaan kandungan N dan P antara kedelai dan mungbean (Vigna radiata) bermikoriza dibanding tanpa mikoriza. Perbedaannya terdapat pada kandungan N dan P tanah dengan MVA yang efektif akan lebih rendah daripada tanpa MVA (Thamsurakul et al., 1991).
Perbedaan toleransi spesies cendawan mikoriza terha- dap kondisi tanah yang berbeda perlu diperhatikan. Tanam- an jagung responsif terhadap MVA pada percobaan pot, tetapi tidak responsif pada percobaan lapang walaupun dengan jenis tanah yang sama (Islam, 1976). Beberapa j enis tanaman mempunyai tingkat kompatibilitas yang tinggi dengan MVA-indigenous. Hal ini ditunjukkan dengan tingkat kolonisasi akar oleh MVA (Khasa et al., 1992). Introduksi tanaman yang cepat tumbuh serta sangat tergan- tung pada mikoriza, dapat meningkatkan populasi MVA- indigenous (Howeler et dl., 1987). Kadar N, P serta se- rapan P pada tanaman kacang panjang (cowpea/Vigna unguila-
ta) lebih tinggi dengan inokulasi MVA dibanding tanpa
inokulasi MVA pada tanah tidak steril. Namun ternyata MVA-indigenous berkembang secara ekstensif dan mampu mengkolonisasi akar sebesar 5 3 , 7 8 % (Chhabra et dl., 1990). Produksi bahan kering hijauan kacang tanah, kacang panjang dan pigeon pea (Cajanus cajan) dengan inokulasi MVA pada tanah steril tidak berbeda dibanding pada tanah tidak
steril. Meskipun kolonisasi akar nyata lebih tinggi pada tanah tidak steril (Khasa et al., 1992).
Beberapa faktor yang menyebabkan penampilan cendawan MVA kurang baik pada kondisi tanah tidak steril, antara lain karena di rhizosfir tersebut juga terdapat cendawan, bakteri dan aktinomisetes (Kehri dan Chandra, 1988). Keadaan tersebut menyebabkan terjadinya kompetisi mikro- bial secara langsung terhadap fungsi mikoriza. Apabila ada patogen yang menginfeksi akar, dapat menyebabkan busuk akar atau penyakit layu daun (Kommedahl dan Windels, 1979). Selanjutnya cendawan MVA tidak akan mengkoloni akar yang sudah terinfeksi nematoda dan sebaliknya nemato- da jarang menginfeksi akar yang sudah di koloni oleh MVA
(Ingham, 1988). Meskipun kadang-kadang patogen akar dan cendawan MVA menempati jaringan akar yang berdekatan tanpa pengaruh yang nyata antara keduanya (Dehne, 1982). Mikro organisme lain pada tanah tidak steril dapat menekan germinasi spora cendawan MVA maupun respons pertumbuhan tanaman terhadap MVA. Sebagai contoh nematoda dapat me- ngurangi potensi MVA dengan memakan hifa eksternal (Linderman, 1992). Dengan demikian adanya mikoparasit dapat menurunkan produksi spora MVA. Hal ini karena spora MVA yang terkontaminasi mikoparasit tidak dapat germinasi dan mati. Dengan demikian mikoparasit berpengaruh terha- dap potensi MVA di lapang (Paulitz dan Menge, 1986).
Tiga faktor utama yang menentukan keberhasilan intro- duksi MVA di lapang yaitu ketergantungan tanaman terhadap MVA, status nutrisi tanah dan potensi inokulum MVA (Thomp- son, 1994)
.
Disamping itu juga kepadatan populasi MVA- indigenous (Abbott et a1.
,
1983).
Di lain pihak, keber- hasilan inokulasi MVA dalam meningkatkan pertumbuhan tanaman di lapang pada tanah tidak steril masih diragu- kan apakah disebabkan karena pengaruh MVA saja atau m e n - pakan efek kumulatif dari inokulum MVA dan MVA-indigenous. Meskipun telah dilakukan inokulasi MVA, namun tidak dapat dijamin bahwa hanya inokulum MVA tersebut yang mengkoloni akar tanaman, karena di lapang terdapat ber- macam-macam populasi cendawan MVA-indigenous (Giovamettiet al., 1988)
.
Akar tanaman dapat terkoloni MVA dari beberapa spesies cendawan MVA dengan keragaman yang lebih luas di lapang. Belum diketahui mekanisme suatu populasi cendawan MVA tersebut membentuk asosiasi simbiotik mikori- za pada sistem perakaran tanaman di lapang pada waktu yang sama (Morton, 1988).
Disamping itu, rhizosfir adalah suatu area yang kompleks dengan aktivitas mikrobial. Dengan demikian ada kemungkinan terjadi interaksi biologik antara cendawan MVA dan mikroorganisme tanah yang mengun- . tungkan (Bagyaraj,
1990).
Hal ini sehubungan dengan adanya asosiasi epipit antara Azotobakter sp dengan spora MVA, Pseudomonas dengan spora MVA yang mempunyai efek merangsang pertumbuhan tanaman (Mosse, 1972). Apabiladibandingkan dengan di pot, maka pertumbuhan tanaman di lapang mempunyai sistem perakaran lebih berlimpah dan siklus pertumbuhan tanaman di lapang lebih panjang sehing- ga meskipun populasi spora MVA-indigenous rendah, akan cukup waktu untuk berkembang membentuk asosiasi mikoriza yang lebih efektif. Oleh karena itu respons terhadap inokulasi MVA cenderung lebih rendah pada kondisi dila- pang dibanding di rumah kaca (Howeler et a l . 1987). Namn
demikian berlimpahnya cendawan MVA-indigenous di lapang bukan merupakan indikasi yang akurat bahwa besarnya kolonisasi akar tanaman karena adanya asosiasi simbiotik dengan cendawan-indigenous tersebut (Scheltema et al., 1987). Pendekatan morfologi merupakan salah satu metode yang dapat digunakan untuk mengenal cendawan MVA dalam akar yaitu morfologi hifa dan vesikular didalam akar (Abbott, 1982; Lopez-Aquilon dan Mosse, 1987; Abbott dan Gazey, 1994)
.
Pada penelitian di lapang tanpa sterilisasi dan tanpa inokulasi MVA, terdapat persentase kolonisasi MVA sebesar 36 %. Hal ini diduga karena adanya kolonisasi oleh spe- sies MVA-indigenous (Adholeya et a1
.
,
1988).
Sebagian besar informasi tentang dinamika dan keragaman cendawan mikoriza di lapang hanya berkisar pada tipe-tipe spora atau besarnya persentase kolonisasi akar oleh MVA (Gemma et al., 1989; Brundrett dan Kendrich, 1990; Johnson et al., 1992). Dengan demikian keberhasilan inokulasi MVA dilapang atau di pot dengan tanah tidak steril, sangat tergantung kepada kemampuan inokulum tersebut untuk hidup dan berkembang dalam lingkungan tumbuh yang baru serta mampu bersaing dengan mikrobia tanah yang ada. Disamping itu juga dipengaruhi oleh populasi dan efektivitas MVA- indigenous (Mosse dan Hayman, 1971; Howeler et al., 1987; Narayanan et dl., 1990).
Interaksi MVA-Fosfor-Tanaman
Famili Gramineae dan Leguminoceae (legum) dapat ber- asosiasi dengan MVA. Meskipun demikian legum lebih res- ponsif terhadap inokulasi MVA dibanding gramineae. Hal ini disebabkan karena tanaman legum mempunyai sedikit bulu-bulu akar (magnolioid roots) sehingga sangat tergan- tung pada asosiasi MVA (Mosse, 1981). Tanaman yang sangat tergantung pada asosiasi MVA mempunyai kebutuhan P ekster- nal sangat tinggi apabila ditanam pada lahan tanpa MVA. Kebutuhan P akan rendah apabila tanaman tersebut diasosi- asikan dengan cendawan MVA yang sangat efektif (Howeler et dl., 1987). Cendawan MVA mampu menekan penggunaan pupuk P dan pestisida sehingga dapat mengatasi masalah ekonomi, polusi air dan air-tanah (Plenchette dan Strullu, 1995).
Struktur MVA dalam akar tanaman mampu meningkatkan luas area untuk pertukaran metabolik antara tanaman inang dan cendawan MVA. Hifa eksternal berperan dalam mening- katkan potensi sistem perakaran untuk mengabsorbsi unsur
hara dan air (Pacovsky, 1986; Linderman, 1992)
.
Hal ini disebabkan karena hifa eksternal berperan dalam serapan dan translokasi unsur-unsur hara ke tanaman yaitu 80% PI 25% N, 10% K, 25% Zn dan 60% Cu (Marschner dan Dell, 1994). Hifa eksternal MVA mampu meningkatkan efisiensi tanaman dalam mengabsorbsi unsur hara dan air dari tanah. Hifa eksternal tersebut berfungsi sebagai jembatan trans- lokasi unsur-unsur imobil menj adi tersedia yaitu P I Zn dan Cu ke akar (Moawad, 1986; Howeler et dl., 1987).
Panjang hifa eksternal mencapai 7-
10 m/g tanah gada penelitian di laboratorium. Apabila di lapang panjang hifa dapat mencapai 5-
50 m/g tanah (Allen et dl., 1992). Kolonisasi MVA pada akar tanaman mempunyai peranan penting dalam meningkatkan pertumbuhan tanaman. Hal ini disebabkan karena meningkatnya serapan unsur hara dan secara tidak langsung juga berpengaruh terhadap modifika- si tingkat transpirasi dan komposisi mikroflora rhizosfir (Marschner dan Dell, 1994). Disamping itu tingkat foto- sintesis lebih tinggi pada tanaman yang dikolonisasi MVA dibanding tanpa MVA (Smith dan Pearson, 1988). Namun demikian tidak ada interaksi antara persentase kolonisasi MVA dalam akar, jumlah spora/100 g tanah dengan produksi bahan kering hi j auan (Furlan et a1.
,
1983).
Pengaruh MVA terhadap peningkatan bahan kering hi- jauan alfalfa diperlihatkan 10 minggu sesudah inokulasi
tanaman yang diinokulasi MVA lebih tinggi dibanding tanpa
MVA (Furlan et dl., 1983). Dilaporkan oleh Gupta et dl., (1990) bahwa MVA mampu meningkatkan produksi bahan kering, demikian juga serapan N dan P hijauan rapeseed. Peningkat- an produksi bahan kering hijauan jagung dan kadar P hi- jauan kedelai bermikoriza masing-masing 100% dan 62% lebih tinggi dibanding kontrol (Hamel et dl.
,
1991).
Demikian pula produksi bahan kering bawang putih dengan inokulasiMVA 41% lebih tinggi daripada tanpa MVA (Furlan dan Cardou, 1989).
Cendawan mikoriza mampu meningkatkan serapan N, PI Zn, Cu (La rue et dl., 1975; Lambert et dl., 1979; Swami-
nathan dan Verma, 1979; Waidyanatha et a1.,1979; Gildon dan Tinker, 1983; Moawad, 1986; Kothari et dl., 1990; Lambert dan Weidensaul, 1991). Hasil-hasil penelitian tersebut dilakukan pada tanaman peach, gandum, j agung
,
kentang, kedelai dan Stylosanthes guyanensis di daerah sub tropika pada tanah steril. Peningkatan serapan unsur-unsur hara tersebut disebabkan meningkatnya konsen- trasi Nl P, Zn dan Cu tanaman bermikoriza dibanding tanpa mikoriza (Hayman dan Mosse, 1972; Pacovsky, 1986; Subramaniam dan Dwivedi, 1988). Sebagai contoh konsentra- si Cu dan N lebih tinggi apabila diinokulasi MVA dibanding tanpa inokulasi, masing-masing pada kedelai (Lambert etKonsentrasi P pada hijauan tanaman bermikoriza adalah 3.7-7 kali lebih tinggi daripada tanpa mikoriza (Furlan
et dl, 1983)
.
Cendawan MVA responsif terhadap pemupukan P yang lambat tersedia seperti halnya BF (Azcon et dl., 1976)
.
Pertumbuhan legum tropika meningkat apabila diinokulasi MVA dengan kombinasi pemupukan BF. Disamping itu, pupuk BF lebih efektif pada tanah masam dan dapat merangsang populasi cendawan MVA-indigenous (Howeler et dl., 1987). Produksi spora cendawan MVA dengan pemupukan BF lebih tinggi daripada dengan superfosfat (Dodd et dl., 1990~). Tanaman legum lebih efektif dalam meningkatkan jumlah spo- ra dibanding rumput (Howeler et dl., 1987).
Pupuk P bervariasi pengaruhnya terhadap asosiasi sim- biotik MVA maupun cendawan MVA itu sendiri (Sylvia dan Neal, 1990 ; Abbott dan Robson, 1991)
.
Penggunaan pupukP dosis tinggi dapat mengubah komposisi dan efisiensi cendawan MVA (Howeler et a1
.
,
1987).
Sebagian besar hasil penelitian menyebutkan bahwa pupuk P dapat menurun- kan kolonisasi MVA. Namun demikian mampu meningkatkan panjang akar yang dikoloni MVA karena pertumbuhan akar ditingkatkan dengan pemupukan P. Tetapi apabila penambah- an pupuk P dilakukan pada tanah yang sangat rendah kandungan fosfatnya maka kolonisasi MVA pada perakaran tanaman meningkat (Bolan et dl., 1984).
Arnijee et dl., (1989) menambahkan bahwa pemupukan P dosis tinggi akanmengurangi pembentukan secondary entry point. Oleh karena itu meningkatnya dosis pupuk P dapat menurunkan persen- tase kolonisasi akar oleh MVA dan menurunkan jumlah spora dalam tanah (Champawat, 1990; Uma Devi dan Sitaramaiah, 1990) serta cenderung menurunkan serapan Zn pada level pupuk 1832 mg P/kg (Khare dan Thompson, 1991). Kolonisasi akar oleh MVA akan berkurang (Amijee et al., 1989; Koide dan Li, 1990) dan menurunkan produksi spora (Menge et al., 1978; Nelsen et al., 1981) apabila ketersediaan P sangat tinggi maupun sangat rendah.
Apabila unsur hara P sebagai faktor pembatas bagi pertumbuhan tanaman, maka pemberian pupuk P mampu meningkatkan konsentrasi P maupun produksi bahan kering hijauan (Howeler et al., 1987; Jones, 1990). Produksi maksimum kedelai bermikoriza dicapai pada dosis pupuk 117 kg P205/ha (Simanungkalit