Contrasting climate- and land-use-driven tree
encroachment patterns of subarctic tundra in
northern Norway and the Kola Peninsula
1
Sigrun Aune, Annika Hofgaard, and Lars So¨derstro¨m
Abstract:High-latitude regions are experiencing substantial climate change, and the forest–tundra transition is assumed to
sensitively track these changes through advancing treeline and increased tundra encroachment. However, herbivores may influence these responses. The present study addresses, through analyses of age structures, growth characteristics, and cli-mate correspondence, how mountain birch (Betula pubescensEhrh. ssp.czerepanovii(Orlova) Ha¨met-Ahti) treelines and sapling cohorts beyond the treeline have responded to the last decade’s warming in six North European subarctic areas with different climate and grazing characters. The results show different response patterns among areas representing ad-vancing, stationary, and possibly retreating treelines. Recruitment was abundant over the last decades in all areas except one, with predominantly arctic conditions, where both tree and sapling cohorts were old. Areas with high annual precipita-tion show advancing birch populaprecipita-tions characterized by young individuals and partly overlapping tree and sapling age dis-tributions. Areas in reindeer herding districts show stationary or retreating birch populations characterized by
nonoverlapping age distributions and low sapling survival. Recruitment patterns beyond the treeline generally corresponded with non-growing-season climate variables, mainly precipitation, indicating the importance of a protecting snow cover. The results highlight the important interplay between abiotic and biotic control over tundra encroachment and treeline dy-namics and the importance of multisite studies when addressing responses to warming.
Re´sume´ :Les re´gions situe´es a` des latitudes e´leve´es subissent d’importants changements climatiques. On assume que la
zone de transition entre la foreˆt et la toundra re´agit de fac¸on perceptible a` ces changements et que cette re´action se traduit par la progression de la limite des arbres et l’augmentation de l’empie`tement sur la toundra. Cependant, les herbivores pourraient influencer ces re´ponses. A` partir de l’analyse des structures d’aˆge, des caracte´ristiques de la croissance et de la concordance avec le climat, cette e´tude examine la fac¸on dont le bouleau pubescent (Betula pubescensEhrh. ssp. czerepa-novii(Orlova) Ha¨met-Ahti) pre´sent a` la limite des arbres et les cohortes de gaules situe´es au-dela` de la limite des arbres ont re´agi au re´chauffement de la dernie`re de´cennie dans six re´gions subarctiques du nord de l’Europe dont les caracte´risti-ques diffe`rent du point de vue du climat et de la paissance. Les re´sultats re´ve`lent diffe´rents patrons de re´ponse qui corres-pondent a` des situations ou` la limite des arbres progresse, demeure stationnaire ou possiblement re´gresse selon la re´gion. Le recrutement a e´te´ abondant au cours des dernie`res de´cennies dans toutes les re´gions, a` l’exception d’une re´gion caracte´-rise´e par des conditions essentiellement arctiques ou` les cohortes d’arbres et de gaules e´taient vieilles. Dans les re´gions ou` la pre´cipitation annuelle est e´leve´e, les populations de bouleau progressent; elles sont caracte´rise´es par de jeunes individus et des distributions d’aˆge des arbres et des gaules qui se chevauchent partiellement. Dans les re´gions ou` il y a des districts d’e´levage de rennes, les populations de bouleau sont stationnaires ou en re´gression et sont caracte´rise´es par des distribu-tions d’aˆge qui ne se chevauchent pas et une faible survie des gaules. Les patrons de recrutement au-dela` de la limite des arbres correspondaient ge´ne´ralement aux variables climatiques de la saison dormante, principalement la pre´cipitation; ce qui illustre l’importance d’un couvert nival protecteur. Les re´sultats font ressortir d’importantes interactions entre les fac-teurs abiotiques et biotiques qui controˆlent l’empie`tement sur la toundra et la dynamique de la limite des arbres ainsi que l’importance d’e´tudier plusieurs re´gions lorsqu’on s’inte´resse aux re´ponses qu’engendre le re´chauffement climatique.
[Traduit par la Re´daction]
Introduction
Over the past half century, temperatures in arctic and sub-arctic regions have increased 1–28C, but with large regional variations (Arctic Council and the International Arctic
Sci-ence Committee (IASC) 2005; Intergovernmental Panel on Climate Change (IPCC) 2007). This temperature increase is predicted to continue during the twenty-first century, with best estimates ranging between 1.8 8C and 4.0 8C, depend-ing on the model used (IPCC 2007). A general agreement
Received 11 February 2010. Accepted 24 March 2010. Published on the NRC Research Press Web site at cjfr.nrc.ca on 3 February 2011.
S. Aune2and L. So¨derstro¨m.Norwegian University of Science and Technology, NO-7491 Trondheim, Norway.
A. Hofgaard.3Norwegian Institute for Nature Research, NO-7485 Trondheim, Norway.
1This article is a contribution to the series Tree recruitment, growth, and distribution at the circumpolar forest–tundra transition. 2Present address: Norwegian Institute for Agricultural and Environmental Research, NO-8860 Tjøtta, Norway.
among scenarios has caused a growing interest in how pre-dicted climate changes might affect the circumpolar forest– tundra boundary (Callaghan et al. 2002; Arctic Council and the IASC 2005; Harsch et al. 2009). The boundary, through its circumpolar distribution, has importance at regional to global scales for its contribution to land–atmosphere interac-tions, C sequestration, and biodiversity (Callaghan et al. 2002; Goodale et al. 2002; Arctic Council and the IASC 2005). One of the most important vegetation shifts in the arctic (observed and predicted) is the expansion of trees and shrubs (Sturm et al. 2001; Forbes et al. 2009; Kullman and O¨ berg 2009). A region-wide tree cover change would re-structure the present tundra and its current services (Chapin et al. 2005; Post and Pedersen 2008).
One distinct component of the forest–tundra boundary is the treeline (i.e., latitudinal and (or) altitudinal limit for tree-sized growth; Holtmeier 2003). This ‘‘line’’ is generally considered a temperature-restricted frontier beyond which tree seedlings may establish but do not reach tree size be-cause of limitations to photosynthesis and growth rate and the destruction of leading shoots (Grace et al. 2002; Sveinb-jo¨rnsson et al. 2002; Holtmeier 2003). Thus, the treeline lo-cation is widely regarded as being sensitive to climate warming, and advances in response to climate change are frequently reported (e.g., Caccianiga and Payette 2006; Wang et al. 2006; Devi et al. 2008). The current treeline ecotone may respond to increased temperature by densifica-tion of the current scattered tree layer (Danby and Hik 2007; Batllori and Gutierrez 2008) and by advance of the existing treeline (Kullman and O¨ berg 2009). An advance is initially associated with a change in height growth of previously es-tablished individuals, but further relocation is dependent on new establishment beyond the current treeline (Kullman 2002; Caccianiga and Payette 2006; Hofgaard et al. 2009). Successful establishment of new advanced individuals de-pends on production, dispersal, and germination of seeds and long-term survival of the emerging seedlings into sap-ling and tree stages. Although temperature is an important factor structuring the treeline ecotone, any change is con-trolled by interplay of both abiotic and biotic factors, includ-ing precipitation, snow cover and duration, topography, and herbivory (Cairns and Moen 2004; Holtmeier and Broll 2005; Sturm et al. 2005). Increased abundance of trees and tree saplings in response to warming will enhance snow-trapping during winter and thus further promote tree growth and establishment (Dalen and Hofgaard 2005; Sturm et al. 2005). However, herbivore activity may inhibit warming-initiated responses (Post and Pedersen 2008; Olofsson et al. 2009; Hofgaard et al. 2010).
The forest–tundra boundary in Scandinavia and the Kola Peninsula is dominated by mountain birch (Betula pubescens Ehrh. ssp. czerepanovii (Orlova) Ha¨met-Ahti), and there is evidence of advancing treelines during the 20th century. Es-tablishment of new birch individuals peaked in the early to mid-20th century in response to climate warming and led to both densification of the birch forest (Kullman 2001) and a peak in establishment of the trees making up the current treeline (Dalen and Hofgaard 2005). The early 20th century warm period was followed by a colder period, 1950s to 1980, characterized by a decline in recruitment and in-creased mortality of saplings (Kullman 2001). Since the late
1980s, annual temperature has increased by about 1.58C but with different effects on treeline position and recruitment between geographical regions (Moen et al. 2008; Hofgaard et al. 2009; Kullman and O¨ berg 2009).
In this study, we use age structure and recruitment pattern of both the current treeline markers (individual treeline trees) and tree saplings beyond the treeline in northern Nor-way and the Kola Peninsula, northwestern Russia, to analyse geographical variation in forest–tundra ecotone responses to recent climate change. Three climatic regions located along the Atlantic Ocean – Arctic Ocean air mass gradient, with decreasing Atlantic air mass and increasing Arctic air mass impacts from west to east (northwestern Norway to the mid-dle of the Kola Peninsula), were selected for the study. In addition to age structure and recruitment pattern data, we used data on tree, sapling, and site characteristics to address the following questions. Do recruitment periods of trees and saplings at and beyond the treeline differ along the Atlantic– Arctic air mass gradient? Is the generally expected climate-driven tundra encroachment supported by current treeline lo-cations and sapling cohort patterns beyond treeline? Are cur-rent sapling cohorts showing a climate–establishment relationship, and if so, does it vary between climate regions?
Methods
Study areas and climate
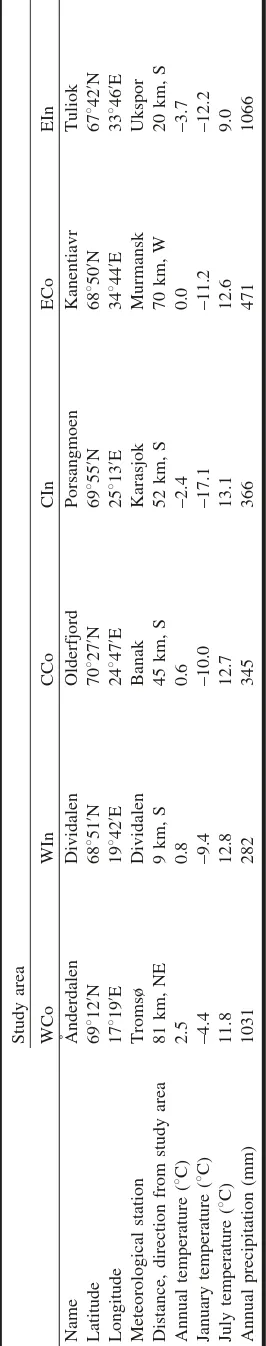
In each region along the Atlantic–Arctic air mass gra-dient, one coastal (Co) and one inland (In) area were used. Two regions are located in northern Norway and one in northwestern Russia, and these are henceforward called western (W), central (C), and eastern (E), respectively (Fig. 1). The western coastal area, A˚ nderdalen (WCo), is sit-uated on the island Senja in Troms County and is character-ized by a rather rugged mountain terrain with surrounding mountain peaks up to 800 m above sea level (a.s.l.), whereas the terrain in the western inland area, Dividalen (WIn), is slightly more gentle but with mountain peaks reaching up to 1500 m a.s.l. The central areas Olderfjord (CCo) and
Por-Fig. 1.Location of the study areas A˚ nderdalen (WCo), Dividalen
sangmoen (CIn), Finnmark County, are situated in a land-scape characterized by rounded mountains. Mountain peaks in these areas reach 400–500 m a.s.l. and 800–1000 m a.s.l., respectively. The eastern inland area, Tuliok (EIn), is lo-cated in the Khibiny Mountains, which is the largest massif on the Kola Peninsula, with several peaks reaching 900– 1100 m a.s.l. Whereas these five areas are located at the transition from forest to subarctic alpine tundra, the coastal easternmost area (Kanentiavr; ECo) is situated at the transi-tion from forest to arctic tundra in a landscape characterized by fairly flat terrain with scattered mires, lakes, and small hills. All six areas are used as foraging domains by a num-ber of herbivores such as moose, hare, grouse, various ro-dents, and insects. In addition, the areas in the central region and western inland area belong to the main reindeer herding districts of northern Norway used for summer graz-ing by large semidomestic reindeer herds.
According to Moen (1999), climate conditions are charac-terized as slightly oceanic in WCo, indifferent in CCo, ECo, and EIn, and slightly continental in WIn and CIn. The west-ern coastal area, which is located close to the Atlantic Ocean, is characterized by high annual precipitation and rather high mean annual temperature (Table 1). Mean annual temperature and mean temperature during the coldest month, January, decrease from the west to east and from the coast to inland areas. The mean temperature for the warmest month, July, shows no west–east gradient and only a weak coast–inland gradient for western and central areas (warmer inland). In the eastern region, the coastal area shows a nota-bly higher mean July temperature and lower annual precipi-tation than the inland area (Table 1).
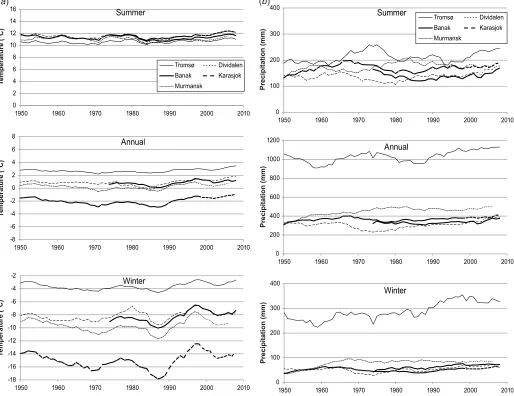
Annual temperature in the regions has increased by ca. 1– 28C since the late 1980s, with emphasis on winter temper-ature (Fig. 2a). Annual precipitation shows an overall weak increasing trend during the same time period, except in the western coastal area because of summer precipitation de-crease in that area (Fig. 2b).
Mountain birch is the dominating tree layer species in the treeline ecotone in all areas. This species is a favoured food source, and birch leaves, buds, and twigs constitute an im-portant part of the diet for many animal species. Birch has an annual seed production with germability up to 60% at its distribution limit across the studied region (A. Hofgaard, un-published data). The shrub layer in the western coastal area is dominated by dwarf birch (Betula nana L.); dwarf birch and willows (Salix spp.) dominate in the other five areas. The field layer in the western inland area is dominated by sedges and low herbs, but in the other five areas, deciduous and evergreen dwarf shrubs dominate together with low herbs.
Sampling design
In each of the six areas, one gentle north-facing mountain slope was chosen for age determination and site characteris-tics of treeline trees and tree recruitment above and (or) be-yond the treeline. The sampling was conducted during summer in 2007 and 2008. In each area, 20 trees from the local treeline (defined as the most advanced trees with a minimum height of 2 m) were selected (Table 2; Fig. 3). The minimum distance horizontally between selected trees
was set to 10 m. These treeline trees were cored at the base Table
Fig. 2.Ten-year running mean for (a) summer, annual, and winter temperatures and (b) total summer, annual, and winter precipitation. Tromsø, Dividalen, Banak, and Karasjok meteorological stations represent study areas WCo, WIn, CCo, and CIn, respectively. Murmansk represents both ECo and EIn. Details for meteorological stations are given in Table 1. Data are from the Norwegian Meteorological Institute (http://www.met.no) and Anonymous (2008).
Table 2.Characteristics for sampled treeline trees in the six study areas (mean values and standard deviations per area are given for all
measured and calculated variables).
Study area
Variable WCo WIn CCo CIn ECo EIn
Altitude (m above sea level) 306 626 310 348 244 534
No. of trees sampled 21 20 20 20 20 20
No. of trees included in age structure: 0 m
17 8 19 17 19 19
No. of trees included in age structure: 2 m
20 16 20 20 18 20
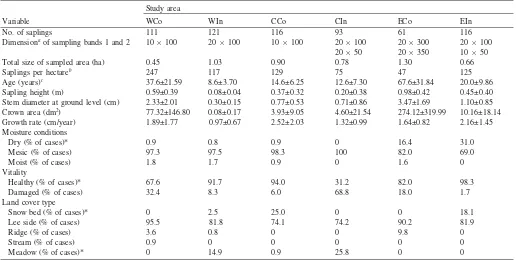
Age at stem base 66.5±29.1 92.4±9.9 52.1±14.8 54.3±19.3 70.8±28.1 33.4±8.0
Age at 2 m 16.8±15.3 59.9±16.0 26.2±12.9 31.1±16.1 24.9±18.2 4.9±1.9
Years to become 2 m 48.1±29.4 32.8±19.3 26.3±8.4 25.7±8.7 46.9±24.0 28.5±7.6
Annual growth rate 0–2 m (cm/year) 5.8±3.4 7.5±2.9 8.3±2.5 9.1±5.3 5.8±4.1 7.6±2.4
Tree height (m) 2.9±1.0 5.8±1.4 4.4±1.1 4.0±1.6 3.0±0.6 2.6±0.4
Stem diameter at 0 m (cm) 9.2±4.8 18.8±5.1 9.7±4.0 10.8±6.3 8.4±3.4 5.1±1.4
Stem diameter at 1.3 m (cm) 3.7±3.3 14.1±5.5 6.0±3.4 6.4±3.4 4.3±2.2 1.8±0.6
(0 m) and at 2 m aboveground for age determination. Trees too thin to be cored at the 2 m level were sampled by cut-ting at that height.
Above and (or) beyond the treeline, all birch saplings (<2 m) within one 10 or 20 m wide sampling band stretch-ing from the sampled treeline and 100 altitudinal metres into the tundra were sampled. Each sampling band started at a sampled treeline tree. The width of the band was determined by local sapling density. An additional band spanning 50 al-titudinal metres was used in areas with low sapling density. Details for each area are given below and in Table 3. In the flat area of Kanentiavr (ECo), two bands 20 m wide 300 and 350 m long, respectively, stretching from the treeline to-wards the north, were used. Because of the different widths (and number) of sampling bands and differences in moun-tain steepness in the six areas, the total sampled area dif-fered between study areas (Table 3). In addition to the sampling within the bands, the highest sapling location was recorded for each mountain slope (except for ECo), and the specimen was sampled.
All saplings were cored (basal stem diameter > 3.5 cm) or cut at the base for age determination. The following varia-bles were recorded for each sampled sapling and treeline tree: height, stem diameter at ground level and, when avail-able, at breast height (1.3 m), crown diameter (two perpen-dicular measures), vitality, ground moisture conditions, land cover type, and GPS position. The vitality was classified as healthy, dead, and damaged by climate, herbivores, or fungi; ground moisture classes used were dry, mesic, and moist; and land cover types were classified as stream, stagnant water, rocky outcrop, meadow, lee side, ridge, and snow bed (i.e., late-laying snow locations).
Core samples were mounted on a wooden support, dried, and brought to the lab, together with cut samples, where their age was determined using a stereomicroscope (6– 40). To increase the contrast between late and early wood, the samples were smoothed with a scalpel and zinc ointment was applied. In total, 121 treeline trees and 618 saplings were sampled and analysed. Stem rot or failure to hit the pith precluded exact age determination of some specimens. For samples with an estimated two–five annual rings miss-ing (estimated by eye usmiss-ing the density of the rmiss-ings near the pith and the approximate distance to the pith), the estimated number was added to the counted age. This was the case for 14.0% and 11.6% of the treeline tree age samples from the 0 m and 2 m levels, respectively, and 1.9% of the sapling samples. Samples with larger estimated missing numbers of rings were omitted from the age structure analysis. The total data set was used in analyses, not including age. The num-ber of treeline trees included in age analyses ranged from eight to 19 (Table 2). Of the 618 sampled saplings, four, two, three, and two samples from WCo, CIn, ECo, and EIn, respectively, were omitted from the age analysis.
Data handling and analyses
The age data for each study area were grouped into classes of 5 years, and height data were grouped into classes of 10 cm. Because of difficulties in finding small individuals (<5 cm) in the field, low numbers of saplings in the young-est age classes and smallyoung-est height classes were expected. The crown area of the trees and saplings was calculated by using following formula:
½1 Crown area¼p½ðdiameter 1þdiameter 2Þ=42
Fig. 3.View of the Dividalen study area (WIn), which is used as summer grazing grounds for reindeer and is characterized by old treeline
This formula uses the two measured diameters of tree and sapling crowns to calculate the mean crown diameter and subsequently the area of the crown by using the formula for an ellipse.
From the treeline age data, the following variables were extracted: age at 0 m, age at 2 m, number of years to be-come tree-sized (age at 0 m – age at 2 m), year of establish-ment, year when the 2 m level was reached, and annual growth rate from 0 m to 2 m (2 m divided by number of years to become tree-sized). For saplings, altitudinal dis-tance from each sampled specimen to the treeline tree at the lower end of the sampling band was calculated using re-corded GPS positions. For the eastern coastal area (ECo), the horizontal distance to the treeline was used. The density of saplings in each area (saplings per hectare) was calcu-lated using number of saplings divided by GPS-recorded horizontal length of the 100 m sampling band multiplied by band width in each area. In the eastern coastal area, both sampling bands were used. Growth rate of saplings (centi-metres per year) was calculated as height divided by age.
For better approximation to normal distribution, the varia-bles age, height, basal diameter, and crown area were log-transformed. Correlations between these variables were tested for using Pearson’s product moment analysis; Spear-man’s rank analysis was used for correlations with distance to treeline, which was non-normally distributed. Differences between study areas were tested for by the use of ANOVA and Tukey’s HSD test for normally distributed variables (log-transformed age, living height, crown area, and basal
stem diameter). Kruskal–Wallis and Berhens–Fisher tests in the npmc (nonparametric multiple comparisons) R-package (R Development Core Team 2007) were used for the non-normally distributed variable distance to treeline. Differen-ces between study areas for each class of the variables mois-ture condition, vitality, and land cover type were tested for using the c2 test with percentage of saplings in each class as input variables.
Principle component analysis (PCA) was used to investi-gate the relationship between sapling establishment and cli-mate (monthly mean temperature and monthly total precipitation for the nearest meteorological station providing monthly data). The period 1940–2007 was used for WIn, CIn, ECo and EIn, and 1966 to 2007 was used for WCo and CCo because of variation in the length of available monthly climate data representing individual areas. Climate data from Murmansk were used for both ECo and EIn as this was the only station with available monthly data for the eastern region.
Results
Treeline
The treeline is located at a higher altitude in inland areas than in coastal areas, but the altitudinal difference varies be-tween the three regions. The differences were 320 and 290 altitudinal metres in the west and east, respectively, but only 38 m in the central region (Table 2). Mean age of sampled treeline trees ranged from 33.4 to 92.4 years, with
Table 3.Characteristics for sampling bands and sampled saplings in the six study areas.
Study area
Variable WCo WIn CCo CIn ECo EIn
No. of saplings 111 121 116 93 61 116
Dimensionaof sampling bands 1 and 2 10100 20100 10100 20100 20300 20100
2050 20350 1050
Total size of sampled area (ha) 0.45 1.03 0.90 0.78 1.30 0.66
Saplings per hectareb 247 117 129 75 47 125
Age (years)c 37.6±21.59 8.6±3.70 14.6±6.25 12.6±7.30 67.6±31.84 20.0±9.86
Sapling height (m) 0.59±0.39 0.08±0.04 0.37±0.32 0.20±0.38 0.98±0.42 0.45±0.40
Stem diameter at ground level (cm) 2.33±2.01 0.30±0.15 0.77±0.53 0.71±0.86 3.47±1.69 1.10±0.85
Crown area (dm2) 77.32±146.80 0.08±0.17 3.93±9.05 4.60±21.54 274.12±319.99 10.16±18.14
Growth rate (cm/year) 1.89±1.77 0.97±0.67 2.52±2.03 1.32±0.99 1.64±0.82 2.16±1.45
Moisture conditions
Dry (% of cases)* 0.9 0.8 0.9 0 16.4 31.0
Mesic (% of cases) 97.3 97.5 98.3 100 82.0 69.0
Moist (% of cases) 1.8 1.7 0.9 0 1.6 0
Vitality
Healthy (% of cases)* 67.6 91.7 94.0 31.2 82.0 98.3
Damaged (% of cases) 32.4 8.3 6.0 68.8 18.0 1.7
Note:Asterisks indicate levels of significance (*,p< 0.05) between study areas for classes of moisture conditions, vitality, and land cover type.
aWidth (m)
length in altitudinal metres, except in ECo where horizontal metres are used.
bBased on band 1 in all but ECo, where both bands were used.
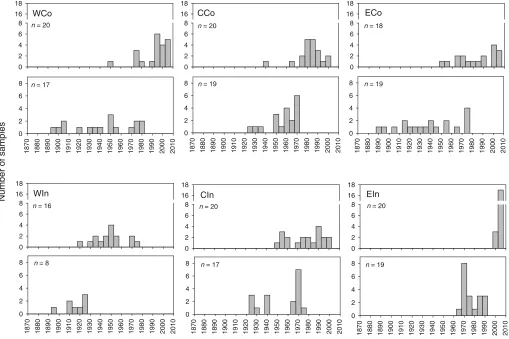
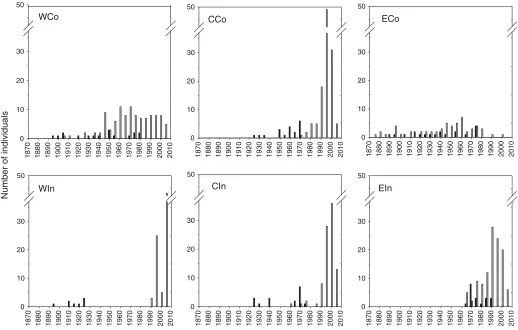
the youngest trees found in the eastern inland area and the oldest found in the western inland area (Table 2). The period of establishment of treeline trees differed noticeably be-tween areas, with establishment over a long period, 1890 to 1980, in WCo and ECo, over short periods in WIn and EIn (ca. 30 years in the first and second half of the 20th century, respectively), and over an intermediate period in the central areas (during ca. 50 years in the mid-20th century) (Fig. 4). A majority of the trees in all areas reached tree size during the last three decades, except in WIn where the majority reached tree size more than 40 years ago. In EIn, all treeline trees were established during the last 45 years and reached tree size less than 15 years ago.
The treeline trees in WIn had the largest stem diameter, crown area, and average tree height (5.8 m) (Table 2). The smallest trees, with an average height of 2.6 m, were found in EIn. Mean annual growth rates from 0 to 2 m were higher in the inland areas than in the coastal areas, and the central region showed a higher overall growth rate than the western and eastern regions (Table 2).
The dominating vitality class for the treeline trees was ‘‘healthy’’ in WCo, CCo, and EIn, whereas 45% and 95% of the treeline trees in WIn and CIn, respectively, were af-fected by herbivores. In ECo, 65% of the treeline trees
showed evidence of damage by climate. Further, the lee side was the most common land cover type among the trees, although 35% of the trees in both WIn and CIn were found in meadow locations. In WCo, CCo, and EIn, 14%, 35%, and 25% of the trees, respectively, were growing in snow bed locations.
Saplings
Spatial distribution
The number of saplings sampled in each of the study areas ranged from 61 in ECo to 121 in WIn (Table 3), and the number of saplings per hectare ranged from 47 (ECo) to 238 (WCo). The saplings with most advanced altitudinal po-sitions were found at 181, 158, 122, 145, and 398 altitudinal metres above the present treeline in WCo, WIn, CCo, CIn, and EIn, respectively. In WCo and CCo, the highest located sapling was found only a few altitudinal metres below the local summit. Most of the saplings were found on the lee sides, although one-fourth of the individuals in CCo and CIn were found in snow bed and meadow locations, respec-tively (Table 3). Mesic ground moisture conditions at sap-ling locations dominated in all areas. Most of the sapsap-lings were classified as healthy, but 69% and 26% of the
individ-Fig. 4.Age distribution of treeline trees at the stem base (0 m level; lower figure sections) and at the 2 m level (upper figure sections) for
uals sampled in CIn and WCo, respectively, showed evi-dence of herbivore damage.
Age and size structure
Mean sapling age ranged from 8.6 years (WIn) to 67.6 years (ECo) and was higher in coastal areas than in in-land areas within each region (Table 3). In CCo and all three inland areas, no individuals older than 50 years were found, whereas WCo and ECo populations consisted of indi-viduals up to 102 and 149 years old, respectively (Fig. 5). The youngest sapling populations were found in WIn, CCo, and CIn, with recruitment peaks at the end of the 1990s and beginning of the 2000s. In EIn, recruitment peaked one dec-ade earlier. WCo and ECo showed no distinct recruitment peaks, but continuous recruitment of about five to 10 indi-viduals per age class from the 1950s until present was found in WCo, and continuous establishment of one to eight sap-lings per age class from the 1870s until the 1980s was found in ECo. In the latter area, only two individuals (3.3%) were younger than 25 years.
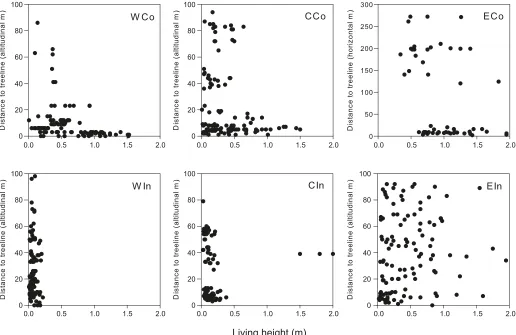
Three areas, WIn, CCo and CIn, show nonoverlapping age distributions for treeline trees and saplings (Fig. 5). The WCo and EIn areas show partly overlapping age distributions, whereas there is total overlap in age of the two groups in ECo. Sapling height and altitudinal distribution differed be-tween areas (Fig. 6). WIn and CIn were characterized by small individuals, <0.3 m and <0.5 m, respectively (except
for three >1.5 m individuals in CIn located in a sheltered habitat surrounded by willows), with no relation to the alti-tudinal gradient in WIn (Table 4). WCo and CCo saplings were all <1.5 m, with the majority found close to the tree-line. Their height decreased with increasing distance to the treeline, but this was only significant in WCo (Table 4). The sapling cohorts of the eastern areas were characterized by a broad height distribution with individuals up to ca. 2 m, but saplings <0.3 m were lacking in ECo. Both areas showed a fairly random sapling height distribution in rela-tion to distance to treeline, although EIn had significantly (weak) taller saplings further away from the treeline (Table 4), and ca. 60% of the ECo saplings were growing less than 30 m from the treeline (Fig. 6).
Mean sapling growth rate among study areas ranged from 0.97 cm/year (WIn) to 2.52 cm/year (CCo) (Table 3), with individual growth rates varying among and within areas (lowest growth rate was 0.06 cm/year in CIn, and the high-est was 14.6 cm/year in WCo).
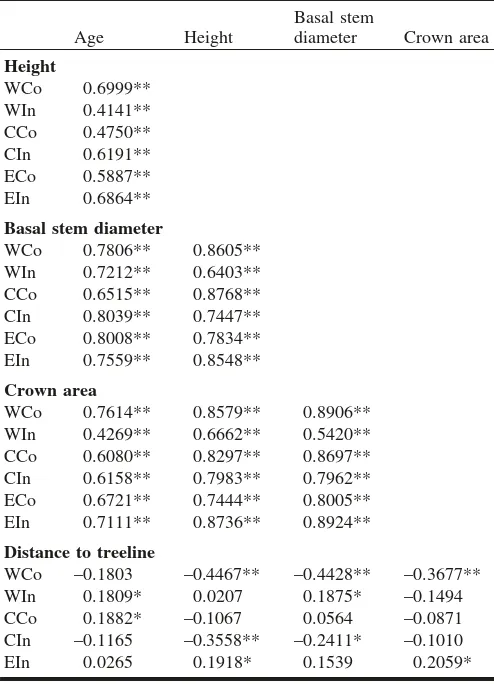
There was a significant positive correlation between sap-ling age, height, basal stem diameter, and crown area in all areas (Table 4), but significant differences among all areas were found for age (F = 146.6, p < 0.01), height (F = 115.0,p< 0.01), basal stem diameter (F = 137.8,p< 0.01), crown area (F = 181.6, p < 0.01), and distance to treeline (c2= 491.79,p< 0.01). Multiple comparisons between pairs of study areas showed nonsignificant differences between
Fig. 5.Age distribution of saplings (shaded bars) and treeline trees (solid bars) in the six study areas, divided into age classes of 5 years.
CCo and CIn for age (p = 0.14), basal stem diameter (p = 0.67), and distance to treeline (p= 0.07), between EIn and CCo for height (p = 0.35), and between WIn and CIn for distance to treeline (p = 0.06). Further, height, basal stem diameter, and crown area in WCo, height and basal stem di-ameter in CIn, and age in WIn and CCo all decreased along the altitudinal gradient (i.e., with increasing distance to tree-line; Table 4).
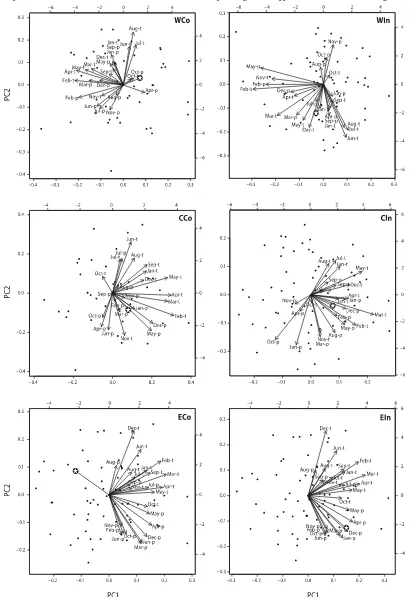
Sapling establishment and climate
Main monthly climate variables corresponding with sap-ling establishment varied between study areas, but impor-tance of non-growing-season months are apparent in all areas (Appendix A, Fig. A1). However, summer tempera-tures (June–August) showed positive correlation with sap-ling establishment in both western areas, even if early winter temperature and precipitation are more closely asso-ciated with establishment in these areas. Sapling establish-ment in both central areas shows a positive relationship to winter and early growing season (May) precipitation as main variables. In addition, February and March tempera-tures are important in CIn. The two eastern sites show al-most opposite correspondence patterns with tested climate variables. EIn sapling establishment corresponds positively with winter and spring precipitation, and ECo corresponds negatively with both winter–spring precipitation and early winter (October) temperature.
Discussion
Treeline
Tree and shrub encroachment of tundra is generally as-sumed to be a consequence of current and future warming at high latitudes (Arctic Council and the IASC 2005; IPCC 2007). Although all from a restricted part of the circumpolar north, the present results highlight the importance of multi-ple study sites representing a diversity of climate sections of the subarctic region to test, adjust, or verify this general as-sumption for a basically climate-driven response at the arctic forest–tundra transition zone. In addition to climate, land-use differences among subarctic regions have to be ac-counted for (Cairns and Moen 2004; Holtmeier and Broll 2007; Olofsson et al. 2009). An increasing number of stud-ies points towards the important role that herbivores play in structuring the current tundra and the treeline zone (Moen and Lagerstro¨m 2008; Post and Pedersen 2008; Hofgaard et al. 2010).
Although the entire study region has experienced a warm-ing trend since the late 1980s and all study areas are situated close to the northern distribution limit of the focal species, studied treelines show different response patterns. Based on the differences in age structure and recruitment to the tree layer, both advancing (WCo and EIn) and stationary (WIn, CCo, and CIn) treelines, and possibly also retreating tree-lines (ECo), were found. Similar results are reported from a
Fig. 6.Altitudinal distance (elevation, metres) of saplings to treeline plotted against sapling height for the study areas. For the flat eastern
78long latitudinal gradient in Scandinavia (Dalen and Hof-gaard 2005). The present study provides strengthened evi-dence for an additional nonclimatic treeline-structuring factor, regionally overriding climate. In the two areas with high annual precipitation (western coastal and eastern in-land), treeline trees were small and healthy and had reached tree size recently. This is indicative of advancing tree popu-lations. However, the age structure for time of establishment points to an advancing position only in the eastern inland area where the trees are young and have reached tree size during the last decade(s). Further, age structure of coastal areas indicates stationary treeline conditions, as inferred from the high age and broad age span of trees, or possibly a slow advance in the west and east as the number of individ-uals reaching tree size has increased somewhat during last decade(s). A J-shaped age distribution at the 2 m level indi-cates a sustained supply of treeline trees, which is typical of a stationary treeline where tree recruitment and mortality are balanced over time (Dalen and Hofgaard 2005). A deviating area is the western inland area characterized by old trees and no recruitment at tree size level since the 1970s. This
pat-tern indicates treeline recession, possibly due to considerable browsing pressure.
Tundra encroachment
Conclusions drawn from analyses of treeline trees are only to some extent supported by the sapling data. Recruit-ment at and beyond the treeline is a crucial trait of ecotone response to climate. Thus, analysis of sapling pool changes is an important tool in predicting potential for treeline ad-vance. However, knowledge of long-term survival of recruits and time required to reach tree size is essential to tune gen-eral scenarios (Arctic Council and the IASC 2005; IPCC 2007) into empirical-based predictions.
If only based on current sapling cohorts, areas dominated by young and small individuals (e.g., the western inland area) could be interpreted as representing a recent (and likely climate driven) change towards increased tundra en-croachment (Kullman 2002; Truong et al. 2007). However, as seed availability is a nonrestricting factor in studied re-gions, the nonoverlapping age distributions of treeline trees and saplings in the western inland and central areas (less clear) indicate prevailing limitations to survival and devel-opment into tree size. As these areas are main summer graz-ing grounds of reindeer, a reasonable conclusion points towards a herbivory-linked control at the sapling stage (Cairns and Moen 2004; Dalen and Hofgaard 2005) inhibit-ing climate-driven saplinhibit-ing expansion under the present graz-ing regime (Hofgaard et al. 2009; Olofsson et al. 2009). This also agrees with the assumption of seedling growth and sur-vival as the main limiting factor for establishment of new birch individuals rather than dispersal and germination of seeds (Kullman 1993).
In the eastern costal area, characterized by arctic condi-tions with very little influence from moist and warm Atlan-tic air masses, sapling data contradict the treeline indication of a possible recent advance in accordance with general sce-narios. The area is characterized by a total overlap in age among saplings and trees, a random height distribution of sapling in relation to distance to treeline, few individuals in the smallest height classes, and a high frequency of climate-caused damage. This all indicates a climate-related station-ary situation or possibly a recession due to the very low number of individuals established during the last three deca-des. Most trees and saplings in this area had a multistemmed shrub-like growth form. This growth form is advantageous in cold harsh arctic environments as it ensures longevity of established individuals through growth of new stems, enhan-ces temperature near the individual through sheltering and effect on snow distribution (Grace et al. 2002; Sturm et al. 2005), and promotes recruitment close to existing individu-als (Sveinbjo¨rnsson et al. 2002).
In the western coastal and eastern inland areas, the abun-dant recruitment of saplings supports and corresponds well with the results from analyses of the treeline trees. Presence of saplings with heights close to tree size indicates a clear potential for relocation of the treeline under the current en-vironmental conditions. This potential for advance is em-phasised in the eastern inland area where 1.5 m tall saplings are present throughout the analysed altitudinal tundra section and also supported by the occurrence of saplings 400 m above the current treeline. Although these areas show clear
Table 4.Correlation coefficients between sapling age, height,
ba-sal stem diameter, crown areas, and distance to treeline (Pearson’s product-moment correlation for log-transformed age, height, basal stem diameter, and crown area; Spearman’s rank correlation for correlations between altitude and the other variables).
Age Height
WCo –0.1803 –0.4467** –0.4428** –0.3677**
WIn 0.1809* 0.0207 0.1875* –0.1494
CCo 0.1882* –0.1067 0.0564 –0.0871
CIn –0.1165 –0.3558** –0.2411* –0.1010
EIn 0.0265 0.1918* 0.1539 0.2059*
evidence of recent tundra encroachment, the overall sapling growth rate is only around 2 cm/year, which gives a time span of a century from seed germination to a tree-sized indi-vidual. However, individual growth rates vary greatly within areas, permitting a more rapid advance even under current environmental conditions.
Establishment and climate
An important climate-related factor for successful estab-lishment is favorable microconditions, including both micro-climate and microtopography (Kja¨llgren and Kullman 1998; Hofgaard et al. 2009). Ridges and exposed microsites with bare soil favor initial establishment but provide harsh and disadvantageous environments for long-term survival (Dalen and Hofgaard 2005; Anschlag et al. 2008), leading to the re-corded situation in which most saplings growing in lee-side locations with mesic moisture conditions. This topoclimati-cally favored pattern is linked to the presence of a protecting snow cover throughout the winter, which in addition enhan-ces winter soil temperatures and moisture supply in the spring (Sturm et al. 2001; Bekker 2005). It is often assumed that the age demography of birch saplings beyond the tree-line corresponds with higher temperatures in the years of es-tablishment (Kullman 2002). This is partly shown for the western areas, but across studied areas, most correspondence signals are for non-growing-season precipitation variables. Precipitation in the non-growing season mainly comes as snow, thus leading to increased temporal stability of a needed uninterrupted duration of the protecting snow cover throughout the winter (Sturm et al. 2005; Hofgaard et al. 2009). However, protection-induced growth enhancement could be counteracted by the reduction in growing-season length caused by the increased snowpack (Shutova et al. 2006; Karlsen et al. 2007). This trade-off between protection and growing-season length varies among areas as indicated by the variation in correspondence with non-growing-season variables.
Concluding remarks
The lack of synchronous recruitment trends, despite a syn-chronous warming trend (as in present study), suggests the presence of different local factors modulating treeline move-ments and tundra encroachment. The results highlight the important and largely overlooked influence of herbivores and the interplay between abiotic and biotic control over tundra encroachment and treeline dynamics. When not con-sidered, response rate to climate warming at forest margins in the circumpolar north could be overestimated. Given the heterogeneous response of treeline areas, the vast expanse of the circumpolar forest–tundra zone, its relevance for tun-dra diversity, and feedbacks resulting in atmospheric heat-ing, analyses of interacting abiotic and biotic factors in multisite studies need further attention.
Acknowledgements
The authors thank Staffan Dova¨rn, Maria Golubeva, Kse-nia Kozlova, Ingrid E. Mathisen, and Erin Sandberg for as-sistance in the field, Mikhail Zimin and Yurate Plyushkyiavichyute for help with Russian climate data, Kari Sivertsen for help with figures, and two anonymous re-viewers for helpful comments on the draft version of the
pa-per. The paper is a product under the IPY core project PPS Arctic (http://ppsarctic.nina.no) financially supported by the Research Council of Norway through grants 176065/S30 and 185023/S50 to A.H.
References
Arctic Council and the International Arctic Science Committee. 2005. Arctic Climate Impact Assessment. Cambridge University Press, Cambridge, UK.
Anonymous. 1988. Nauchnyj i prakticheskij spravochnik po kli-matu SSSR. Gidrometeoizdat, Leningrad, Russia.
Anonymous. 2008. Federalnaya sluzhba Rossii po gidrometeorolo-gii i monitoringu okruzhajuschei spedy. Roshydromet, Moscow, Russia.
Anschlag, K., Broll, G., and Holtmeier, F.K. 2008. Mountain birch seedlings in the treeline ecotone, subarctic Finland: variation in above- and below-ground growth depending on microtopogra-phy. Arct. Antarct. Alp. Res.40(4): 609–616. doi:10.1657/1523-0430(07-087)[ANSCHLAG]2.0.CO;2.
Batllori, E., and Gutierrez, E. 2008. Regional tree line dynamics in response to global change in the Pyrenees. J. Ecol.96(6): 1275– 1288. doi:10.1111/j.1365-2745.2008.01429.x.
Bekker, M.F. 2005. Positive feedback between tree establishment and patterns of subalpine forest advancement, Glacier National Park, Montana, USA. Arct. Antarct. Alp. Res. 37(1): 97–107. doi:10.1657/1523-0430(2005)037[0097:PFBTEA]2.0.CO;2. Caccianiga, M., and Payette, S. 2006. Recent advance of white
spruce (Picea glauca) in the coastal tundra of the eastern shore of Hudson Bay (Quebec, Canada). J. Biogeogr. 33(12): 2120– 2135. doi:10.1111/j.1365-2699.2006.01563.x.
Cairns, D.M., and Moen, J. 2004. Herbivory influences tree lines. J. Ecol.92(6): 1019–1024. doi:10.1111/j.1365-2745.2004.00945.x. Callaghan, T.V., Crawford, R.M.M., Eronen, M., Hofgaard, A.,
Payette, S., Rees, W.G., Skre, O., Sveinbjo¨rnsson, B., Vlassova, T.K., and Werkman, B.R. 2002. The dynamics of the tundra– taiga boundary: an overview and suggested coordinated and in-tegrated approach to research. Ambio Special Report,12: 3–5. Chapin, F.S., 3rd, Sturm, M., Serreze, M.C., McFadden, J.P., Key,
J.R., Lloyd, A.H., McGuire, A.D., Rupp, T.S., Lynch, A.H., Schimel, J.P., Beringer, J., Chapman, W.L., Epstein, H.E., Eu-skirchen, E.S., Hinzman, L.D., Jia, G., Ping, C.-L., Tape, K.D., Thompson, C.D.C., Walker, D.A., and Welker, J.M. 2005. Role of land-surface changes in arctic summer warming. Science (Washington, D.C.), 310(5748): 657–660. doi:10.1126/science. 1117368. PMID:16179434.
Dalen, L., and Hofgaard, A. 2005. Differential regional treeline dy-namics in the Scandes Mountains. Arct. Antarct. Alp. Res.37(3): 284–296. doi:10.1657/1523-0430(2005)037[0284:DRTDIT]2.0. CO;2.
Danby, R.K., and Hik, D.S. 2007. Variability, contingency and ra-pid change in recent subarctic alpine tree line dynamics. J. Ecol. 95(2): 352–363. doi:10.1111/j.1365-2745.2006.01200.x. Devi, N., Hagedorn, F., Moiseev, P., Bugmann, H., Shiyatov, S.,
Mazepa, V., and Rigling, A. 2008. Expanding forests and chan-ging growth forms of Siberian larch at the Polar Urals treeline during the 20th century. Glob. Change Biol. 14(7): 1581–1591. doi:10.1111/j.1365-2486.2008.01583.x.
Forbes, C.B., Macias Fauria, M., and Zetterberg, P. 2009. Russian arctic warming and ‘greening’ are closely tracked by tundra shrub willows. Glob. Change Biol. 16(5). doi: 10.1111/j.1365-2486.2009.02047.x.
S.R., Nabuurs, G.J., Nilsson, S., and Shvidenko, A.Z. 2002. For-est carbon sinks in the Northern Hemisphere. Ecol. Appl.12(3): 891–899. doi:10.1890/1051-0761(2002)012[0891:FCSITN]2.0. CO;2.
Grace, J., Berninger, F., and Nagy, L. 2002. Impacts of climate change on the tree line. Ann. Bot. (Lond.), 90(4): 537–544. doi:10.1093/aob/mcf222.
Harsch, M.A., Hulme, P.E., McGlone, M.S., and Duncan, R.P. 2009. Are treelines advancing? A global meta-analysis of tree-line response to climate warming. Ecol. Lett. 12(10): 1040– 1049. doi:10.1111/j.1461-0248.2009.01355.x. PMID:19682007. Hofgaard, A., Dalen, L., and Hytteborn, H. 2009. Tree recruitment
above the treeline and potential for climate-driven treeline change. J. Veg. Sci. 20(6): 1133–1144. doi:10.1111/j.1654-1103.2009.01114.x.
Hofgaard, A., Løkken, J.O., Dalen, L., and Hytteborn, H. 2010. Comparing warming and grazing effects on birch growth in an alpine environment — a 10-year experiment. Plant Ecol. Diver-sity,3: 19–27. doi: 0.1080/17550871003717016.
Holtmeier, F.-K. 2003. Mountain timberlines: ecology, patchiness, and dynamics. Kluwer Academic Publishers, Dordrecht, the Netherlands.
Holtmeier, F.-K., and Broll, G. 2005. Sensitivity and response of northern hemisphere altitudinal and polar treelines to environ-mental change at landscape and local scales. Glob. Ecol. Bio-geogr. Lett. 14(5): 395–410. doi:10.1111/j.1466-822X.2005. 00168.x.
Holtmeier, F.-K., and Broll, G. 2007. Treeline advance — driving processes and adverse factors. Landscape Online, 1: 1–32. doi:10.3097/LO.200701.
Intergovernmental Panel on Climate Change. 2007. Climate change 2007: the physical science basis — summary for policymakers 10th session of Working group 1 of the IPCC. IPCC, Paris, France.
Karlsen, S.R., Solheim, I., Beck, P.S.A., Høgda, K.A., Wielgolaski, F.E., and Tømmervik, H. 2007. Variability of the start of the growing season in Fennoscandia, 1982–2002. Int. J. Biomet. 51(6): 513–524. doi:10.1007/s00484-007-0091-x.
Kja¨llgren, L., and Kullman, L. 1998. Spatial patterns and structure of the mountain birch tree-limit in the southern Swedish Scandes — a regional perspective. Geog. Ann. Ser. A Phys.80(1): 1–16. doi:10.1111/1468-0459.00023.
Kullman, L. 1993. Tree limit dynamics of Betula pubescens ssp. tortuosain relation to climate variability — evidence from cen-tral Sweden. J. Veg. Sci.4(6): 765–772. doi:10.2307/3235613. Kullman, L. 2001. 20th century climate warming and tree-limit rise
in the southern Scandes of Sweden. Ambio, 30(2): 72–80. PMID:11374309.
Kullman, L. 2002. Rapid recent range-margin rise of tree and shrub species in the Swedish Scandes. J. Ecol. 90(1): 68–77. doi:10. 1046/j.0022-0477.2001.00630.x.
Kullman, L., and O¨ berg, L. 2009. Post-Little Ice Age tree line rise and climate warming in the Swedish Scandes: a landscape eco-logical perspective. J. Ecol.97(3): 415–429. doi:10.1111/j.1365-2745.2009.01488.x.
Moen, A. 1999. National atlas of Norway: vegetation. Norwegian Mapping Authority, Hønefoss, Norway.
Moen, J., and Lagerstro¨m, A. 2008. High species turnover and de-creasing plant species richness on mountain summits in Sweden: reindeer grazing overrides climate change? Arct. Antarct. Alp.
Res. 40(2): 382–395. doi:10.1657/1523-0430(07-031)[MOEN]2.
0.CO;2.
Moen, J., Cairns, D.M., and Lafon, C.W. 2008. Factors structuring the treeline ecotone in Fennoscandia. Plant Ecol. Diversity,1(1): 77–87. doi:10.1080/17550870802246664.
Olofsson, J., Oksanen, L., Callaghan, T., Hulme, P.E., Oksanen, T., and Suominen, O. 2009. Herbivores inhibit climate-driven shrub expansion on the tundra. Glob. Change Biol.15(11): 2681–2693. doi:10.1111/j.1365-2486.2009.01935.x.
Post, E., and Pedersen, C. 2008. Opposing plant community re-sponses to warming with and without herbivores. Proc. Natl. Acad. Sci. U.S.A. 105(34): 12353–12358. doi:10.1073/pnas. 0802421105. PMID:18719116.
R Development Core Team. 2007. R: a language and environment for statistical computing. R foundation for Statistical Comput-ing, Vienna, Austria. ISBN 3-900051-07-0. Available at http:// www.R-project.org.
Shutova, E., Wielgolaski, F.E., Karlsen, S.R., Makarova, O., Ber-lina, N., Filimonova, T., Haraldsson, E., Aspholm, P.E., Flø, L., and Høgda, K.A. 2006. Growing seasons of Nordic mountain birch in northernmost Europe as indicated by long-term field studies and analyses of satellite images. Int. J. Biometeorol.
51(2): 155–166. doi:10.1007/s00484-006-0042-y. PMID:
16832653.
Sturm, M., McFadden, J.P., Liston, G.E., Chapin, F.S., III, Racine, C.H., and Holmgren, J. 2001. Snow–shrub interactions in Arctic tundra: a hypothesis with climatic implications. J. Clim. 14(3): 336–344. doi:10.1175/1520-0442(2001)014<0336:SSIIAT>2.0. CO;2.
Sturm, M., Schimel, J., Michaelson, G., Welker, J.M., Oberbauer, S.F., Liston, G.E., Fahnestock, J., and Romanovsky, V.E. 2005. Winter biological processes could help convert arctic tundra to shrubland. Bioscience, 55(1): 17–26. doi:10.1641/0006-3568(2005)055[0017:WBPCHC]2.0.CO;2.
Sveinbjo¨rnsson, B., Hofgaard, A., and Lloyd, A. 2002. Natural causes of the tundra–taiga boundary. Ambio Special Report, 12: 23–29.
Truong, C., Palme´, A.E., and Felber, F. 2007. Recent invasion of the mountain birch Betula pubescens ssp. tortuosa above the treeline due to climate change: genetic and ecological study in northern Sweden. J. Evol. Biol. 20(1): 369–380. doi:10.1111/j. 1420-9101.2006.01190.x. PMID:17210030.
Wang, T., Zhang, Q.B., and Ma, K.P. 2006. Treeline dynamics in relation to climatic variability in the central Tianshan Moun-tains, northwestern China. Glob. Ecol. Biogeogr. Lett. 15(4): 406–415. doi:10.1111/j.1466-822X.2006.00233.x.