TINJAUAN PUSTAKA
Tanaman Padi
Tanaman padi termasuk ke dalam divisio spermatophyta, sub divisio Angiospermae, Kelas Monocotyledoneae, Ordo Graminales, Famili Gramineae, dan Genus Oryza. Terdapat dua spesies budidaya yaitu Oryza sativa dan Oryza
glaberrima. Oryza sativa ditanam oleh banyak negara di dunia tetapi Oryza glaberrima hanya ditanam di sebagian besar Afrika Barat. Oryza sativa kemudian
diklasifikasikan menjadi tiga sub spesies berdasarkan distribusi secara geografi dan karakter morfologi yaitu japonica, indica, dan javanica (Takahashi 1984).
Padi memiliki jumlah kromosom bervariasi diantara 24-48 dengan n=12. Berdasarkan perpasangan kromosom pada fase meiosis, padi mempunyai genom: AA, BB, CC, EE dan FF untuk spesies diploid dan BBCC dan CCDD untuk spesies tetraploid. Spesies padi diploid 2n=24 telah dibudidayakan pada berbagai kondisi lingkungan. O. sativa banyak dibudidayakan di Asia Tenggara dan Asia Selatan dan spesies O. rufipogon dan O. nivara yang merupakan tetua dari O.
glaberrima banyak dibudidayakan di Afrika Barat (Vaughan 1989).
Tipe japonika mempunyai karakter agronomis seperti berumur lebih panjang, struktur tanaman tinggi, mempunyai bulu pada ujung gabah, dan biji berukuran agak besar. Sedangkan tipe indica mempunyai ciri sebaliknya yaitu umur tanaman lebih genjah, biji berukuran lebih kecil dan ramping, tanaman pendek dan tidak berbulu pada bagian palea. Javanica memiliki ciri diantara keduanya (Matsuo et al. 1995).
Pertumbuhan tanaman dibagi menjadi tiga tahapan yaitu tahapan pertumbuhan vegetatif, reproduktif dan pengisian biji atau tahap pematangan. Tahapan vegetatif dimulai dari perkecambahan, kemunculan daun, hingga membentuk anakan. Pemanjangan batang, munculnya daun bendera, fase bunting,
heading dan pembungaan termasuk pada tahapan reproduktif, sedangkan tahapan
pengisian biji merupakan tahapan pematangan yang merupakan akhir dari pertumbuhan padi. Bagian vegetatif terdiri dari tiga bagian yaitu akar, batang dan daun (Counce et al. 2000).
Pertumbuhan padi sangat dipengaruhi oleh faktor lingkungan, terutama suhu. Respon padi terhadap variasi suhu pada fase pertumbuhan (Yoshida 1977) disajikan pada Tabel 1.
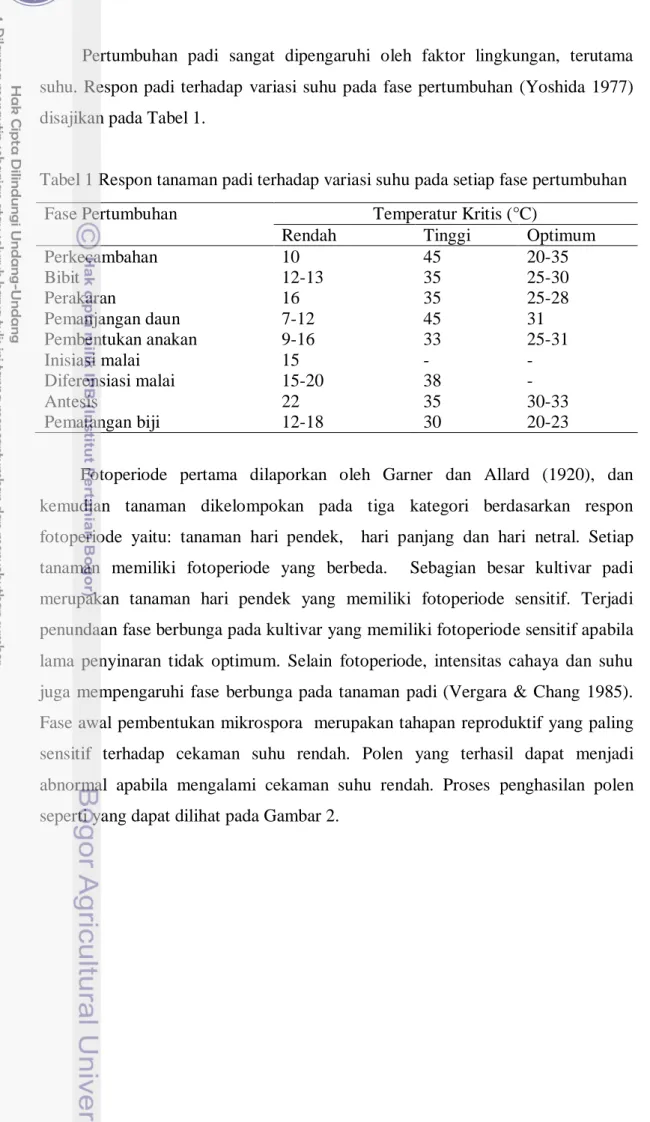
Tabel 1 Respon tanaman padi terhadap variasi suhu pada setiap fase pertumbuhan
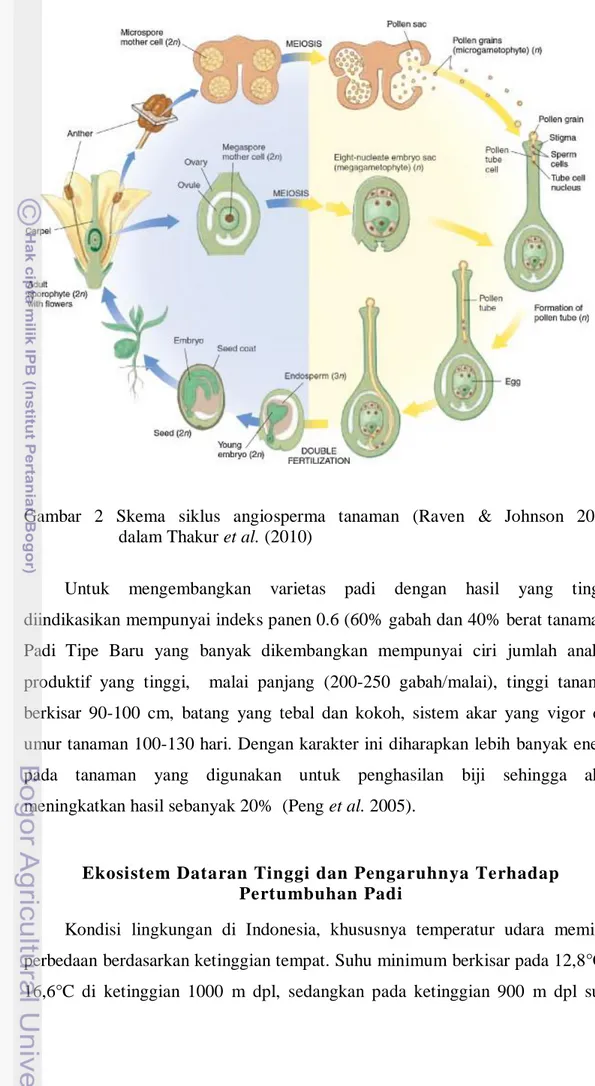
Fotoperiode pertama dilaporkan oleh Garner dan Allard (1920), dan kemudian tanaman dikelompokan pada tiga kategori berdasarkan respon fotoperiode yaitu: tanaman hari pendek, hari panjang dan hari netral. Setiap tanaman memiliki fotoperiode yang berbeda. Sebagian besar kultivar padi merupakan tanaman hari pendek yang memiliki fotoperiode sensitif. Terjadi penundaan fase berbunga pada kultivar yang memiliki fotoperiode sensitif apabila lama penyinaran tidak optimum. Selain fotoperiode, intensitas cahaya dan suhu juga mempengaruhi fase berbunga pada tanaman padi (Vergara & Chang 1985). Fase awal pembentukan mikrospora merupakan tahapan reproduktif yang paling sensitif terhadap cekaman suhu rendah. Polen yang terhasil dapat menjadi abnormal apabila mengalami cekaman suhu rendah. Proses penghasilan polen seperti yang dapat dilihat pada Gambar 2.
Fase Pertumbuhan Temperatur Kritis (°C)
Rendah Tinggi Optimum
Perkecambahan 10 45 20-35 Bibit 12-13 35 25-30 Perakaran 16 35 25-28 Pemanjangan daun 7-12 45 31 Pembentukan anakan 9-16 33 25-31 Inisiasi malai 15 - - Diferensiasi malai 15-20 38 - Antesis 22 35 30-33 Pematangan biji 12-18 30 20-23
Gambar 2 Skema siklus angiosperma tanaman (Raven & Johnson 2002) dalam Thakur et al. (2010)
Untuk mengembangkan varietas padi dengan hasil yang tinggi, diindikasikan mempunyai indeks panen 0.6 (60% gabah dan 40% berat tanaman). Padi Tipe Baru yang banyak dikembangkan mempunyai ciri jumlah anakan produktif yang tinggi, malai panjang (200-250 gabah/malai), tinggi tanaman berkisar 90-100 cm, batang yang tebal dan kokoh, sistem akar yang vigor dan umur tanaman 100-130 hari. Dengan karakter ini diharapkan lebih banyak energi pada tanaman yang digunakan untuk penghasilan biji sehingga akan meningkatkan hasil sebanyak 20% (Peng et al. 2005).
Ekosistem Dataran Tinggi dan Pengaruhnya Terhadap Pertumbuhan Padi
Kondisi lingkungan di Indonesia, khususnya temperatur udara memiliki perbedaan berdasarkan ketinggian tempat. Suhu minimum berkisar pada 12,8°C – 16,6°C di ketinggian 1000 m dpl, sedangkan pada ketinggian 900 m dpl suhu
minimum berkisar antara 14,4°C - 21,0°C (Harahap 1979). Terjadi penurunan suhu sebesar 0,6°C setiap kenaikan ketinggian tempat 100 m (Lockwood 1974). Tanaman padi memiliki suhu kritis berkisar 10°C - 20°C terutama pada waktu antesis dan pada fase mikrosporogenesis (Cruz et al. 2006). Selain dipengaruhi oleh suhu, padi dataran tinggi tumbuh sebagai pertanian yang tergantung hujan, membutuhkan curah hujan lebih dari 750 mm di atas periode 3-4 bulan dan tidak bertoleransi terhadap kekeringan. Di Asia Tenggara kebutuhan rata-rata air untuk irigasi padi adalah 1200 mm per sekali tanam atau 200 mm curah hujan per bulan (Harahap 1979).
Menurut Darmawan dan Baharsjah (2010) pengaruh lingkungan terhadap pertumbuhan yaitu:
a. Curah hujan, besarnya curah hujan mempengaruhi kadar air tanah sehingga sangat berpengaruh terhadap pertumbuhan tanaman.
b. Tinggi dari permukaan laut. Ketinggian suatu tempat mempengaruhi suhu, intensitas cahaya matahari dan curah hujan.
c. Keadaan tanah, yang terdiri dari keadaan fisik tanah yang ditentukan oleh tekstur, struktur tanah dan keadaan kimia tanah yang berkaitan dengan kandungan zat hara di dalam tanah.
d. Suhu, mempengaruhi kecepatan pertumbuhan maupun sifat dan struktur tanaman. Pertumbuhan tanaman padi memiliki suhu minimum 5°C, suhu optimum 25-30°C dan suhu maksimum berkisar antara 35-40°C. Tetapi suhu kardinal (suhu minimum, suhu optimum dan maksimum) sangat dipengaruhi oleh jenis tanaman dan berbeda menurut umur tanaman. Fluktuasi suhu udara siang dan malam hari juga berpengaruh terhadap pertumbuhan.
e. Cahaya matahari yang terdiri dari intensitas cahaya, kualitas cahaya (panjang gelombang) dan lamanya penyinaran (panjang hari) juga sangat berpengaruh terhadap pertumbuhan tanaman. Pengaruh dari ketiga sifat cahaya tersebut terhadap pertumbuhan tanaman adalah melalui pembentukan klorofil, pembentukan stomata, pembentukan antocyanin (pigmen merah), perubahan suhu daun atau batang, penyerapan hara, permeabilitas dinding sel, transpirasi dan gerakan protoplasma.
Faktor yang menyebabkan bahaya cekaman suhu rendah terhadap tanaman padi berasal dari temperatur udara yang rendah dan suhu air irigasi yang dingin. Kedua komponen ini dapat digunakan dalam skrining tanaman toleran suhu rendah. Suhu tertentu akan menyebabkan kerusakan bergantung pada genotipe dan fase pertumbuhan padi (Kaneda & Beachell 1974).
Mekanisme yang tejadi pada tanaman selama cekaman suhu rendah yaitu terjadinya low temperature photoinhhibition, dimana terhambatnya pemanfaatan cahaya oleh tanaman pada proses fotosintesis. Fotoinhibisi berkaitan langsung dengan kompleks protein fotosistem II (PS II). Untuk mengurangi fenomena ini, maka diperlukan asam lemak tak jenuh phosphatidylglycerol yang terdapat dalam membran tilakoid yang berkaitan dengan kompleks fotosistem II (Taiz & Zeinger 2002).
Fotosintesis dalam peranannya pada proses diferensiasi dan perkembangan tanaman yaitu memperbesar kapasitas sink dengan menyediakan bahan dan energi yang diperlukan untuk peningkatan hasil. Setelah pembentukan malai, fotosintesis akan berperanan dalam proses pengisian gabah dengan cara menyediakan karbohidrat (Murata & Nishida 1989). Tekanan yang terjadi pada fotosistem II pada proses fotosintesis, dapat mempengaruhi morfologi tanaman dan ekspresi gen yang terlibat selama aklimasi suhu rendah (Gray et al. 1997).
Fase berbunga diatur oleh gen dan faktor lingkungan. Cahaya (fotoperiode dan kualitas cahaya) dan suhu merupakan faktor pengatur utama pada fase pembungaan tanaman. Fotoperiode (panjang hari) merupakan signal lingkungan yang paling penting untuk transisi pembungaan. Tanaman dapat mengenali dan mengukur perubahan panjang hari secara akurat untuk mengatur waktu berbunga. Bahaya cekaman suhu rendah berbeda untuk setiap kultivar, pada suhu 15°C selama 4 hari pada fase awal pembentukan mikrospora meningkatkan sterilitas gabah pada kultivar yang toleran cekaman suhu rendah, sedangkan untuk kultivar yang rentan terhadap cekaman suhu rendah mempunyai suhu kritis pada 17-19 °C. Suhu rendah (12°C) selama dua hari tidak menyebabkan sterilitas, tetapi jika lebih dari 6 hari menyebabkan sterilitas sebesar 100% (Satake 1969).
Waktu heading tanaman padi pada suhu rendah (23°C) berlangsung lebih lambat dibandingkan dengan suhu normal (27°C). Penelitian sebelumnya menunjukkan bahwa gen Hd3a dan Hd1 memiliki peran penting selama fase pembungaan. Ekspresi gen Hd3a berkurang pada suhu rendah selama fase berbunga tetapi ekspresi gen Hd1 tidak dipengaruhi oleh perlakuan suhu yang berbeda (23°C dan 27°C). Hal ini mengindikasikan bahwa penekanan ekspresi gen Hd3a oleh suhu rendah menyebabkan terlambatnya waktu pembungaan (Luan
et al. 2009).
Waktu berbunga adalah tahapan yang paling sensitif terhadap cekaman suhu rendah seperti yang telah dilaporkan oleh Enomoto (1933) dan Sakai (1937). Terao et al. (1940) dalam percobaanya melaporkan bahwa tingkat sterilitas maksimum terjadi pada cekaman suhu rendah pada fase meiosis yang terjadi pada polen dan fase awal pembentukan malai. Penelitian lain menyatakan bahwa tahapan yang paling sensitif terhadap cekaman suhu rendah yaitu pada fase bunting, sekitar 11 hari sebelum heading (Kakizaki 1938). Tanaka (1962) melaporkan bahwa pembungaan akan terhambat apabila suhu maksimum di bawah 25°C dan yang paling mempengaruhi yaitu suhu minimum per hari.
Jumlah gabah hampa berkorelasi nyata dengan cekaman suhu rendah. Jumlah polen per anter yang terhasil, lebih sedikit dan ukuran stigma lebih kecil pada cekaman suhu rendah sehingga meningkatkan jumlah gabah hampa (Farrell
et al. 2006). Telah dibuktikan bahwa spikelet dan malai merupakan organ yang
paling sensitif terhadap cekaman suhu rendah selama fase bunting, terutama anter (Nishiyama et al. 1969). Posisi spikelet pada malai juga turut menentukan tingkat sterilitas pada kondisi suhu rendah, spikelet yang berada pada ujung malai lebih banyak yang steril dibandingkan dengan di bagian bawah (Nishimura 1987).
Beberapa gen pada padi seperti Ehd1, Ghd7 dan RID1/ Ehd2/OsId1 yang berperan penting dalam fase berbunga telah dapat diidentifikasi, namun mekanisme molekular respon tanaman terhadap suhu pada fase berbunga masih belum diketahui secara jelas (Li & Jiang 2012). Ketahanan tehadap cekaman suhu rendah dikendalikan oleh 5-7 gen dominan yang bersifat aditif (Toriyama 1962).
Cekaman suhu rendah selama tahap reproduksi pada padi dapat mengurangi jumlah gabah, eksersi malai yang tidak sempurna dan meningkatnya sterilitas
gabah sehingga berakibat pada berkurangnya hasil (Han et al. 2006). Hal yang serupa disampaikan oleh Lee (2001) yang menyatakan bahwa faktor lingkungan utama yang menyebabkan panjangnya umur tanaman dan persentase gabah hampa yang tinggi yaitu besarnya cekaman suhu rendah pada kawasan dataran tinggi yang berakibat pada sterilitas polen, fase vegetatif menjadi lebih panjang dan terhambatnya proses pengisian biji pada tanaman padi.
Kultivar yang memiliki ketahanan terhadap cekaman suhu rendah dapat diidentifikasi pada suhu kritis tanaman padi yaitu 19°C. Suhu di bawah 15°C selama fase vegetatif berdampak terhadap klorosis daun, jumlah anakan sedikit dan penundaan waktu berbunga (Jena & Jeung 2004). Bobot 1000 butir dan ukuran gabah lebih kecil pada kondisi cekaman suhu rendah, sedangkan kandungan protein lebih tinggi. Pengaruh cekaman suhu rendah bervariasi untuk setiap genotipe (Zhao et al. 2009).
Angka pertumbuhan yang sama diperoleh dengan membandingkan kultivar yang tahan dan rentan cekaman suhu rendah pada tingkat suhu dan durasi perlakuan yang berbeda. Kultivar yang toleran suhu rendah masih dapat tumbuh setelah perlakuan selama 5 hari pada suhu 12-14°C, sedangkan pada kultivar yang rentan suhu rendah angka pertumbuhan yang sama didapatkan setelah diperlakukan dengan suhu 17-20°C selama 4 hari (Nishiyama et al. 1969). Suhu malam di bawah 5°C tidak menyebabkan kerusakan yang berarti. Oleh karena itu, cekaman suhu rendah selama beberapa jam tidak menyebabkan sterilitas. Selain tingkat cekaman suhu rendah, durasi berlangsungnya cekaman suhu rendah juga merupakan faktor yang penting (Kashibuchi 1968).
Dua akibat utama yang disebabkan cekaman suhu rendah yaitu sterilitas dan terlambatnya pertumbuhan. Sterilitas merupakan gagalnya penyerbukan yang disebabkan adanya cekaman suhu rendah selama fase bunting dan pembungaan. Sedangkan jenis kerusakan terlambatnya pertumbuhan ditandai dengan matang yang tidak sempurna. Suhu kritis yang menyebabkan sterilitas bervariasi berdasarkan durasi cekaman suhu rendah, perbedaan suhu siang dan malam, kondisi lingkungan sebelum dan sesudah fase kritis, metode dan jumlah pupuk yang digunakan dan jenis kultivar yang digunakan (Matsuo et al. 1995).
Bentuk kerusakan pada tanaman padi akibat cekaman suhu rendah berbeda pada suatu daerah dengan daerah lainnya. Seperti di Korea, cekaman suhu rendah berpengaruh terhadap fase bibit dan pematangan, di Nepal dan India suhu rendah berpengaruh terhadap jumlah anakan dan fase berbunga. Di negara tropis dimana penanaman dilakukan secara terasering, perbedaan suhu bergantung pada ketinggian tempat. Oleh karena itu perlu dikembangakan varietas padi dataran tinggi yang spesifik lokasi karena beragamnya kerusakan yang ditimbulkan cekaman suhu rendah pada berbagai fase pertumbuhan yang berdampak terhadap penurunan hasil (Nanda & Seshu 1979).
Di Indonesia, penanaman beberapa varietas padi pada ketinggian 700-1000 m dpl, memiliki hasil yang rendah dengan sterilitas tinggi yang disebabkan oleh cekaman suhu rendah selama fase bunting dan berbunga (Harahap 1979). Perbedaan tingkat cekaman suhu rendah akan mempengaruhi hasil dan komponen hasil padi yang ditanam di berbagai kondisi lingkungan, khususnya dengan suhu rendah yang bervariasi (Jiang et al. 2010).
Selain cekaman suhu rendah, lama penyinaran juga mempengaruhi sterilitas tanaman. Diperlukan intensitas cahaya yang lebih banyak selama proses fotosintesis untuk menghasilkan tanaman yang baik. Pengaruh cekaman suhu rendah pada proses fotosintesis sangat kecil pada kondisi kurang cahaya, disebabkan karena tingkat reaksi ditentukan oleh fotokimia. Fotosintesis dapat berlangsung optimal pada kisaran suhu 15°C – 30°C. Aktivitas RuBP karboksilase yang terlibat dalam fiksasi CO2 selama proses fotosintesis dapat bereaksi pada
kisaran suhu 10 – 40 °C, aktivitas meningkat seiring dengan peningkatan suhu (Ishii et al. 1977). Laju fotosintesis berkurang drastis pada suhu di bawah 18°C dan di atas 33°C, tetapi tidak terdapat perbedaan pada kisaran suhu 18 - 33°C, namun demikian suhu optimum fotosintesis relatif berbeda untuk setiap genotipe (Yamada et al. 1955).
Laju fotosintesis dipengaruhi oleh beberapa faktor diantaranya; (a) kadar CO2 (b) suhu, apabila intensitas cahaya cukup tinggi, maka suhu semakin tinggi
dan laju fotosintesis semakin meningkat (c) cahaya, terdiri dari tiga komponen yaitu intensitas cahaya, kualitas cahaya dan lama penyinaran (d) air tanah (e) kadar O2 (f) kandungan hara dalam tanaman dan (g) kandungan klorofil.
Kaitan antara fotosintesis dan respirasi yang berpengaruh terhadap komponen hasil seperti jumlah anakan, jumlah malai, jumlah gabah per malai, persentase gabah isi dan bobot gabah telah dipelajari oleh Matsushima 1957. Fotosintesis berfungsi menyediakan bahan dasar yang berkontribusi terhadap setiap komponen hasil, di sisi lain respirasi menyediakan energi yang diperlukan selama tahapan diferensiasi dan pertumbuhan organ pada komponen hasil. Peranan fotosintesis selama periode pembentukan sink dan akumulasi karbohidrat sebagai sumber sink lebih besar dibandingkan peranannya pada periode pertumbuhan. Kurangnya cahaya berdampak terhadap berkurangnya jumlah bunga yang terhasil dan persentase gabah isi (Yoshida 1981).
Pengaruh cekaman suhu rendah terhadap tanaman lain juga telah dilaporkan pada sorgum (Yu et al. 2004), tembakau (Kodama et al. 1994) dan strawberi (Rajashekar et al. 1999).
Keragaan Karakter Agronomi Padi Dataran Tinggi
Beberapa karakter yang perlu diperbaiki untuk meningkatkan produksi padi yaitu jumlah malai, kapasitas sink, jumlah gabah total, panjang malai, efisiensi pengisian gabah yang stabil dan potensi hasil yang tinggi (Chen 2008). Tingginya persentase gabah bernas dan jumlah gabah total berkorelasi positif terhadap produksi. Karakter utama yang menentukan produksi di lingkungan dengan cekaman suhu rendah adalah umur tanaman dan persentase gabah bernas. Oleh karena itu perlu ditingkatkan keragaan karakter agronomi diantaranya memiliki umur lebih genjah dan persentase gabah bernas, jumlah malai dan panjang malai yang lebih baik (Rasyad et al. 2012).
Daun merupakan organ utama yang berfungsi dalam menghasilkan dan mengangkut asimilat, oleh karena itu permukaan daun yang lebih luas akan menghasilkan gabah yang lebih banyak, disamping itu posisi daun bendera juga memainkan peranan penting terutama tiga daun pertama dengan sudut yang kecil akan berpengaruh terhadap fotosintesis (Chen et al. 2002).
Kemampuan membentuk anakan produktif dipengaruhi oleh interaksi antara genotipe dan lingkungan tumbuhnya. Pada ekosistem dataran tinggi, pembentukan anakan padi terhambat diakibatkan oleh suhu dan intensitas cahaya matahari yang
rendah (Endrizal 2012). Fageria (2007) melaporkan bahwa kultivar dengan jumlah anakan yang banyak akan lebih baik dibandingkan dengan yang memiliki anakan sedikit pada lingkungan bercekaman, karena dapat mengimbangi produksi, namun pada lingkungan optimal tidak memiliki pengaruh nyata. Kemampuan tanaman menghasilkan anakan mempunyai pengaruh besar terhadap pembentukan malai yang berkorelasi kuat dengan hasil (Miller et al. 1991). Jumlah anakan sangat dipengaruhi oleh kondisi lingkungan seperti cahaya, suhu, densitas tanaman dan nutrisi (Wu et al. 1998).
Kontribusi karakter agronomi lainnya yaitu persentase gabah bernas yang memiliki kontribusi di lingkungan optimal sebesar 4%, sedangkan di lingkungan marginal jauh lebih besar yaitu sebesar 69%, dapat diartikan bahwa karakter persentase gabah bernas merupakan karakter utama di dataran tinggi dengan cekaman suhu rendah. Struktur tanaman yang pendek akan meningkatkan indeks panen dan meningkatkan biomasa hasil (Khush 1999). Kepadatan malai, jumlah gabah per malai, bobot gabah dan jumlah gabah bernas merupakan komponen hasil utama yang berkontribusi terhadap hasil (Fageria 2007).
Pemuliaan Tanaman Padi
Berbagai metode pemuliaan tanaman telah digunakan untuk meningkatkan keragaman genetik tanaman di antaranya dengan teknik persilangan buatan, pemuliaan mutasi dan bioteknologi (Poespodarsono 1988). Program pemuliaan berupaya melakukan perbaikan terhadap keragaan dan produktivitas tanaman padi. Penentuan ideotipe tanaman dalam pemuliaan sangat diperlukan untuk meningkatkan potensi genetik karakter yang diinginkan dengan memodifikasi karakter tersebut secara spesifik (Roy 2000).
Hibridisasi (persilangan) pada tanaman padi dapat menyebabkan terjadinya kombinasi alela-alela yang dapat meningkatkan keragaman genetik. Penentuan tetua merupakan tahap yang sangat penting karena akan menentukan keberhasilan dari tujuan perolehan karakter yang diinginkan. Tetua yang digunakan harus memiliki karakter yang diinginkan dan mempunyai adaptasi yang baik. Keragaman yang tinggi dapat dihasilkan dengan menggunakan tetua yang mempunyai kekerabatan yang jauh (Allard 1960).
Pemuliaan mutasi terhadap tanaman padi telah dimulai pada tahun 1980. Di beberapa negara teknik ini banyak digunakan untuk menghasilkan tanaman dengan hasil yang lebih baik untuk berbagai karakter dan ketahanan terhadap hama dan penyakit tanaman. Di Cina sebanyak 145 varietas telah dihasilkan sejak 1966 dengan menggunakan sinar gamma dan perbaikan dilakukan terhadap karakter agronomi dan fenotipik kualitas gabah (Carena. 2009).
Sebanyak 2541 varietas telah dilepas, sebagian besar dihasilkan dari hasil mutasi induksi. Jenis tanaman yang paling banyak dihasilkan melalui mutasi induksi yaitu sereal (1212 varietas) diikuti oleh kekacangan dan tanaman industri. Padi merupakan tanaman jenis sereal yang terbanyak dihasilkan dari teknik mutasi yaitu sebanyak 525 varietas diikui oleh barley (303 varietas) dan gandum (200 varietas) (Guimaraes 2010). Karakter utama pada padi yang mengalami banyak perubahan yaitu umur panen, tinggi tanaman dan ketahanan terhadap penyakit (Maluszynski et al. 1998).
Korelasi Genetik Karakter Agronomi
Koefisien korelasi genetik dapat bernilai positif atau negatif. Korelasi antara dua sifat mengindikasikan bahwa perubahan pada suatu sifat juga turut memberikan andil bagi perubahan lainnya. Terjadi peningkatan keragaan secara bersama-sama bila koefisien bernilai positif dan berbanding terbalik bila koefisien korelasi negatif. Perubahan pada suatu sifat tidak memberikan andil bagi perubahan sifat lainnya jika tidak terdapat korelasi (Steel et al. 1977).
Limbongan (2008) melaporkan hasil penelitian tanaman padi pada ketinggian 750 m dpl dan 1500 m dpl. Kedua lokasi memiliki bobot gabah relatif lebih tinggi pada tanaman yang berumur genjah dan hal sebaliknya terjadi pada tanaman yang berumur lebih panjang. Tingginya persentase gabah isi dan jumlah gabah total per malai berkorelasi positif terhadap produksi (Rasyad et al. 2012).
Schnier et al. (1990) melaporkan karakter tinggi tanaman dan umur panen berkorelasi negatif dan nyata. Sedangkan karakter panjang malai, persentase gabah isi, dan bobot 1000 butir berkorelasi positif dan nyata terhadap hasil. Hasil berkorelasi kuat dengan persentase gabah isi, tetapi berkorelasi lemah dengan jumlah gabah per malai. Jumlah anakan yang kurang merupakan faktor pembatas
bagi pembentukan sink. Jumlah malai per meter per segi merupakan komponen hasil terpenting yang berpengaruh sebesar 89% dari variasi hasil (Jones & Synder 1987).
Terdapat pengaruh langsung yang positif di antara karakter jumlah gabah bernas per malai terhadap hasil, namun merupakan pengaruh terpenting kedua setelah pengaruh langsung kepadatan malai. Pengaruh langsung jumlah gabah hampa per malai terhadap hasil bersifat negatif, tetapi bila berasosiasi dengan persentase gabah isi, maka pengaruhnya lebih besar (Gravois & Helms 1992).
Interaksi Genotipe x Lingkungan
Adanya interaksi genotipe x lingkungan (G x E) menunjukkan kegagalan genotipe yang diuji memperlihatkan keragaan yang relatif sama dari satu lingkungan ke lingkungan lainnya (Fehr 1987). Informasi menenai GxE sangat berguna dalam menentukan apakah dapat dilakukan pengembangan suatu kultivar di semua lingkungan yang diinginkan atau perlu dilakukan pengembangan kultivar spesifik untuk lingkungan target yang spesifik (Bridges 1989).
Urutan relatif suatu varietas akan berubah dari tempat ke tempat dan dari musim ke musim dengan adanya interaksi G x E. Suatu genotipe akan dapat tumbuh dan berproduksi dengan sama baiknya di berbagai tempat atau lingkungan pertumbuhannya jika tidak terdapat interaksi G x E sehingga varietas atau galur dapat dikatakan stabil. Varietas yang stabil sangat penting untuk mengurangi resiko akibat perubahan lingkungan yang sukar diramalkan seperti kesuburan tanah, perubahan cuaca yang menyolok serta serangan hama dan penyakit (Limbongan 2008).
Pentingnya interaksi GxE dalam analisis stabilitas telah banyak dilaporkan pada sereal (Saeed & Francis. 1984; Oosterom et al. 1993; Harsanti et al. 2003) dan barley (Ceccarelli 1987) yang menyatakan bahwa suhu rendah sangat mempengaruhi hasil dan komponen hasil pada tanaman barley.
Menurut Nasrullah (1981), bahwa interaksi G x E dapat dipergunakan untuk mengukur stabilitas suatu genotipe, karena stabilitas penampilan pada suatu kisaran lingkungan tergantung dari besarnya interaksi tersebut. Pada uji daya hasil galur-galur seringkali terjadi interaksi antara galur dengan lingkungan. Perbedaan ini dapat
mengakibatkan perubahan daya hasil antara suatu tempat dengan tempat lainnya. Mengingat perbedaan hasil sangat dipengaruhi oleh perbedaan genetik dan lingkungan, maka perlu memilih galur-galur yang unggul dengan hasil yang stabil (Sutjihno 1993).
Adanya variasi lingkungan tumbuh makro tidak akan menjamin suatu genotipe atau varietas tanaman akan tumbuh baik dan memberikan hasil panen tinggi di semua wilayah dalam kisaran area yang luas, atau sebaliknya. Hal tersebut terkait dengan kemungkinan ada atau tidak adanya interaksi antara genotipe atau genotipe-genotipe tanaman dengan kisaran variasi lingkungan yang luas (Baihaki & Wicaksono 2005). Informasi menenai GxE sangat berguna dalam menentukan apakah dapat dilakukan pengembangan suatu kultivar di semua lingkungan yang diinginkan atau perlu dilakukan pengembangan kultivar spesifik untuk lingkungan target yang spesifik (Bridges 1989). Cara yang dapat dilakukan untuk mengetahui daya adaptasi suatu genotipe yaitu dengan pengujian galur harapan pada beberapa lingkungan. Hasil analisis variansnya akan menunjukkan besarnya interaksi GxE, sehingga lebih mudah menentukan galur yang ideal (Eberhart & Russel 1966).
Stabilitas dan Adaptabilitas
Interaksi G x E dapat digunakan untuk mengukur stabilitas suatu genotipe (Nasrullah 1981), karena stabilitas penampilan pada suatu kisaran lingkungan tergantung dari besarnya interaksi G x E. Stabilitas adalah kemampuan tanaman untuk mempertahankan daya hasil terhadap perubahan kondisi lingkungan. Stabilitas hasil merupakan karakter yang diwariskan melalui daya saing populasi yang secara genetik heterogen (Nor & Cady 1979). Salah satu metode yang dapat digunakan dalam menduga adaptabilitas dan stabilitas fenotipik seperti hasil adalah dengan cara melakukan pengujian berulang pada berbagai lingkungan tumbuh yang bervariasi (Singh & Chaudhary 1979).
Lin et al. (1986) membagi konsep stabilitas menjadi tiga tipe. Tipe 1 yaitu suatu genotipe dianggap stabil bila ragam lingkungannya kecil. Stabilitas tipe ini digunakan oleh Francis dan Kannenberg (1978). Tipe 2 yaitu suatu genotipe dikatakan stabil jika respon lingkungan paralel terhadap rata-rata respon semua genotipe percobaan. Tipe 3 yaitu suatu genotipe dikatakan stabil bila kuadrat tengah sisa model regresi pada indeks lingkungan kecil.
Pemilihan genotipe untuk lingkungan spesifik didasarkan pada nilai duga interaksi G x E yang nyata, menggambarkan kemampuan suatu genotipe mengekspresikan gen-gen yang menguntungkan pada lingkungan tertentu sehingga diperoleh hasil tinggi. Sebaliknya, pemilihan genotipe yang beradaptasi pada lingkungan luas didasarkan pada nilai duga interaksi G x E yang tidak nyata, yang menggambarkan kemampuan suatu genotipe berpenampilan sama pada kondisi lingkungan berbeda (Baihaki & Wicaksono 2005). Cara yang paling umum dilakukan untuk mengenali galur ideal adalah dengan menguji seperangkat galur harapan pada beberapa lingkungan. Berdasarkan hasil analisis variansnya, akan diketahui interaksi G x E (Eberhart & Russel 1966).
Finlay dan Wilkinson (1963) menggunakan koefisien regresi sebagai ukuran stabilitas sedangkan Eberhart dan Russel (1966) menggunakan rata-rata jumlah kuadrat simpangan regresi. Shukla (1972) menggunakan besaran yang disebut varians stabilitas untuk menyatakan genotipe yang stabil. Gauch (1992) menggunakan model AMMI untuk menyatakan genotipe stabil berdasarkan gabungan antara analisis ragam dan analisis komponen utama.
Adaptabilitas adalah kemampuan tanaman untuk menyesuaikan diri terhadap kondisi lingkungan pertumbuhannya. Tanggapan genotipe terhadap lingkungannya dikelompokkan menjadi dua yaitu kelompok pertama, yang menunjukkan kemampuan adaptasi pada lingkungan yang luas berarti interaksi G x E kecil. Kelompok kedua adalah yang menunjukkan kemampuan adaptasi sempit atau beradaptasi khusus dan berperagaan baik pada suatu lingkungan tetapi berperagaan buruk pada lingkungan yang berbeda, dapat dikatakan memiliki interaksi G x E besar (Soemartono & Nasrullah 1988).
Analisis Stabilitas Finlay dan Wilkinson (bi)
Parameter stabilitas Finlay dan Wilkinson diduga dengan menggunakan nilai koefisien regresi tiap genotipe (bi) yang dikelompokkan menjadi tiga :
1. Jika bi ≈ 1, memiliki stabilitas rata-rata.
2. Jika bi > 1, stabilitas berada di bawah rata-rata. Genotipe peka terhadap perubahan lingkungan dan beradaptasi khusus pada lingkungan yang menguntungkan.
3. Jika bi < 1, stabilitas berada di atas rata-rata. Genotipe beradaptasi pada lingkungan marginal.
Analisis Stabilitas menurut Eberhart dan Russel (1966)
Eberhart dan Russel (1966) menggunakan parameter koefisien regresi (bi) dan simpangan regresi (Sdi2). Suatu genotipe dikatakan stabil jika kuadrat tengah sisa dari model regresi pada indeks lingkungannya kecil. Indeks lingkungan adalah rata-rata hasil dari semua genotipe pada masing-masing lokasi dikurangi rataan total dari semua genotipe di semua lokasi.
Analisis Stabilitas menurut Francis dan Kannenberg (1978)
Francis dan Kannenberg (1978) menggunakan parameter koefisien keragaman (CVi) untuk masing-masing galur sebagai parameter stabilitas dan
keragaman genotipe terhadap lingkungan. Kriteria nilai koefisien keragaman menurut Moedjiono dan Mejaya (1994) yaitu rendah (0 < x < 25%), agak rendah (25% < x < 50%), cukup tinggi (50% < x < 75%), dan tinggi (75% < x < 100%). Nilai CVi yang semakin kecil menunjukan bahwa genotipe tersebut lebih stabil pada lingkungan yang diuji.
Analisis Stabilitas AMMI
Model AMMI (Additive Main Effects and Multiplicative Interaction) menggabungkan analisis ragam aditif bagi pengaruh utama perlakuan dengan analisis komponen utama ganda dengan pemodelan bilinier bagi pengaruh interaksi. Model AMMI dapat digunakan untuk menganalisis percobaan lokasi ganda. Asumsi yang mendasari pengujian ini adalah perlakuan dan lingkungan bersifat aditif, ragam yang homogen dan galat bebas (Mattjik dan Sumertajaya 2008).
Gauch (1992) menggunakan model AMMI dengan menyatakan genotipe yang stabil berdasarkan gabungan antara analisis ragam dan analisis komponen utama. Genotipe yang stabil dapat digambarkan dengan model biplot (Yang 2000). Model AMMI secara lengkap:
Ygen = μ+αg+βe+∑√λnφgnρen+δge+εgen Keterangan: g=1,2,…,a; e=1,2,…,b; n=1,2,…,m
Parameter √λn adalah nilai singular untuk komponen bilinier ke-n. Pengaruh ganda genotipe ke-g melalui komponen bilinier ke-n dilambangkan dengan φgn, dan ρen merupakan pengaruh ganda lokasi ke-e melalui komponen bilinier ke-n. Asumsi-asumsi yang mendasari analisis ragam adalah galat percobaan menyebar saling bebas mengikuti sebaran normal dengan ragam homogen (εijk ~ N (0,σ2ε) (Mattjik dan Sumertajaya 2008).