Brain Research 885 (2000) 133–136
www.elsevier.com / locate / bres
Short communication
Autoradiographic analysis of dopamine receptor-stimulated
35
[ S]GTP
g
S binding in rat striatum
*
Li He, Donato A. Di Monte, J. William Langston, Maryka Quik
The Parkinson’s Institute, 1170 Morse Avenue, Sunnyvale, CA 94089, USA
Accepted 29 August 2000
Abstract
35
Autoradiographic analysis of [ S]GTPgS binding was used to investigate functional activation of dopamine receptors in rat striatum.
35
Dopamine-stimulated [ S]GTPgS binding was observed with a maximal increase of 38% over basal activity. A similar stimulatory response was obtained with the D agonist quinpirole, but not SKF-238393, a D agonist. The effect of dopamine was blocked by the D2 1 2
antagonist raclopride, but was unaffected by SCH-23990, a D1 antagonist. There appeared to be a differential distribution of
35
dopamine-stimulated [ S]GTPgS binding, with the lowest activity obtained in the medial portion of the caudal striatum. These results
35
demonstrate, using an autoradiographic approach, (i) that dopamine stimulated [ S]GTPgS binding in the rat striatum occurs through activation of D receptors, and (ii) that the effects of dopamine activation vary in different areas of the rat striatum.2 2000 Elsevier Science B.V. All rights reserved.
Theme: Neurotransmitters, modulators, transporters, and receptors
Topic: Catecholamine receptors
35
Keywords: Autoradiography; Dopamine receptor; Rat; [ S]GTPgS binding; Striatum
The central and peripheral effects of dopamine are adrenergic receptors [20], cannabinoid receptors [14], mediated via dopamine receptors, of which there are at adenosine receptors [8] and opioid receptors [18,21], as least five different subtypes. They belong to the superfami- well as dopamine receptors [2–5,7,13]. This assay relies ly of neurotransmitter receptors, which couple to their on agonist-promoted GDP/ GTP exchange occurring at the specific effector functions via G-proteins [9,15]. Mem- level of the G-protein within the receptor / G-protein
com-35
brane-associated G-proteins have been long recognized as plex with [ S]GTPgS binding used as an indicator of a key component in the dopamine receptor signal transduc- receptor activation. In addition to being sensitive and tion pathway and control a variety of intracellular effec- reproducible [1], this technique can also be used in tors, including the activity of adenylyl cyclase, phospho- autoradiographic analysis [16], making it possible to diesterase, and some ionic channel conductance. Therefore, identify and quantitate the functional activity of G-protein-information concerning the functional status of G-proteins coupled receptors in specific brain regions [17]. The is essential for a better understanding of dopamine re- present study was designed to evaluate and compare the ceptor-mediated functions. efficiency of dopamine and dopamine receptor agonists in Receptor activation of G-proteins can be monitored by activating G proteins coupled to striatal dopamine
re-35
several methods [11,12]. This includes agonist-stimulated ceptors using an autoradiographic analysis of [ S]GTPgS
35 35
guanosine-59-O-(3-[ S]thio)triphosphate ([ S]GTPgS) binding. The anatomical distribution of
dopamine-stimu-35
binding, which is widely used for assaying the coupling of lated [ S]GTPgS binding sites was also determined various neurotransmitter receptors to G-proteins, such as throughout the rat striatum.
Male Sprague–Dawley rats, weighing 200–250 g, were purchased from Simonsen Laboratories (Gilroy, CA). All
*Corresponding author. Tel.: 11-408-542-5601; fax: 1
1-408-734-experiments were carried out in accordance with the
8522.
E-mail address: [email protected] (M. Quik). National Institutes of Health Guide for the Care and Use of
134 L. He et al. / Brain Research 885 (2000) 133 –136
Laboratory Animals and were approved by the Institutional Animal Care and Use Committee at the Parkinson’s Institute. Rats were killed by CO2 inhalation and brains were quickly removed and immersed in isopentane on dry ice. Coronal sections (18 mm) were cut through the striatum on a cryostat at (2168C) and thaw-mounted onto poly-L-lysine-subbed slides. Sections were stored at2808C
for 3 to 7 days.
Sections were preincubated in assay buffer (50 mM Tris–HCl, 3 mM MgCl , 0.2 mM EGTA and 100 mM2
NaCl, pH 7.4) at 258C for 15 min followed by a second
Fig. 1. Representative autoradiogram depicting dopamine-stimulated
preincubation for 15 min in the same buffer, containing 2 35
[ S]GTPgS binding in rat striatum. Coronal sections were incubated in
mM GDP [16]. Sections were incubated for 1 h at 258C in 35 buffer (see Methods) also containing 2 mM GDP and 50 pM [ S]GTPgS 35
the above buffer with 50 pM [ S]GTPgS (1255 Ci / mmol, in the absence (Basal) or presence of 100 mM dopamine. Note the
35
New England Nuclear, Boston, MA) in the presence or significant increase in dopamine-stimulated [ S]GTPgS binding in striatum.
absence of dopamine and other dopamine receptor ligands. The sections were then washed twice for 3 min each in ice-cold 50 mM Tris–HCl (pH 7.4) followed by a brief rinse in ice-cold distilled water. After drying, the sections
raclopride was much more effective than the D antagonist were exposed to MP film for 3 to 4 days. 1
35
Autoradiographic images were analyzed using Im- SCH-23390 in blocking dopamine-stimulated [ S]GTPgS ageQuant software (Molecular Dynamics, Sunnyvale, CA). binding. Similar results were obtained using rat striatal Agonist-stimulated activity in rat brain sections was calcu- synaptic membranes using the GTPase assay [12]. The lated by subtracting the optical density in basal sections relative functional unresponsiveness of D1 receptors to (incubated with GDP in the absence of agonists) from that dopamine stimulation is somewhat surprising given the of agonist-stimulated sections. Data were expressed as % fact that these receptors are quite abundant in the rat stimulation over basal, where striatum [19]. However, this apparent discrepancy supports the notion that the receptor binding properties of neuronal stimulated2basal
membranes do not necessarily correlate with functional
]]]]]
%stimulation5 3100
basal coupling to intracellular signal transduction mechanisms Statistical significance was determined using Student’s
t-test, with P,0.05 considered significant. Competition curves were generated using the program Prism (Grap-hPAD, San Diego, CA).
The first goal of this study was to characterize the receptor subtypes which mediate dopamine stimulated
35
[ S]GTPgS binding in the rat striatum using an
au-35
toradiographic approach. Dopamine increased [ S]GTPgS binding (Fig. 1) in a dose-dependent manner with a maximum stimulatory effect of 38% above basal level (Fig. 2). A similar, albeit somewhat lower (31%) dose-dependent stimulation was observed with quinpirole (Fig. 2), an agonist selective for D2 receptors. SKF-38393, a highly selective D1 receptor agonist, resulted in only a
35
nonsignificant increase (15%) in [ S]GTPgS binding. We next tested the effect of selective dopamine receptor antagonists (Fig. 3). The D receptor antagonist raclopride2
35
significantly blocked dopamine-stimulated [ S]GTPgS binding with a maximum inhibitory effect of 87%. In contrast, no statistically significant inhibition of
dopamine-35
stimulated [ S]GTPgS binding was obtained with the D1
Fig. 2. Dose–response curves for dopamine-, the selective D receptor
receptor antagonist SCH-23390. These results indicate that 1 35 agonist SKF-28389- and the selective D receptor agonist
quinpirole-2
the dopamine-stimulated increase in [ S]GTPgS binding 35
stimulated [ S]GTPgS binding to rat striatal sections. The maximum
occurs primarily through D receptor activation, in agree-2 stimulation produced by dopamine was 38%, and quinpirole 31%, with no
ment with Rinken and colleagues [13]. Using a membrane significant stimulation by SKF-38393. Each value represents the
L. He et al. / Brain Research 885 (2000) 133 –136 135
35
stimulated [ S]GTPgS binding may reflect the sub-re-gional distribution of D2 receptors within the striatum since a similar lateral-to-medial gradient for this receptor has been observed using receptor binding [6].
The present results are, to our knowledge, the first to characterize dopamine receptor-stimulated GTPgS binding using autoradiographic analysis. Our results clearly
dem-35
onstrate the ability of dopamine to increase [ S]GTPgS binding above basal levels, thus confirming previous data obtained using striatal membrane preparations [5,13] and sections [7]. The dose-dependent and saturable nature of dopamine action shows that dopamine-stimulated
35
[ S]GTPgS binding is to a dopamine receptor, most likely of the D2 subtype. Furthermore, this study demonstrates the feasibility of using an autoradiographic approach to obtain information concerning the precise neuroanatomical
35
Fig. 3. Inhibition of dopamine (100mM)-stimulated [ S]GTPgS binding
distribution of G-protein activation in response to
dopa-by dopamine receptor antagonists. The D receptor antagonist raclopride2
mine receptor agonists. This is important since receptor
produced a significant inhibitory effect on dopamine-stimulated 35
[ S]GTPgS binding. No significant block was observed with the D1 binding properties do not necessarily correlate with re-antagonist SCH-23390. The data represent the mean6S.E.M. of four ceptor-mediated functional activity. The present technique sections and are representative of two separate experiments. *P,0.05,
offers a novel approach not only to study the functional
**P,0.01 compared with dopamine alone.
neuroanatomy of receptors, but also to localize changes in dopamine receptor subtype selective functions associated [10]. The latter are ultimately responsible for receptor- with pathophysiological conditions.
mediated biological functions.
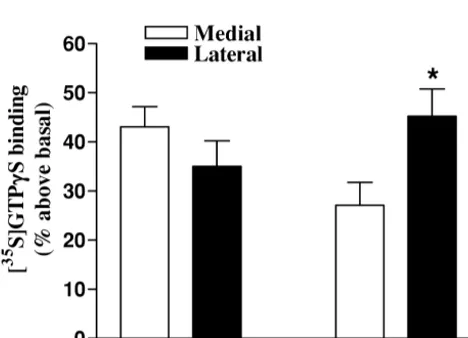
We subsequently investigated the distribution of
dopa-35
mine-stimulated [ S]GTPgS binding in rat striatum. As Acknowledgements shown in Fig. 4, lateral-to-medial and rostral-to-caudal
gradients were observed. Stimulation was significantly This work was supported by NIH Grant NS34886-01A2. higher in the lateral than the medial part of the caudal
striatum, whereas no differences were observed at rostral
levels. Further anatomical and functional studies will be References necessary to clarify the significance of this finding.
How-ever, it is quite possible that the pattern of dopamine- 35
[1] K. Befort, L. Tabbara, B.L. Kieffer, [ S]GTPgS binding: a tool to evaluate functional activity of a cloned opioid receptor transiently expressed in COS cells, Neurochem. Res. 21 (1996) 1301–1307. [2] V. Coronas, S. Krantic, F. Jourdan, E. Moyse, Dopamine receptor
coupling to adenylyl cyclase in rat olfactory pathway: a combined pharmacological-radiographic approach, Neuroscience 90 (1999) 69–78.
[3] B.R. Gardner, D.A. Hall, P.G. Strange, Pharmacological analysis of 35
dopamine stimulation of [ S]-GTPgS binding via human D2shortand D2long dopamine receptors expressed in recombinant cells, Br. J. Pharmacol. 118 (1996) 1544–1550.
[4] B.R. Gardner, D.A. Hall, P.G. Strange, Agonist action at D2(short ) dopamine receptors determined in ligand binding and functional assays, J. Neurochem. 69 (1997) 2589–2598.
[5] M. Geurts, E. Hermans, J.M. Maloteaux, Assessment of striatal D1 and D2 dopamine receptor-G protein coupling by agonist-induced
35
[ S]GTP gamma S binding, Life Sci. 65 (1999) 1633–1645. [6] J.N. Joyce, S.K. Loeschen, J.F. Marshall, Dopamine D-2 receptors
in rat caudate–putamen: the lateral to medial gradient does not correspond to dopaminergic innervation, Brain Res. 338 (1985) 209–218.
35
Fig. 4. Differential stimulation of [ S]GTPgS binding by dopamine (100 [7] S.M. Khan, T.S. Smith, J.P. Bennett Jr., Effects of single and
136 L. He et al. / Brain Research 885 (2000) 133 –136
[8] A. Lorenzen, H. Lang, U. Schwabe, Activation of various subtypes [16] L.J. Sim, D.E. Selley, S.R. Childers, In vitro autoradiography of of G-protein alpha subunits by partial agonists of the adenosine A1 receptor-activated G proteins in rat brain by agonist-stimulated
35
receptor, Biochem. Pharmacol. 56 (1998) 1287–1293. guanylyl 59-[g-[ S]thio]-triphosphate binding, Proc. Natl. Acad. [9] C. Missale, S.R. Nash, S.W. Robinson, M. Jaber, M.G. Caron, Sci. USA 92 (1995) 7242–7246.
Dopamine receptors: from structure to function, Physiol. Rev. 78 [17] L.J. Sim, D.E. Selley, S.I. Dworkin, S.R. Childers, Effects of (1998) 189–225. chronic morphine administration on m opioid receptor-stimulated
35
[10] P.C.G. Nijssen, T. Sexton, S.R. Childers, Opioid-inhibited adenylyl [ S]GTPgS autoradiography in rat brain, J. Neurosci. 16 (1996) cyclase in rat brain membranes: lack of correlation with high-affinity 2684–2692.
opioid receptor binding sites, J. Neurochem. 59 (1992) 2251–2262. [18] P.G. Szekeres, J.R. Traynor, Delta opioid modulation of the binding 35
[11] Y. Odagaki, K. Fuxe, Functional coupling between A1 adenosine of guanosine-59-O-(3-[ S]thio)triphosphate to NG108-15 cell mem-receptors and G-proteins in rat hippocampal membranes assessed by branes: characterization of agonist and inverse agonist effects, J. high-affinity GTPase activity, Br. J. Pharmacol. 116 (1995) 2691– Pharmacol. Exp. Ther. 283 (1997) 1276–1284.
2697. [19] F.I. Tarazi, A. Campbell, S.K. Yeghiayan, R.J. Baldessarini, Locali-[12] P. Onali, M.C. Olianas, Pharmacological and biochemical characteri- zation of dopamine receptor subtypes in corpus striatum and nucleus zation of dopamine receptors mediating stimulation of a high affinity accumbens septi of rat brain: comparison of D1-, D2-, and D4-like GTPase in rat striatum, Biochem. Pharmacol. 36 (1987) 2839–2845. receptors, Neuroscience 83 (1998) 169–176.
[13] A. Rinken, U.B. Finnman, K. Fuxe, Pharmacological characteriza- [20] W.N. Tian, E. Duzic, S.M. Lanier, R.C. Deth, Determinants of 35
tion of dopamine-stimulated [ S]-guanosine 59-(g-thiotriphosphate) a2-adrenergic receptor activation of G proteins: evidence for a 35
([ S]GTPgS) binding in rat striatal membranes, Biochem. Phar- precoupled receptor / G protein state, Mol. Pharmacol. 45 (1994)
macol. 57 (1999) 155–162. 524–531.
[14] D.E. Selley, S. Stark, L.J. Sim, S.R. Childers, Cannabinoid receptor [21] J. Zhu, L. Luo, J. Li, C. Chen, L.Y. Liu-Chen, Activation of the 35
stimulation of guanosine-59-O-(3-[ S]thio)triphosphate binding in cloned human kappa opioid receptor by agonists enhances 35
rat brain membranes, Life. Sci. 59 (1996) 659–668. [ S]GTPgS binding to membranes: determination of potencies and [15] D.R. Sibley, F.J. Monsma Jr., Molecular biology of dopamine efficacies of ligands, J. Pharmacol. Exp. Ther. 282 (1997) 676–684.
![Fig.1. Representativeautoradiogramdepictingdopamine-stimulated[ S]GTP35gS binding in rat striatum](https://thumb-ap.123doks.com/thumbv2/123dok/3138505.1382597/2.612.311.546.435.662/fig-representativeautoradiogramdepictingdopamine-stimulated-s-gtp-binding-rat-striatum.webp)