Oseana, Volume XXV, Nomor 1, 2000 : 1-11 ISSN 0216- 1877
PRODUKTIVITAS DILAMUN
oleh
Muhammad Husni Azkab 1)
ABSTRACT
PRODUCTIVITY IN SEAGRASSES. Productivity is one of the aspects of
seafrass ecology. It is important in analyses of productivity that values be obtained by comparable methods or techniques. Therefore, these values should be expressed in such a way as to comparable among different ecosystems, among different com-munity components of the same system, or among responses of the components to natural or perturbed environmental variables. This paper will discuss the definitions and units which will be used for seagrass productivity, and also present a review of the productivity methods or techniques.
PENDAHULUAN
Pada tahun-tahun terakhir ini, ekosistem lamun telah diketahui sebagai salah satu ekosistem terkaya dan paling produktif, bila dibandingkan dengan produktivitas dari hasil usaha pertanian tropis (McROY 1973; PHILLIPS 1974; ZIEMAN 1975; THAYER
et al. 1975).
Dengan adanya produksi primer yang tinggi ini, maka dapat dikatakan bahwa salah satu fungsi lamun adalah menjaga atau memelihara produktivitas dan stabilitas pantai pesisir dan ekosistem estuaria. Hal ini dapat diartikan bahwa lamun merupakan unsur utama dalam proses-proses siklus yang cukup rumit dan memelihara tingginya produktivitas daerah estuaria dan pantai pesisir.
Peranan lamun dalam siklus tersebut telah disarikan dan disimpulkan oleh WOOD
et al. (1969) sebagai berikut: (a) Lamun mempunyai produktivitas dan kecepat- an tumbuh yang tinggi. Nilai produksi 500-1000 gC/m2/tahun merupakan tipe area dengan produktivitas tinggi bahkan dapat lebih dari dua kali lipat. (b) Daun lamun menyum- bangkan sejumlah besar organisme epifit. Hal ini bahkan sering biomassa epifit sama dengan biomassa daun lamun. (c) Beberapa oraganisme memakan langsung daun-daun lamun. Beberapa dari organisme tersebut memakan langsung material epifit, tetapi umumnya daun-daun yang sudah kering/mati, dan biasanya dikonsumsi sebagai detritus, (d). Banyak material serasah tersimpan dalam sedimen, termasuk material akar yang berguna
1) Balitbang Biologi Laut, Puslitbang Oseanologi-LIPI, Jakarta.
untuk memelihara aktivitas siklus sulfur, (e). Daun-daun yang lebat akan memperlambat arus yang dekat dengan sedimen sehingga akan terjadi sedimentasi. (f). Sistem perakaran dan rimpang (rhizome) akan mengikat sedimen dan dengan bantuan daun akan mencegah erosi.
Produksi primer padang lamun sangat rumit dan memberikan kontribusi problem yang unik untuk mengukur produktivitas di laut. Semua komponen dari total produktivitas harus diukur secara keseluruhan untuk menghitung besarnya produktivitas dalam suatu sistem. Komponen-komponen tersebut termasuk algae bentik, algae epifit, fitoplankton dan lamun. Tiap metode atau tehnik khusus mulai dari tehnik pengukuran produktivitas fitoplankton sampai yang masih dalam percobaan pengukuran produktivitas lamun termasuk epifit yang berasosiasi dengan lamun akan mengalami kerumitan. Hal ini karena pengetahuan tentang produktivitas lamun belum banyak bergeser dari apa yang telah dipelajari oleh PETERSON pada tahun 1913, 87 tahun yang lalu (McROY 1977). Di samping itu, problema utama yang muncul untuk menentukan akurasi produksi lamun akan menyangkut penyimpanan gas-gas metabolik dalam kolom air yang kosong. HARTMAN & BROWN (1967) menunjukkan bahwa tumbuhan laut berbunga (angios- permae) menimbun produksi metabolik O2 dan
CO2 , kemudian akan menggunakan kembali
gas-gas tersebut secara internal. Jadi pengukuran produktivitas sebagai suatu fungsi dari perubahan dalam oksigen terlarut dan 14C tidak begitu akurat karena tidak ada hubungan langsung antara perubahan O2 dan CO2 pada
suatu media dan produksi dari tanaman. Tulisan ini merupakan tinjauan dari hasil produktivitas lamun dengan meng-gunakan beberapa metode atau tehnik pengukuran.
DEFINISI DAN SATUAN
Suatu hal yang sangat penting dalam menganalisis produktivitas lamun dengan cara memperbandingkan beberapa hasil atau nilai yang didapat dari suatu tehnik atau metode yang digunakan. Di samping itu, nilai-nilai tersebut menunjukkan suatu hal yang dapat dibandingkan antara ekosistem yang satu dengan ekosistem yang lain. Pada saat ini sering kita temukan banyaknya istilah dan definisi produktivitas yang dikelompokkan dalam suatu elemen secara umum (WEST- LAKE 1974; WETZEL 1975).
Beberapa definisi dan satuan yang perlu diketahui berkaitan dengan produktivitas yaitu :
1. Hasil yang dipanen (Standing crop) Istilah "standing crop" dapat disamakan dengan "standing stock" yaitu berat material organik yang dapat dipanen pada keadaan nor-mal pada waktu tertentu dari suatu area. Istilah tersebut berasal dari terminologi yang digunakan dalam pertanian yaitu "crop". Standing crop biasanya tidak termasuk pada seluruh populasi tanaman dari suatu jenis tertentu, dengan kata lain umumnya berdasarkan hanya pada bagian atas sedimen dari suatu tanaman air.
2. Hasil (Yield)
Istilah "yield" yaitu suatu hasil panen berdasarkan kecepatan.
3. Biomassa (Biomass)
Biomassa yaitu berat dari semua mate-rial yang hidup pada suatu satuan luas tertentu. Berbeda dengan "standing crop", biomassa termasuk semua tanaman dengan bagian akarnya.
4. Produksi (Production)
Produksi adalah berat bahan organik yang baru pada periode waktu tertentu ditambah dengan setiap kehilangan pada periode tersebut. Dengan demikian produksi merujuk kepada meningkatnya biomassa yang
diamati pada suatu periode ditambah dengan setiap kehilangan oleh respirasi, ekskresi, sekresi, "grasing" dan kematian. Produksi primer secara kuantitas adalah materi organik yang baru yang dihasilkan dari fotosintesa atau penimbunan energi dimana materi tersebut ada. 5. Produktivitas (Productivity)
Produktivitas yaitu kecepatan produksi yang merupakan hasil dari produksi per satuan waktu. Produktivitas biasanya merupakan rata-rata kecepatan pada waktu itu dari beberapa waktu tertentu, seperti satu hari atau satu tahun, karena faktor-faktor aturan dari sistem alam yang menentukan seperti kecepatan, frekuensi, dan perubahan yang tidak teratur pada kecepatan pada saat itu, sehingga dapat diartikan bahwa hanya kecepatan rata-rata yang dapat ditentukan pada studi yang konvensional.
Hilangnya produksi muncul sebagai akibat bukan kehilangan karena predator (kegiatan respirasi ke bentuk CO2 dan
pemanasan, ekskresi dan sekresi materi organik terlarut, patah dan mati) serta kehilangan oleh "grazing". Produktivitas kotor (gross productivity) berdasarkan pada pengamatan biomassa ditambah semua preda-tor dan non-predapreda-tor yang hilang dibagi dengan waktu tertentu. Sedangkan produktivitas bersih (net productivity) adalah kecepatan produksi kotor dari materi organik baru atau fiksasi dari energi dibagi dengan waktu tertentu.
Produktivitas juga merupakan kecepatan produksi dari lamun yang sangat umum dihitung dari perubahan biomassa pada waktu tertentu dengan koreksi dari materi- materi yang hilang.
6. Berat (Weight)
Berat segar (fresh weight) adalah berat dari tanaman tanpa kandungan air. Hal ini biasanya juga diartikan dengan berat basah. Berat basah dapat dikonversi ke berat kering (dry weight).
BIOMASSA
Walaupun lamun tumbuh dalam skala luas dari kerapatannya, tetapi pada kondisi dibawah optimal beberapa jenis dapat menjadi bentuk yang besar dan merupakan padang lamun, yang rapat pada laut dangkal. Lamun mempunyai biomassa daun yang besar, tetapi mayoritas dari biomassa lamun yang di dalam sedimen dan agak sulit untuk mengambilnya karena sistem perakarannya yang dalam (JONES 1986; ZIEMAN 1975).
Beberapa penelitian tentang biomassa telah dipublikasikan, tetapi secara umum data tersebut hanya terbatas pada delapan jenis lamun, dan secara khusus hanya dua jenis yaitu
Zostera marina dan Thalassia testudinum.
Pada daerah semi tropik, 73% dari biomassa dan pengukuran produktivitas adalah jenis T. testudinum, sedangkan pada daerah urgahari (temperate) 92% dilaporkan dari jenis Z. ma-rina. Jadi untuk mahasiswa atau peneliti hanya mendapatkan informasi dari kedua jenis tersebut, sehingga masih tersisa kurang lebih 40 jenis yang belum diteliti secara rinci.
Beberapa pengukuran yang komplit dari biomassa telah dilaporkan, dan mayoritas dari literatur tersebut terdiri hanya dari biomassa daun lamun (standing crop). Daun- daun tersebut adalah sejumlah variabel berat tanaman yang tergantung pada kedaiaman, substrat, nutrien dan musim. Problema utama dalam menentukan total biomassa lamun adalah dalam mengoleksi material akar. Walaupun mayoritas dari material akar didapat dalam lapisan atas sedimen, ZIEMAN (1972) menemukan bahwa akar Thalassia dapat mencapai 4 m di padang lamun yang ber- batu di selatan Florida. Lebih lanjut BURKHOLDER et al. (1959) yang melakukan studi lamun Thalassia di Purto Rico menemukan dari substrat lumpur ke substrat pasir terjadi peningkatan total biomassa dan terjadi penurunan rasio antara daun dan akar/
rimpang. Mereka menemukan rasio daun dengan akar/rimpang 1:3 untuk lumpur halus, 1:5 pada lumpur dan pasir dan 1:7 pada pasir kasar. Berat biomassa daun tertinggi ditemukan pada sedimen pasir kasar/krikil.
Beberapa studi menunjukkan bahwa daun Thalassia biasanya mengandung 15-22% dari total berat kering tanaman, walaupun proporsi ini bervariasi antara 10-45% (ZIEMAN 1975). JONES (1968) pada penelitiannya di Teluk Biscayne menemukan rasio relatif tetap yaitu 3:2:2 untuk daun:rimpang:akar. BURKHOLDER & DOHENY (1968) pada studi mereka terhadap jenis Zostera di Pulau Long menemukan rasio daun: rhizome: akar adalah 2:1:2 pada substrat pasir sedang pada substrat lumpur rasionya adalah 10:1:2.
Pada dua jenis lamun diatas (Thalassia
dan Zostera) rasio daun/akar meningkat pada perubahan subtrat pasir kasar ke substrat lumpur halus. Hal ini menunjukkan bahwa ada efek positif dari lumpur organik terhadap perkembangan tanaman atau sebaliknya suatu kebutuhan dalam perkembangan sistem penyerapan nutrien pada sedimen yang kasar dimana secara umum ada kecenderungan penurunan nutrien dan material organik. Jadi modifikasi efek dari lingkungan sedimen pada rasio dan ukuran daun harus diperhatikan tentang fenologi tanaman. Walaupun demikian respons jenis Thalassia dan Zostera berbeda. Jika Zostera berkembang secara maksimum yang mengindikasikan berat pada sedimen kasar, dimana Thalassia angka maksimumnya pada substrat halus.
Biomassa dan produktivitas lamun merupakan suatu fungsi dari beberapa faktor interaksi yang berarti interpertasi dari beberapa literatur untuk rata-rata dan maksimum "stand-ing crop" adalah sulit. Beberapa hasil penelitian tentang "standing crop" disajikan pada Tabel l. McROY & McMILLAN (1977) telah membuat daftar dari mayoritas publikasi
untuk biomassa dan produktivitas lamun. Mengingat variasi habitat tempat pengambilan contoh (sample), perbedaan waktu pengambilan contoh, jumlah ulangan dan perbedaan tujuan penelitian, maka sangat sukar untuk mengsintesa dari hasil-hasil yang tercantum dalam publikasi tersebut. Hal ini cukup jika kondisi dalam keadaan normal, luas padang lamun yang padat, kesinambungan padang lamun dengan jarak yang besar. Tercatat biomassa maksimun untuk Thalassia lebih dari 8000 g/m2 dan untuk Zostera sekitar 5000 g/ m2 (Tabel 1)
Biomassa atau "standing crop" dari lamun sangat penting dari beberapa aspek. Sebagai produksi primer berarti berfungsi sebagai salah satu sumber makanan. Walaupun beberapa fakta menunjukkan bahwa secara umum lamun bukan berfungsi sebagai makanan primer atau sebagai sumber "graz-ing", tetapi lamun tersebut dapat dimakan langsung atau masuk ke dalam siklus makanan sebagai detritus.
METODE-METODE PRODUKTIVITAS 1. Metode Biomassa (Biomassa methods)
Perhitungan produksi dari komunitas tanaman dengan pengukuran "standing", ma-terial yang hidup telah ditemukan pada beberapa lingkungan, terutama pada komunitas makrofita air tawar dan air laut (WESTLAKE 1965, WETZEL 1965). WESTLAKE (1965) telah mencatat bahwa jika tidak ada material tanaman yang hilang kecuali respirasi antara dua waktu sampling, maka produksi bersih sesuai dengan definisi adalah sama dengan biomassa. Metode ini lebih baik untuk mengukur produksi tanaman tahunan (annual plants), dimana kehilangan akan mudah diukur. Problema akan muncul dan rumit bila dilakukan pada tanaman yang bertahun-tahun (perennial plants).
Tabel 1. Beberapa hasil penelitian tentang biomassa (berat kering dalam gram/m2) (ZIEMAN & WEZEL1980).
Metode biomassa yang lebih mudah, jika hanya mengukur biomassa maksimum atau yang dikehendaki, misalnya biomassa pada satu suksesi waktu seluruh musim pertumbuhan atau perhitungan produksi berdasarkan penambahan-penambahan (WESTLAKE 1965). Metode ini akan lebih akurat jika berdasarkan metode "split-plot" dari WEIGERT & EVANS (1964). Metode ini akan menghitung kehilangan dari mated yang mati dan serasah. Metode ini telah dikembangkan pada studi beberapa tanaman dan aplikasinya sukses pada tanaman payau atau laut.
Ada dua problema dalam menggunakan metode ini pada lamun yaitu: (1). Beberapa
lamun baik yang tumbuh di daerah tropis maupun yang tumbuh di daerah kutub adalah tanaman bertahun-tahun. (2). Lamun tumbuh pada daerah yang arus sedang sampai arus tinggi, sehingga material yang mati dapat terbawa. Kedua hal tersebut dapat menyebabkan produksi tidak dapat dihitung.
PETERSON pada penelitiannya tahun 1913 (ZIEMAN & WETZEL 1980) telah menghitung produksi bersih dari Zostera
ma-rina dengan mengduakalikan biomassa
maksimun pada akhir musim panas. Lebih lanjut GRONTVED (1958) menghitung hasil
Zostera marina per tahun dengan nilai biomassa pada musim semi dari maksimum
akhir musim panas, tetapi dia tidak menggunakan perhitungan koreksi untuk menghitung daun yang hilang selama periode penelitian. Karena hasil ini tidak akurat, maka metode ini hanya untuk perhitungan kasar dari produktivitas dan biasanya jarang digunakan. Dengan pendekatan yang berbeda dari metode di atas, PATRIQUIN (1973) telah menggunakan beberapa assosiasi parameter statistik dengan padang lamun untuk perhitungan kecepatan tumbuh, produksi dan umur Thalassia testudium. Dia menemukan bahwa produksi bersih sama dengan 0,0037 x biomassa (standing crop) berat basah pada suatu area. Dia juga memperoleh beberapa variasi hubungan secara statistik, termasuk perhitungan pertumbuhan rimpang. Tetapi walaupun demikian metode ini harus diuji pada beberapa area yang berbeda untuk membuktikan keakuratannya sehingga dapat diterima secara umum.
2. Metode Penandaan (Marking Methods) Hasil pengkuran produksi dari metode penandaan adalah sebuah peningkatan yang besar dari nilai biomassa mentah. Proses penandaan mengatur (menyediakan) sejumlah biomassa pada waktu tertentu dan dibolehkan pengukuran lebih tepat dari pertumbuhan tanaman pada interval waktu.
Setelah penentuan sebuah plot pada padang lamun yang terpilih, maka area penelitian tersebut ditandai dengan menekan bingkai kawat/besi ke dalam sedimen. Ukuran bingkai bervariasi tergantung jenis lamun yang akan kita teliti dan kerapatan dari padang lamun tersebut. Semua tegakan lamun dalam bingkai tersebut ditandai dengan menggunakan stapler kecil. Hal ini biasanya dimodifikasi untuk menghindari terjadinya patahan pada daun (ZIEMAN 1968, 1975). Setelah 2 atau 4 minggu, semua daun dalam bingkai diambil dengan memotongnya (memakai gunting atau pisau) dan dimasukkan ke dalam kantong plastik untuk dibawa ke laboratorium untuk dianalisis.
Di laboratorium daun-daun tersebut dibagi dua grup, grup pertama adalah daun- daun baru yaitu daun-daun yang muncul sejak mulai waktu penandaan sehingga tidak ada bekas stapler. Grup kedua adalah daun-daun tua yaitu semua daun-daun yang mempunyai tanda stapler. Kemudian daun-daun tersebut dihitung, diukur panjang dan lebar, dicatat daun yang masih mempunyai ujung daun atau tidak. Umumnya daun-daun yang sudah diberi tanda (tidak semuanya) tumbuh antara pangkal daun dengan daun yang di stapler, tetapi untuk daun-daun baru pertumbuhannya adalah panjang daun seluruhnya.
Setelah pengukuran, daun-daun tua dipotong persis dibawah yang di stapler dan dipisahkan dari daun muda. Panjang pertumbuhan yaitu antara yang ditandai dengan pangkal daun. Kemudian dicuci dan dibersihkan dari karbonat dan epifit dan dikeringkan dengan oven 105°C, kemudian ditimbang. Prosedur ini sama perlakuannya pada daun baru. Jangan lupa dicatat jumlah tegakan dan lembar daun dalam bingkai yang diberi tanda dan yang dikoleksi.
Pergantian daun dapat dihitung dengan cara mengstapler semua seludang daun mulai yang tegakan pendek bersama dengan yang lain pada pangkal daun. Hal ini nantinya akan menunjukan bahwa semua yang diberi tanda akan tumbuh dan terlihat perubahannnya dengan memperhatikan pada bagian yang distapler.
Jumlah daun baru yang muncul selama priode penandaan akan diketahui dengan metode penandaan ini biasanya periode koleksi dapat 10, 20, atau 30 hari, sehingga kisaran waktu pada daun yang muncul dapat diketahui. Jika rata-rata berat daun diketahui, maka rata-rata produktivitas daun dapat dihitung.
Penandaan dengan stapler memberi- kan hasil yang cukup baik untuk daun- daun yang lebar seperti Thalassia dan Zostera,
tetapi sangat sulit untuk daun-daun yang sempit seperti Halodule atau Syringodium.
SAND-JENSEN (1975) telah menggunakan spidol tahan air (Penol 70) untuk menandai daun Zostera di Denmark, tehnik ini akan lebih mudah digunakan untuk Halodule atau
Syringodium.
3. Metode Metabolisme
Metode ini merupakan proses foto-sintesa yang merupakan dasar dari reakasi redoks dengan bentuk:
CO2 + 2AH2 h . v (HCOH) + 2A + H2O,
dimana AH2 merupakan sumbangan hidrogen
di air yang merupakan sistem reseptor pigmen (h) untuk energi cahaya (v). Pada fotosintesa yang konvensional di lamun, H2O dan hasil
O2. Secara prakteknya, evolusi dari oksigen
dan pengambilan karbon merupakan segmen utama dari persamaan tersebut, dimana dapat diukur berdasarkan kecepatan fotosintesa.
Metode botol ruang terang-gelap sangat umum digunakan untuk menghitung produktivitas primer dari fitoplankton. Metode ini hanya digunakan pada analisis in situ untuk tanaman berpembuluh. Pada beberapa karakteristik metabolisme dari perubahan gas yang spesifik untuk tumbuhan air berbunga (seperti penimbunan dan penggunaannya), saat ini metode oksigen hasilnya kurang akurat sehingga tidak digunakan. Metode CO2
memerlukan peninjauan kembali sebelum digunakan untuk analisis in situ.
Pada metode oksigen, sampel populasi makrofita tertutup dari pencahayaan, sehingga konsentarsi oksigen disekitar air (C1) ada
kecendrungan menjadi rendah (c2) oleh
respirasi tanaman pada ruang yang gelap, dan meningkat tinggi konsentrasinya (c3) jika
ruang terang. Hal ini akan memberikan respon yang berbeda antara kegiatan produksi fotosintesa dan konsumsi respirasi dari tanaman. Perbedaan (c1-c2) merupakan
aktivitas respirasi pada interval waktu dari inkubasi, sedangkan perbedaan (c3-c2) adalah
kegiatan fotosintesis bersih. Jadi (c3-c1) + (c1,-
c2) = (c3-c2), kegiatan fotosintesa kotor.
Beberapa asumsi telah dibuat bahwa metode oksigen dapat menyebabkan kesalahan besar. Kecepatan respirasi tidak penting pada saat yang sama dengan terang dan gelap, jika fotorespirasi jelas muncul pada tumbuhan berbunga dan algae. Proses lain seperti hewan, bakteri dan algae, dan foto-oksidasi dari senyawa organik menggunakan oksigen secara terpisah dari respirasi tanaman. Lebih lanjut oksigen dapat diproduksi oleh plankton dan algae yang menempel. Sering diassumsikan bahwa sumberdaya produksi oksigen dan pemanfaatannya adalah kecil dalam analisis komunitas dari produksitivitas yang berkaitan dengan makrofita, dimana kadang-kadang kurang tepat untuk lamun yang mempunyai epifit melimpah. Telah diasumsikan bahwa produksi oksigen dari makrofita yang dibuang dari tanaman masuk ke sekitar perairan pada proporsi umum sebagai kecepatan fotosintesa. Selamaperiode fotosintesa, akumulasi oksigen dalam antar sel dan hanya berdiffusi lambat (keluar-masuk) ke perairan (HARTMAN & BROWN 1967). Kecepatan relatif diffusi oksigen yang masuk ke air tidak berhubungan langsung dengan intensitas fotosintesa. Akumulasi dalam antar sel dapat digunakan untuk fotorespirasi dan respirasi gelap mitokhondria oleh kedua sistem dedaunan dan akar pada periode fotosintesa dan dalam keadaan gelap, tanpa adanya efek terhadap konsentrasi oksigen sekitar media.
Menurut ZIEMAN (1975) bahwa gelembung udara, oksigen yang banyak sering dibuang dari tumbuhan air selama periode fotositentesa yang intesif pada air hangat dimana daya larut menurun. Banyak produksi oksigen yang digunakan dalam metabolisme respirasi atau diffusi masuk ke air dalam bentuk larutan. Beberapa metode separti metode O2
pada air mengalir atau metode kurva diurnal dan perubahan penggunanan pada pH air yang berdasarkan prinsip-prinsip bio-kimia.
Pada sistem terbuka, tehnik air mengalir telah dikembangkan oleh ODUM & HOSKIN
(1958) dengan menggunakan dasar asumsi tehnik ruangan. Tetapi asumsi ini pada air yang banyak pada suatu ruangan. Hanya saja, hal ini sangat rumit dimana O2 dan CO2 yang
berdiffusi dari udara ke air. Pada metode kurva diurnal, stasiun harus diseleksi dan konsentrasi O2 di atas air pada stasiun penelitian diukur
pada setiap interval 1-2 jam. Data temperatur dan kedalaman serta beberapa pengukuran arus juga harus diketahui. Dari informasi tersebut, respirasi pada malam had dan angka diffusi O2 dapat dihitung dan metabolisme komunitas
dapat dikalkulasi. Metode ini lebih sederhana, tetapi perhitungan dari diffusi agak sulit dan semua metode di atas dalam penyediaan oksigen kurang tepat (error).
Beberapa penelitian telah menghitung produksi padang lamun dengan mengukur perubahan pada pH dan alkalinitas di perairan terbuka atau sistem air mengalir (PARKE et al. 1958). Metode ini agak lebih akurat dari metode O2 sistem terbuka, tetapi lebih sulit
digunakan dalam praktek secara luas karena perbedaan antara kandungan CO2 ditentukan
oleh pH dan pengukuran alkaliniti pada sistem air laut. SMITH (1974) telah memperlihatkan perubahah kecepatan O2 dan CO2 mungkin
digunakan untuk menghitung produksi yang akurat pada komunitas laut perairan dangkal dengan mengukur pH dan alkaliniti di air laut. Sampel diambil secara terus menerus dengan transek di terumbu karang selama priode arus rendah dan produksi dihitung dari konsumsi CO2. Dengan beberapa kekurangan metode ini
lebih dapat diterima dibandingkan metode yang lain di atas.
Semua metode di atas harus di uji atau diteliti kembali, kecuali untuk metode 14C. Metode 14C, berkembang lebih cepat untuk menghitung produktivitas fitoplankton yang merupakan metode yang pertama sukses diadaptasi untuk mengukur produtivitas tanaman air berpembuluh (WETZEL 1965). Metode 1 4C telah diaplikasikan untuk pengukuran produktivitas beberapa tumbuhan
laut berbunga dan tumbuhan air tawar (DILLON 1971; BRYLINSKY 1971; McROY 1974; BITTAKER & IVERSON 1976). Lebih lanjut metode 14C ini telah dilakukan pada
Thalassia oleh BITTAKER & IVERSON (1976) dan membandingkannya dengan metode penandaan. Perbedaan pengukuran pada kedua metode ini sangat berarti dimana metode 14C sangat akurat yaitu untuk kehilangan an organik (13%), inkubasi absorpsi energi (14%) dan total perbedaan penyinaran dari percoban (7,7%). Hasil ini menunjukkan bahwa pengamatan dengan metode 14C hampir sama dengan produktivitas bersih.
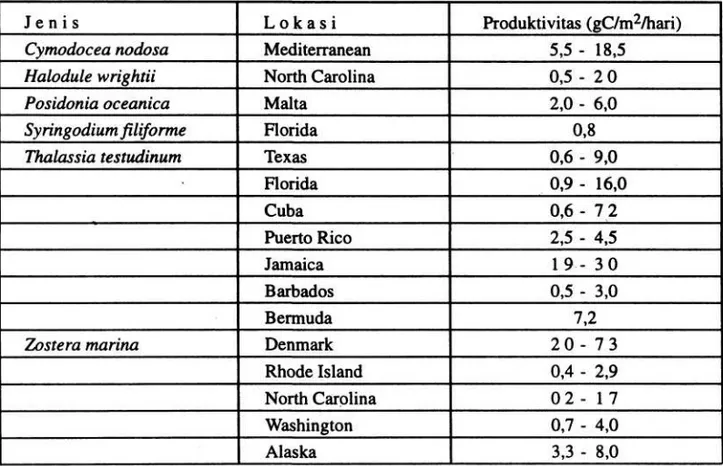
PRODUKTIVITAS
Salah satu aspek dari ekologi lamun telah lama diketahui adalah potensi tingginya produktifitas dari padang lamun. Cukup banyak penelitian tentang produktivitas lamun, tetapi seperti yang diterangkan sebelumnya bahwa hanya terkonsentrasi pada dua jenis yaitu
Zostera marina dan Thalassia testudinum. Pada Tabel 2 disajikan nilai (harga) produktivitas lamun dari beberapa hasil penelitian. Telah dikemukakan bahwa nilai- nilai pada literatur tersebut menunjukkan bahwa produktivitas lamun lebih besar dari suatu area pertanian (WESTLAKE 1963). Pada Z. marina,
produksinya dapat mencapai sampai 8 gC/m2/ hari dan produksi tahunannya adalah 500gC/m2. Disebabkan adanya radiasi yang lebih besar dan lamanya musim pertumbuhan, maka lamun di daerah tropis dapat naik lebih besar produktivitasnya. Komunitas T. testudinum
telah dilaporkan memproduksi sampai 16 gC/m2/hari. Tetapi kita harus berhati-hati karena tingginya angka produktivitas tersebut terutama untuk Thalassia disebabkan menggu- nakan metode oksigen (McROY 1970). Hal ini bertolak belakang dengan perhitungan produksi yang menggunakan metode penandaan untuk jenis yang sama.
Tabel 2. Beberapa hasil produktivitas lamun (ZIEMAN & WETZEL 1980).
Metabolisme relatif dari daun-daun dan bagian di bawah sedimen dapat juga dihitung.
ZIEMAN (1975) menemukan daun Thalassia
biasanya 15-20% dari total berat kering tanaman. Sedangkan PATRIQUIN (1973) telah menghitung pada tanaman yang pendek dan rimpang hanya 10-13% dari produksi bersih. Hal ini merupakan suatu hal yang diluar dugaan, karena tanaman lamun pertum- buhannya mendasar dengan kecendrungan rebah. SAND-JENSEN (1973) menemukan
Zostera di Denmark, rimpangnya meningkat 100-200 g berat kering/m2 selama musim pertumbuhan, sedangkan daun-daun dan tanaman yang berbunganya meningkat 50-230 g berat kering/m2.
DAFTAR PUSTAKA
BIRTTAKER, H. F. and R.L. IVERSON 1976.
Thalassia testudinum productivity: A field comparison of measurement meth- ods. Mar.Biol 37: 39-46.
BURKHOLDER, P.R., L.M. BURKHOLDER and J.A RIVERO 1959. Some chemi- cal constituents of turtle grass,
Thalassia testudinum. Bull. Torre Bot.Club 82 (2): 88
BURKHOLDER, P.R. and T.E. DOHENY 1968. The biology of eelgrass, with special reference to Hempstead and South Oyster Bays, Nassau County,
Long Island, New York, Contr. No.3, Dep.Conser. and Waterways, Town Hempstead, LIN : 120 pp.
DILLON, C.R. 1971. A comparative study of the primary production of estuarine phytoplankton and macrobenthic plants. Ph.D. dissertation, University of North Carolina. Chapel Hill, N.C.: 112 pp. GREENWAY, M. 1974. The effects of
crop-ping on the growth of Thalassia testudinum in Jamaica. Aquaculture 4: 199-206.
GRONTVED, J. 1958. Investigations on the phytoplankton and the primary production in an an oyster culture in Linstjord. Mee.Fra.Dan.Fish.Og. Havundersog Wiser 2 (17): 1-17.
HARTMAN, R.T. and D.L. BROWN 1967. Changes in internal atmosphere of sub-mersed vascular hydrophytes in relation to photosynthesis. Ecology 48: 252-258. JONES, J.A. 1968. Primary productivity by the
tropical marine turtle grass Thalassia testudinum Konig, and its epiphytes. Doctoral dissertation, University of Miami, Fla. : 196 pp.
McROY, C.P. 1970. Standing stocks and other feutures of eelgrass (Zostera marina)
populations on the coast of Alaska.
J.Fish.Res.Bd.Canada 27: 1811-1821. McROY, C.P. 1973. Seagrass ecosystems:
rec-ommendations for research programs.
In :Proc. Intl. Seagrass Workshop, Leiden. C.P. McROY (ed.).
McROY, C.P. 1974. Seagrass productivity: carbon uptake expriments in eelgrass, Zostera marina. Aquaculture 4: 131-137 McROY, C.P. and C. McMILLAN 1977.
Pro-duction ecology and physiology of
seagrasses. In: Seagrass Ecosystems: A scientific perspective (C.P.McRoy and C. Helfferich, eds.). Marcel Dekker,Newyork.Ch.2: 53-88.
ODUM, H.T. 1963. Productivity measure-ments in Texas turtle grass and the ef- fects of dredging an intracoastal chan- nel. Publ.Inst.Mar.Sci.Texas 9 :48-58. ODUM, M. and P.J. MARIS 1958. Comparative
studies on the metabolism of marine waters. Publ.Inst.Mar.Sci.Texas 9: 48-58. PARK, K., D.W.HOOD and H.T. ODUM 1958.
Diurnal pH variation in Texas Bays and its application to primary production estimation. Publ. Inst. Mar. Sci. Texas 5: 47-68.
PATRIQUIN, D.G. 1973. Estimation of growth rate, production and age of the marine angiosperm, Thalassia testudinum
Konig. Carib.J.Sci. 13: 111-123. PHILLIPS, R.C. 1974. Temperate grass plots. In:
Coastal ecological systems of the United States, Vol. 2 (H. T. Odum, B . J. Copeland and E.A. McMahan, eds.). Conservation - Foundation, Washington, D.C.: 244-299.
SAND-JENSEN, K. 1975. Biomass, net pro-duction and growth dynamics in an ee-lgrass (Zostera marina L.) population in Vellerup Vig, Denmark, Ophelia 14: 185-201
SMITH, S.V. 1973.Coral reef carbon dioxide flux. Proc.2nd Int.Coral Reef Symp.
Great Barrier Reef Comm. Brisbane.: 77-85.
THAYER, G.W., D.A. WOLFE and R.B. WIL-LIAMS 1975.The impact of man on seagrass systems. Amer.Scientist 63: 288-296
WESTLAKE, D.F. 1965. Theoretical aspects of comparability of productivity data.
Mem.Ist. Ital.Idrobiol 18 (suppl.): 313- 322.
WESTLAKE, D.F. 1974. Macrophytes. In:
Manual on methods for measuring pri-mary production in aquatic environ- ment, 2nd (R.A. Vollenweider, ed.) Int.Biol.Program Handbook 12. Blackwell Scientific Publ. Oxford.: 25-40.
WETZEL, R.G. 1965. Techniques and problems of primary productivity measurements in higher aquatic plants and periphyton.
Mem.Ist.Itai.Idrobiol. 18 (suppl.):
249-267.
WETZEL, R.G. 1975. Limnology. Saunders, Philadelphia, 743 pp.
WIEGERT,R.G. and F.C. EVANS 1964. Primary production and the disappearance of dead vegetation in an old field in southeastern Michigan. Ecology 45:49-63.
WOOD, E.J.F., W.E. ODUM and J.C. ZEMAN 1969. Influence of seagrass on the pro-ductivity of coastal lagoons. In: Coastal
Lagoons, A Symposium, UNAM- UNESCO. Univ.Nacional Autononia Mexico, MexicO7 D.F.: 495-502. ZIEMAN, J.C. 1968. A study og the growth
and decomposition of the seagrass Thalassia testudinum. M.S. thesis, Univ.Miami, Inst.Mar.Sci.: 217pp. ZEMAN, J.C. 1972. Origin of circular beds of
Thalassia (Spermatophyte: hydro- charitaceae) in South Biscayne Bay, Florida, and their relationship to man-grove hammocks. Bull.Mar. Sci.22
(3):559-574.
ZEMAN, J.C. 1975. Quantitative and dynamics aspects of the ecolofy of turtle grass, Thalassia testudinum. In: Estuarine Reserach, Vol.1, Chemistry, Biology and the Estuarine System. Academic Press, New York.: 541-562.
ZIEMAN, J.C. and ng. wetzel 1980. Produc-tivity in seagrasses: methods and rates. In: Handbook of seagrass biology: an ecosystem perspective. (R.C. Phillips and C.P. McRoy eds.) Garland Publ.Inc. New York.: 87-115.
--- * * * * * ---