Keanekaragaman Burung Berdasarkan Gradien Elevasi di Hutan
Mbeliling dan Sano Nggoang, Flores, Nusa Tenggara Timur
Achmad Ridha Junaid1, Wisnu Wardhana1,Nurul L. Winarni2 1Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam,
Universitas Indonesia Depok, Jawa Barat 2
Research Center for Climate Change – University of Indonesia (RCCC – UI)
achmad.ridha.junaid@gmail.com
Abstrak
Penelitian tentang keanekaragaman burung berdasarkan gradien elevasi telah dilakukan di Hutan Mbeliling dan Sano Nggoang, Flores, Nusa Tenggara Timur. Tujuan penelitian ini adalah untuk mengetahui keanekaragaman dan komposisi burung pada zona elevasi rendah, tengah, dan tinggi. Pengambilan data dilakukan pada bulan Mei dan Juli 2013 di tiga area, yaitu Wae Ndae, Dencang Mese, dan Lengkong Ra’beng. Metode pengambilan data yang digunakan adalah metode titik hitung (point count) dengan jarak antar titik-titik pengamatan sebesar 200 m dan interval waktu pengamatan 10-15 menit. Keseluruhan titik sampel yang diperoleh dikelompokkan menjadi tiga, yaitu titik sampel pada zona elevasi rendah (400-600 mdpl), tengah (700-900 mdpl), dan tinggi (>1.000 mdpl). Berdasarkan hasil pengamatan, ditemukan sebanyak 4.381 individu dari 70 spesies, diantaranya terdapat 18 spesies BST (Burung Sebaran Terbatas) Nusa Tenggara, 4 spesies endemik Flores, dan 8 sub-spesies endemik Flores. Keanekaragaman pada masing-masing zona elevasi berbeda-beda dan menunjukkan pola berbanding terbalik dengan keanekaragaman tertinggi terdapat pada zona elevasi rendah (H’= 3,35; J’= 0,83; D= 0,045), kemudian zona elevasi tengah (H’= 3,22; J’= 0,81; D= 0,055), dan keanekaragaman terendah pada zona elevasi tinggi (H’= 3,10; J’= 0,83; D= 0,065). Spesies burung endemik Flores seperti Serindit flores (Loriculus flosculus), Gagak flores (Corvus florensis), dan Kehicap flores (Monarcha sacerdotum) dan beberapa beberapa spesies endemik lainnya ditemukan di sepanjang zona elevasi, sehingga kawasan Hutan Mbeliling dan Sano Nggoang dari elevasi rendah hingga elevasi tinggi miliki arti penting untuk konservasi spesies burung endemik Flores.
Kata kunci:Keanekaragaman; burung; gradien elevasi; Flores.
Abstarct
Research on bird diversity along elevational gradient has been conducted in the Mbeliling and Sano Nggoang Forests, Flores, East Nusa Tenggara. The aim of this research was to determine the diversity and composition of birds at several elevation zones. Research was conducted on May and July 2013 in three areas, Wae Ndae, Dencang Mese, and Lengkong Ra'beng. Bird survey was conducted using point count method with points spaced at 200 m point interval and was conducted for 10-15 minutes observation at each points. The point samples were classified into three elevation zones, low (400-600 mdpl), middle (700-900 mdpl), and high (> 1,000 mdpl) elevation zones. Seventy bird species and 4.381 individuals were recorded, including 18 species of BST (Burung Sebaran Terbatas) Nusa Tenggara, 4 species endemic to Flores, and 8 sub-species endemic to Flores. Bird diversity varies along elevation zones and bird diversity showed a decrease at higher elevation zones. The highest diversity was found in the low elevation zone (H ' = 3,35; J ' = 0,83; D = 0,045), then the middle elevation zone (H ' = 3,22; J ' = 0.81; D = 0.055), and lowest bird diversity in the high elevation zone (H ' = 3.10; J ' = 0,83; D = 0,065). Endemic bird species such as the Flores hanging-parrot (Loriculus flosculus), Flores crow (Corvus florensis), and Flores monarch (Monarcha sacerdotum) were found at every elevation zones and such results make the Mbeliling and Sano Nggoang Forest have significant importance for the conservation of endemic bird species of Flores.
Pendahuluan
Keberadaan spesies burung pada suatu habitat berkorelasi positif terhadap struktur vegetasi kawasan tersebut (Karr & Roth 1971; Zhijun & Young 2002). Temperatur,
komposisi tumbuhan, gangguan habitat, dan produktivitas habitat juga dapat menjadi faktor yang berpengaruh. Sementara itu, faktor-faktor tersebut dapat sangat bervariasi sepanjang gradien lingkungan, khususnya gradien elevasi (ketinggian tempat) (Able & Noon 1976). Kenaikan elevasi berpengaruh terhadap perubahan fisik lingkungan, jenis, dan jumlah sumber daya yang dimanfaatkan spesies burung untuk melakukan aktivitasnya seperti kawin dan mencari makan, sehingga keanekaragaman dan komposisi burung dapat berbeda seiring peningkatan elevasi (Able & Noon 1976; Cody 1981).
Telah banyak penelitian yang dilakukan terkait keanekaragaman burung berdasarkan gradien elevasi. Beberapa penelitian diantaranya dilakukan oleh Archaya dkk. (2001), Blake & Loiselle (2000), Finch (1989), Navarro (1992), Thakur (2013), dan Zou dkk (2012)
menunjukkan adanya pola keanekaragaman yang dapat terbentuk seiring peningkatan elevasi, yaitu pola berbanding terbalik, hump-shaped, dan pola berbanding lurus. Namun belum banyak informasi yang diketahui tentang keanekaragaman burung berdasarkan gradien elevasi pada kawasan Indonesia, khususnya kawasan Wallacea yang menjadi kawasan dengan tingkat endemisitas burung tertinggi di Indonesia (Coates dkk. 2000).
Salah satu kawasan Wallacea yang penting untuk diketahui keanekaragaman burungnya berdasarkan gradien elevasi adalah Flores, karena 85% DBE (Daerah Burung Endemik) di Nusa Tenggara, terdapat di pulau ini. Kondisi topografi pulau yang bergunung-gunung dengan puncak tertinggi mencapai 2.375 mdpl menyebabkan spesies burung endemik dan non-endemik di Flores tersebar mengikuti gradien elevasi (Sudjatnika dkk. 1995; Trainor dkk. 2000; Rombang dkk. 2002).
Tujuan penelitian ini adalah untuk mengetahui dan membandingkan keanekaragaman burung pada zona elevasi yang berbeda dan mengetahui komposisi spesies burung pada masing-masing zona elevasi. Pengetahuan terkait keanekaragaman dan komposisi burung di zona elevasi berbeda dapat menjadi informasi mendasar dalam penentuan dan evaluasi kawasan prioritas konservasi berdasarkan ketinggian tempat di kawasan Flores.
Tinjauan Teoritis
Keanekaragaman spesies menggambarkan jumlah dan kelimpahan spesies yang ada pada suatu habitat (Begon dkk. 2006; Shukor dkk. 2001). Perbedaan keanekaragaman spesies antar habitat dipengaruhi dua faktor utama, yaitu: variasi ruang (spatial carrying factor) yang meliputi produktivitas habitat, heterogenitas habitat, luas habitat (habitat area), dan kondisi lingkungan yang ekstrem (environmental harshness), serta variasi temporal (temporal carying factor) yang meliputi variasi iklim dan tahapan suksesi (Begon dkk. 2006). Perbedaan keanekaragaman spesies dapat digunakan sebagai indikator keanekaragaman hayati pada suatu bentang alam yang di dalamnya memuat informasi tentang perbedaan kondisi lingkungan antar regional (keanekaragaman beta) (Sharma dkk. 2009).
Perubahan keanekaragaman terkadang memiliki pola sesuai dengan perubahan faktor abiotik yang dikenal sebagai gradien diversitas (Shukor dkk. 2001). Faktor-faktor yang terkait terhadap perubahan gradien diversitas secara umum dapat berhubungan dengan aspek geografis seperti lintang (latitude), ketinggian (altitude), dan kedalaman (depth), atau aspek temporal seperti musim, kondisi suksesi, dan perubahan iklim. Spesies-spesies yang
memiliki batasan terhadap faktor-faktor tersebut menyebabkan spesies tertentu hanya dapat hidup pada relung yang sesuai dan membentuk gradien diversitas (Begon dkk. 2006)
Ricklefs (1997) (dalam Rockwood 2006) mendefinisikan relung (niche) sebagai rentang kondisi dan sumber daya yang memungkinkan bagi suatu organisme untuk dapat bertahan. Secara informal, relung didefinisikan sebagai peran ekologis suatu spesies dalam komunitas. Lebih spesifik, relung dibagi menjadi dua, yaitu fundamental niche dan realized niche. Menurut G.E. Hutchinson (dalam Rockwood 2006), fundamental niche adalah dimensi yang ditempati oleh suatu spesies karena kemampuan toleransi terhadap suatu kondisi fisik dan kimia, serta kemampuan spesies tersebut untuk memenuhi kebutuhan sumber daya tanpa adanya kompetisi dengan spesies lain (Rockwood 2006). Realized niche merupakan bagian yang lebih kecil dari fundamental niche yang dapat ditempati setelah adanya interaksi dengan spesies lain yang bertindak sebagai kompetitor atau predator. Selain dibatasi oleh spesies lainnya, realized niche juga dipengaruhi oleh kemampuan dispersal dari suatu spesies untuk dapat menempati relung yang sesuai (Futuyma 2001; Begon dkk. 2006).
Pola Keanekaragaman Berdasarkan Gradien Elevasi
Ketinggian tempat merupakan salah satu gradien lingkungan yang telah banyak diteliti, khususnya terkait keanekaragaman burung dan mamalia tidak terbang (Guo dkk. 2013). Pola berbanding terbalik atau perubahan keanekaragaman spesies yang menurun seiring
yang memungkinkan ditemukan di alam yaitu pola berbanding terbalik, pola berbanding lurus, dan pola berbukit (hump-shaped) (Rahbek 1997; Begon dkk. 2006). Pola berbanding terbalik menunjukkan adanya perubahan keanekaragaman spesies yang berkurang sering peningkatan elevasi. Penelitian ornitologi yang dilakukan oleh Finch (1989), Navarro (1992), Thakur (2013), dan Blake & Loiselle (2000) menunjukkan adanya pola berbanding terbalik. Faktor utama yang menyebabkan terbentuknya pola tersebut adalah berkurangnya luas area pada elevasi yang lebih tinggi sehingga elevasi tinggi menjadi lebih terisolasi dibandingkan dengan ekosistem yang serupa pada elevasi rendah. Selain itu, berkurangnya sumber daya yang dapat dimanfaatkan, berkurangnya produktivitas primer habitat, dan kondisi iklim ekstrem yang menimbulkan stres fisiologis menjadi faktor yang juga dapat berperan dalam terbentuknya pola berbanding terbalik (Begon dkk. 2006).
Pola berbanding lurus merupakan pola kebalikan dari pola berbanding terbalik. Pola berbanding lurus menunjukkan adanya peningkatan keanekaragaman spesies seiring dengan peningkatan elevasi (Rahbek 1995). Pola berbanding lurus merupakan pola yang jarang ditemukan di alam. Salah satu penelitian ornitologi yang memperlihatkan pola berbanding lurus dilakukan oleh Zou dkk di Pulau Hainan, Cina pada rentang elevasi 200--1.000 mdpl (2012). Menurut McCain (2007), pola berbanding lurus mungkin terjadi saat tidak ada pengurangan luas area habitat seiring peningkatan elevasi. Selain itu, McCain (2009) menyebutkan bahwa faktor yang memungkinkan pola berbanding lurus terbentuk adalah temperatur yang mendukung, ketersediaan air, sejarah evolusi, dan kurangnya gangguan manusia.
Pola yang kerap kali ditemukan di alam adalah pola berbukit (hump-shaped). Pola berbukit memperlihatkan keanekaragaman yang tinggi tidak pada elevasi rendah ataupun elevasi yang tinggi (Begon dkk. 2006). Salah satu penelitian ornitologi yang menunjukkan adanya pola tersebut dilakukan oleh Archarya dkk. (2011). Faktor iklim dan variabel habitat sangat berperan penting dalam terbentuknya pola berbukit. Selain itu, luas area yang
mencukupi dapat memungkinkan peningkatan variasi struktur komunitas dan produktivitas habitat pada elevasi menengah. Umumnya keanekaragaman tinggi terjadi di area perbatasan antara dua habitat, seperti perbatasan antara hutan daratan rendah dan hutan dataran tinggi atau daerah yang memiliki faktor abiotik pendukung yang cukup seperti daerah plateau
Metode Penelitian Tempat dan Waktu
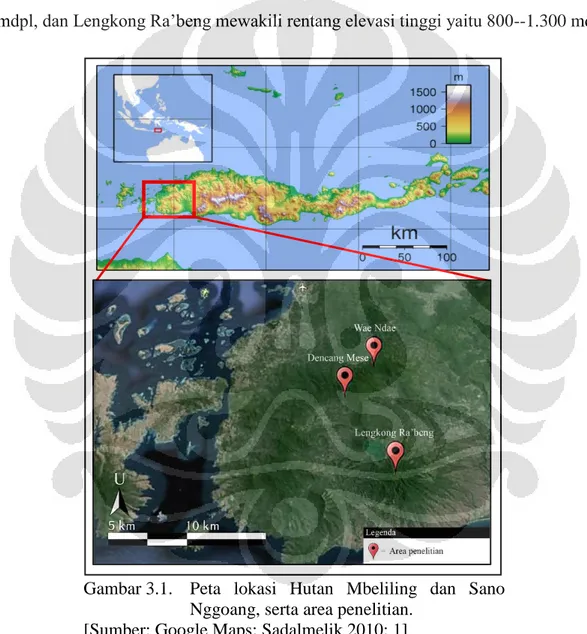
Penelitian dilakukan di Hutan Mbeliling dan Sano Nggoang, Flores (Gambar 1). Area penelitian dibagi menjadi 3 dengan waktu 6 hari pada masing-masing area penelitian. Area tersebut diantaranya Dencang Mese (1-6 Juli 2013) dan Wae Ndae (14-19 Juli 2013) di kawasan Hutan Mbeliling dan Lengkong Ra’beng (8-13 Mei 2013) di kawasan Hutan Sano Nggoang. Area penelitian Wae Ndae mewakili rentang elevasi rendah hingga menengah yaitu 400--950 mdpl, Dencang Mese mewakili rentang elevasi menengah hingga tinggi yaitu 530--1277 mdpl, dan Lengkong Ra’beng mewakili rentang elevasi tinggi yaitu 800--1.300 mdpl.
Gambar 3.1. Peta lokasi Hutan Mbeliling dan Sano Nggoang, serta area penelitian.
[Sumber: Google Maps; Sadalmelik 2010: 1]
Alat
Alat yang digunakan dalam penelitian ini yaitu binokular, kamera, alat tulis menulis, buku panduan spesies burung (Coates dkk. 2000), jam tangan, lembar data pengamatan, GPS
(menentukan koordinat lokasi dan ketinggian tempat), alat perekam suara, dan pita putih (sebagai penanda transek dan titik pengamatan).
Metode Survei Burung
Metode yang digunakan adalah titik hitung (point count). Ditentukan sebanyak 4 transek dengan arah yang berbeda pada setiap area penelitian. Setiap transek terdapat masing-masing 10 titik pengamatan dengan jarak antar titik sebesar 200 m, sehingga setiap transek memiliki panjang 2 km. Interval waktu pengamatan selama 10-15 menit. Pengamatan dilakukan dengan mencatat ketinggian tempat, kode titik, koordinat titik, kondisi cuaca, tipe habitat (hutan sekunder/ hutan primer/ kebun campuran/ savana), waktu awal pengamatan, dan waktu akhir pengamatan, nama spesies dan jumlah individu burung yang ditemui. Analisis Data
Keseluruhan titik sampel yang diperoleh dikelompokkan menjadi tiga berdasarkan tingkat elevasi, yaitu titik sampel pada zona elevasi rendah, tengah, dan tinggi. Zona elevasi dibagi menjadi tiga mengikuti pembagian elevasi Zou (2012), yaitu zona elevasi rendah (400--600 mdpl), zona elevasi tengah (700--900 mdpl), dan zona elevasi tinggi (>1.000 mdpl).
Analisis dilakukan dengan membuat kurva akumulasi spesies untuk melihat kekayaan spesies setiap zona elevasi, kemudian menghitung nilai indeks keanekaragaman Shannon-Wiener, indeks kemerataan Shannon, dan indeks dominansi Simpson. Uji t Hutchenson’s dilakukan untuk menguji signifikansi perbedaan antar nilai indeks keanekaragaman Shannon-Wiener yang diperoleh (dihitung dengan software statistik PAST). Perhitungan dominansi relatif juga dilakukan untuk mengetahui spesies apa yang mendominasi pada suatu zona elevasi. Kesamaan zona elevasi berdasarkan komposisi spesies burung dihitung dengan indeks kesamaan Sorensen. Perhitungan ini dilakukan untuk mengevaluasi kesamaan kondisi habitat pada dua zona elevasi berbeda berdasarkan komposisi spesies burung yang terdapat pada zona elevasi tersebut.
Hasil
Sebanyak 4.381 individu dari 70 spesies yang termasuk ke dalam 32 famili burung tercatat selama penelitian. Sejumlah 18 spesies yang ditemukan adalah BST Nusa Tenggara (92% dari keseluruhan BST yang terdapat di Nusa Tenggara) dan empat spesies endemik Flores (100% dari keseluruhan spesies endemik Flores), serta delapan sub-spesies endemik Flores (88,9% dari keseluruhan sub-spesies endemik Flores). Empat spesies endemik Flores adalah Serindit flores (Loriculus flosculus), Celepuk flores (Otus alfredi), Gagak flores
(Corvus florensis), dan Kehicap flores (Monarcha sacerdotum). Sedangkan delapan sub-spesies endemik Flores antara lain Perkici pelangi (Trichoglossus haematodus weberi), Brencet kerdil (Pnoepyga pusilla everetti), Tesia timor (Tesia everetti everetti), Cinenen gunung (Orthotomus cuculatus everetti), Cikrak dada-kuning (Seicercus montis floris), Sikatan-rimba ayun (Rhinomyias oscillans oscillans), Kipasan flores (Rhipidura diluta diluta), dan Kancilan flores (Pachycephala nudigula nudigula). Informasi lengkap tentang spesies yang ditemukan selama penelitian dapat dilihat pada lampiran 1.
Jumlah spesies yang ditemukan dalam penelitian ini relatif lebih sedikit dibandingkan dengan jumlah spesies yang ditemukan Trainor dan Lesama (2000) di kawasan Mbeliling dan Sano Nggoang. Sedikitnya jumlah spesies burung yang ditemukan pada penelitian ini
dipengaruhi oleh keterbatasan wilayah penelitian. Penelitian ini dilakukan hanya terbatas pada kawasan hutan dan tepi hutan sehingga spesies burung perairan sebagian besar tidak ditemukan. Tutupan tajuk hutan yang rapat juga menyebabkan spesies burung pemangsa (umumnya termati saat terbang tanpa mengepakkan sayap pada siang hari) menjadi susah untuk diamati. Akibatnya, spesies yang teramati dan dianalisis terhadap gradien elevasi umumnya merupakan spesies hutan.
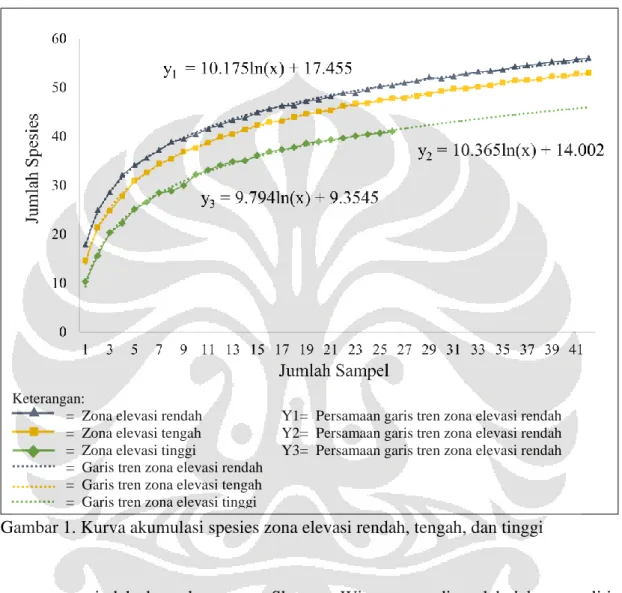
Kurva akumulasi spesies menunjukkan bahwa kekayaan spesies tertinggi terdapat pada zona elevasi rendah (56 spesies), kemudian zona elevasi tengah (53 spesies), dan paling rendah pada zona elevasi tinggi (41 spesies) (gambar 2). Secara umum, kurva zona elevasi rendah, tengah, dan tinggi masih cenderung naik, sehingga memungkinkan ditemukannya spesies baru jika dilakukan penambahan jumlah sampel. Akan tetapi laju pertambahan spesies sudah menurun drastis. Menurut Handerson (2003), jika dilakukan penambahan jumlah sampel, kurva yang cenderung naik akan menyebabkan pertambahan jumlah spesies baru, akan tetapi dengan laju yang sangat rendah. Hal tersebut sekaligus menandakan bahwa jumlah sampel yang diperoleh dianggap cukup untuk menggambar sebagian besar spesies yang terdapat pada suatu habitat.
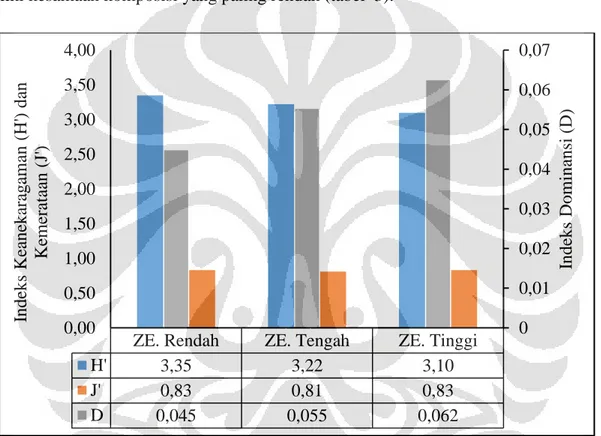
Hasil perhitungan indeks keanekaragaman menunjukkan nilai keanekaragaman nilai yang berbeda-beda antar zona elevasi. Namun demikian, kemerataan jumlah spesies cenderung sama antar zona elevasi (gambar 3). Berdasarkan uji t Hutchenson’s, diketahui nilai indeks keanekaragaman Shannon-Wiener antar zona elevasi berbeda secara signifikan (tabel 1). Dengan demikian dapat dinyatakan bahwa zona elevasi dengan keanekaragaman tertinggi hingga terendah secara berturut-turut adalah zona elevasi rendah, tengah, dan tinggi.
Secara umum, indeks keanekaragaman Shannon-Wiener yang diperoleh dalam penelitian ini tergolong tinggi. Beberapa penelitian ornitologi seperti yang dilakukan oleh Surtikati &
Bahri (2012) dan Swastikaningrum dkk. (2012) menggunakan indeks keanekaragaman Shannon-Wiener untuk kuantifikasi keanekaragaman burung suatu habitat. Dalam penelitian tersebut, dijelaskan bahwa nilai H’ > 3 dianggap tinggi yang menggambarkan habitat
keanekaragaman burung pada habitat tersebut sangat beragam.
Gambar 1. Kurva akumulasi spesies zona elevasi rendah, tengah, dan tinggi
Secara umum, indeks keanekaragaman Shannon-Wiener yang diperoleh dalam penelitian ini tergolong tinggi (gambar 3). Beberapa penelitian ornitologi seperti yang dilakukan oleh Surtikanti & Bahri (2012) dan Swastikaningrum dkk. (2012) menggunakan indeks
keanekaragaman Shannon-Wiener untuk kuantifikasi keanekaragaman burung suatu habitat. Dalam penelitian tersebut, dijelaskan bahwa nilai H’ > 3 dianggap tinggi yang
menggambarkan habitat keanekaragaman burung pada habitat tersebut sangat beragam. Indeks dominansi menunjukkan nilai yang cukup rendah pada setiap zona elevasi namun dengan nilai yang cenderung meningkat pada zona elevasi yang lebih tinggi (gambar 3). Secara umum, rendahnya nilai indeks dominansi dipengaruhi oleh kemerataan jumlah individu antar spesies yang cukup tinggi dengan nilai pada masing-masing zona elevasi
Keterangan:
= Zona elevasi rendah Y1= Persamaan garis tren zona elevasi rendah = Zona elevasi tengah Y2= Persamaan garis tren zona elevasi rendah = Zona elevasi tinggi Y3= Persamaan garis tren zona elevasi rendah = Garis tren zona elevasi rendah
= Garis tren zona elevasi tengah = Garis tren zona elevasi tinggi
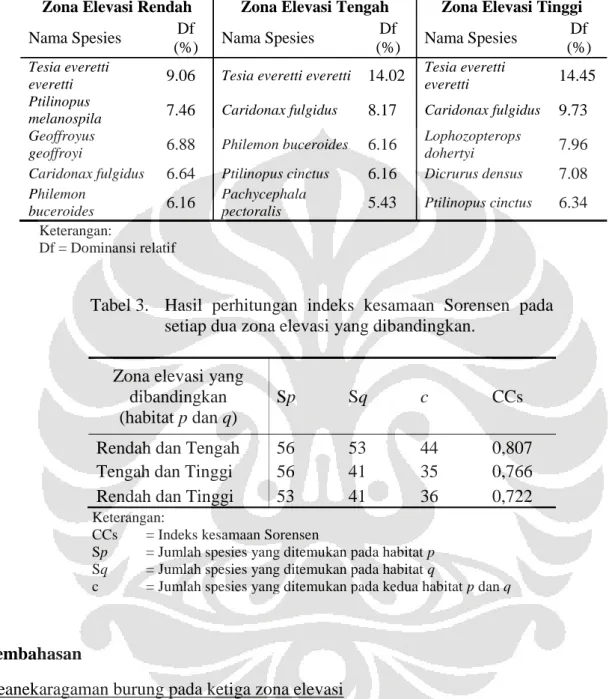
mendekati 1 (gambar 3). Hasil perhitungan indeks dominansi relatif menunjukkan Tesia timor (Tesia everetti everetti) merupakan spesies yang mendominasi pada zona elevasi rendah hingga tinggi. Lima spesies yang mendominasi pada masing-masing zona elevasi dapat dilihat pada tabel 2. Tingginya dominansi relatif Tesia timor diduga dikarenakan penggunaan relung yang luas oleh spesies tersebut serta rendahnya kompetisi dengan spesies yang lain.
Berdasarkan indeks kesamaan Sorensen, diketahui zona elevasi rendah dan tengah memiliki kesamaan komposisi yang paling besar. Sedangkan zona elevasi rendah dan tinggi memiliki kesamaan komposisi yang paling rendah (tabel 3).
Gambar 2. Diagram indeks keanekaragaman Shannon-Wiener, kemerataan Shannon, dan dominansi Simpson pada setiap zona elevasi.
Tabel 1. Hasil uji t Hutcheson’s indeks keanekaragaman zona elevasi rendah, tengah, dan tinggi.
ZE. Rendah ZE. Tengah ZE. Tinggi
H' 3,35 3,22 3,10 J' 0,83 0,81 0,83 D 0,045 0,055 0,062 0 0,01 0,02 0,03 0,04 0,05 0,06 0,07 0,00 0,50 1,00 1,50 2,00 2,50 3,00 3,50 4,00 Inde ks Dominansi (D ) Inde ks Ke ane ka ra g aman (H ') da n Ke mer ataa n (J ') Indeks Keanekaragaman
Shannon-Wiener t hitung df p(same) t tabel α
(H' vs H')
H' 1 vs H' 2 4,3517 3324,8 1.39×10-05 1,64531 0,05
H' 2 vs H' 3 3,1972 1263,0 1.42×10-03 1,64606 0,05
H' 1 vs H' 3 6,5615 1057,3 8.33×10-11 1,64630 0,05
Keterangan:
H’ 1 = Indeks keanekaragaman Shannon-Wiener zona elevasi rendah H’ 2 = Indeks keanekaragaman Shannon-Wiener zona elevasi tengah H’ 3 = Indeks keanekaragaman Shannon-Wiener zona elevasi tinggi
Tabel 2. Lima spesies dengan nilai dominansi relatif tertinggi pada masing-masing zona elevasi.
Tabel 3. Hasil perhitungan indeks kesamaan Sorensen pada setiap dua zona elevasi yang dibandingkan.
Pembahasan
Keanekaragaman burung pada ketiga zona elevasi
Keanekaragaman suatu habitat dipengaruhi oleh dua faktor yaitu jumlah spesies dan kemerataan spesies. Kemerataan merupakan gambaran spesies penyusunnya. Semakin banyak jumlah spesies dan jumlah individu masing-masing spesies tersebar secara merata (kemerataan tinggi), maka keanekaragaman juga akan semakin tinggi (Brewer 1994; Krebs 1985). Berdasarkan indeks kemerataan Shannon (J’) yang ditunjukkan pada gambar 3, individu spesies burung di zona elevasi rendah, tengah, dan tinggi tersebar merata. Menurut Begon dkk. (200) dan Magurran (2004) nilai indeks kemerataan Shannon berkisar antara 0 hingga 1, semakin mendekati 1 maka kemerataan akan tinggi atau jumlah individu setiap
Zona Elevasi Rendah Zona Elevasi Tengah Zona Elevasi Tinggi
Nama Spesies Df (%) Nama Spesies Df (%) Nama Spesies Df (%) Tesia everetti
everetti 9.06 Tesia everetti everetti 14.02
Tesia everetti
everetti 14.45 Ptilinopus
melanospila 7.46 Caridonax fulgidus 8.17 Caridonax fulgidus 9.73 Geoffroyus
geoffroyi 6.88 Philemon buceroides 6.16
Lophozopterops
dohertyi 7.96 Caridonax fulgidus 6.64 Ptilinopus cinctus 6.16 Dicrurus densus 7.08
Philemon
buceroides 6.16
Pachycephala
pectoralis 5.43 Ptilinopus cinctus 6.34
Keterangan:
Df = Dominansi relatif
Zona elevasi yang dibandingkan (habitat p dan q)
Sp Sq c CCs
Rendah dan Tengah 56 53 44 0,807
Tengah dan Tinggi 56 41 35 0,766
Rendah dan Tinggi 53 41 36 0,722
Keterangan:
CCs = Indeks kesamaan Sorensen
Sp = Jumlah spesies yang ditemukan pada habitat p
Sq = Jumlah spesies yang ditemukan pada habitat q
spesies yang terdapat di suatu habitat tersebar merata. Keanekaragaman yang tinggi dapat tercapai jika jumlah individu setiap spesies tersebar merata. Dengan demikian, indeks kemerataan Shannon yang tinggi juga menyebabkan tingginya indeks keanekaragaman Shannon-Wiener yang diperoleh di setiap zona elevasi.
Keanekaragaman burung paling rendah terdapat pada zona elevasi tinggi (H’= 3,10) dibandingkan dengan zona elevasi tengah (H’= 3,22) dan rendah (H’= 3,35) mengindikasikan bahwa adanya batasan-batasan tertentu bagi beberapa spesies burung terhadap zona elevasi tinggi. Hal tersebut juga terlihat dari rendahnya kekayaan spesies burung pada zona elevasi tinggi, sehingga menyebabkan adanya pola penurunan keanekaragaman dan kekayaan spesies burung seiring peningkatan elevasi. Pola tersebut sama dengan pola yang ditemukan pada kawasan Halmahera dan Sulawesi (Coates dkk. 2000; Poulsen & Lambert 2000). Pola yang serupa juga terjadi pada kawasan Papua Nugini, pada elevasi rendah setidaknya terdapat 463 spesies burung daratan, namun mengalami penurunan jumlah spesies seiring peningkatan elevasi. Penurunan kekayaan spesies pada kawasan tersebut merupakan efek dari
berkurangnya area hutan seiring dengan peningkatan elevasi dan terjadinya perubahan struktur dan komposisi hutan (Whitmore & Burnham 1986).
Menurunnya kekayaan spesies burung di zona elevasi yang lebih tinggi dimungkinkan terjadi akibat perbedaan faktor suhu dan variasi tipe habitat. Suhu udara menurun seiring dengan peningkatan elevasi (penurunan 0,56 ºC/kenaikan 100 m) (Terborgh 1977). Hal tersebut menyebabkan suhu udara pada zona elevasi tinggi jauh lebih dingin dibandingkan dengan zona elevasi rendah. Suhu yang rendah dapat menyebabkan stres fisiologis pada beberapa jenis burung sehingga terjadi pembatasan fisiologis (physiological limitation) tertentu terhadap suatu spesies untuk berada pada zona elevasi tinggi (Begon dkk. 2006; MacCain 2009). Kondisi tersebut berdampak pada penurunan jumlah spesies pada zona elevasi tinggi.
Variasi tipe habitat berkorelasi terhadap luas area. Seiring peningkatan elevasi, luas area semakin sempit sehingga variasi habitat pada zona elevasi tinggi lebih rendah dibandingkan dengan zona elevasi rendah dan tengah (Rafe dkk. 1985). Hal tersebut sesuai dengan hasil pengamatan yang menunjukkan zona elevasi rendah memiliki tipe habitat yang lebih
bervariasi, sedikitnya terdapat tiga tipe habitat yaitu hutan primer, hutan sekunder, dan kebun campuran. Sedangkan pada zona elevasi tinggi, luas area yang sempit menyebabkan ragam vegetasi lebih rendah, seperti pada kawasan penelitian yang hanya terdiri dari habitat hutan primer. Kawasan yang memiliki variasi habitat yang tinggi umumnya mampu menyediakan variasi sumber daya yang lebih banyak untuk spesies burung, sebagai tempat mencari makan,
berlindung dari predator, maupun berkembang biak sehingga dapat menurunkan kompetisi pada spesies-spesies yang memanfaatkan sumber daya yang sama (Begon dkk. 2006: 605--606; Nugroho dkk. 2013: 7).
Perbedaan struktur dan komposisi hutan antar zona elevasi juga terlihat memengaruhi penurunan keanekaragaman burung seiring peningkatan elevasi. Kondisi tersebut terlihat dari penurunan ketinggian dan variasi tajuk pada zona elevasi yang lebih tinggi. Trainor dan Lesamana (2000) menyatakan bahwa zona elevasi rendah pada kawasan Mbeliling dan Sano Nggoang merupakan hutan-lembab tropika basah luruh-daun yang memiliki tajuk utama yang rapat dan lapisan tajuk atas yang disusun oleh pohon-pohon emergent. Namun seiring
peningkatan elevasi, pohon emergent semakin jarang sehingga lapisan tajuk atas berkurang. Spesies-spesies pohon pada zona elevasi tinggi juga lebih sedikit dibandingkan dengan zona elevasi rendah. Keanekaragaman burung dipengaruhi oleh variasi tajuk, semakin tinggi dan bervariasinya lapisan tajuk, maka keanekaragaman burung juga akan semakin meningkat. Selain itu, keanekaragaman burung akan semakin meningkat pada kawasan dengan keanekaragaman tumbuhan yang lebih tinggi, hal tersebut terkait dengan penggunaan tanaman tertentu oleh spesies burung sebagai pakan, tempat bertengger, bersarang, ataupun tempat berlindung dari predator (Karr & Roth 1971; Goerck 1999; Joshi dkk. 2012; Zhijun & Young 2002).
Perbedaan struktur dan komposisi habitat juga dapat dilihat melalui komposisi burung yang mendiami habitat tersebut. Kesamaan komposisi burung antar area yang berbeda dapat mengindikasikan area tersebut memiliki kesamaan struktur dan komposisi habitat yang hampir serupa. Berdasarkan hasil perhitungan indeks kesamaan Sorensen, masing-masing zona elevasi yang dibandingkan memiliki kesamaan komposisi spesies burung yang tinggi (nilai mendekati 1). Kesamaan komposisi burung paling besar terjadi pada zona elevasi rendah dan tengah (0,807), sedangkan kesamaan komposisi burung paling kecil terjadi pada zona elevasi rendah dan tinggi (0,722). Hal tersebut mengindikasikan bahwa zona elevasi yang berdekatan cenderung masih memiliki komposisi spesies burung yang sama. Besar atau kecilnya kesamaan komposisi spesies pada kedua zona elevasi yang dibandingkan
dipengaruhi oleh kesamaan struktur vegetasi dan komposisi tumbuhan pada kedua zona elevasi tersebut (Joshi dkk. 2012).
Zona elevasi yang berdekatan cenderung memiliki struktur dan komposisi tumbuhan tidak jauh berbeda. Perubahan struktur dan komposisi tumbuhan pada zona elevasi yang berdekatan terjadi secara bertahap, sehingga spesies burung yang menghuni suatu zona elevasi masih memungkinkan untuk menggunakan sumber daya yang terdapat pada zona
elevasi yang dekat. Sebagai contoh, Kancilan flores (Pachycephala nudigula nudigula) memiliki kelimpahan tertinggi pada zona elevasi rendah (pi= 0,023) namun masih dapat ditemukan pada zona elevasi tengah (pi= 0,018) karena masih memiliki sumber daya yang dapat digunakan pada zona elevasi tengah. Sementara zona elevasi rendah dan tinggi secara jelas telah memiliki struktur vegetasi dan komposisi tumbuhan yang berbeda. Perbedaan yang jelas adalah tinggi tajuk pohon dan kehadiran pohon emergent. Zona elevasi rendah memiliki tinggi tajuk pohon antara 25 m hingga 45 m dan masih ditemukan beberapa pohon emergent, sedangkan pada zona elevasi rendah tinggi tajuk hanya 15 m hingga 30 meter dan sudah tidak ditemukan lagi pohon emergent. Akibatnya, kesamaan komposisi spesies burung pada kedua zona elevasi tersebut paling rendah.
Komposisi spesies burung endemik berdasarkan zona elevasi
Komposisi spesies burung sebaran terbatas Nusa Tenggara, spesies endemik Flores, dan sub-spesies endemik Flores tersebar hampir merata pada pada zona elevasi rendah dan tengah, namun jumlahnya menurun pada zona elevasi tinggi (lihat lampiran 2). Celepuk flores (Otus alfredi) adalah spesies endemik Flores yang tidak ditemukan di zona elevasi rendah, sedangkan Serindit flores (Loriculus flosculus), Gagak flores (Corvus florensis), dan Kehicap flores (Monarcha sacerdotum) dapat ditemui sepanjang zona elevasi di area
penelitian. Belum banyak informasi yang diketahui tentang ekologi Celepuk flores, namun tiga spesimen untuk keperluan taksonomi yang ditemukan oleh Alfred Everett pada elevasi 3.500 kaki (1.066 mdpl) mengindikasikan bahwa Celepuk flores merupakan burung
pegunungan (Mess 2006; Verhoye & Holmes 1998). Berdasarkan hasil pengamatan, Celepuk flores ditemui sebanyak dua kali pada elevasi 870 mdpl di habitat hutan primer sehingga diduga spesies tersebut sangat bergantung pada habitat hutan. Hal yang serupa diutarakan oleh Trainor & Lesmana (2000) bahwa Celepuk flores hanya hidup di hutan tropika pegunungan bawah (1000-1.800 mdpl) dan sangat rentan terhadap perubahan habitat terutama akibat penebangan pohon oleh aktivitas manusia.
Zona elevasi tinggi secara umum memiliki jumlah spesies endemik yang lebih rendah dibandingkan dengan zona elevasi rendah dan tengah, akan tetapi memiliki jumlah sub-spesies endemik Flores yang lebih banyak. Hal tersebut dikarenakan Brencet kerdil (Pnoepyga pusilla everetti) dan Cikrak dada-kuning (Seicercus montis floris) hanya ditemukan pada zona elevasi tinggi. Menurut Verhoye & Holmes (1998), kedua spesies tersebut umum pada elevasi lebih dari 1.200 mdpl, sedangkan elevasi terendah yang pernah tercatat spesies tersebut ditemukan yaitu pada elevasi 900 mdpl. Hasil pengamatan selama
penelitian juga menunjukkan bahwa Brencet kerdil dan Cikrark dada-kuning hanya ditemukan pada elevasi di atas 1.100 mdpl.
Dengan demikian, diketahui bahwa zona elevasi rendah hingga tinggi memiliki jumlah spesies endemik relatif sama, sehingga habitat hutan sepanjang elevasi memiliki nilai penting untuk konservasi spesies burung endemik di kawasan Hutan Mbeliling dan Sano Nggoang. Terutama untuk empat jenis endemik Flores yang dapat ditemukan pada habitat zona elevasi rendah hingga tinggi. Zona elevasi rendah, tengah, dan tinggi memiliki tingkat gangguan habitat yang relatif sama. Selain itu terdapat potensi gangguan habitat yang mungkin dapat merusak eksistensi spesies-spesies burung baik endemik maupun non-endemik terutama spesies-spesies yang sangat rentan terhadap perubahan lingkungan di Hutan Mbeliling dan Sano Nggoang seperti Celepuk flores dan Sikatan-rimba ayun. Tekanan tersebut berupa pembalakan kayu secara ilegal, perambahan hutan untuk perluasan lahan perkebunan, kebakaran, hingga tekanan yang langsung berdampak pada spesies burung, yaitu perburuan liar (Trainor & Lesmana 2000). Sehingga dibutuhkan upaya bersama dengan masyarakat setempat, organisasi yang terkait, dan pemerintah untuk menjalankan upaya konservasi pada Hutan Mbeliling dan Sano Nggoang dari elevasi rendah hingga tinggi.
Kesimpulan
Terdapat sekitar 70 spesies burung yang dapat ditemukan di Hutan Mbeliling dan Sano Nggoang, 56 spesies diantaranya ditemukan di zona elevasi rendah, 53 spesies di zona elevasi tengah , dan 41 spesies di zona elevasi tinggi. Keanekaragaman burung antara zona elevasi di Hutan Mbeliling dan Sano Nggoang berbeda satu sama lain dan menurun seiring dengan peningkatan zona elevasi. Keanekaragaman tertinggi terdapat pada zona elevasi rendah (H’: 3,35), kemudian zona elevasi tengah (H’: 3,22), dan keanekaragaman terendah pada zona elevasi tinggi (H’: 3,10). Penurunan kekayaan spesies dan keanekaragaman burung di Hutan Mbeliling dan Sano Nggoang diduga merupakan efek dari berkurangnya area, penurunan suhu, perubahan struktur dan komposisi hutan, serta penurunan kompleksitas habitat pada zona elevasi pada zona elevasi yang lebih tinggi. Selain itu spesies burung endemik yang relatif tersebar merata pada berbagai zona elevasi menggambarkan pentingnya kawasan Hutan Mbeliling dan Sano Nggoang dari elevasi paling rendah hingga tertinggi bagi konservasi spesies endemik.
Saran
1. Perlu dilakukan penelitian lebih lanjut mengenai keanekaragaman burung dan
hubungannya terhadap struktur vegetasi sepanjang gradien elevasi di Hutan Mbeliling dan Sano Nggoang. Pengambilan data burung dan data struktur vegetasi ataupun data kondisi habitat diharapkan dapat menjelaskan faktor utama yang menyebabkan penurunan
kekayaan dan keanekaragaman burung pada zona elevasi yang lebih tinggi.
2. Perlindungan spesies dan habitat burung di Hutan Mbeliling dan Sano Nggoang harus dilakukan dari zona elevasi rendah hingga tinggi karena habitat sepanjang gradien elevasi memiliki nilai penting untuk konservasi burung endemik dan non-endemik di kedua hutan tersebut.
Daftar Referensi
Able, K.P. & B.R. Noon. 1976. Avian Community structure along elevational gradients in the Northeastern United States. Dalam: F.L. Waterhouse, M.H. Mather & D. Seip. 2002. Distribution and abundance of birds relative to elevation and biogeoclimatic zones in coastal old-growth forests in southern British Columbia. Journal of Ecosystems and Management2(2): 1--13.
Archarya. 2011. Elevational gradients in bird diversity in the Eastern Himalaya: an evaluation of distribution patterns and their underlying mechanisms. PLoS ONE6(12): 1--14.
Begon, M., C.R. Townsend & J.L. Harper. 2006. Ecology: from individual to ecosystems. 4th ed. Blackwell Publishing, Oxford: xii+738 hlm.
Blake, J.G. & B.A. Loiselle. 2000. Diversity of birds long an elevational gradien in the Cordillera Central, Costa Rica. The Auk117(3): 663--686.
Brewer, R. 1994. The science of ecology. 2nd ed. Sounders Collage Publishing, Washington DC: xviii+773 hlm.
Coates, B.J., K.D. Bishop & D. Gardner. 2000. Panduan lapangan burung-burung di kawasan
Wallacea: Sulawesi, Maluku, dan Nusa Tenggara. Edisi Bahasa Indonesia. BirdLife
International-Indonesia Programme & Dove Publication Pty. Ltd., Bogor: xiv+247 hlm.
Finch, D.M. 1989. Habitat use and habitat overlap of riparian birds in three elevational zones. Ecology
70(4): 866--880.
Goerck, J.M. 1999. Distribution of birds along an elevational gradient in the Atlantic forest of Brazil: implications for the conservation of endemic and endangered species. Bird Conservation
International 9: 235--253.
Guo, Q., Douglas A.K., Zhongyu S., Hongxiao L., Liangjun H., Hai R. & Jun W. 2013. Global variation in elevational diversity patterns. Scientific Reports3: 1--7.
Joshi, K.K., D. Bhatt & A. Thapliyal. 2012. Avian diversity and its association with vegetation structure in different elevational zones of Nainital district (Western Himalayan) of Uttarakhand.
International Journal of Biodiversity and Conservation4(11): 364-376.
Karr, J.R. & R.R. Roth. 1971. Vegetation structure and avian diversity inseveral New World Areas.
The American Naturalist105: 423--435.
Krebs, C.J. 1985. Ecology: The experimental analysis of distribution and abundance. 3rd ed. Harper Collins Publishers, Inc., New York: xv+800 hlm.
Magurran, A.E. 2004. Measuring biological diversity. Blackwell Publishing, Victoria: viii+256 hlm.
Forktail18: 127--146.
McCain, C.M. 2005. Elevational gradients in diversity of small mammals. Ecology86(2): 336--372. McCain, C.M. 2007. Area and mammalian elevational diversity. Ecology88(1): 76--86.
McCain, C.M. 2009. Global analysis of bird elevational diversity. Global Ecology and Biogeography
18: 346--360.
Mees, G.F. 2006. The avifauna of Flores (Lesser Sunda Islands). Zool. Med. Leiden25(viii): 1--261 Navarro S., A.G. 1992. Altitudinal distribution of bird in the Sierra Madre Del Sur, Guerrero, Mexico.
The Condor94: 29--39.
Nugroho, M.S., S. Ningsih & M. Ihsan. 2013. Keanekaragaman jenis burung pada areal Dongi-Dongi di kawasan Taman Nasional Lore Lindu. Warta Rimba1(1): 1--10.
Poulsen, M.K. & F.R. Lambert. 2000. Altitudinal distribution and habitat preferences of forest Bird on Halmahera and Buru, Indonesia: implications for conservation of Mollucan avifaunas. Ibis142: 566--586.
Rafe, R. W., M. B. Usher, & R. G. Jefferson. 1985. The influence of area and habitat on species richness. Journal of Applied Ecology22(2): 327--335.
Rahbek, C. 1995. The elevational gradient of spesies richness: a uniform pattern?. Ecography18: 200--205.
Rahbek, C. 1997. The relationship among area, elevation, and regional species richness in Neotropical Birds. The American Naturalist149(5): 875--902.
Rockwood, L.L. 2006. Introduction to population ecology. Blackwell Publishing, Victoria: xi+339 hlm.
Rombang, W.M., C. Trainor & D. Lesmana. 2002. Daerah penting bagi burung: Nusa Tenggara. PHKA/BirdLife Indonesia, Bogor: vi+158 hlm.
Sadalmelik. 2010. 1 hlm. Flores tophography. http://commons.wikimedia.org/wiki/ File:Flores_Locator_Topography.png. 25 Februari 2014, pk. 13:38 WIB.
Sharma, C.M., S. Suyal, S. Gairola & S.K. Ghildiyal. 2009. Secies richness and diversity long an altitudinal gradien in Moist Temperas Forest of Garhwal Himalaya. Journal of American Science
Shukor, Md. N., Z. Batin & Z. Akbar. 2001. Elevational diversity pattern of non-volan small
mammals on Mount Nuang, Hulu Langkat, Selangor. Online Journal of Biological Science 1(11): 1081--1084.
Sudjatnika, P. Jepson., T.R. Soehartono, M.J. Crosby & A. Mardiastuti. 1995. Conserving indonesian
biodiversity: the endemic bird area approach. BirdLife International Indonesia Programme.
Bogor: xi+221 hlm.
Surtikati, H.K. & S. Bahri. 2012. Environmental factors and biodiversity reflecting water quality: case study of raising livestock. Journal of Life Sciences and Technology1(1):1-7
Swastikaningrum, H., B. Irawan & S. Hariyanto. 2012. Keanekaragaman jenis burung pada berbagai tipe pemanfaatan lahan di kawasan muara Kali Lamong, perbatasan Surabaya - Gresik. ?(?) : 1-13. Terborgh, J. 1977. Bird species diversity on an andean elevational gradient. Ecology58(5):
1007--1019.
Thakur, M.L. 2013. Bird species composition along the altitudinal gradient in Himachal Pradesh (Western Himalaya), India. International Jurnal of Advance Biological Research3(4): 556--562. Trainor, C. & D. Lesmana. 2000. Gunung berapi, burung-burung khas, tikus raksasa, dan tenun ikat
yang menawan: identifikasi kawasan yang memiliki arti penting bagi keanekaragaman hayati
global di Flores, Nusa Tenggara. Report No. 11. PKA/BirdLife/WWF, Bogor: xvi+269 hlm.
Verhoye, J. & D.A. Holmes. 1998. The bird of the island of flores: A Review. Kukila10: 3--59. Whitmore, T.C. & C.P. Burnham. 1986. Tropical rainforest of the far east. Oxford University Press,
Great Britain: xvi+350 hlm.
Zhijun, W. & S.S. Young. 2002. Differences in bird diversity between two swidden agricultural sites in mountainous terrain, Xishuangbanna, Yunnan, China. Elsevier: 1-13.
Zou, F.S., G.Z. Chen, Q.F. Yang & Y.D. Li. 2012. Bird species richness along elevational gradien in a forest at Jiangfengling, Hainan Island, China. Zoological Studies51(3): 362--371.
Lampiran 1. Daftar spesies burung yang ditemukan selama penelitian di Hutan Mbeliling dan Sano Nggoang
No. Nama Lokal Nama Ilmiah Presensi
IUCN Red L is t (v er 3 .1 ) CIT E S Ca teg o ry 2011 Status Perlindungan (PP No. 7 Tahun 1999) 1 2 3 Accipitridae
1 Elang flores Nisaetus floris × × CR ya
Megapodidae
2 Gosong kaki-merah Megapodius reinwardt reinwardt LC ya
Phasianidae
3 Ayam-hutan hijau Gallus varius × LC tidak
4 Delimukan zamrud Chalcophaps indica indica LC tidak
5 Pergam hijau Ducula aenea × × LC tidak
6 Pergam
punggung-hitam Ducula lacernulata sasakensis LC tidak
7 Uncal buau Macropygia emiliana × × LC tidak
Columbidae
8 Uncal loreng Macropygia unchall × × LC tidak
9 Uncal Macropygia sp. × × LC tidak
10 Walik putih Ptilinopus cinctus LC tidak
11 Walik kembang Ptilinopus melanospila LC tidak
12 Tekukur biasa Streptophelia chinensis × × LC tidak
Psittaculidae
13 Nuri pipi-merah Geoffroyus geoffroyi LC Appdx II tidak
14 Serindit flores Loriculus flosculus EN Appdx II tidak
15 Betet-kelapa paruh-besar Tanygnathus megalorynchos × × LC Appdx II tidak 16 Perkici pelangi Trichoglossus haematodus
weberi LC Appdx II tidak
Cuculidae
17 Wiwik unciung Cacomantis sepulcralis LC tidak
18 Bubut alang-alang Centropus bengalensis × LC tidak
19 Kangkok sunda Cuculus lepidus × × LC tidak
Strigidae
20 Celepuk flores Otus alfredi × × EN Appdx II tidak
21 Celepuk maluku Otus magicus albiventris × × LC Appdx II tidak
22 Celepuk wallacea Otus silvicola × LC Appdx II tidak
Apodidae
23 Walet sapi Collocalia esculenta LC tidak
24 Walet sarang-putih Collocalia fusciphaga × × LC tidak
Alcedinidae
25 Raja-udang erasia Alcedo atthis × × LC ya
26 Cekakak tunggir-putih Caridonax fulgidus LC ya
27 Udang-merah api Ceyx erithacus × LC ya
Meropidae
29 Kirik-kirik australia Merops ornatus × LC tidak
Picidae
30 Caladi tilik Dendrocopos moluccensis LC tidak
Pittidae
31 Paok la'us Pitta elegans LC ya
Campephagidae
32 Kepudang-sungu sumba Coracina dohertyi LC tidak
33 Kepudang-sungu topeng Coracina personata × × LC tidak
34 Sepah kerdil Pericrocotus lansbergei LC tidak
Dicruridae
35 Srigunting wallacea Dicrurus densus LC tidak
Oriolidae
36 Kepudang kuduk-hitam Oriolus chinensis LC tidak
Corvidae
37 Gagak flores Corvus florensis EN tidak
38 Gagak kampung Corvus macrorhynchos × × LC tidak
Turdidae
39 Anis nusa-tenggara Zoothera dohertyi × NT tidak
40 Anis kembang Zoothera interpres NT tidak
Timaliidae
41 Brencet kerdil Pnoepyga pusilla everetti × × LC tidak
Acanthizidae
42 Remetuk laut Gerygone sulphurea × × LC tidak
Cettiidae
43 Tesia timor Tesia everetti everetti LC tidak
Cisticolidae
44 Cici merah Cisticola exilis × × LC tidak
Sylviidae
45 Cinenen gunung Orthotomus cuculatus everetti × × LC tidak
46 Cikrak dada-kuning Seicercus montis floris × × LC tidak
Zosteropidae
47 Opior paruh-tebal Heleia crassirostris LC tidak
48 Opior jambul Lophozosterops dohertyi
subcristatus LC tidak
49 Kacamata biasa Zosterops palpebrosus LC tidak
50 Kacamata wallacea Zosterops wallacei × LC tidak
Muscicapidae
51 Sikatan kepala-kelabu Culicicapa ceylonensis × LC tidak
52 Sikatan dada-merah Ficedula dumetoria NT tidak
53 Sikatan-rimba ayun Rhinomyias oscillans oscillans × LC tidak
Monarchidae
54 Kehicap ranting Hypothymis azurea prophata LC tidak
55 Kehicap flores Monarcha sacerdotum EN tidak
56 Seriwang asia Terpsiphone paradisi LC tidak
57 Kipasan flores Rhipidura diluta diluta LC tidak
58 Kipasan dada-hitam Rhipidura rufifrons × × LC tidak
Pachycephalidae
59 Kancilan flores Pachycephala nudigula nudigula × LC tidak
60 Kancilan emas Pachycephala pectoralis
fulvitincta LC tidak
Sturnidae
61 Tiong emas Gracula religiosa LC Appdx II ya
Meliphagidae
62 Cikukua tanduk Philemon buceroides LC tidak
Nectarinidae
63 Burung-madu kelapa Anthreptes malacensis × × LC ya
64 Burung-madu sriganti Nectarinia jugularis LC ya
65 Burung-madu matari Nectarinia solaris solaris LC ya
Dicaeidae
66 Cabai gesit Dicaeum agile × × LC tidak
67 Cabai emas Dicaeum annae LC tidak
68 Cabai dahi-hitam Dicaeum igniferum LC tidak
Estrildidae
69 Bondol taruk Lonchura molucca × × LC tidak
70 Bondol peking Lonchura punctulata × × LC tidak
Keterangan:
1= Zona elevasi rendah 2= Zona elevasi tengah 3= Zona elevasi tinggi
Lampiran 2. Kehadiran spesies endemik pada zona elevasi tertentu di Hutan Mbeliling dan Sano Nggoang
Nama Ilmiah Z.E.
Rendah Z.E. Tengah Z.E. Tinggi BST Nusa Tenggara Nisaetus floris
Ducula lacernulata sasakensis
Otus silvicola Caridonax fulgidus Coracina dohertyi Pericrocotus lansbergei Dicrurus densus Zoothera dohertyi
Tesia everetti everetti*
Heleia crassirostris
Lophozosterops dohertyi subcristatus*
Zosterops wallacei
Rhinomyias oscillans oscillans*
Rhipidura diluta diluta*
Pachycephala nudigula nudigula*
Nectarinia solaris solaris
Dicaeum annae
Dicaeum igniferum
Spesies Endemik Flores
Loriculus flosculus
Otus alfredi
Corvus florensis
Monarcha sacerdotum
Sub-spesies Endemik Flores
Trichoglossus haematodus weberi
Pnoepyga pusilla everetti
Orthotomus cuculatus everetti
Seicercus montis floris
JUMLAH 22 22 19
Keterangan:
BST = Burung Sebaran Terbatas