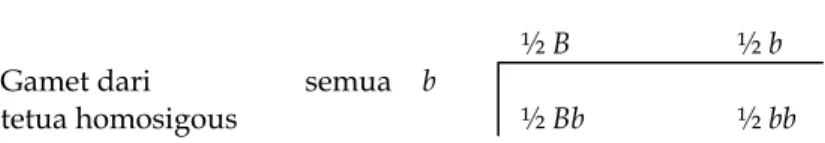G
ENETIKA
D
ASAR
ab ini memberikan tinjauan tentang genetika dasar. Ini terfokus pada prinsip-prinsip umum mengenai genetika yang terjadi pada hewan normal yang sehat. Pengecualian atau penyimpangan dari prinsip-prinsip ini seringkali merupakan landasan tentang penyakit-penyakit keturunan, yang akan didiskusikan pada bab-bab berikutnya.
B
Kromosom
Jika biak sel darah putih yang membelah dengan cepat diperlakukan dengan alkaloid kolkisin (yang menghentikan pembelahan sel), dan sel tersebut kemudian diwarnai dan dilihat di bawah mikroskop cahaya, struktur yang disebut kromosom menjadi dapat dilihat secara jelas. Kromosom tersebut tersebar secara acak dalam kelompok-kelompok, dan setiap kelompok mengandung semua kromosom yang hanya berasal dari satu sel. Area genetika yang terkait erat dengan kromosom dinamakan sitogenetika.
Untuk mempelajari kromosom secara lebih mendalam, sekelompok kromosom dipilih dan difoto untuk diperoleh gambarnya, seperti ditunjukkan hasilnya pada Gambar 1.1a. Tiap unit pada gambar tersebut terdiri atas dua struktur seperti batang yang digabung bersama pada satu titik sempit. Tiap struktur seperti batang itu adalah kromatid, dan penyempitan tersebut adalah sentromer. Dua kromatid yang digabung pada sentromer baru saja terbentuk dari satu kromosom asli. Jika pembelahan sel tadi dibiarkan terus berlangsung, sentromer akan memisah dan masing-masing kromatid yang terpisah kemudian dinamakan kromosom baru. Untuk lebih mudahnya, kita bicara tentang tiap pasang kromatid yang
digabung pada sentromernya sebagai satu kromosom, yang sebenarnya merupakan kromosom yang baru saja menimbulkannya.
Dari hasil cetakan fotograf, semua kromosom dipotong secara individual dengan gunting, dan selanjutnya diatur secara berurutan berdasarkan ukurannya pada selembar kertas. Pengaturan semacam ini memberikan suatu gambaran komplemen kromosom secara lengkap atau kariotipe dari satu sel (Gambar 1.1b). Apabila pengaturan kromosom seperti di atas dilakukan pada banyak individu sehat yang normal dari kedua jenis kelamin spesies mamalia atau burung, maka terdapat dua fakta yang jelas: tiap spesies mempunyai kariotipe yang khas, dan dalam setiap spesies, setiap jenis kelamin mempunyai kariotipe yang khas.
Kariotipe dari spesies yang berbeda mempunyai bentuk, ukuran dan jumlah kromosom yang berbeda juga. Pada setiap spesies, semua kromosom dalam sel selalu berpasangan. Pada individu-individu dari satu jenis kelamin tertentu, kedua kromosom dari setiap pasangan mempunyai bentuk dan ukuran yang sama. Pada jenis kelamin yang lain, semua kromosom juga selalu berpasangan, tetapi ada satu pasang kromosom terdiri dari dua kromosom yang bentuk dan ukurannya berbeda. Pada sepasang kromosom ini, satu kromosom mempunyai bentuk dan ukuran sama seperti salah satu pasang kromosom pada jenis kelamin yang disebutkan pertama.
Perbedaan kariotipe antara dua jenis kelamin tersebut merupakan kunci untuk penentuan jenis kelamin. Pada mamalia, sepasang kromosom yang bentuk dan ukurannya berbeda terdapat pada jantan, dan disebut kromosom X dan Y. Satu dari semua pasangan kromosom dalam sel mamalia betina, terdiri dari dua kromosom X. Jadi pada mamalia, individu jantan adalah XY dan individu betina adalah XX. Kromosom X dan Y dikenal sebagai kromosom kelamin. Pada burung, kromosom kelamin mempunyai nama yang berbeda, dan berkaitan dengan jenis kelamin, penamaannya berlawanan dengan mamalia: dimana burung jantan adalah ZZ dan burung betina adalah ZW. Untuk memudahkan pemahaman, kita akan menggunakan sistem penamaan pada mamalia dalam diskusi-diskusi berikutnya, walaupun semua pernyataan dapat digunakan pada burung jika penamaan jenis kelaminnya dibalik.
Kromosom selain kromosom kelamin dinamakan autosom. Pada setiap spesies, jantan dan betina mempunyai satu set autosom yang sama, dalam bentuk berpasangan. Kromosom kelamin dan autosom yang secara bersama-sama terdapat dalam satu sel disebut genom, yang merupakan total set kromosom dalam satu sel. Genom yang terdiri dari pasangan-pasangan kromosom dikatakan diploid, dan dua kromosom dalam setiap pasangan dinamakan homolog. Untuk menekankan bahwa kromosom-kromosom tersebut selalu berpasangan, jumlah total kromosom dikatakan sebagai 2n, dimana n adalah jumlah pasangan. Sebagai contoh, jumlah kromosom kariotipe yang diilustrasikan pada Gambar 1.2 adalah 2n = 38. Agar dapat
mengidentifikasi setiap pasang kromosom dalam satu kariotipe, pasangan-pasangan autosom diberi label sesuai dengan aturan main yang telah disetujui secara internasional, seperti dapat dilihat pada Gambar 1.1b. Dua kromosom kelamin biasanya ditempatkan pada urutan terakhir.
(b) A B C F E D
Gambar 1.1. (a) Kromosom kucing jantan, sebagaimana dilihat melalui mikroskop
cahaya. (b) Kariotipe kucing jantan, sebagaimana diperoleh dengan penataan kembali potongan kromosom secara individu dari cetakan fotografi kromosom (a).
Untuk menjelaskan kariotipe secara lebih lengkap, kromosom seringkali dikelompokkan menurut apakah sentromer berada pada satu ujungnya (akrosentrik), lebih dekat ke satu ujung daripada ujung lainnya (sub-metasentrik) atau di tengah (metasentrik). Pada buku ini, kita akan mengikuti kebiasaan praktis dalam menggunakan metasentrik untuk mencakup baik metasentrik maupun sub-metasentrik. Tangan pendek dari tiap kromosom diberi simbul p (petite = small = kecil), dan tangan panjang diberi simbul q. (Jika sentromer berada di tengah kromosom, simbul dari tangan yang mana yang disebut p adalah bersifat bebas (arbitrary), tetapi disetujui oleh konvensi internasional, untuk kromosom akrosentrik, misalnya autosom sapi, tidak perlu membedakan antar tangan, dan tidak juga p atau q yang digunakan.) Penjelasan ringkas tentang kariotipe dari hewan domestik disajikan pada Tabel 1.1. Kariotipe burung berbeda dengan kariotipe mamalian, karena selain beberapa autosom berukuran besar, mereka memiliki banyak autosom berukuran sangat kecil yang dinamakan mikrokromosom.
Tabel 1.1. Penjelasan ringkas mengenai karyotype dari beberapa mamalia domestik
Pasangan autosom Spesie Jumlah
diploid total
(2n= ) Metacentrics Acrocentrics
Kucing, Felis catus 38 16 2 Anjing, Canis familiaris 78 0 38 Babi, Sus scrofa domesticus 38 12 6 Kambing, Capra hircus 60 0 29
Domba, Ovis aries 54 3 23
Sapi, Bos taurus 60 0 29
Kuda, Equus caballus 64 13 18 Keledai, Equus asinus 62 24 6 Onta, Camelus dromedarius 74 10 62 Alpaca, Lama pacos
Llama, Lama glama
74 16 20 Kelinci, Oryctolagus cuniculus 44 34 8
Pemitaan
Ketika kariotipe pertama kali ditemukan, pasangan individu kromosom dapat diidentifikasi hanya berdasarkan ukuran dan bentuknya.
Sejak itu, berbagai metode pewarnaan kromosom telah dikembangkan, yang memunculkan wilayah terang dan gelap yang dinamakan pita. Tipe utama pita tersebut secara luas diklasifikasikan sebagai G, Q, R, C, T, dan N.
Sebagai contoh pemitaan, pita-G sapi diilustrasikan pada Gambar 1.2. Karena posisi, lebar, dan jumlah pita biasanya berbeda untuk setiap pasang kromosom, tiap pasangan kromosom dapat diidentifikasi oleh pola pitanya. Dengan mempelajari banyak sel yang diperlakukan dengan cara yang sama, dimungkinkan menggambar idiogram, yang merupakan pencerminan dari pola pita yang khas untuk tiap pasang kromosom. Pita pita tersebut secara unik diidentifikasi menurut suatu konvensi yang dikenal sebagai the
International System for Cytogenetic Nomenclature of Domestic Animals
(ISCNDA). Tiap tangan dibagi menjadi sejumlah kecil wilayah yang diberi nomor secara berurutan mulai dari sentromer. Kemudian, pada tiap wilayah, pita-pita tersebut diberi nomor secara berurutan mulai yang terdekat dengan sentromer. Misalnya, pita ke dua pada wilayah ke tiga dari kromosom 1 pada sapi diberi simbul 132, sedangkan pita ke dua pada wilayah ke empat dari tangan panjang kromosom X adalah Xq42. Idiogram ISCNDA untuk sapi diilustrasikan pada Gambar 1.3. Kariotipe berpita dari spesies domestik diilustrasikan pada Lampiran 1.1.
X Y
Gambar 1.3. Idiogram sapi standar, yang menunjukkan baik pita-G (kiri) maupun
pita-R (kanan).
Meiosis dan Mitosis
Selama beribu-ribu tahun, manusia telah menemukan dua fenomena yang berkaitan dengan penentuan jenis kelamin pada hewan: pertama, adanya keragaman pada jumlah jenis kelamin di antara anak-anak yang dilahirkan dari pasangan-pasangan orang tua; dan kedua, disamping keragaman ini, secara keseluruhan jumlah jenis kelamin jantan dan betina kira-kira sama.
Seperti telah disebutkan di atas, perbedaan kromosom kelamin pada dua jenis kelamin merupakan kunci dalam penentuan jenis kelamin. Alasan mengapa individu XX adalah betina dan XY adalah jantan juga akan diterangkan pada Bab 4. Untuk saat ini, kita hanya akan bertanya mengapa
terdapat keragaman jumlah individu XX dan XY pada anak-anak yang dilahirkan dari pasangan-pasangan orang tua, dan mengapa jumlah setiap jenis kelamin secara keseluruhan kurang lebih sama? Jawaban tersebut dapat diterangkan dalam proses pembentukan gamet.
Meiosis
Meiosis merupakan proses pembentukan gamet dimana gamet jantan (sel-sel sperma) dibentuk di dalam testes hewan jantan, dan gamet betina (sel-sel telur) dibentuk di dalam ovarium hewan betina. Hasil utama meiosis adalah bahwa setiap sperma dan setiap telur hanya mengandung satu dari setiap pasang kromosom. Dengan mengandung separuh dari jumlah kromosom diploid, gamet dikatakan sebagai haploid. Penyatuan satu sperma dengan satu telur pada saat pembuahan/fertilisasi menghasilkan zygot dengan jumlah normal kromosom diploid.
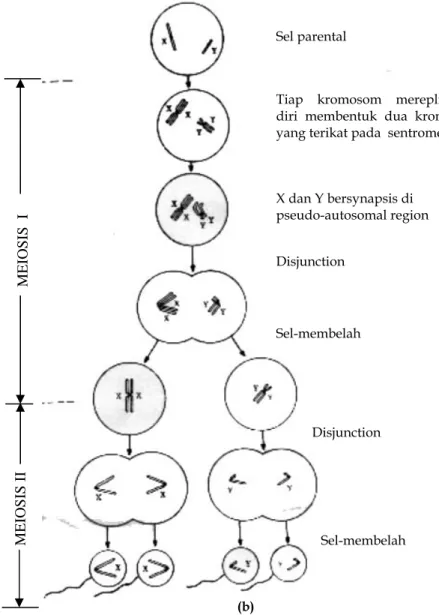
Proses meiosis berawal dari satu sel normal yang mengandung satu set kromosom diploid. Untuk menerangkan secara lebih mudah, kita akan mempertimbangkan apa yang terjadi pada satu pasang kromosom (yaitu kromosom kelamin) pada satu jenis kelamin (yaitu betina), seperti diilustrasikan pada Gambar 1.4a. Untuk membedakan dua kromosom kelamin X pada hewan betina, kita akan menyebutnya sebagai Xp (paternal: yang berasal dari pejantan) dan Xm (maternal: yang berasal dari induk).
Meiosis terjadi dalam dua tahap. Meiosis I mulai dengan proses penggandaan pada setiap kromosom, yang menghasilkan dua kromatid identik yang saling terpaut pada sentromer. Kemudian kromosom homolog, dalam hal ini Xp dan Xm, berdekatan satu sama lain secara vertikal di tengah sel, dalam suatu proses yang dikenal sebagai berpasangan atau sinapsis. Ini difasilitasi oleh struktur protein yang disebut synaptonemal complex, yang menyandingkan dua homolog tersebut secara bersama. Karena setiap kromosom telah mengalami penggandaan menjadi dua kromatid, maka terdapat empat kromatid yang berjajar satu dengan lainnya di dalam sel; dua kromatid Xp dan dua kromatid Xm. Kedua kromatid Xp masih saling terpaut pada sentromernya, demikian juga kedua kromatid Xm. Pada tahap ini, suatu proses yang disebut rekombinasi atau crossing-over terjadi, dimana kromatid homolog masing-masing pecah pada tempat yang sama, dan dalam proses penggabungan kembali, terjadi pertukaran segmen. Ini menghasilkan struktur yang nampak seperti tanda silang yang dikenal sebagai kiasmata. Untuk menyederhanakan diskusi, kita akan melanjutkan penggunaan kromatid-kromatid tersebut sebagai Xp atau Xm, dengan menyadari bahwa, sebagai akibat terjadinya pindah silang, setiap kromatid mungkin membawa segmen-segmen dari kromatid Xp dan Xm. (Diskusi lengkap mengenai implikasi genetika terjadinya pindah silang, disajikan kemudian pada Bab ini.) Pada tahap berikutnya dari meiosis I, dua
sentromer tersebut ditarik ke ujung-ujung yang berlawanan atau ke arah kutub dari sel, sehingga dua kromatid Xp bergerak ke satu kutub dan dua kromatid Xm bergerak ke kutub satunya lagi. Karena proses ini melibatkan terpisahnya dua kromatid yang tadinya selalu berpasangan, proses ini disebut sebagai disjungsi. Pada tahap akhir meiosis I, sel membelah menjadi dua sel anak; satu sel anak mengandung dua kromatid Xp, yang masih saling terpaut pada sentromer, dan sel anak lainnya mengandung dua kromatid Xm, yang juga masih saling terpaut pada sentromernya.
Mengikuti disjungsi pada hewan betina, hanya satu sel anak yang terus berfungsi secara normal; sedangkan sel anak lainnya mengalami degenerasi menjadi suatu struktur yang berwarna gelap dan tidak aktif yang dikenal sebagai tubuh polar pertama (first polar body). Kejadian tersebut, yaitu penentuan satu di antara kedua sel anak yang terus dapat berfungsi, merupakan suatu peluang saja. Oleh karena itu, terdapat peluang yang sama besarnya antara kedua kromatid Xp atau kedua kromatid Xm untuk membentuk sel anak yang dapat terus berfungsi secara normal. (Pada Gambar 1.4a., telah terjadi bahwa kromatid Xp terus dapat bertahan.)
Pada Meiosis II, dua kromatid yang berada dalam sel normal saling menjauh (disjoin) dan selanjutnya sel tersebut membelah menjadi dua sel anak, dimana setiap sel anak mengandung satu kromatid yang disebut kromosom. Sekali lagi, hanya satu dari dua sel anak tersebut yang dapat terus berfungsi; sel anak yang lain mengalami degenerasi menjadi tubuh polar ke dua (second polar body). Dan sekali lagi, ini juga masalah peluang, yang mana dari kedua sel anak tersebut akan terus berfungsi dan yang mana akan menjadi tubuh polar ke dua.
Jelaslah bahwa pada hewan betina, dari setiap sel yang mengalami meiosis, hanya satu gamet yang dapat terus berfungsi. Juga jelas ditunjukkan bahwa tanpa melihat sel anak yang dapat terus berfungsi, semua gamet yang dihasilkan oleh hewan betina adalah sama, dimana tiap gamet mengandung satu kromosom X. Untuk alasan ini, hewan betina juga disebut hewan berjenis kelamin homogamet.
Pada hewan jantan, proses meiosis sama seperti yang diterangkan diatas: disjungsi yang kemudian diikuti dengan pembelahan sel dalam meiosis I, dan dalam meiosis II (Gambar 1.4b). Akan tetapi, ada dua perbedaan penting. Perbedaan pertama adalah bahwa kromosom X dan Y hanya mempunyai sedikit segmen homolog pada ujung dari satu tangannya (dinamakan pseudo-autosomal region) dimana sinapsis terjadi; untuk sisa panjangnya, tangan tersebut tidak digabung bersama. Walaupun aturan yang tidak biasa ini, disjungsi berikutnya berjalan normal, dan menghasilkan dua sel anak yang dapat berfungsi setelah meiosis I berakhir: satu sel anak mengandung dua kromatid X yang masih terpaut pada sentromernya, dan satu sel anak lainnya mengandung dua kromatid Y yang juga masih terpaut pada sentromernya. Perbedaan ke dua antara proses
meiosis pada hewan betina dan jantan adalah bahwa tubuh polar tidak dibentuk pada hewan jantan. Sebagai gantinya, kedua sel anak yang dibentuk pada akhir meiosis I mengalami pembelahan sel pada meiosis II, yang menghasilkan empat gamet (sperma) yang semuanya dapat berfungsi normal, dua di antaranya masing-masing mengandung satu kromosom X dan dua lainnya masing-masing mengandung satu kromosom Y. Karena hewan jantan menghasilkan dua macam gamet yang berbeda, hewan ini juga disebut hewan berjenis kelamin heterogamet.
Sel parental
(a)
Tiap kromosom mereplikasi-diri membentuk dua kromatid yang terikat pada sentromer
Disjunction Chiasmata tampak Polar body prtm Sel-membelah Kromosom homolog bersynapsis Sel-membelah Disjunction
Polar body kedua
S OS I MEI II S OS I MEI I
(b)
Gambar 1.4. Meiosis pada betina (a) dan pada jantan (b), yang diilustrasikan dalam
hal kromosom kelamin. Dengan pengecualian pemasangan tak biasa pada jantan, proses yang persis sama terjadi untuk semua pasangan autosom.
Setelah proses pembentukan gamet, tahap berikutnya adalah pembuahan (fertilisasi) yang hasilnya, pada umumnya, juga merupakan faktor peluang.
MEIOSIS II
MEIOSIS I
Sel parental
Tiap kromosom mereplikasi-diri membentuk dua
yang terikat pada sentromer kromatid
Disjunction Sel-membelah X dan Y bersynapsis di pseudo-autosomal region Sel-membelah Disjunction
Peluang dan Keragaman
Karena semua gamet betina mengandung satu kromosom X, peluang satu gamet betina mengandung kromosom X adalah satu. Akan tetapi, hewan jantan menghasilkan gamet yang mengandung kromosom X dan gamet yang mengandung kromosom Y dengan jumlah sama. Oleh karena itu, ada peluang ½ bahwa sperma tertentu membawa kromosom X dan peluang yang sama bahwa sperma membawa kromosom Y. Ini berarti bahwa peluang untuk menghasilkan zigot XY adalah 1 X ½, yang sama dengan ½. Dengan cara yang sama, peluang untuk memperoleh zigot XX adalah 1 X ½ = ½. Kita dapat menunjukkan hasil pendugaan yang demikian dengan menggunakan suatu cara yang dinamakan checkerboard atau Punnett square, dimana proporsi pada bagian kolom (gamet jantan) dikalikan dengan proporsi pada bagian lajur (gamet betina), sehingga menghasilkan proporsi anak yang diharapkan pada bagian tengah checkerboard:
Gamet jantan ½ X ½ Y Gamet betina semua X ½ XX ½ XY
Sekarang kita telah mengetahui bagaimana meiosis dapat menghasilkan proporsi yang diharapkan dari setiap jenis kelamin, yang merupakan satu dari hasil observasi kita sendiri. Bagaimana kita dapat menerangkan hasil observasi kita yang ke dua, dengan mempertimbangkan adanya keragaman jumlah setiap jenis kelamin diantara anak-anak yang dihasilkan dari pasangan-pasangan orang tua yang berbeda? Ini hanyalah merupakan satu fakta yang memungkinkan kita dapat menerangkan adanya keragaman itu: tiap fertilisasi merupakan satu kejadian independen. Dengan cara ini kita dapat mengartikan bahwa tanpa mengetahui apakah sperma yang membawa kromosom X atau Y berhasil membuahi telur, hasil fertilisasi itu tidak berpengaruh pada fertilisasi berikutnya, walaupun fertilisasi tersebut terjadi pada saat yang hampir bersamaan. Sebagai contoh, pada hewan betina yang menghasilkan empat ovum, peluang bahwa ovum terakhir dibuahi oleh sperma yang membawa Y adalah ½ tanpa peduli jenis sperma yang mana yang membuahi ovum lainnya. Pada kenyataannya, urutan jenis kelamin, misalnya JJBJ, seharusnya sama dengan uirutan lainnya, misalnya BBBB.
Sekarang kita telah dapat memberikan penjelasan yang cukup untuk setiap observasi yang diterangkan terdahulu. Dalam hal ini, kita telah mendiskusikan kromosom, penurunan sederhana dan peluang, yang masing-masing merupakan dasar untuk dapat memahami ilmu genetika.
Untuk dapat melengkapi siklus reproduksi yang telah kita singgung ketika mendiskusikan meiosis, kita perlu membicarakannya, melalui proses yang dikenal sebagai mitosis, mulai dari zigot sampai dewasa yang dapat menghasilkan gametnya sendiri.
Mitosis
Perkembangan zigot mulai dari bersel satu sampai menjadi dewasa yang mempunyai banyak sel melibatkan suatu mekanisme dimana jumlah sel dapat berkembang biak secara cepat, yang dijamin bahwa setiap sel yang dihasilkan hanya mempunyai satu set kromosom yang persis sama seperti yang dipunyai zigot bersel satu. Mitosis merupakan mekanisme yang menerangkan kejadiannya. Untuk memudahkan pemahaman, kita hanya akan mempertimbangkan dua kromosom (yaitu kromosom kelamin) pada hewan jantan; tetapi proses yang terjadi persis sama untuk semua kromosom baik dari hewan jantan maupun betina.
Gambar 1.5. Mitosis pada jantan, yang diilustrasikan dalam hal kromosom kelamin.
Proses tersebut persis sama untuk semua kromosom, dan dalam semua sel dari tiap jenis kelamin.
Seperti ditunjukkan pada Gambar 1.5, mitosis berawal ketika tiap kromosom menggandakan diri/dupliaksi untuk membentuk dua kromatid yang masih terpaut pada sentromernya. Tiap kromosom yang digandakan bergerak menuju ke tengah sel tetapi tidak seperti pada proses meiosis, mengalami sinapsis dengan homolognya. Tahap ini, yang disebut metafase, merupakan satu-satunya tahap dimana kromosom dapat dilihat. Oleh karena itu, kariotipe merupakan kromosom pada tahap metafase. Setelah metafase, sentromer terbagi dua dan kedua kromatid memisah (disjoin), yang masing-masing menuju ke tiap kutub di dalam sel. Satu pembelahan terjadi di bagian tengah sel dan selanjutnya dua sel anak terbentuk, dimana tiap sel anak mengandung kedua kromosom X dan Y. Dengan cara ini, dua sel anak mempunyai satu set kromosom yang persis sama dengan yang dipunyai sel aslinya.
Pada kedua proses meiosis dan mitosis, kita telah mengetahui bahwa kromosom dapat menggandakan diri sendiri. Bagaimana kromosom tersebut dapat melakukannya? Untunglah, kita sekarang mempunyai cukup pengetahuan mengenai sifat biokimia kromosom untuk dapat menjawab pertanyaan ini dan menerangkan juga beberapa proses lainnya.
Biokimia Sifat Keturunan
Secara kimiawi, kromosom terdiri dari banyak asam deoksiribonukleat/deoxyribonucleic acid (DNA) dan sedikit protein yang disebut histon. Protein ini mempunyai fungsi secara struktural dan fungsi pengikatan pada saat molekul DNA membentuk informasi genetika yang diturunkan dari satu sel kepada sel anak hasil penggandaan, dan dari satu generasi ke generasi berikutnya, melalui meiosis.
DNA
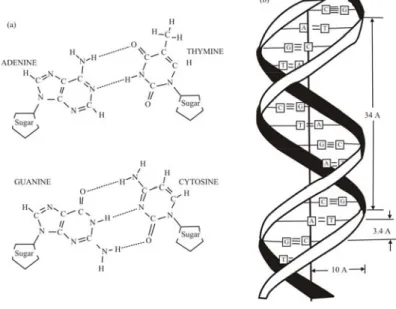
DNA terdiri atas dua rangkaian, yang masing-masing merupakan urutan nukleotida secara linier. Semua nukleotida DNA mengandung satu molekul gula (deoksiribosa) dan satu gugus fosfat. Komponen ke tiga, yaitu satu basa nitrogen, yang muncul dalam empat bentuk (adenin: A; guanin: G; timin: T; sitosin: C), yang menghasilkan mepat macam nukleotida, seperti diilustrasikan pada Gambar 1.6a.
Basa A dan G mempunyai struktur sama dan dinamakan purin; T dan C mempunyai struktur sama dan dinamakan pirimidin. Satu rangkaian nukleotida dihubungkan bersama dengan ikatan kovalen antara fosfat pada
karbon 5' dari satu nukleotida dan OH pada karbon 3' dari nukleotida di dekatnya, seperti ditunjukkan pada Gambar 1.6b. Itu berarti bahwa serangkaian DNA mempunyai 5'-fosfat pada satu ujungnya (dinamakan ujung 5') dan 3'-OH pada ujung lainnya (dinamakan ujung 3').
Gambar 1.6. (a) Struktur kimia dari empat nukleotida yang merupakan pembentuk
DNA, (b) Struktur dasar serangkaian DNA.
Dua rangkaian yang membentuk satu molekul DNA dihubungkan oleh ikatan hidrogen yang sangat spesifik antara purin dan pirimidin (A dengan T, dan G dengan C; Gambar 1.7a), yang menghasilkan pasang basa A:T dan G:C. Karena A hanya berikatan dengan T, dan G hanya dengan C, satu rangkaian DNA bersifat komplemen dengan rangkaian lainnya; urutan basa pada salah satu rangkaian DNA dapat diduga berdasarkan urutan basa dari rangkaian komplemennya. Konsekuensi lebih jauh dari pengaturan pasangan semacam ini adalah bahwa dua rangkaian tersebut tersusun bersama dalam bentuk heliks. Karena melibatkan dua rangkaian, susunan rangkaian yang demikian disebut heliks ganda (double-helix) (Gambar 1.7b). Panjang dari sepotong DNA yang pendek biasanya diukur berdasarkan jumlah pasang basa (pb). Potongan yang lebih panjang diukur berdasarkan kilobasa ( 1kb = 1.000 basa) atau bahkan megabasa (1 Mb = 1.000 kb).
Gambar 1.7. (a) Dua tipe pasangan basa pyrimidin:purin yang dibentuk oleh ikatan
hidrogen antara dua rangkaian DNA. (b) Heliks ganda DNA. Dua pita yang merepresentasikan ‘backbone’ gula-fosfat. Struktur tersebut berulang setiap 10 pasang basa.
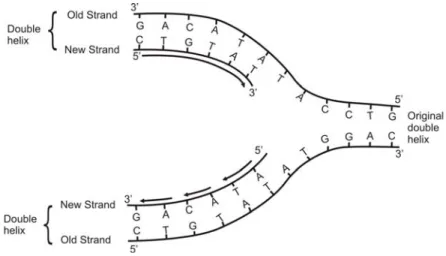
Aspek yang paling penting dari struktur molekul DNA adalah bahwa model ini diharapkan dapat menerangkan mekanisme replikasi. Apabila susunan heliks ganda mulai terurai yang diawali dari satu ujung molekul DNA sehingga dua rangkaian penyusunnya saling memisah diri, maka nukleotida-nukleotida yang sesuai dan telah tersedia dalam sel akan diikatkan dengan basa-basa pada setiap rangkaian yang tidak sedang berpasangan, yang selanjutnya membentuk satu rangkaian baru dan merupakan rangkaian komplemen bagi tiap-tiap rangkaian aslinya. Jadi, pada saat pemisahan dua rangkaian terus berlanjut (Gambar 1.8), dua heliks ganda akan dihasilkan dari satu heliks ganda aslinya; DNA telah direplikasi. Pembentukan tiap rangkaian baru oleh penambahan nukleotida diselesaikan dengan bantuan enzim DNA polimerase. Akan tetapi, enzim ini dapat menambah nukleotida hanya pada ujung 3´ dari rangkaian yang sedang memanjang, yang berarti bahwa replikasi hanya dapat terjadi dengan arah 5´ ke 3´. Konsekuensinya, satu rangkaian baru (Gambar 1.8 atas) disintesis secara berurutan, sedangkan rangkaian lainnya (Gambar 1.8. bawah) dibentuk melalui potong kecil-kecil (dinamakan Okazaki fragments) yang masing-masing disintesis dengan arah 5´ ke 3´. Okazaki fragments digabung bersama secara berurutan oleh enzim lainnya yang disebut DNA ligase. Kemampuan kedua enzim ini untuk menunjukkan fungsinya telah digunakan secara baik dalam biologi molekuler, seperti dijelaskan pada Bab 2.
Gambar 1.8. Replikasi DNA.
Sekarang dengan pengetahuan tersebut di atas, pengetahuan kita mengenai struktur molekul DNA dapat dikaitkan dengan struktur
kromosom seperti yang terlihat melalui mikroskop. Panjang total DNA pada sel mamalia adalah 1,74 meter, yang merupakan 7.000 kali lebih panjang dari total panjang kromosom metafase yang diamati melalui mikroskop! Oleh karena itu, jelaslah bahwa kromosom terdiri dari DNA yang tersusun seperti benang kusut yang padat dan kuat. Ini menimbulkan pertanyaan mengenai bagaimana dua rangkaian yang membentuk molekul DNA saling memisah setiap kali kromosom mengalami replikasi diri sendiri. Sudah barang tentu, protein histon dilibatkan dalam proses replikasi kromosom, tetapi mekanisme yang sebenarnya belum diketahui. Struktur DNA merupakan kunci untuk pemahaman mengenai suatu cara bagaimana informasi genetika disimpan dalam kromosom, dan dipindahkan ke sel dengan suatu cara sehingga menghasilkan suatu pengaruh tertentu. Pada kenyataannya, urutan basa dalam satu molekul DNA mempunyai arti spesifik yang dicatat dalam bentuk kode.
Kode genetika
Protein merupakan gugusan kimia yang mempunyai berbagai macam peranan spesifik pada berbagai organisme hidup. Beberapa dilibatkan dalam proses pengangkutan (misalnya hemoglobin), proses pendukung (misalnya kolagen), atau kekebalan (misalnya antibodi); beberapa merupakan enzim yang mengkatalisis reaksi-reaksi biokimia yang terjadi dalam sel hidup (misalnya alcohol dehidrogenase). Beberapa berupa hormon (misalnya hormon pertumbuhan); beberapa berupa reseptor untuk hormon (misalnya estrogen reseptor). Beberapa mengontrol aliran molekul atau ion keluar-masuk sel (misalnya calcium release channel). Selain itu, produk-produk komersial yang umumnya diperoleh dari hewan, baik yang hampir semuanya terdiri dari protein, misalnya daging dan wul, atau yang mempunyai protein sebagai komponen penting, misalnya susu dan telur.
Protein terdiri dari satu atau lebih polipeptida, dimana tiap-tiap polipeptida terdiri dari serangkaian asam amino. Ada 20 asam amino yang berbeda. Tiap polipeptida tertentu mempunyai suatu urutan spesifik asam amino yang menunjukkan satu set sifat kimia dan sifat fisika secara spesifik.
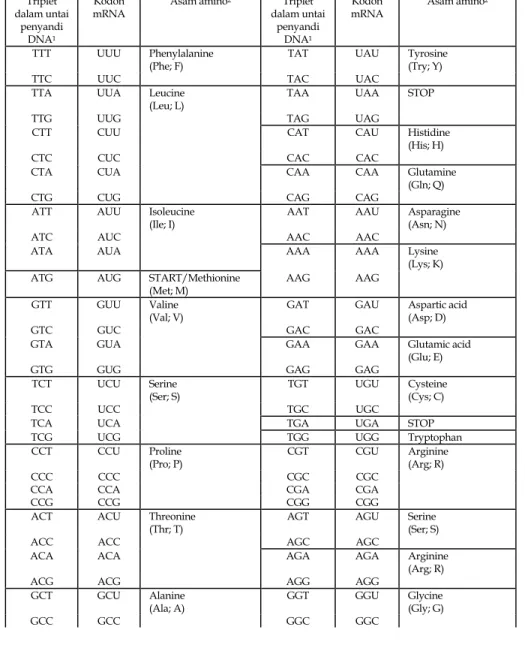
Informasi yang diperlukan untuk menghasilkan suatu urutan asam amino spesifik dibawa dalam bentuk kode dalam urutan basa pada satu segmen DNA. Kode ini, yang disebut kode genetika, ada sebagai triplet basa (Tabel 1.2). Dengan 4 X 4 X 4 = 64 kombinasi triplet dan hanya 20 asam amino, jelas ada beberapa pengulangan (redundancy); pada kenyataannya, dua basa pertama dari triplet tersebut seringkali cukup untuk mengidentifikasi satu asam amino tertentu, misalnya triplet GTT, GTC, GTA, dan GTG semua untuk valin. Tiga triplet (TAA, TAG, dan TGA) tidak memberikan kode untuk semua asam amino, dan dikenal sebagai triplet stop; triplet ini mengakibatkan berakhirnya suatu rantai polipeptida. Satu
triplet lainnya (ATG) beraksi sebagai tanda start untuk sintesis polipeptida. (Ini juga menyandi untuk metionin.) DNA antara dan termasuk triplet start dan triplet stop dinamakan open reading frame atau ORF, dimana urutan basa dibaca dalam triplet, yang masing-masing menyandi asam amino.
Tabel 1.2. Kode genetik
Triplet dalam untai
penyandi DNA1
Kodon
mRNA Asam amino
2 Triplet
dalam untai penyandi
DNA1
Kodon
mRNA Asam amino
2
TTT UUU Phenylalanine
(Phe; F) TAT UAU Tyrosine (Try; Y) TTC UUC TAC UAC
TTA UUA Leucine
(Leu; L) TAA UAA STOP TTG UUG TAG UAG
CTT CUU CAT CAU Histidine (His; H) CTC CUC CAC CAC
CTA CUA CAA CAA Glutamine (Gln; Q)
CTG CUG CAG CAG ATT AUU Isoleucine
(Ile; I) AAT AAU Asparagine (Asn; N) ATC AUC AAC AAC
ATA AUA AAA AAA Lysine (Lys; K) ATG AUG START/Methionine
(Met; M) AAG AAG GTT GUU Valine
(Val; V) GAT GAU Aspartic (Asp; D) acid GTC GUC GAC GAC
GTA GUA GAA GAA Glutamic acid (Glu; E)
GTG GUG GAG GAG TCT UCU Serine
(Ser; S) TGT UGU Cysteine (Cys; C) TCC UCC TGC UGC
TCA UCA TGA UGA STOP TCG UCG TGG UGG Tryptophan CCT CCU Proline
(Pro; P) CGT CGU Arginine (Arg; R) CCC CCC CGC CGC
CCA CCA CGA CGA CCG CCG CGG CGG ACT ACU Threonine
(Thr; T) AGT AGU Serine (Ser; S) ACC ACC AGC AGC
ACA ACA AGA AGA Arginine (Arg; R) ACG ACG AGG AGG GCT GCU Alanine
(Ala; A) GGT GGU Glycine (Gly; G) GCC GCC GGC GGC
GCA GCA GGA GGA GCG GCG GGG GGG
Keterangan:
1. Mengikuti konvensi biasa, sekuens DNA ditulis dalam DNA-equivalent of RNA, yang adalah sekuens pada untai (penyandi) non-template.
2. Simbol di dalam kurung setelah setiap nama asam amino adalah tiga huruf standard dan singkatan satu-huruf.
Dilengkapi dengan kode genetika seperti ini, kita sekarang dapat mengikuti proses yang dilibatkan dalam sintesis protein.
Sintesa protein
Seperti disajikan pada Gambar 1.9, sintesis polipeptida dimulai dengan bagian DNA yang mengurai, dan dua benang yang saling memisah. Runutan basa DNA pada satu dari benang tersebut (disebut benang tatakan/template) bertindak sebagai tatakan untuk sintesis asam nukleat yang berbeda (asam ribonukleat, RNA, dinamakan demikian karena nukleotidanya mengandung ribosa ketimbang deoksiribosa). Sintesis tersebut dikatalisis oleh enzim RNA polimerase, yang, seperti DNA polimerase, menambah nukleotida pada ujung 3' dari benang yang sedang dicetak, dengan kata lain RNA juga disintesis pada arah 5' ke 3'. Tiga dari basa-basa RNA sama seperti DNA, dan basa ke empat, urasil (U), terbentuk sebagai ganti dari timin (T). Pembentukan benang komplemen dari RNA pada tatakan DNA dinamakan transkripsi (karena runutan basa dalam DNA telah ditranskrip ke RNA).
Gambar 1.9. Sitesis polipeptida pada eukaryot, dengan cara transkripsi dan translasi.
Sebelum tahap berikutnya dapat dimulai, RNA tersebut harus pindah dari inti sel, tempat kromosom berada, ke suatu struktur yang disebut ribosom di dalam sitoplasma, tempat polypeptida disintesis. (Sudah barang tentu, tahap ini hanya diperlukan dalam organisme yang selnya mempunyai inti sel, yaitu eukariot. Pada prokariot, yang tidak mempunyai inti sel, ribosom tertempel secara langsung ke RNA bahkan sebelum transkripsi selesai.) Karena RNA tersebut di atas membawa sandi antara DNA dan protein, RNA ini disebut messenger RNA atau mRNA. Tripletnya dinamakan kodon (disajikan pada Tabel 1.2).
Seperti juga disajikan pada Gambar 1.9, tahap ke dua dari sintesis protein melibatkan RNA tipe ke dua yang dikenal sebagai transfer RNA atau tRNA. Untuk setiap dari 20 asam amino, ada satu atau lebih tRNA spesifik yang mengikat asam amino yang sesuai dan yang mempunyai triplet nukleotida (triplet ini dinamakan antikodon) yang bersifat komplemen terhadap kodon mRNA. Karena bersifat komplemen, antikodon tRNA berpasangan dengan kodon mRNA yang sesuai, yang membawa asam amino yang tepat ke posisinya pada rantai polipeptida. Tahap ke dua dan tahap akhir dari sintesis protein ini disebut translasi, karena runutan basa tersebut telah diterjemahkan (melalui sandi genetika) ke runutan asam amino.
Apakah Gen Itu?
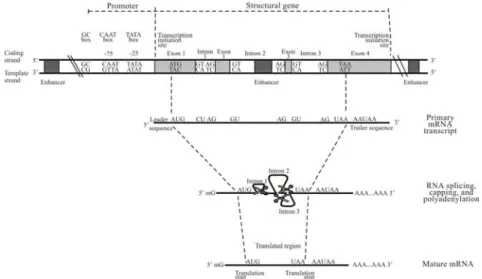
Dari penjelasan di atas, jelaslah bahwa bagian tertentu dari DNA menyandi polipeptida tertentu, yang dihasilkan melalui mRNA. Bagian DNA termasuk semua nukleotida yang ditranskrip menjadi mRNA disebut gen struktur (Gambar 1.10).
Karena yang sebenarnya diterjemahkan menjadi polipeptida itu adalah runutan mRNA, dan karena translasi dimulai pada ujung 5' mRNA, ada kesepakatan bahwa runutan basa dari gen ditulis sebagai DNA yang ekuivalen dengan runutan mRNA, pada arah 5' ke 3'. Mengingat bahwa runutan basa dari mRNA bersifat komplemen terhadap runutan pada benang DNA yang ditranskrip, itu berarti bahwa benang DNA yang runutannya ekuivalen dengan runutan mRNA bukanlah benang tatakan, tetapi benang yang satunya lagi. Karena alasan ini, benang bukan tatakan dinamakan benang penyandi (coding) atau sense, sedangkan benang tatakan disebut benang anti penyandi (anticoding) atau antisense.
Dengan demikian gen struktur mencakup semua nukleotida yang secara pasti diterjemahkan menjadi polipeptida, yaitu semuanya mulai nukleotida pertama dari triplet pemulai sampai nukleotida dari triplet
pemberhenti. Tetapi itu mencakup lebih dari ini. Kenyataannya, transkripsi dimulai sebelum triplet pemulai dan berakhir setelah triplet pemberhenti. Ini berarti bahwa mRNA mempunyai untranslated region pada tiap ujungnya. Daerah yang terjadi sebelum triplet pemulai (runutan pemandu/leader sequence) adalah tempat ribosom melekat. Daerah tak-terjemahkan pada ujung lainnya (runutan penjejak/trailer sequence) diperlukan untuk memproses mRNA. Definisi gen struktur mencakup bagian-bagian DNA yang berkaitan dengan daerah-daerah tersebut. Jadi, nukleotida pertama dari gen struktur adalah nukleotida pada suatu titik tempat transkripsi dimulai, yaitu pada tempat mulai transkripsi atau transcription initiation site. Akhir dari gen struktur disebut tempat akhir transkripsi atau transcription termination site. Melalui kesepakatan, nukleotida dari gen struktur dinomori dari awal tempat mulai transkripsi, dan basa-basa yang mendahului tempat tersebut dinomori negatif, yaitu –1, –2, dan seterusnya.
Gambar 1.10. Karakteristik penting dari gen dan mRNA-nya.
Daerah pendahulu yang dekat, yaitu usptream dari, tempat mulai transkripsi merupakan daerah penting, karena itu merupakan tempat melekatnya RNA polimerase sebelum dimulainya transkripsi. Daerah ini disebut promoter. Ini mengandung runutan spesifik yang sangat konservatif, yaitu runutan basa yang sama atau sangat mirip yang terdapat pada sebagian besar gen. Dengan mempelajari runutan pada daerah konservatif ini pada banyak gen dari banyak spesies, runutan konsensus (consensus sequence) dapat dihasilkan, yang terdiri atas basa yang paling sering pada
tiap posisi. Jenis pertama runutan semacam itu pada daerah promoter dari gen eukariot adalah TATAAAA (disebut kotak TATA), yang terletak kira-kira 25 basa upstream, yaitu pada sekitar posisi –25 . Lebih jauh kedepan adalah runutan GGCCAATCT (kotak CAAT) pada sekitar posisi –75, dan GGGCGG (kotak GC) pada sekitar –90. Kotak-kotak tersebut merupakan tempat untuk pengenalan dan pengikatan protein pengatur (regulatory
protein) yang disebut faktor-faktor transkripsi (transcription factor), yang
memungkinkan RNA polimerase ditempatkan secara tepat untuk memulai transkripsi. Dengan cara ini, mereka melakukan pengontrolan terhadap pentranskripsian. Prokariot juga mempunyai runutan konservatif pada daerah promoternya, yaitu TATAAT dan TTGACA.
Akhir dari pentranskripsian masih kurang dimengerti dengan baik daripada awalnya. Akan tetapi, itu diketahui bahwa pentranskripsian sebenarnya berkembang di atas apa yang kita namakan tempat akhir tersebut. Kemudian enzim tak teridentifikasi memotong transkrip (hasil transkripsi) pada tempat akhir. Tidak terdapat runutan konservatif yang berkaitan dengan tempat ini, tetapi terdapat daerah yang sangat konservatif dengan runutan konsensus AATAAA (AAUAAA pada mRNA) yang terletak 10--30 basa sebelum tempat akhir, yang tampaknya merupakan tempat pengenalan pada mRNA untuk faktor yang mengontrol pemotongan. Gen split
Sampai tahun 1977, dioerkirakan bahwa gen struktur berukuran cukup panjang untuk menyandi mRNA yang dilibatkan dalam pentranslasian. Tetapi kini diketahui bahwa pada eukariot, sebagian besar gen struktur berukuran lebih panjang dari itu--mereka mengandung bagian-bagian yang tidak diwujudkan dalam mRNA pada saat mRNA tersebut mengalami translasi. Bagian tersebut dinamakan intron (karena mereka awalnya dinamakan intragenic region). Bagian yang diwujudkan dalam versi akhir dari mRNA (mature/matang) dinamakan exon (expressed regions). Gen yang mengandung intron dikatakan merupakan split. mRNA muasal (primary) merupakan cetakan dari seluruh gen struktur, yaitu exon dan
intron. Sebelum mRNA pindah dari inti sel ke sitoplasma, intron dihilangkan
dan exon disambung bersama (dalam suatu proses yang disebut penyambungan RNA/RNA splicing), sehingga mRNA matang tersebut hanya terdiri dari exon saja. Pengenalan dan penghilangan intron dibantu oleh kenyataan bahwa pada tiap intron, dua basa pertama pada ujung 5' dan dua basa terakhir pada ujung 3' sangat konservatif--basa yang sama (masing-masing GU dan AG dalam mRNA) terdapat pada tempat-tempat tersebut pada sebagian besar, jika tidak semua, intron-intron. Mengingat bahwa runutan biasanya diekspresikan berdasarkan benang DNA penyandi, ini disebut aturan GT-AG. Runutan basa yang terdekat pada kedua tempat
runutan yang sangat konservatif tersebut, yaitu dekat perbatasan antara
intron dan exon, juga sangat konservatif, tetapi dengan derajat yang lebih
rendah.
Perbatasan tersebut dinamakan tempat penyambungan (splice site), dengan tempat pada ujung 5' disebut tempat donor (donor site) dan tempat pada ujung 3' disebut tempat penerima (acceptor site).
Gen struktur berukuran antara sekitar 1.000 basa (1kb) sampai lebih besar dari dua juta basa (2.000kb), dengan rataan sekitar 100.000 basa (100kb). Sebaliknya, jumlah asam amino dalam polipeptida berkisar dari sekitar 200 sampai sekitar 5.000, dengan rataan sekitar 330, yang berarti bahwa mRNA-matang berukuran antara sekitar 600 basa (0,6kb) sampai 15.000 basa (15kb), dengan rataan sekitar 1.000 basa (1kb). Jelas dari data tersebut bahwa exon hanya membentuk proporsi kecil dari gen struktur; kebanyakan DNA dalam gen struktur berupa intron. Keberadaan dari begitu banyak DNA 'non-fungsional’ tetap merupakan satu dari misteri biologi terbesar yang belum terpecahkan. Akan tetapi, kita sebaiknya tak perlu heran, jika solusi terhadap misteri tersebut adalah bahwa benar-benar mempunyai fungsi penting, yang hanya sedang menunggu untuk diketemukan. (Kita telah mempunyai beberapa petunjuk, yang diantaranya
intron cenderung berfungsi sebagai pengatur jarak antara unit-unit yang
berfungsi, yaitu banyak exon berkaitan dengan unit-unit berfngsi yang dapat dibuat dalam kombinasi berbeda untuk menghasilkan polipeptida berbeda. Tetapi ini bukan penjelasan yang memuaskan untuk jumlah DNA yang berada dalam introns.)
Disamping penghilangan intron, transkrip mRNA utama juga dimodifikasi dalam dua cara lain. Ujung (depan) 5'-nya dilindungi oleh penambahan bungkus-5' (5' cap), yang terdiri dari nukleotida guanin yang termetilasi. Pada ujung lainnya, ekor poli-A (poly-A tail) ditambahkan, yang terdiri dari nukleotida adenin yang bervariasi jumlahnya (biasanya 100--200). Karena bungkus dan ekor tersebut merupakan karakteristik yang penting dari mRNA-matang, tempat awal dan akhir pentranslasian dari gen kadang-kadang disebut tempat bungkus (cap site) dan tempat poli-A (poly-A site). Runutan (poly-A(poly-AU(poly-A(poly-A(poly-A yang terletak 10--30 basa di depan tempat poli-(poly-A disebut tanda poliadenilasi (polyadenylation signal).
Jenis gen yang diterangkan di atas merupakan yang paling umum. Akan tetapi, penting disadari bahwa ada bagian DNA yang fungsi utamanya adalah produksi, melalui transkripsi, tRNA atau ribosomal RNA (rRNA, yang merupakan penyusun utama ribosom). Itu juga dinamakan gen. Karena sejumlah besar rRNA diperlukan untuk pembentukan ribosom yang cukup untuk memenuhi setiap kebutuhan sel untuk translasi, ada ratusan gen untuk rRNA di dalam genom. Mereka terbentuk dalam beberapa kelompok dari gen rRNA yang berulang secara tandem. Setiap kelompok menghasilkan nukleolus, yang merupakan struktur terpisah yang
ditemukan di dalam nukleus (lihat Gambar1.13), dan yang terutama terdiri dari RNA ribosom plus enzim-enzim yang diperlukan untuk pembentukan ribosom. Sekelompok gen rRNA dinamakan nucleolar organizer region (NOR).
Kita dapat menggabungkan semua tipe gen yang berbeda tersebut menjadi definisi tunggal dengan mengatakan bahwa gen merupakan serangkaian DNA yang menghasilkan molekul RNA yang berfungsi.
Regulasi Gen
Tentunya akan ada kekacaubalauan jika semua gen ditranskrip dalam semua sel sepanjang waktu. Kenyataannya, hanya sebagaian kecil gen ditranskrip pada satu waktu dalam setiap satu sel. Dari saat fertilisasi sampai kematian, perkembangan dari tiap organisme hidup ditentukan oleh gen yang dinyalakan atau dimatikan pada saat-saat yang tepat dalam sel-sel yang tepat. Penyetelan ini diatur oleh bermacam-macam protein (kadang-kadang berkaitan dengan hormon steroid) yang melekat ke, atau dilepaskan dari, runutan DNA spesifik yang seringkali sangat konservatif. Kita telah menyebutkan tiga runutan seperti itu di dalam promoter. Runutan yang mempunyai fungsi serupa tapi yang tidak terletak di dalam promoter dinamakan enhancer. Ini terletak di depan (upstream), di belakang (downstream) dan kadang-kadang bahkan di dalam gen struktur (yaitu di dalam intron). Beberapa terletak dekat sekitar gen struktur, tetapi beberapa lainnya terletak cukup jauh (20kb atau bahkan lebih).
Protein-protein pengatur yang melakukan pengontrolan terhadap pentranskripsian melalui pengikatan promoter atau enhancer mempunyai satu atau lebih runutan asam amino yang serupa (dinamakan motif-motif asam amino) yang diberi nama agak eksotik. Sebagai contoh, zinc finger merupakan struktur yang muncul dari kejadian berulang sepasang molekul sistein yang dipisahkan oleh dua atau tiga asam amino lain, diikuti sekitar 10 asam amino kemudian oleh sepasang molekul histidin, juga dipisahkan oleh dua atau tiga asam amino lain. Berkombinasi dengan atom zinc, dua sistein tersebut bergandengan dengan dua histidin, dan asam amino yang mengintervensinya membentuk lekukan seperti jari (finger) yang berikatan dengan DNA. Contoh lainnya adalah leucine zipper, yang mengandung ulangan periodik dari leusin setiap asam amino ke tujuh, yang menimbulkan heliks dengan leusin disejajarkan sepanjang satu permukaan. Dua molekul yang tersebut siap bergabung bersama (dalam suatu cara zipper yang menciptakan dimer. Satu ujung dari dimer ini mengikat DNA.
Secara umum, itu merupakan pengikatan protein pengatur (kadang-kadang berkaitan dengan hormon steroid) ke promoter atau enhancer yang memungkinkan RNA polimerase melekat ke promoter dari suatu gen, dan yang oleh karenanya menimbulkan pengontrolan terhadap tahap pertama dalam pentranskripsian. Ini adalah cara penyalaan dan pematian gen.
Misalnya, perkembangan embrio tahap awal pertama dikontrol oleh sekelompok gen segmental (yang membagi embrio yang belum terbagi-bagi menjadi segmen-segmen) dan kemudian oleh sekelompok gen homeotik (yang masing-masing menentukan nasib perkembangan dari satu segmen, misalnya hindbrain atau spinal cord). Protein yang disandi oleh gen segmental dan homeotik mempunyai motif pengikatan DNA seperti zinc finger atau
leucine zippers. Banyak dari gen-gen tersebut juga mempunyai daerah
sepanjang 180pb yang sangat konservatif, yang disebut homeobox, yang menyandi motif pengikatan-DNA yang dinamakan homeodomain yang sangat konservatif pada seluruh eukariot. Jadi gen yang mengontrol perkembangan embrio tahap awal menyandi protein pengatur yang menyalakan dan mematikan gen dalam cara yang terkontrol.
Contoh spesifik dari regulasi gen kemudian dalam kehidupan disajikan oleh cara estrogen mengontrol pentranskripsian gen ovalbumin pada ayam. Molekul dari hormon tersebut masuk sel dan berikatan dengan protein yang disebut reseptor estrogen. Kompleks estrogen plus reseptor kemudian mengikat ke daerah yang kira-kira 250 basa upstream dari kotak TATA dari gen ovalbumin, yang menyalakannya.
Tidak mengherankan, kelompok-kelompok gen yang semuanya perlu dikontrol dalam cara yang sama mempunyai daerah serupa pada promoter-nya. Daerah ini dinamakan elemen respon. Misalnya, semua gen yang perlu diaktifkan oleh glukokortikoid mempunyai elemen respon glukokortikoid, tempat receptor glucocorticoid mengikat, setelah pengaktifan oleh glukokortikoid. Runutan konsensusnya adalah TGGTACAAATGTTCT.
Dari diskusi di atas, jelaslah bahwa runutan pada kedua sisi suatu gen juga sama pentingnya dengan gen itu sendiri. Kenyataannya, jika kata gen digunakan terhadap gen itu sendiri, nama itu sering diambil untuk memasukkan daerah promoter dan enhancer serta daerah di antaranya.
Mutasi
Kita telah melihat bagaimana DNA mereplikasi, dan bagaimana DNA membentuk protein. Walaupun proses yang dilibatkan sangat baik dan biasanya berlangsung sempurna, kesalahan terjadi dari waktu ke waktu. Banyak kesalahan tidak berdampak sama sekali, karena mereka diperbaiki oleh mekanisme perbaikan yang dimiliki sel. Akan tetapi, kesalahan yang tak diperbaiki dalam pereplikasian DNA menyebabkan perubahan pada DNA tersebut paling tidak pada satu di antara sel anakannya. Dan karena pereplikasian DNA biasanya berlangsung sempurna, DNA yang mengalami perubahan tersebut akan tetap diwariskan ke semua sel turunannya; sampai kesalahan berikutnya terjadi. Kesalahan yang tidak diperbaiki dalam pereplikasian DNA dinamakan mutasi.
Kita akan mulai dengan mempertimbangkan point mutation (juga disebut mutasi gen), yang melibatkan substitusi satu nukleotida dengan nukleotida lainnya, atau penambahan atau penghilangan satu atau beberapa nuikleotida. Tipe mutasi lainnya akan didiskusikan pada bab berikutnya.
Ada beberapa akibat mutasi gen yang mungkin berbeda. Pada satu ekstrim, substitusi basa dapat mengubah triplet fungsional menjadi triplet stop (disebut mutasi nonsense). Misalnya, TAT menyandi tirosin; tetapi jika T pada posisi ke tiga diganti dengan A, tripletnya menjadi (TAA) yang berarti stop (cek ini pada Tabel 1.2). Jika triplet stop baru tersebut terjadi sebelum triplet stop biasanya, polipeptida yang dihasilkan lebih pendek dari biasanya, dan oleh karena itu mungkin tidak fungsional. Jika substitusi basa mengubah triplet sehingga menyebabkan substitusi asam amino, itu dinamakan mutasi mis-sense. Misalnya, mensubstitusi A untuk T pada posisi ke tiga dari CAT (histidin) menyebabkan CAA (glutamin).
Pada ekstrim lainnya, banyak substitusi basa tidak mempunyai pengaruh pada urutan asam amino dari suatu produk gen, karena triplet mutan terjadi dengan hasil asam amino yang sama seperti triplet aslinya. Mutasi yang disebut mutasi silent ini merupakan konsekuensi langsung dari pengulangan dalam kode genetika. Sebagai contoh, mensubstitusi C untuk T pada posisi ke tiga dari CAT (histidin) menyebabkan CAC, yang masih menyandi histidin (cek ini pada Tabel 1.2).
Tipe mutasi gen lainnya melibatkan penghilangan atau penyisipan satu atau dua basa. Ini dinamakan mutasi frameshift, karena tiap triplet yang terjadi downstream dari tempat suatu mutasi digeser keluar dari open
reading frame aslinya. Terakhir, mutasi frameshift menyebabkan urutan asam
amino yang sangat berbeda downstream dari tempat mutasi. Misalnya, pertimbangkan kasus berikut (cek lagi pada Tabel 1.2):
Rangkaian penyandi asli TCCGAGTATCAGTCCCAG... Urutan asam amino Ser Glu Tyr Gln Ser Gln... Jika basa ke dua dihilangkan, kita mempunyai:
Rangkaian penyandi mutan TCGAGTATCAGTCCCAG... Urutan asam amino Ser Ser Ile Ser Pro...
Urutan asam amino mutan jelas sangat berbeda dengan urutan aslinya. Pada beberapa kasus, satu dari triplet ‘baru’ adalah triplet stop, yang menyebabkan terminasi prematur dari proses translasi. Apakah triplet stop baru diciptakan atau tidak, sangat mungkin bahwa polipeptida mutan akan berfungsi.
Jika mutasi terjadi di dalam sel selain mereka yang menimbulkan sel kelamin, maka yang demikian itu dinamakan mutasi somatik. Tahap
perkembangan dari individu ketika mutasi somatik muncul akan menentukan jumlah total sel yang mengandung DNA mutan atau DNA terubah; makin awal mutasi terjadi, makin besar jumlah sel yang dipengaruhi.
Sebaliknya, mutasi yang terjadi di dalam sel yang menimbulkan sel kelamin dikenal sebagai mutasi nutfah (germ-line mutation), yang barangkali menyebabkan pembentukan gamet yang mengandung DNA terubah. Jika gamet ini berhasil dalam fertilisasi, mutasi akan diwariskan ke keturunan yang dihasilkan, dalam setiap sel yang itu semua akan direproduksi.
Gen, Alel, dan Lokus
Bentuk berbeda dari bagian DNA yang berada pada tempat tertentu di kromosom dinamakan alel. Tempat atau posisi tertentu dari gen di kromosom dinamakan lokus (jamak, loki). Kata 'gen' umumnya digunakan dalam hal alel atau lokus. Jika digunakan pada cara ini, arti yang cocok untuk kata tersebut biasanya mudah dimengerti dari konteksnya.
Jika keturunan berasal dari penyatuan sperma dengan DNA normal dan sel telur dengan DNA terubah atau DNA mutan pada satu dari kromosomnya, maka keturunan itu akan mempunyai satu kromosom normal dan satu kromosom mutan, yang membentuk pasangan homolog yang sesuai. Secara lebih spesifik, akan ada satu alel normal dan satu alel mutan pada lokus yang relevan. Kita akan menandai dua alel ini dengan lambang B dan b masing-masing. Hewan dengan dua alel yang berbeda pada lokus tertentu dikatakan bersifat heterozigot pada lokus itu. Sebaliknya, jika hewan mempunyai dua kopi alel yang sama maka hewan itu bersifat homozigot pada lokus tersebut.
Meskipun kedua hewan dapat mempunyai maksimal hanya dua alel yang berbeda pada suatu lokus, jumlah alel yang berbeda dalam populasi hewan dapat jauh lebih besar dari dua. Jika lebih dari dua alel berada dalam populasi pada lokus tertentu, maka lokus itu dikatakan mempunyai alel ganda (multiple-allele).
Pewarisan Sederhana atau Mendel
Penurunan gen dari satu generasi ke generasi berikutnya dinamakan pewarisan (inheritance). Satu dari terobosan utama dalam ilmu pengetahuan adalah kesadaran bahwa hasil pewarisan dapat diprediksi. Orang pertama yang memformulasikan prediksi itu adalah pendeta
Augustinian, Gregor Mendel, yang melakukan penelitiannya di Brün (sekarang Brno di Republik Czech) pada pertengahan abad lalu.
Lokus tunggal
Sebagai contoh, anggap perkawinan dari heterozigot (Bb) dengan homozigot (bb). Ini persis sama dengan situasi pada kromosom kelamin. Sebagai akibatnya, hasil perkawinan Bb X bb dapat diterangkan dengan cara yang sama seperti yang digunakan untuk pewarisan kelamin yang didiskusikan lebih awal, dengan bantuan checkerboard:
Gamet dari tetua heterosigous
½ B ½ b
Gamet dari semua b
tetua homosigous ½ Bb ½ bb
Hasil perkawinan Bb X bb diharapkan mempunyai proporsi yang sama antara keturunan Bb dan bb. Pemisahan alel pada lokus selama meiosis dinamakan segregasi, dan rasio dari jenis keturunan yang berbeda sebagai akibat dari perkawinan dari tetua tertentu dinamakan rasio segregasi. Untuk perkawinan Bb X bb, rasio segregasinya adalah ½ Bb : ½ bb, yang sering ditulis sebagai 1 Bb : 1 bb.
Checkerboard dapat digunakan untuk menduga hasil dari sembarang
perkawinan tertentu yang melibatkan lokus tunggal. Rasio segregasi yang diharapkan dari semua jenis perkawinan yang mungkin dengan asumsi lokus tunggal terdaftar pada Tabel 1.3.
Tabel 1.3. Rasio segregasi yang diharapkan pada keturunan yang timbul dari semua
tipe perkawinan yang mungkin dalam hubungannya dengan lokus autosom tunggal, sebagaimana diperoleh dari checkerboard
Tipe perkawinan Rasio segregasi
BB Bb bb
BB × BB 1 : 0 : 0
BB × bb 0 : 1 : 0
Bb × Bb 1 : 2 : 1
Bb × bb 0 : 1 : 1
bb × bb 0 : 0 : 1
Lebih dari satu lokus
Dalam ketiadaan bukti terhadap hal yang sebaliknya, itu dianggap bahwa segresasi pada satu lokus bersifat bebas dari segregasi pada lokus yang lain. Ini merupakan asumsi yang dibuat Mendel, dan itu benar pada berbagai situasi yang diamati pada hewan saat ini. Jika segregasi pada tiap lokus bersifat bebas dari segregasi pada lokus lain, maka peluang memperoleh gamet dengan alel tertentu (katakan B) pada lokus pertama dan alel tertentu (katakan d) pada lokus ke dua merupakan produk dari kemungkinan yang berkaitan dengan tiap alel secara bebas. Sebagai misal, jika individu bersifat heterozigot pada dua lokus (BbDd), maka ada empat tipe gamet yang mungkin, BD, Bd, bD, dan bd, yang akan dihasilkan dengan frekuensi sama. Hasil dari segregasi bebas pada dua lokus ditunjukkan pada
checkerboard:
Gamet dari satu tetua
¼ BD ¼ Bd ¼ bD ¼ bd
Gamet ¼ BD BBDD BBDd BbDD BbDd
dari ¼ Bd BBDd BBdd BbDd Bbdd
tetua ¼ bD BbDD BbDd bbDD bbDd
lainnya ¼ bd BbDd Bbdd bbDd bbdd
Menggabungkan semua sel pada checkerboard yang mempunyai keturunan identik, dan menyadari bahwa keturunan pada tiap sel terjadi dengan frekuensi ¼ x ¼ = 1/16, rasio segregasinya adalah:
1 BBDD : 2 BBDd : 1 BBdd : 2 BbDD: 4 BbDd : 2 Bbdd : 1 bbDD : 2 bbDd : 1 bbdd
Meskipun checkerboard menjadi agak besar, pada dasarnya itu dapat digunakan untuk menurunkan rasio segregasi yang diharapkan untuk semua tipe perkawinan yang melibatkan sembarang jumlah lokus yang mengalami segregasi secara bebas.
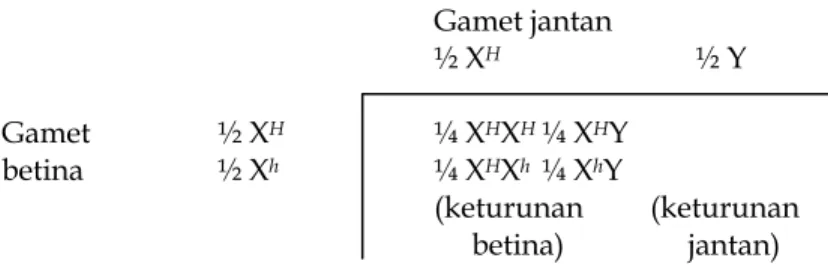
Pola pewarisan di atas tersebut mengilustrasikan pewarisan autosom sederhana karena pola tersebut menerangkan apa yang terjadi berkaitan dengan lokus pada autosom. Akan tetapi, beberapa lokus berada pada kromosom kelamin dan akibatnya mempunyai pola pewarisan yang berbeda. Lokus yang demikian dikatakan terpaut kelamin (sex-linked). Pola pewarisan lokus terpaut-X dapat diilustrasikan pada checkerboard:
Gamet jantan ½ XH ½ Y Gamet ½ XH ¼ XHXH ¼ XHY betina ½ Xh ¼ XHXh ¼ XhY (keturunan (keturunan betina) jantan)
dan diringkas pada Tabel 1.4. Sangat sedikit jumlah lokus yang diidentifikasi pada kromosom Y (terpaut-Y).
Tabel 1.4. Rasio segregasi yang diharapkan dari semua tipe perkawinan yang mungkin
dalam hubungannya dengan lokus terpaut-X, sebagaimana diperoleh dari checkerboard
Rasio segregasi
Diantara betina Diantara jantan
Tipe perkawinan XHXH XHXh XhXh XHY XhY XHXH × XHY 1 : 0 : 0 1 : 0 XHXh × XHY 1 : 1 : 0 1 : 1 XhXh × XHY 0 : 1 : 0 0 : 1 XHXH × XhY 0 : 1 : 0 1 : 0 XHXh × XhY 0 : 1 : 1 1 : 1 XhXh × XhY 0 : 0 : 1 0 : 1
Pada bagian awal seksi ini ditunjukkan bahwa kadang-kadang bukti yang baik diperoleh untuk menunjukkan bahwa segregasi pada dua lokus atau lebih tidaklah semuanya bersifat bebas. Kita sekarang akan menguji mengapa hal ini terjadi.
Sedikitnya ada ribuan gen berbeda, tetapi hanya ada sejumlah kecil kromosom. Oleh karena itu, sangatlah jelas tiap kromosom terdiri atas banyak gen berbeda, yang masing-masing mempunyai posisi (lokus) spesifik pada kromosom itu. Jika kromosom diwariskan sebagai unit kesatuan, maka untuk semua lokus pada kromosom tertentu, alel yang berada pada kromosom itu akan selalu bersegregasi bersama. Misalnya, anggap satu kromosom yang mengandung alel B pada suatu lokus dan alel D pada lokus lainnya, dan masing-masing homolognya mengandung alel b dan d. Jika kromosom bersegregasi sebagai unit kesatuan, maka hanya dua tipe gamet yang akan menghasilkan, yaitu BD dan bd, dengan frekuensi yang sama.
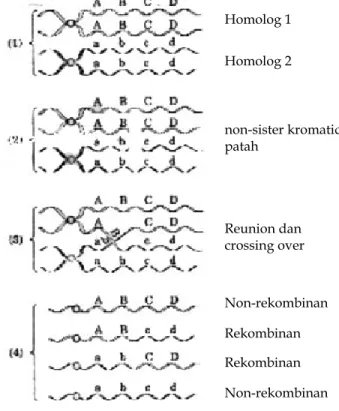
Pada prakteknya, kromosom tidak diwariskan sebagai unit kesatuan. Sebagai gantinya, seperti diterangkan lebih awal, rekombinasi atau pindah silang terjadi ketika kromosom homolog mengalami sinapsis selama tahap pertama meiosis. Selama sinapsis, pematahan dan penyambungan-kembali kromatid terjadi. Jika dua segmen dari kromatid yang patah menyambung kembali, kromatid itu masih akan diwariskan sebagai unit kesatuan. Akan tetapi, jika patahan terjadi pada posisi yang sama pada dua kromatid yang berdekatan, maka kadang-kadang segmen tersebut mengubah pasangannya, yang membentuk kromatid rekombinan. Jika dua kromatid berasal dari satu homolog, yaitu tergabung pada sentromernya (dinamakan sister chromatid), pindah silang tidak mempunyai pengaruh, karena sister chromatid merupakan kembaran dari satu sama lainnya. Akan tetapi, jika dua
kromatid tersebut adalah non-sister chromatid (satu dari homolognya dan satu dari yang lain), pinda silang menyebabkan pertukaran seimbang dari gen antara kromosom homolog, seperti ditunjukkan pada Gambar 1.11. Seberapa jauh bahwa pematahan terjadi lebih-kurang secara acak di sepanjang setiap kromosom, ada hubungan langsung antara jarak nyata yang memisahkan dua lokus pada satu kromosom, dan jumlah rata-rata pinda silang di antara mereka. Sayangnya, jumlah ini tidak dapat diukur secara langsung. Akan tetapi, dengan mengobservasi keturunan dari perkawinan tertentu, kita dapat menghitung fraksi rekombinasi, yang merupakan proporsi gamet dari satu tetua yang hanya dihasilkan dari pindah silang selama meiosis pada tetua itu. Gambar 1.12 mengilustrasikan konsep fraksi rekombinasi, untuk dua kasus ekstrim dari keterpautan sempurna dan independen, dan untuk satu contoh data aktual.
Jika dua lokus berada sangat berdekatan pada kromosom yang sama, maka fraksi rekombinasi yang diamati sangat rendah dan lokus tersebut dikatakan bersifat terpaut erat. Semakin jauh jarak dua lokus pada kromosom yang sama, semakin besar peluang terjadinya pindah silang di antara keduanya, dan oleh karena itu semakin besar fraksi rekombinasinya. Untuk lokus yang terletak sangat berjauhan pada kromosom yang sama, gamet rekombinan sama frekuensinya dengan gamet non-rekombinan, yang menyebabkan nilai fraksi rekombinasi maksimum 50%. (Alasan mengapa
frekuensi rekombinan maksimum adalah 50% merupakan bukti dari Gambar 1.11, yang menunjukkan bahwa pindah silang menyebabkan dua gamet rekombinan dan dua gamet non-rekombinan.) Lokus yang letaknya cukup berjauhan pada kromosom yang sama sehingga mempunyai fraksi rekombinasi 50% dikatakan tak-terpaut secara efektif walaupun mereka sebenarnya berada pada satu kromosom. Mereka dikatakan tak-terpaut secara efektif karena mereka bersegregasi secara bebas, seolah-olah mereka berada pada kromosom yang berbeda.
Homolog 1 Homolog 2 non-sister kromatid patah Reunion dan crossing over Non-rekombinan
Gambar 1.11. Empat tahap yang dilibatkan dalam pindah silang antara sepasang
kromosom homolog.
Hubungan antara fraksi rekombinasi dan jarak antara dua lokus memungkinkan pembuatan peta keterpautan (linkage map), dimana lokus ditempatkan berdasarkan fraksi rekombinasi di antara mereka. Pada peta seperti itu, jarak antara lokus diekspresikan sebagai jarak peta (map distance), (dalam unit dinamakan centimorgan, cM), yang sama dengan 100 kali fraksi rekombinasi. Dengan menghitung fraksi rekombinasi di antara banyak pasangan lokus dalam spesies, kelompok dari lokus terpaut
Rekombinan Rekombinan
(kelompok keterpautan) menjadi dapat dimengerti. Karena lokus dalam tiap grup adalah terpaut, mereka pasti terletak pada kromosom yang sama. Dengan demikian, itu berarti bahwa jika sejumlah lokus digunakan dalam analisis keterpautan dalam suatu spesies, jumlah kelompok keterpautan sama dengan jumlah pasangan kromosom. Konstruksi peta keterpautan, yang mempunyai aplikasi praktis sangat penting, didiskusikan pada Bab 2.
Gambar 1.12. Konsep fraksi rekombinasi, yang diilustrasikan dengan checkerboard.
Garis pertama dan kedua pada bagian bawah menunjukkan bahwa harapan secara teoritis untuk rekombinasi 0% dan 50%. Garis terakhir menunjukkan beberapa data aktual, yang memberikan fraksi rekombinasi 18%.
Inaktivasi
Inaktivasi-X dan kompensasi dosis
Di antara banyak warna bulu yang dilihat pada kucing, mosaik warna oranye dan non-oranye, yang dikenal sebagai tortoiseshell (Gambar 1.13a) merupakan satu dari yang paling menarik. Rambut oranye disebabkan oleh alel O yang terpaut X, yang mencegah produksi pigmen gelap (hitam dan coklat), tetapi memungkinkan produksi pigmen kuning. Rambut non-oranye
adalah karena alel normal (tipe liar/wild-type) pada lokus yang sama, o, yang memungkinkan produksi pigmen gelap, dengan cara apapun yang ditentukan oleh alel pada lokus bulu tubuh berwarna lainnya. (Lihat Bab 12 untuk informasi lebih jauh mengenai genetika warna bulu tubuh.) Karena kedua alel harus ada agar dapat menghasilkan mosaik oranye dan non-oranye, kucing tortoiseshell tentunya bersifat heterozigot, XOXo, pada lokus
terpaut-X ini. Tetapi mengapa beberapa bagian dari badan tersebut menampilkan pengaruh dari alel oranye, sedangkan bagian lain menampilkan pengaruh dari alel non-oranye? Dan mengapa pola dari oranye dan non-oranye kira-kira sama pada keseluruhan area, dan mengapa mereka tersebar lebih- kurang secara acak ke seluruh bulu tubuh?
Jawaban untuk pertanyaan ini sebagian terdapat pada pengamatan lain yang pertama kali dibuat pada kucing, oleh Barr dan Bertram, yang pada tahun 1949 melaporkan bahwa nukleus dari sel syaraf yang tidak membelah pada betina biasanya mengandung tubuh berwarna gelap, sedangkan hal yang sama pada jantan tidak (Gambar 1.13b). Tubuh berwarna gelap, yang sekarang dikenal sebagai Barr body atau kromatin sex. Walaupun itu telah diamati oleh banyak peneliti sebelumnya, Barr dan Bertram adalah peneliti pertama yang mencatat bahwa tubuh berwarna gelap terjadi hanya pada sel betina. Dalam upaya untuk menerangkan observasinya, mereka menduga bahwa itu mungkin kromosom X yang telah menjadi sangat padat dan kompak. Peneliti lain menunjukkan bahwa mereka benar; Barr body, pada kenyataannya, adalah kromosom X yang terlambat mereplikasi selama mitosis.
Mengambil contoh dari pengamatan serupa pada mencit, Mary Lyon menyatakan pada tahun 1961 bahwa kromosom X yang sangat padat dan kompak yang dilihat pada sel betina merupakan hasil dari satu di antara kromosom X (dipilih secara acak) yang menjadi tidak aktif pada tiap sel dari semua embrio betina pada tahap awal dari perkembangan. Ini dikenal sebagai hipotesis Lyon. (Pada kenyataannya, sekarang telah diketahui bahwa tidak semua gen pada kromosom X yang inaktif adalah inaktif; gen yang berada pada dan dekat daerah pseudo-autosomal tetap berfungsi pada kedua kromosom X.)
Karena hipotesis Lyon menyimpulkan bahwa pemilihan X untuk inaktivasi seluruhnya bersifat acak, itu menunjukkan bahwa setiap kromosom X pada betina normal akan bersifat aktif pada kira-kira separuh dari semua sel betina.
Proses inaktivasi-X secara acak memberikan penjelasan yang cukup untuk warna bulu tubuh tortoiseshell; tiap pola warna oranye mencerminkan sel yang diwariskan dari sel tempat di mana alel non-oranye tidak diaktifkan, dan sebaliknya. Selain itu, penyebaran pola yang nampak acak tersebut, dan area total oranye dan non-oranye yang kira-kira sama, dapat diharapkan jika X yang tak diaktifkan terpilih secara acak.
Dalam pewarisan, perlu dicatat bahwa karena kucing tortoiseshell bersifat heterozigot pada lokus terpaut-X, mereka tentunya mempunyai dua kromosom X, pada kasus itu mereka semestinya betina. Kucing jantan normal, yang hanya mempunyai satu kromosom X, bisa oranye (XOY) atau
bukan-oranye (XoY), tetapi tidak tortoiseshell. Jadi, dugaan yang cukup
aman bahwa setiap kucing tortoiseshell adalah betina. Kejadian yang jarang untuk tortoiseshell jantan pernah dilaporkan, tetapi mereka biasanya berubah menjadi jantan yang tidak normal yang mempunyai kromosom X ekstra, seperti diterangkan pada Bab 4.
Hasil dari inaktivasi-X secara acak adalah bahwa setiap betina bersifat mosaik, yang terdiri atas dua populasi sel berbeda yang berasal dari sumber yang sama; pada satu populasi sel kromosom X maternal (yaitu kromosom X yang berasal dari ibu) bersifat tidak aktif, dan pada populasi sel lain, X
paternal bersifat tidak aktif.
Satu-satunya pengecualian terhadap inaktivasi-X acak yang sampai kini tercatat terjadi pada kanguru, dimana kromosom X paternal yang tidak diaktifkan. Alasan untuk ini tidak diketahui.
Jelaslah hasil akhir dari inaktivasi-X pada betina adalah bahwa tiap sel betina mempunyai jumlah produk gen yang sama dari gen terpaut-X seperti pada sel jantan. Jadi, inaktivasi-X merupakan mekanisme yang menukaralihkan untuk perbedaan dalam 'dosis' gen antara jantan dan betina berkaitan dengan gen terpaut-X. Pengaruh inaktivasi-X ini dinamakan kompensasi dosis.
Gambar 1.13. (a) Seekor kucing tortoiseshell dengan totol-totol putih. Totol-totol
putih tersebut karena satu alel pada lokus autosom (lihat Bab 12). (b) Motor neurones dari nukleus hypoglossal dari kucing betina dewasa (kiri) dan kucing jantan dewasa (kanan). Tubuh yang berwarna gelap pada tiap sel adalah nukleolus. Tubuh berwarna terang kecil (panah) pada sel betina adalah Barr body.
Akhirnya, perbedaan penting antara mamalia dan burung harus dicatat: ketika inaktivasi-X tampak terjadi pada semua mamalia, inaktivasi-Z tidak terjadi pada burung. Alasan untuk ini tidak diketahui.
Perekaman (imprinting)
Inaktivasi tidak terbatas pada kromosom X saja. Pada lokus tertentu di kromosom lain, sejauh mana suatu alel diekspresikan (atau bahkan apakah itu diekspresikan semuanya) tergantung pada tetua asalnya. Ekspresi berbeda dari elel ini dinamakan perekaman genom (genomic imprinting). Seperti mungkin dibayangkan, ini dapat menjadi sumber frustasi dalam
upaya menetukan model pewarisan abnormalitas, karena perekaman dapat menyebabkan pola pewarisan tak khas.
Inaktivasi akibat dari metilasi
Pada level molekuler, inaktivasi berhubungan dengan penambahan grup metil (CH3) ke molekul sitosin yang terjadi dekat dengan sisi 5´
molekul guanin, yaitu inaktivasi terkait dengan metilasi sitosin pada suatu yang disebut CpG island, dimana p kependekan dari ikatan fosfat antara dua basa yang berdekatan. Dalam individu seekor hewan, semua turunan dari setiap sel tempat kejadian pertama inaktivasi mempunyai gen atau kromosom inaktif yang sama, karena setelah setiap replikasi dari rangkaian DNA termetilasi, rangkaian baru tersebut secara otomatis termetilasi pada tempat CpG yang sama seperti pada rangkaian aslinya. Akan tetapi, pada meiosis atau pada perkembangan embrio awal, pola metilasi diatur kembali.
Tidak semua pola metilasi merupakan satu set sepanjang kehidupan hewan. Pada kenyataannya, pada daerah yang dijadikan inaktivasi-X atau perekaman, metilasi CpG island pada promoter merupakan karakter umum dari gen inaktif, dan metilasi merupakan prasyarat untuk transkripsi banyak gen. Jadi metilasi merupakan cara lain gen diregulasi.
Tipe DNA
Walaupun pentingnya gen sudah sangat jelas, tidak semua DNA terdiri atas gen. Pada kenyataannya, hanya proporsi kecil dari genom hewan terdiri atas gen; mungkin kurang dari 10% dan mungkin hanya sekitar 1% nya. Bagaimana kita dapat mengkategorikan DNA total, dan dimanakah posisi gen?
Kategori DNA yang paling umum berupa sekuen unik (unique) atau kopi tunggal (single-copy), yang kira-kira membentuk 60--70% dari genom mamalia. Sekuen kopi tunggal ini tersebar di seluruh genom. Proporsi kecil dari DNA ini sebagian besar merupakan gen.
Beberapa gen terjadi dalam bentuk multigene family, yang trediri atas sekumpulan gen yang sangat mirip atau identik, yang jumlah inidividualnya biasanya tersebar di seluruh genom, atau, dalam beberapa kasus, terjadi sebagai sekelompok gen yang berdekatan. Tidaklah mengherankan, gen yang terjadi dalam bentuk multigene family adalah gen yang produknya diperlukan dalam jumlah yang relatif besar, misalnya histon, keratin, kolagen, ribosomal RNA, dan transfer RNA.
Kategori utama ke tiga dari DNA adalah repetitive DNA, yang terdiri atas kopi-kopi multiple dari sekuen tertentu yang dinamakan unit ulangan (repeat unit), yang ukurannya berkisar dari basa tunggal sampai