TINJAUAN PUSTAKA
2.1. Masalah Resistensi
Populasi suatu serangga yang dikendalikan, pada mulanya rentan terhadap insektisida yang digunakan untuk memberantasnya. Pada beberapa generasi, keampuhan dari insektisida itu semakin menurun sebab serangganya semakin toleran terhadap insektisida dan akhirnya tidak berdayaguna lagi sebab serangga yang diberantas sudah menjadi resisten terhadap insektisida yang digunakan (Brown dan Pal, 1971).
Menurut Darwin (dalam Villee, 1957; Bishop, 1982) di alam terjadi seleksi alamiah terhadap makhluk hidup, yang dapat meningkatkan daya penyesuaian populasi terhadap lingkungannya. Hal ini mungkin terjadi bila faktor-faktor yang menentukan daya penyesuaian itu mempunyai keragaman. Darwin mengatakan bahwa faktor-faktor itu juga diwariskan kepada keturunannya. Mendel (dalam Villee, 1957) menyatakan bahwa hanya karakter yang diatur oleh gen yang dapat diwariskan. Fisher (dalam Bishop, 1982) menyatakan bahwa laju pertambahan daya penyesuaian sebanding dengan keragaman genetik yang mengatur daya penyesuaian itu.
Toleransi suatu spesies serangga terhadap insektisida sangat beragam, dapat terbukti dengan terjadinya berbagai presentasi kematian bila beberapa kelompok serangga dari spesies yang sama, dipaparkan dengan berbagai dosis atau konsentrasi insektisida. Toleransi itu berkisar antara 0 dan 100% yang merupakan distribusi kumulatif normal yang disebut sebaran toleransi sedangkan Macnair (1981) menyebut differential susceptibility. Wood (1981) telah mengumpulkan fakta-fakta yang menunjukkan bahwa toleransi diatur oleh gen R. Misalnya resistensi terhadap DDT pada A. aegypti diatur oleh gen RDDT1 dan gen RDDT2 . Resistensi terhadap dieldrin diatur oleh gen Rdl , terhadap pyretroid oleh gen Rpy dan terhadap HCH/cyclodien diatur oleh gen y. Gen-gen RDDT1 , y dan Rdl terdapat pada kromosom II sedangkan gen-gen RDDT2 dan Rpy terdapat pada kromosom III. Alel lainnya dari gen R adalah gen (+). Oleh karena kromosom berpasangan maka gen juga berpasangan, sehingga dari gen R dan (+) diperoleh tiga kombinasi genotip
yaitu RR, R(+) dan (+)(+), yang paling toleran adalah RR sedangkan yang paling rentan adalah (+)(+). Fenotip dari R(+) tergantung kepada kombinasi dominasi gen R. Bila gen R dominan maka fenotip dari R(+) menyerupai RR, bila gen R resesif maka fenotip dari R(+) menyerupai (+)(+) dan bila gen intermediate maka fenotip dari R(+) beragam diantara RR dan (+)(+). Oleh karena toleransi terhadap pestisida beragam dan diatur oleh gen maka toleransi ini akan mengalami seleksi bila kontak dengan pestisida sehingga menjadi lebih toleran untuk kemudian menjadi resistensi.
Teori Darwin (dalam Villee, 1957) menyatakan bahwa seleksi alam menyebabkan punahnya individu-individu yang daya penyesuainnya lemah, sedangkan yang daya penyesuainnya baik akan terus mempertahankan eksistensi populasinya. Seperti halnya dengan gen (+) yang (+) akan punah sedangkan gen RR akan mampu hidup bila kontak dengan insektisida. Untuk gen R(+) keadaannya tergantung kepada dominasi gen R, bila gen R dominan maka R(+) akan terus hidup bersama-sama RR, sedang bila gen R resesif maka R(+) akan punah bersama-sama (+)(+). Bila gen R bersifat intermediate maka nasib R(+) tergantung pada dosis yang digunakan. Culex pipiens pipiens mempunyai gen R dominan, Anopheles sp mempunyai gen R resesif sedangkan A. aegypti mempunyai gen R yang intermediate (Wood dan Mani, 1981; Wood, 1981).
2.2. Faktor yang mempengaruhi kerentanan
Faktor – faktor seperti suhu, kelembaban nisbi, umur, jenis kelamin, dan berat badan dapat mempengaruhi kerentanan serangga terhadap insektisida (Bainbridge et al. 1982)
Ogushi et al. (1968) meneliti pengaruh suhu atas kerentanan larva Culex pipiens terhadap beberapa insektisida. Uji kerentanan terhadap dieldrin dan DDT menunjukkan peningkatan konsentrasi letalitas 50% (KL50) bila suhu semakin tinggi. Sebaliknya bila menggunakan fenthion, fenitrothion, fenchlorphos, diazinon, malathion, trichlorfon, dichlorvos, naled dan lindane menunjukkan penurunan KL50 dengan meningkatnya suhu.
Faktor lain yang mempengaruhi kerentanan serangga terhadap insektisida adalah umur. Brown dan Pal (1971) meneliti KL50 larva A. aegypti mulai umur satu hari sampai umur 8 hari terhadap DDT dan dieldrin dengan waktu kontak 1 dan 24 jam. Bila waktu kontak 1 jam maka KL50 terhadap DDT berkisar antara 2,2- 3,00
ppm sedang bila waktu kontaknya 24 jam maka KL50 terhadap DDT berkisar antara 1,2 pada hari keempat sampai 30 ppm pada hari kedelapan.
Kondisi pemeliharaan ternyata besar pengaruhnya terhadap kerentanan. Bila kepadatan meningkat dari 200 menjadi 2000 larva pernampan maka KL50 akan menurun 20 kali yang berarti larvanya semakin rentan (Garm dalam Brown dan Pal, 1971). Bila larva lapar selama 4-6 jam maka KL50nya mengecil menjadi seperduanya (Jones, 1971).
Penelitian Rosen (1967) menunjukkan bahwa kerentanan larva Culex pipiens pipiens terhadap DDT dan dieldrin serta gamma HCH di Rangoon (Burma), paling rendah pada musim kemarau dan paling tinggi pada peralihan musim. KL50 mempunyai keragaman paling tinggi pada musim angin muson (Monsoon). Toleransi A. taeniorhynchus terhadap beberapa pestisida pada musim semi, 2 – 8 kali toleransinya pada musim panas (Keller dalam Brown dan Pal, 1971).
Bainbrigde et al. (1982) meneliti kerentanan larva dan dewasa nyamuk A. aegypti dari galur rentan dan resisten terhadap DDT selama 12 jam periode gelap dan 12 jam periode terang secara bergantian dan bersambung. Galur rentan menunjukkan tiga saat paling rentan yang ditunjukkan oleh angka kematian yang paling tinggi dengan kisaran antara 32% sampai 37%. Galur ini menunjukkan dua saat yang paling toleran dengan kematian 5% dan 7%. Saat yang paling rentan adalah dua jam setelah gelap, empat jam setelah waktu gelap dan saat peralihan periode gelap menjadi terang. Saat paling toleran terjadi pada peralihan periode terang menjadi gelap, dan empat jam sebelum terang. Galur resisten hanya menunjukkan satu saat yang paling rentan yang terjadi pada pertengahan periode terang.
2.3. Mekanisne resistensi
Resistensi serangga terhadap insektisida diartikan sebagai kemampuan peningkatan daya tahan suatu populasi serangga terhadap insektisida yang biasanya mematikan. Pada suatu populasi serangga, individu-individu yang pada dasarnya sudah resisten terhadap suatu insektisida akan menghasilkan keturunan yang resisten (Brown dan Pal, 1971).
Adapun kerugian penggunaan insektisida adalah timbulnya galur-galur serangga sasaran yang resisten terhadap insektisida. Mekanisme resistensi serangga
terhadap insektisida dapat terjadi melalui perubahan fisiologi dan perubahan tingkah laku (O’Brien, 1967, Brown dan Pal, 1971). Resistensi fisiologi pada serangga memperlihatkan kenyataan yang lebih penting. Hal ini disebabkan karena faktor-faktor 1) daya absorbsi insektisida yang sangat lambat sehingga serangga tidak mati; 2) daya penyimpanan insektisida dalam jaringan yang tidak vital, seperti jaringan lemak sehingga alat-alat vital terhindar dan serangga tidak mati; 3) daya ekskresi insektisida yang cepat sehingga tidak sampai membunuh serangga; 4) daya beberapa enzim yang bersifat meniadakan efek keracunan (detoksifikasi) yang menyebabkan serangga tidak mati seperti dehidroklorinase untuk senyawa DDT (Tarumingkeng, 1992).
Mekanisme resistensi perubahan tingkah laku disebabkan karena faktor-faktor 1) perubahan habitat serangga sehingga terhindar dari pengaruh insektisida dan keturunannya akan mempertahankan habitat yang baru tersebut; 2) meningkatnya kepekaan serangga terhadap insektisida, dengan demikian serangga tersebut dapat menghindarkan diri dari pengaruh insektisida sehingga tidak terbunuh, tanpa mengubah habitatnya.
Lalat rumah yang resisten terhadap DDT mempunyai kutikula yang lebih tebal pada pulvilusnya dari pada galur yang rentan. Kutikula berpengaruh terhadap penetrasi DDT, hal ini diatur oleh gen organotonin-R atau tin (Plapp dan Hoyer, 1968). Ketebalan ini dapat menyebabkan lalat rumah menjadi lebih toleran terhadap pestisida lainnya (Plapp, 1970)
Finney (1964) menyatakan bahwa toleransi suatu spesies serangga terhadap insektisida adalah beragam. Toleransi berkisar antara 0 sampai 100% yang merupakan distribusi komulatif normal yang disebut sebagai sebaran toleransi. Hal ini dapat ditunjukkan bila beberapa kelompok serangga dari spesies yang sama dikontakkan dengan berbagai dosis insektisida yang sama.
Senyawa fosfor organik dan karbamat bekerja sebagai penghalang kolinesterase, suatu enzim yang berfungsi dalam rangsangan syaraf dan bekerja pada sinap kolinergik. Adapun senyawa hidrokarbon berklor bekerja menimbulkan kerusakan pada komponen selubung sel-sel syaraf menyebabkan fungsinya terganggu (Matsumura, 1975). Brown dan Pal (1971) menyatakan bahwa resistensi
mengandung gen-gen yang tahan terhadap insektisida.
Detoksikasi diatur oleh gen madya (intermediate) yang menghasilkan resistensi melalui peningkatan kemampuan detoksikasi insektisida (Plapp, 1970). Dehidroklorinase adalah faktor utama pada detoksikasi DDT. Detoksikasi senyawa orgonfosfat (OP) dengan karboksilesterase, fosfotriesterase, asetilkolinesterase (AChE), glutation dependen transferase dan mixed function oxidase (MFO) (Georghiou dan Pasteuer, 1978). Dehidroklorinase pada larva A.aegypti galur yang resisten menghasilkan DDT lebih banyak dibanding dengan yang dihasilkan oleh galur yang rentan (Abedi et al. 1963). Kimura dan Brown (1964) menyatakan bahwa dehidroklorinase mempunyai korelasi kuat dengan tingkat resistensi A.aegypti galur Amerika Tengah, tetapi lemah pada galur Asia, Rathor dan Wood (1981) menemukan bahwa DDT dihasilkan lebih banyak oleh galur resisten dari pada galur rentan, bukan hanya pada galur Amerika tetapi juga pada galur Asia dan Afrika.
Senyawa OP dan karbamat berfungsi menghambat pekerjaan enzim AChE. Bila serangga mampu menggurangi pengaruh itu melalui seleksi alam, maka serangga tersebut akan menjadi resisten terhadap OP dan Karbamat (Wood, 1981). Hemingway dan Georghiou (1983) menyimpulkan bahwa pada larva Anopheles albimanus yang resisten, AChE semakin sukar dihambat dibanding dengan AChE pada galur yang rentan dan ada korelasi yang kuat antara laju hambat AChE dengan tingkat resistensi serangganya.
Sifat merangsang (iritasi) beberapa insektisida dapat menyebabkan serangga meninggalkan permukaan yang ada lapisan insektisidanya, sebelum memperoleh dosis yang mematikan. Hal ini menyebabkan kontak berulang-ulang untuk menghasilkan kematian. Sifat ini dikenal pada DDT terutama terhadap Anopheles sp. (Brown dan Pal, 1971).
Selain itu pula resistensi silang (cross resistance) dapat terjadi bila suatu spesies serangga resisten terhadap beberapa insektisida dalam satu golongan. Hal ini dapat terjadi karena secara umum daya kerja insektisida di dalam satu kelas adalah sama, misalnya malation dan paration atau fosfor organik yang lain. Suatu spesies serangga juga dapat mengalami resistensi majemuk (multiple resistance) yaitu
berbeda (Brown dan Pal, 1971; WHO, 1976).
Faktor-faktor yang mempengaruhi laju perkembangan resistensi adalah 1) faktor genetik; 2) faktor ekologi; 3) faktor fisiologi/biokimia dan 4) frekuensi penggunaan insektisida (Metcalf dan Luckman, 1975; WHO, 1976)
Laju perkembangan resistensi untuk setiap spesies organisme berbeda. Resistensi dapat berkembang sangat cepat pada suatu spesies tapi lebih lambat pada spesies yang lain. Menurut Fisher semakin besar keragaman genetik yang mengatur resistensi, maka semakin cepat pula tercapainya status resistensi. Wood dan Mani (1981) mengemukakan bahwa bila gen R dominan maka resistensi semakin cepat tercapai dibanding dengan bila gen R intermediate atau resesif. Demikian pula bila gen R intermediate , resistensi akan lebih cepat tercapai dari pada bila gen R resesif. Faktor ekologi yang penting dalam mempengaruhi laju resistensi adalah tingkah laku isolasi, migrasi dan mobilitas dari individu-individu yang bersifat rentan (WHO, 1975). Faktor fisiologi dan biokimia sangat berpengaruh terhadap laju resistensi suatu populasi serangga. Kecepatan menghasilkan keturunan merupakan faktor yang penting, semakin besar jumlah generasi per tahun yang dihasilkan maka makin cepat terjadi evolusi resistensi.
Anopheles stephensi di Irak hanya membutuhkan waktu selama sekali perlakuan dengan DDT untuk menjadi resistensi terhadap DDT. Spesies yang sama di Arab Saudi membutuhkan waktu 5 tahun untuk menjadi resistensi terhadap DDT (Davidson, 1958). Culex pipiens pipiens setelah mengalami kontak selama 40 generasi, toleransinya meningkat 100 kali semula (RF = 100), sedang bila kontak dengan triprene selama 27 generasi RF nya hanya 10 (WHO, 1980). RF (resistance factor (Wood, 1981)atau resistance ratio (Brown dan Pal, 1971) adalah rasio antara KL50 atau KL90 dari satu spesies yang telah mengalamai seleksi dengan KL50 atau KL90 dari populasinya yang normal.
Status dan ukuran kerentanan serangga terhadap insektisida dapat diukur dengan uji kerentanan (Susceptibility). Uji ini didasarkan atas kontak serangga dengan insektisida yang menghasilkan respon berupa kematian. Status kerentanan dapat dilihat dari hasil kontak selama periode waktu tertentu, misalnya satu jam dengan masa pengamatan 24 jam (WHO, 1975; Dit. Jen. PPM & PLP, 1986).
jurkan untuk menggunakan dosis diagnosa tentative yang besarnya tergantung pada spesies, jenis insektisida dan stadium hidup serangganya. Untuk menentukan status kerentanan nyamuk A. aegypti dewasa terhadap malation dengan konsentrasi 5%.
Pelacakan untuk mengetahui timbulnya galur serangga yang resistensi perlu dilakukan di berbagai tempat secara periodik. Hal ini disebabkan karena waktu timbulnya resistensi tidak dapat diduga dengan tepat sebab tergantung pada tempat, waktu, spesies serangga sasaran, dan insektisida yang digunakan. Faktor-faktor seperti suhu, kelembaban nisbi, umur, jenis kelamin, dan berat badan mempengaruhi kerentanan serangga terhadap insektisida (Bainbridge et al. 1982).
Brown dan Pal (1971), mengatakan bahwa hingga akhir dekade enam puluhan, belum ada laporan adanya galur A. aegypti yang resisten terhadap senyawa OP. Tetapi WHO (1980) telah mencatat adanya galur spesies ini yang resisten terhadap senyawa OP seperti malathion, temephos dan fenitrithion di Kepulauan Karibia, malation di India, Malaysia, Thailand dan Vietnam.
Masalah resistensi sudah dilaporkan di beberapa wilayah Indonesia seperti Jakarta, Yogyakara, Surabaya dan Palembang menunjukkan populasi nyamuk A. aegypti asal Jakarta dan Surabaya, dengan kematian 92,8% dan 91% sudah mulai toleran terhadap malation (Soekirno et al, 1990). Di wilayah Jabotabek, populasi A. aegypti dari Jakarta Pusat telah resisten terhadap malation 5%, populasi dari Bogor dan Tangerang masih rentan sedangkan populasi dari Jakarta Utara, Jakarta Barat, Jakarta Timur, Jakarta Selatan, dan Kota Bekasi telah toleran terhadap insektisida organophosphat dan pyretroid (Shinta et al, 2006). Seperti halnya di wilayah endemik Kel. Duren Sawit Jakarta Timur penggunaan insektisida malation dengan konsentrasi 0,8% dan 5% belum terindikasi resisten, tetapi menuju ke arah toleran (Marisa. 2007).
Di Yogyakarta dari 14 kelurahan (8 kecamatan) nyamuk A.aegypti masih rentan (RR) sebesar 96,7% dan 3,3% (RS) toleran terhadap insektisida piretroid, di Kab. Bantul dari 14 kelurahan (9 kecamatan), beragam status resistensinya terhadap insektisida organofosfat dengan mekanisme peningkatan aktivitas enzim esterase non - spesifik (Mardihusodo, 1995). Sementara populasi nyamuk A. aegypti di Kota Bandung belum terjadi resistensi oleh insektisda malation (Sulianti et al, 2008).
Resistensi di Brazil dilaporkan oleh Lima et al, (2003) adanya tanda-tanda resistensi larva dan dewasa A. aegypti terhadap beberapa jenis insektisida, sampel diambil dari 10 kota di Rio de Janeiro dan Espirito Santo, delapan kota yang resisten terhadap temefos (0,012 mg/L), dengan tingkat mortalitasnya adalah 23,5% - 74%.
Menurut Dep. Kes R.I (1986), macam-macam resisten yang terjadi pada serangga yang disebabkan oleh insektisida adalah; 1) Resistensi fisiologis (physiological resistance), resisten yang disebabkan oleh mekanisme fisiologis suatu gen sehingga menurun dimana populasi nyamuk sebagai mahluk hidup akan mengadakan reaksi akibat adanya tekanan racun serangga, dengan cara menghasilkan enzym untuk menawarkan daya racun serangga, mengikat racun serangga dalam jaringan lemak, memblokir racun serangga dalam tubuh, atau segera mengeluarkan racun serangga dari dalam tubuhnya; 2) resistensi perilaku (behaviouristic resistance). resisten yang disebabkan oleh kepekaan terhadap adanya rangsangan dari racun serangga yang menyebabkan nyamuk menghindari kontak dengan racun tersebut sehingga yang sensitif akan hidup dan kurang sensitif akan mati; 3) resistensi bersifat toleransi (toleransi resistance), resisten bukan karena faktor genetik, karena variasi musiman seperti bentuk yang lebih besar, kutikula menebal, kenaikan kandungan lemak, sehingga konsentrasi insektisida tidak cukup untuk mematikannya; 4) resistensi terhadap kelompok insektisida yang sama (cross resistance), kekebalan yang terjadi pada racun serangga lain dalam kelompok yang sama, misalnya penyemprotan dieldrin menyebabkan serangga kebal terhadap DDT atau gamexane.
2.4. Penyakit Demam Berdarah
Penyakit demam berdarah merupakan masalah kesehatan di Asia Tenggara sejak tahun 1950. Kasus penyakit demam berdarah pertama kali dilaporkan di Filipina tahun 1954, kemudian Thailand 1958, Singapura 1960, Vietnam 1961, Malaysia 1962, India 1963 dan Indonesia 1968 (Kho et al, 1969; Hammon, 1973). Hsiech et al. (1982) melaporkan bahwa demam berdarah dengue merupakan masalah yang sangat serius di Asia Tenggara. Pada tahun 1981 terjadi wabah di Taiwan yang disebabkan oleh virus dengue tipe 2. Selanjutnya tahun 1987 sampai
1988 terjadi wabah yang disebabkan oleh virus dengan tipe 1, dengan jumlah kasus sebanyak 10,420 orang, Di Taiwan virus tipe 1 ini telah diketahui sejak tahun 1945.
Penyakit demam berdarah adalah penyakit menular yang disebabkan oleh virus dengan tipe 1, 2, 3 dan 4 yang ditandai dengan demam mendadak 2 sampai 7 hari, lemah atau lesu disertai tanda pendarahan di kulit berupa bintik-bintik kadang-kadang mimisan, muntah darah, berak darah, kesadaran menurun atau shock (Krupp dan Chatton, 1976; Soedarmo, 1988; WHO, 1986).
Virus dengue memperbanyak diri dan menyebar ke seluruh tubuh nyamuk, termasuk ke kelenjar liurnya, virus dengue ini dapat dibedakan dalam tipe 1, 2, 3 dan 4 (Jawetz, 1982; Kettle, 1984). Infeksi virus ini pada manusia mengakibatkan suatu manifestasi klinis yang bervariasi (WHO, 1986) yaitu : a) Demam Dengue (Dengue klasik atau “Silent Dengue Infection”); b) Demam berdarah dengue (“Dengue Haemorragic Fever”); dan c)Dengue dengan renjatan (“Dengue Shock Syndrome DSS”). Orang yang terinfeksi virus dengue, tidak semuanya akan sakit demam berdarah dengue. Ada yang menderita demam ringan yang akan sembuh dengan sendirinya atau bahkan ada yang sama sekali tanpa gejala sakit, tetapi semuanya merupakan pembawa virus dengue selama kurang lebih tujuh hari sehingga dapat menularkan kepada orang lain di berbagai wilayah yang ada nyamuk penularnya (Ramalingam, 1974).
Di Indonesia kasus demam berdarah ditemukan pertama kali pada tahun 1968 di Surabaya dan Jakarta dan dari tahun ke tahun ada kecenderungan peningkatan kasus. Dalam tahun 1988 diketahui ada 47,573 orang penderita dan sebanyak 1,527 orang meninggal atau 3,2 % (Suroso, 1991). Seluruh wilayah mempunyai resiko untuk kejangkitan penyakit demam berdarah dengue, namum tempat yang potensial bagi penyebaran penyakit adalah rumah-rumah maupun tempat-tempat umum, kecuali yang ketinggiannya lebih dari 1000 meter dari permukaan laut. Semua tipe virus penyebab demam berdarah telah diisolasi di Indonesia (Suharyono, 1990).
Nyamuk A. aegypti sebagai vektor utama penyakit demam berdarah tersebar luas di seluruh Indonesia. Meskipun nyamuk ini banyak ditemukan di daerah perkotaan yang padat penduduk, namun ditemukan juga di daerah pedesaan. Faktor lingkungan fisik yang penting dalam mempengaruhi endemisitas penyakit demam berdarah terutama berkaitan dengan keberadaan nyamuk A. aegypti.
Kelembaban udara akan mempengaruhi umur nyamuk. Nyamuk A. aegypti hanya dapat menularkan penyakit demam berdarah bila umurnya lebih dari sepuluh hari, karena masa inkubasi ekstrinsik virus dengue di dalam tubuh nyamuk antara delapan sampai sepuluh hari. Disamping itu faktor curah hujan akan menambah genangan air sebagai tempat perindukan, dengan demikian populasi vektor akan bertambah dan kemungkinan terjadinya penularan penyakit demam berdarah lebih besar lagi (Soedarmo, 1988; Dit.Jen. PPM & PLP, 1990).
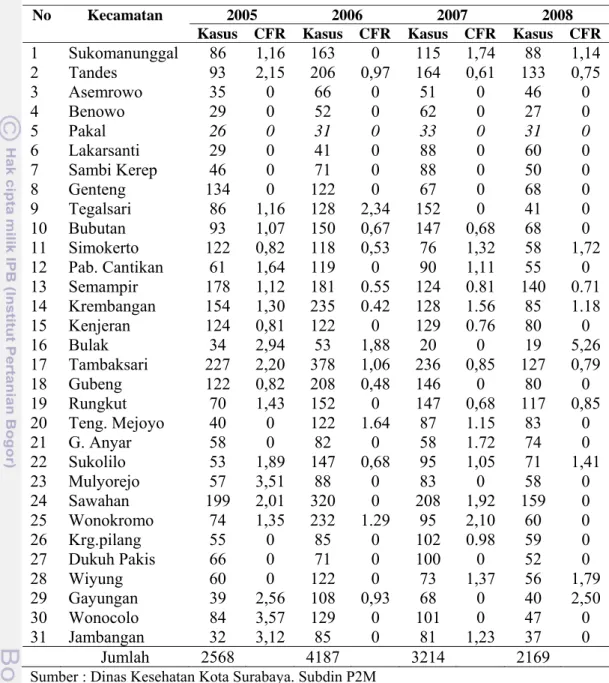
Di Kota Surabaya, dilaporkan angka kesakitan dan angka kematian penyakit demam berdarah oleh Subdin P2M Dinas Kesehatan Kota dari tahun 2005-2008 seperti lokasi dengan kasus tinggi di Kecamatan Sawahan, tahun 2005 (CFR =2,01%) dan tahun 2007 (CFR =1,92%) sedangkan di Kecamatan Tambaksari secara berurutan angka CFR adalah 2,20%, 1,06%, 0,85% dan 0,79%. Pada lokasi kasus sedang di wilayah Wiyung, tahun 2007 (CFR =1,37%) dan tahun 2008 (CFR =1,78%). Pada Wilayah Wonocolo, tahun 2005 angka (CFR =3,57%). Sementara diwilayah Bulak, tahun 2005 (CFR = 2,94%), tahun 2006 (CFR=1,88%), dan tahun 2008 (CFR = 5,26%) pada (Lampiran 3). Gambaran kasus penyakit demam berdarah oleh Subdin P2M Dinas Kesehatan Kota Surabaya Tahun 2005-2008 ditunjukkan pada Tabel 1.
2.5. Biologi Aedes aegypti
Siklus hidup vektor yang berperan dalam penularan demam berdarah dengue adalah nyamuk family Culicidae, subfamily Culicinae, genus Aedes spesies aegypti (Gubler et al. 1978). Nyamuk ini mengalami metamorfosis sempurna (holometabola) ditunjukkan pada Gambar 1 berikut :
Telur Larva Pupa Dewasa Gambar 1 Siklus hidup nyamuk A. aegypti (www.google.com)
Tabel 1 Insiden dan Case Fatality Rate Demam Berdarah di Kota Surabaya Tahun 2005-2008
No Kecamatan 2005 2006 2007 2008
Kasus CFR Kasus CFR Kasus CFR Kasus CFR 1 Sukomanunggal 86 1,16 163 0 115 1,74 88 1,14 2 Tandes 93 2,15 206 0,97 164 0,61 133 0,75 3 Asemrowo 35 0 66 0 51 0 46 0 4 Benowo 29 0 52 0 62 0 27 0 5 Pakal 26 0 31 0 33 0 31 0 6 Lakarsanti 29 0 41 0 88 0 60 0 7 Sambi Kerep 46 0 71 0 88 0 50 0 8 Genteng 134 0 122 0 67 0 68 0 9 Tegalsari 86 1,16 128 2,34 152 0 41 0 10 Bubutan 93 1,07 150 0,67 147 0,68 68 0 11 Simokerto 122 0,82 118 0,53 76 1,32 58 1,72 12 Pab. Cantikan 61 1,64 119 0 90 1,11 55 0 13 Semampir 178 1,12 181 0.55 124 0.81 140 0.71 14 Krembangan 154 1,30 235 0.42 128 1.56 85 1.18 15 Kenjeran 124 0,81 122 0 129 0.76 80 0 16 Bulak 34 2,94 53 1,88 20 0 19 5,26 17 Tambaksari 227 2,20 378 1,06 236 0,85 127 0,79 18 Gubeng 122 0,82 208 0,48 146 0 80 0 19 Rungkut 70 1,43 152 0 147 0,68 117 0,85 20 Teng. Mejoyo 40 0 122 1.64 87 1.15 83 0 21 G. Anyar 58 0 82 0 58 1.72 74 0 22 Sukolilo 53 1,89 147 0,68 95 1,05 71 1,41 23 Mulyorejo 57 3,51 88 0 83 0 58 0 24 Sawahan 199 2,01 320 0 208 1,92 159 0 25 Wonokromo 74 1,35 232 1.29 95 2,10 60 0 26 Krg.pilang 55 0 85 0 102 0.98 59 0 27 Dukuh Pakis 66 0 71 0 100 0 52 0 28 Wiyung 60 0 122 0 73 1,37 56 1,79 29 Gayungan 39 2,56 108 0,93 68 0 40 2,50 30 Wonocolo 84 3,57 129 0 101 0 47 0 31 Jambangan 32 3,12 85 0 81 1,23 37 0 Jumlah 2568 4187 3214 2169
Sumber : Dinas Kesehatan Kota Surabaya. Subdin P2M
Tahap Telur. Telur A. aegypti berbentuk ellips atau oval memanjang, warna hitam, ukuran 0,5 – 0,8 mm, permukaan polygonal, tidak memiliki alat pelampung dan diletakkan satu per satu pada benda – benda yang terapung atau pada dinding bagian dalam tempat penampungan air (TPA) yang berbatasan langsung dengan air. Dilaporkan bahwa dari telur yang lepas, sebanyak 85% melekat di dinding TPA, sedangkan 15% lainnya jatuh ke permukaan air .
Tahap Larva. Larva nyamuk A. aegypti tubuhnya memanjang dan tanpa kaki dengan bulu – bulu sederhana yang tersusun bilateral simetris. Ciri utama larva A. aegypti adalah bentuk siphon oval agak gemuk dan berwarna kecoklatan. Larva ini dalam pertumbuhan dan perkembangannya mengalami empat kali pergantian kulit (ecdysis), dan larva yang terbentuk berturut – turut disebut larva instar I, II, III dan IV .
Tahap Pupa. Pupa nyamuk A. aegypti bentuk tubuhnya bengkok, dengan bagian kepala – dada (cephalothorax) lebih besar bila dibandingkan dengan bagian perutnya, sehingga tampak seperti tanda baca “koma”. Pada bagian punggung (dorsal) dada terdapat alat bernafas seperti terompet. Pada ruas perut ke-8 terdapat sepasang alat pengayuh yang berguna untuk berenang. Alat pengayuh tersebut berjumbai panjang dan bulu di nomer 7 pada ruas perut ke-8 tidak bercabang. Pupa adalah bentuk tidak makan, tampak gerakannya lebih lincah bila dibandingkan dengan larva. Waktu istirahat posisi pupa sejajar dengan bidang permukaan air.
Tahap Dewasa. Nyamuk A. aegypti tubuhnya tersusun dari tiga bagian, yaitu kepala, dada dan perut. Pada bagian kepala terdapat sepasang mata majemuk dan antena yang berbulu. Alat mulut nyamuk betina tipe penusuk – penghisap (piercing – sucking) dan termasuk lebih menyukai manusia (anthropophagus), sedangkan nyamuk jantan bagian mulut lebih lemah sehingga tidak mampu menembus kulit manusia, karena itu tergolong lebih menyukai cairan tumbuhan (phytophagus). Nyamuk betina mempunyai antena tipe-pilose, sedangkan nyamuk jantan tipe mulut plumose.
Dada nyamuk ini tersusun dari 3 ruas, prothorax, mesothorax dan metathorax. Setiap ruas dada ada sepasang kaki yang terdiri dari femur (paha), tibia (betis) dan tarsus (tampak). Pada ruas – ruas kaki ada gelang – gelang putih, tetapi pada bagian tibia kaki belakang tidak ada gelang putih. Pada bagian dada juga terdapat sepasang sayap tanpa noda – noda hitam. Bagian punggung (mesonotum) ada gambaran garis – garis putih yang dapat dipakai untuk membedakan dengan jenis lain.
Perut terdiri dari 8 ruas dan pada ruas-ruas tersebut bintik-bintik putih. Waktu istirahat posisi nyamuk A. aegypti, tubuhnya sejajar dengan bidang permukaan yang dihinggapinya.
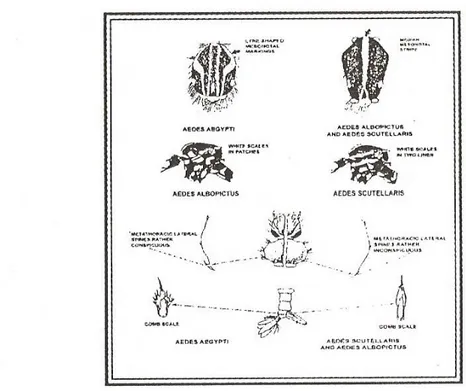
Gambar 2 Ciri-ciri diagnostik untuk membedakan beberapa jenis nyamuk Aedes yang penting di bidang kesehatan (Taboada, 1967).
Morfologi Larva. Larva A. aegypti tubuhnya memanjang tanpa kaki dengan bulu – bulu sederhana yang tersusun bilateral simetris. Larva ini dalam pertumbuhan dan perkembangan mengalami 4 (empat) kali pergantian kulit (ecdysis), dan larva yang terbentuk berturut-turut disebut larva instar I, II, III, dan IV sebagai berikut (Soegijanto, 2003):
Larva instar I memiliki ciri-ciri tubuhnya sangat kecil, warna transparan,
panjang 1 – 2 mm, duri-duri (spinae) pada dada (thorax) belum begitu jelas, dan corong pernafasan (siphon) belum menghitam.
Larva instar II memiliki ciri-ciri tubuhnya tambah besar, ukuran 2,5 – 3,8
mm, duri belum jelas, dan corong pernafasan sudah berwarna hitam.
Larva instar III Pada stadium ini telah memasuki siklus perkembangan larva
menjadi pupa atau berumur sekitar 4 hari.
Larva instar IV Memiliki ukuran tubuh 5 mm dan telah lengkap struktur
anatominya, tubuh dibagi menjadi bagian kepala (chepal), dada (thorax), dan perut (abdomen).
Pada bagian kepala terdapat sepasang mata majemuk, sepasang antenna tanpa duri-duri, dan alat-alat mulut tipe pengunyah (chewing). Bagian dada tampak paling besar dan terdapat bulu-bulu yang simetris. Perut tersusun atas 8 ruas. Ruas perut ke-8, ada alat untuk bernafas yang disebut corong pernafasan. Corong pernafasan tanpa duri-duri, berwarna hitam, dan ada seberkas bulu-bulu (tuft). Ruas ke-8 juga dilengkapi dengan seberkas bulu-bulu sikat (brush) di bagian ventral dan gigi-gigi sisir (comb) yang berjumlah 15 -19 gigi yang tersusun dalam 1 (satu) baris. Gigi-gigi sisir dengan lekukan yang jelas membentuk gerigi. Larva ini tubuhnya langsing dan bergerak sangat lincah, bersifat fototaksis negatif, dan waktu istirahat membentuk sudut hampir tegak lurus dengan bidang permukaan air.
Lamanya perkembangan larva akan tergantung pada suhu, ketersediaan makanan, dan kepadatan larva pada sarang. Habitat alami larva jarang ditemukan, tetapi dapat mencakup lubang pohon, pangkal daun, dan tempurung kelapa. Di daerah yang yang panas dan kering, tanki air di atas, tanki penyimpanan air di tanah, dan septic tank bisa menjadi habitat utama larva. Di wilayah yang persediaan airnya tidak teratur, penghuni menyimpan air untuk kegunaan rumah tangga sehingga semakin memperbanyak jumlah habitat yang ada untuk larva (WHO. 2004).
Habitat. Tempat berkembang biak larva A. aegypti adalah kontainer buatan yang berada di lingkungan perumahan. Habitat larva buatan manusia yang banyak ditemukan di dalam rumah dan sekitar lingkungan perkotaan (rumah tangga, lokasi pembangunan dan pabrik), misalnya botol minuman, pot bunga, bak mandi, tong kayu dan logam, ban, kaleng, pipa saluran (WHO, 2004).
Soegijanto (2003) melaporkan nyamuk Aedes hanya senang di air yang cukup bersih dan tidak langsung beralaskan tanah. Tempat perkembangbiakan utamanya ialah tempat penampungan air di dalam atau di sekitar rumah atau tempat-tempat umum biasanya tidak melebihi jarak 100 meter dari rumah . Habitat larva yang alami seperti pada lubang pohon, bambu, ketiak daun, dan tempurung kelapa merupakan habitat utama larva Aedes .
Agustina (2006), melaporkan bahwa A. aegypti dapat hidup di air terkontaminasi deterjen dengan perolehan telur tertinggi 2,7 ppm, kaporit dengan konsentrasi 10 ppm ditemukan perolehan telur tertinggi, pada tanah konsentrasi 30
gm/ml juga memperoleh jumlah telur tertinggi sedangkan air terkontaminasi feses ayam, perolehan telur tertinggi pada konsentrasi 10 gr/ml.
Aktifitas menghisap darah. A. aegypti dewasa betina menghisap darah
manusia umumnya pada siang hari yang dilakukan baik di dalam rumah maupun di luar rumah. Aktifitas menghisap darah pada nyamuk A. aegypti ini memperlihatkan dua puncak waktu yaitu pukul 08.00 sampai pukul 12.00 dan pukul 15.00 sampai pukul 17.00. Nyamuk ini mempunyai kebiasaan mengisap darah berulang, yaitu aktifitas menghisap darah dilakukan pada beberapa orang dalam waktu singkat secara bergantian. Hal ini disebabkan karena nyamuk ini sangat sensitif dan mudah terganggu. Pada siang hari manusia aktif melakukan pekerjaan dan selalu bergerak, nyamuk belum kenyang darah, orang sudah bergerak lagi maka nyamuk akan terbang untuk menggigit orang lain sampai cukup mendapatkan darah untuk pertumbuhan dan perkembangan telurnya. Keadaan seperti ini sangat membantu nyamuk tersebut dalam transmisi atau memindahkan virus dengue ke beberapa orang (Dit.Jen PPM & PLP, 1990). Perubahan suhu dan kelembaban udara dan angin dapat mempengaruhi aktifitas nyamuk dalam hal menghisap darah (Bruce Chwatt, 1980).
Menurut Marchoux dan Simond (dalam Christopher, 1960) nyamuk A. aegypti pada suhu 15 0C tidak dapat melakukan aktifitas menghisap darah. Umumnya jarak terbang A. aegypti sekitar 50 meter (Horsfall, 1955; Dit.Jen. PPM & PLP. 1990) dengan demikian populasi nyamuk ini penyebarannya akan terpencar – pencar atau tersebar. Menurut Howard, 1923 (dalam Christopher 1960) pada suhu 13 - 14 0C nyamuk akan susah terbang dan kakinya akan lemah.
Jangka hidup. Lama hidup A. aegypti ditentukan terutama oleh tinggi rendahnya suhu dan kelembaban udara, persediaan air dan makanan. Makanan di sini darah dan air juga merupakan kebutuhan pokok bagi serangga karena air sangat penting untuk kelangsungan hidupnya (Christopher, 1960).
Gubler et al. (1978) melaporkan bahwa pada suhu 20 0C dan kelembaban 70% umur nyamuk betina dapat mencapai lebih kurang 100 hari dan nyamuk jantan sekitar 35 hari. Nyamuk yang tidak menghisap darah umurnya lebih pendek daripada yang menghisap darah. Lama hidup A. aegypti di laboratorium dipengaruhi oleh jenis makanan. Menurut Fielding (dalam Christopher, 1960)
nyamuk yang dipelihara pada suhu optimum tanpa diberi makan dapat hidup selama tujuh hari, diberi makan larutan gula dapat hidup sampai 20 hari, dengan pemberian makan susu di campur gula dapat hidup sampai 19 hari, dengan pisang sampai 68 hari, sedangkan diberi makan dengan darah manusia mencapai umur 93 hari.
Dilaporkan oleh Gould et al. (1970) bahwa rata-rata lama hidup nyamuk A. aegypti dewasa di Malaysia adalah 3 – 6 minggu pada suhu 28 0C dan kelembaban nisbi antara 80% sampai 90%. Suhu yang tinggi yaitu 35 0C akan mengurangi umur nyamuk A. aegypti, sedangkan suhu rendah antara 15– 20 0C dengan kelembaban 90% akan memperpanjang lama hidupnya.
2.6. Peranan A. aegypti sebagai vektor penyakit.
Chen et al. (1993) di Taiwan mengatakan bahwa A. albopictus dan A. aegypti mempunyai kemampuan sebagai virus dengue 1 dan virus ini telah di isolasi dari A. aegypti yang terdapat di Taiwan sedangkan A. albopictus belum dilakukan (Lien et at, 1975). Kemudian dilanjutkan pula dengan percobaan menginfeksikan virus dengue 1 kepada nyamuk A. aegypti dan A. albopictus. Pada hari ke 7 (tujuh) positif terinfeksi dari 30 ekor A. albopictus dan 30 ekor A. aegypti masing-masing 3,3%. Pada hari ke 35,24% dari 28 A. albopictus positif terinfeksi dan pada hari ke 21 terdapat 100% dari 6 ekor nyamuk A. aegypti positif terinfeksi.
Selain virus dengue A. aegypti dapat pula terinfeksi oleh virus Sinbis. Virus Sinbis (Sin) adalah suatu alphavirus yang termasuk Famili Togoviridae (Westaway et al. 1985). Virus Sinbis ini dapat merupakan grup yang penting karena dapat menyebabkan penyakit ensefalitis pada manusia dan kuda (Jackson dan Johnson, 1987).
2.7. Toksikologi Insektisida.
Toksisitas pada suatu organisme selalu dinyatakan dalam istilah LD50 (lethal dose) yang berarti jumlah racun per unit berat organisme yang dibutuhkan untuk membunuh 50% populasi percobaan. Satuan dari LD50 dinyatakan dalam mg insektisida per Kg berat organisme. Pada kondisi bahan kimia/insektisida digunakan untuk serangga, maka LD50 dinyatakan dalam mikrogram insektisida per serangga ( μ g/serangga ).
Konsentrasi bahan kimia yang digunakan secara eksternal dapat membunuh 50% hewan dinamakan LC50 (lethal concentration). Nilai ini digunakan ketika dosis
yang pasti pada serangga tidak dapat ditentukan. Istilah LT50 (lethal time) adalah waktu yang dibutuhkan sehingga menyebabkan kematian 50% hewan percobaan pada dosis dan konsentrasi tertentu (Perry et al. 1998). Metode ini digunakan ketika jumlah hewan percobaan terbatas dan sering digunakan pada pengujian lapangan dimana sulit mengumpulkan jumlah serangga yang cukup untuk suatu pengujian. Pada kasus tertentu digunakan nilai KD50 (knockdown dose) dan KT50 (knockdown time).
Beberapa cara untuk melakukan pengujian pada serangga dan metode yang paling banyak digunakan adalah aplikasi topikal, karena insektisida dilarutkan dalam pelarut yang relatif tidak toksik seperti aseton dan larutan yang dihasilkan diteteskan pada permukaan tubuh serangga (Perry et al. 1998). Metode lain yaitu metode injeksi yang menggunakan jarum suntik yang halus terbuat dari baja tahan karat 20-30 gauge (diameter 0,41 atau 0,3 mm), yang membutuhkan gelas kecil untuk wadah insektisida yang dilarutkan dalam propiles glikol atau minyak kacang tanah dan injeksi dilakukan ke dalam rongga tubuh (intraperitoneal). Metode pencelupan digunakan ketika aplikasi topical dipandang tidak praktis untuk dilaksanakan.
Pengujian menggunakan metode kontak atau residu dengan cara insektisida dilarutkan dalam pelarut yang mudah menguap (volatile) kemudian dimasukkan pada kontainer gelas. Pelarut yang mengandung insektisida tersebut akan menguap dan ditampung dalam kontainer yang diputar-putar sehingga menghasilkan lapisan residu pada dinding gelas. Alternatf lain insektisida ditempatkan pada kertas saring, panel kayu atau jenis material bangunan lainnya dan dibiarkan mengering sebelum dipajankan pada serangga percobaan. Deposit residu insektisida tersebut dinyatakan sebagai miligram ramuan aktif per meter persegi (mg atau g AI/m2).
Insektisida Organofosfat. Menurut Foley (2005), fungsi sistem syarat adalah
perantaraan komunikasi antara sel syaraf dengan sel-sel lain dalam suatu organisme. Komunikasi diawali dengan melepaskan senyawa kimia yang disebut neurotransmiter dari pre sinap sel-sel syaraf. Senyawa ini berdifusi sepanjang sinap diantara sel-sel syaraf dan sel-sel pengontak (postsynaptic) dan berikatan pada protein reseptor di dalam membran sel. Ikatan tersebut menstimulasi perubahan saluran ion dalam membran yang difasilitasi oleh ion-ion spesifik (Na+, K+, Ca2+
atau Cl-) yang mengalir sepanjang membran bagian bawah dari gradien konsentrasinya masuk atau keluar dari sel. Gradien konsentrasi ini dapat memicu ataupun menghambat tergantung pada perubahan muatan ion di bagian dalam sel. Pada keadaan absennya neurotransmiter muatan di luar sel menjadi negatif. Pemicu neurotransmiter oleh sel terjadi melalui aliran ion Na+ dan dilipatgandakan melalui pembukaan tegangan listrik sensitif pada saluran Na+ disepanjang akson sel-sel syaraf. Aliran ini membukakan tegangan listrik sensitif saluran Ca2+ pada ujung sel syaraf. Aliran Ca2+ selanjutnya menstimulasi perbedaan neurotransmiter berikutnya yang menghasilkan kontraksi otot. Inhibisi neurotransmiter pada sel disebabkan oleh aliran ion K+ atau ion Cl- yang menghasilkan suatu sel lebih resisten dalam mendepolarisasika aliran Ca2+.
Asetilkolin adalah suatu neurotransmiter yang menstimulasikan pembukaan saluran Na+ dan K+ . Asetilkolin memberikan sinyal pada sinap yang diakhiri melalui suatu enzim asetilkolinesterase (AchE) yang berfungsi mengkatalisis reaksi hidroksil asetilkolin menjadi kolin tidak aktif dan asetat seperti ditunjukkan pada reaski berikut :
AChE
(CH3)NCH2CH2OCOCH3 CH3)3NCh2CH2OH + CH3CO2H
Asetilkolin kolin asam asetat
Enzim AchE merupakan kelompok serin esterase yang mengandung sisi aktif serin (Ser), histidin (His) dan residu asam amino glutamat (Glu) yang bersama-sama mengkatalisis reaksi hidrolisis asetilkolin. Ikatan H antara gugus Glu karboksilat dan N-1 pada histidin meningkatkan kemampuan N-3 His untuk bertindak sebagai basa yang menghilangkan H dari gugus hidroksil Ser. Hal ini menyebabkan oksigen pada Ser merupakan neukleofilik dan mampu menyerang gugus karboksil dari asetilkolin.
Organofosfat bekerja dengan cara menghambat enzim AchE, sehingga enzim ini tidak dapat menghidrolisis Ach. Senyawa orgaonfosfat secara luas telah digunakan menggantikan insektisida organoklorin. Senyawa ini merupakan ester atau derifat amida dari asam fosfat dan merupakan zat toksik untuk serangga maupun vertebrata melalui inhibisi enzim kolinesterase.
ENZIM-OH + Z-P-(O)(OR)2 ENZIM-O-P(O)(OR)2 + ZH Enzim kolinesterase Senyawa organofosfat enzim terinhibisi
Senyawa organofosfat diproduksi pada suhu tinggi (150 – 200 oC) sehingga pada umumnya mengandung isomer atau produk samping yang menyebabkan bau tidak enak (Perry et al. 1998).
Gejala keracunan pada serangga mengikuti pola umum dari peracunan syaraf, misalnya keresahan, hyperexitibility, gemetaran, kejang, lumpuh dan mati. Untuk mengetahui mekanisme aksi toksik senyawa organofosfat adalah interaksi enzim asetilkoline dengan asetilkolinesterase (AChE). Pertama-tama AChE menghasilkan kompleks E-OH – Ach (Asetilkolin – Enzim) yang merupakan intermediasi antara enzim dengan substrat kompleks tersebut bersifat reversibel yang membentuk asetilase AChE, selanjutnya asetil AChE dihidrolisis kembali menghasilkan AChE (Perry et al. 1998).
Reaksi tersebut sangat cepat sehingga tidak terjadi akumlasi asetilkolin sepanjang sinap atau pada sambungan neuromuscular. Bagian kolin kemudian dihilangkan dan asam asetat berkombinasi lagi (dengan bantuan enzim lain) membentuk asetilkolin dan siklus berulang.
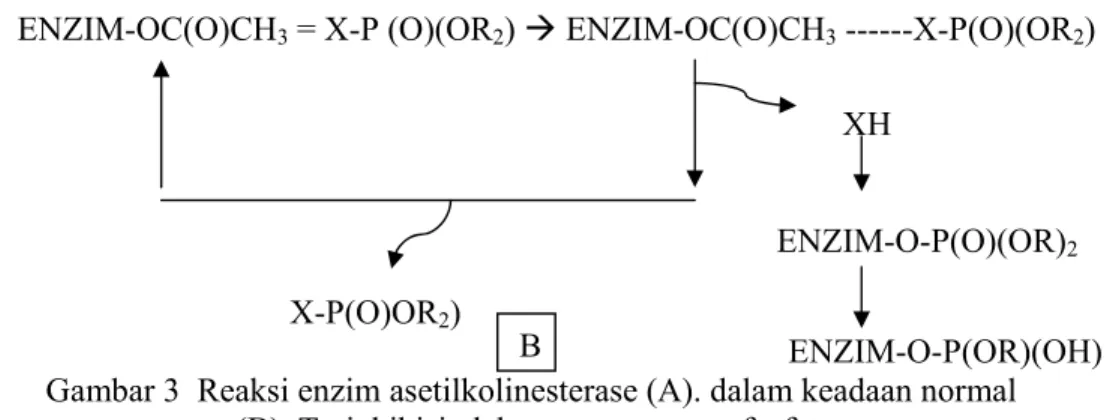
Insektisida organofosfat yang diaplikasikan pada serangga bereaksi dengan AChE pada kondisi yang sama. Tahap pertama enzim membentuk kompleks reversibel dengan senyawa organofosfat, kemudian kompleks putus menghasilkan organofosfat dan enzim yang terinhibisi (enzim fosforilate). Inhibisi tersebut menghasilkan fosforilat yang eksistensinya cukup sama dengan tahapan terakhir. Kemudian terjadi hidrolisis dan AChE dibebaskan dengan sebuah residu fosfat (sebagai dimetil asam fosfat). Tahapan hidrolisis tersebut sangat lambat dibanding dengan kondisi normal substrat asetilkolin sehingga enzim tidap dapat berfungsi secara efektif dengan peningkatan asetilkolin yang masuk ke sinap hasil akhir dari aktivitas syaraf. Mekanisme reaksi inhibisi tersebut ditunjukkan pada Gambar 3. ENZIM –OH++ (CH3)3N+CH2OC(O)CH3 ÆENZIM-OH ---(CH3)3N+CH2OC(O)CH3
(CH3)3N+CH2OC(O)CH3 HOC(O)CH3 ENZIM-OC(O)CH3
ENZIM-OC(O)CH3 = X-P (O)(OR2) Æ ENZIM-OC(O)CH3 ---X-P(O)(OR2) XH ENZIM-O-P(O)(OR)2 X-P(O)OR2) ENZIM-O-P(OR)(OH) Gambar 3 Reaksi enzim asetilkolinesterase (A). dalam keadaan normal
(B). Terinhibisi oleh senyawa organofosfat.
Insektisida Malation. Malation termasuk ke dalam golongan insektisida
organofosfat, yang pertama kali dibuat di Jerman pada tahun 1934 oleh Schrader. Insektisida ini termasuk jenis yang aman bagi mamalia dengan nilai LD50 oral akut 900-5800 mg/kg berat badan mempunyai tekanan uap 1.25 x 10-4 mm Hg pada suhu 20 oC. Malation berwujud cair, tidak berwarna dengan titik didih 156 – 157 oC, larut dalam hampir semua pelarut organik dan sedikit larut dalam air. Malation memiliki gugus karboksil yang menyebabkan insektisida ini mudah terhidrolisis dalam tubuh mamalia (Matsumura, 1975).
Malation merupakan insektisida organofosfat yang telah digunakan sejak tahun 1950 di Amerika Serikat. Pemakaian malation sebagai insektisida di Amerika Serikat mencapai 30 juta pound per tahun. Malation mulai digunakan di Indonesia sejak tahun 1972 dengan metode pengasapan (fogging) ( Suroso, 1991 dan Heodojo, 1993). Malation (O,O-dimetyl dithiophosphate of diethyl mercaptosuccinate) adalah insektisida yang digunakan untuk berbagai tanaman sayuran, buah dan lain-lain. Rumus empirik malation adalah C10H19O6PS2 dengan berat molekul 330,4 (Cox, 2003). Rumus bangun malation (gambar 4).
Gambar 4. Rumus bangun malation (www.google.com) B
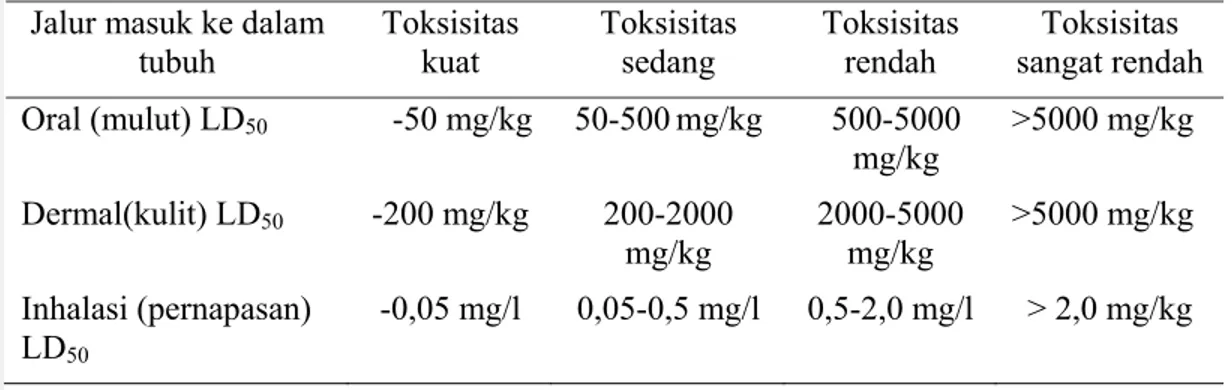
Malation membunuh serangga dengan cara dalam tubuh serangga malation diubah menjadi malaoxon yang menghambat enzim asetilkolinesterase. Tahun 2000 malation diduga sebagai bahan karsinogenik, tetapi tidak cukup berdampak karsinogenik terhadap manusia (Cox, 2003). Toksisitas malation dibagi menjadi empat jenis sesuai dengan dosisnya dan ditunjukkan pada Tabel 2 berikut.
Tabel 2 Empat kriteria toksisitas malation berdasarkan nilai LD50 Jalur masuk ke dalam
tubuh Toksisitas kuat Toksisitas sedang Toksisitas rendah Toksisitas sangat rendah Oral (mulut) LD50 -50 mg/kg 50-500 mg/kg 500-5000 mg/kg >5000 mg/kg Dermal(kulit) LD50 -200 mg/kg 200-2000 mg/kg 2000-5000 mg/kg >5000 mg/kg Inhalasi (pernapasan) LD50 -0,05 mg/l 0,05-0,5 mg/l 0,5-2,0 mg/l > 2,0 mg/kg