www.elsevier.com / locate / bres
Research report
Long-term reduction of benzodiazepine receptor density in the rat
cerebellum by acute seizures and kindling and its recovery 6 months
later by a pentylenetetrazole challenge
a ,
*
a b b bA.S. Bazyan
, V.V. Zhulin , M.N. Karpova , N.Y. Klishina , R.N. Glebov
a
Institute of Higher Nervous Activity and Neurophysiology, Russian Academy of Sciences, Butlerov St. 5A, GSP-7, Moscow 117865, Russia b
Institute of General Pathology and Pathophysiology, Russian Academy of Medical Sciences, Moscow, Russia Accepted 26 September 2000
Abstract
Seizures induced by an acute pentylenetetrazole (50 mg / kg) injection were accompanied by a long-term (at 1–48 h, but not on day 7)
3
decrease in the density (Bmax) of [ H]-diazepam binding to benzodiazepine receptors in rat cerebellar cortex with no change in affinity (K ). Kindling for 24 days by daily administrations of pentylenetetrazole (20 mg / kg) led to the same decrease in benzodiazepine receptord
density (at 1–48 h, but not on day 7) as that observed after a single dose of pentylenetetrazole (50 mg / kg). This suggests a common mechanism for both acute and kindling-induced seizures, dependent on the long-term receptor changes. The increased susceptibility to seizures persisted for 6 months after the termination of kindling, with BDZ receptor density in cerebellar cortex reduced almost by half. In
3
age-matched controls, an acute dose of PTZ (30 mg / kg) induced seizures and decrease in both Bmaxand K of [ H]-diazepam binding. Ind
kindled rats, at 6 months post-kindling, the same dose of PTZ (30 mg / kg) restored the benzodiazepine receptor density to the level found 6 months before, at the time of termination of kindling. Also, the severity of seizures was enhanced in the kindled rats. The results are discussed in terms of a balance of inhibitory and excitatory processes, in which the reduced BDZ receptor density at 6 months post-kindling may represent a compensatory reaction to outbalance some alterations in excitatory systems that have been reported to be induced by kindling. 2001 Elsevier Science B.V. All rights reserved.
Keywords: Kindling; Cerebellum; Pentylenetetrazole; Benzodiazepine; GABA
36 2
1. Introduction BDZ receptors and GABA-stimulated Cl influx into
cortical membrane preparations [29,40,49–52,55]. BDZ It is known that GABA , benzodiazepine (BDZ), pic-A agonists manifest a marked anticonvulsive effect on
2
rotoxin-barbiturate (PCT-BB) receptors and Cl channels clonic–tonic seizures in PTZ-kindled rats, with a marked are assembled in a single supramolecular complex. Both dose-effect dependence [3,5,13,34,53,54].
BDZ and PCT-BB receptors act as allosteric sites for the Chronic subthreshold stimulation of some brain struc-GABA receptor and vice versa. Pentylenetetrazole (PTZ)A tures, or chronic subconvulsive doses of convulsants result interacts with the PCT-BB receptor of the GABA / BDZ /A in kindling which is maintained for a long time PCT-BB receptor complex, inhibiting the GABAA and [18,29,42,67]. It is known that electrical kindling develops
2
BDZ receptors, closing the Cl channels and provoking with the active involvement of long-term potentiation-seizures in convulsive doses [5,11,12,25,35,41,46,47, induced NMDA receptors [10,26,37,38,48,61]. While the
50,55,65,69]. long-term suppression of the GABA / BDZ / PCT-BB re-A
In subconvulsive doses, chronic administration of PTZ ceptor activity by kindling has been described in a number induces kindling phenomena and modifies the GABA /A of studies, little is known about the initial long-term BDZ / PCT-BB receptor complex, suppressing GABA ,A responses. Is the long-term response attributable to kindl-ing development, or initial attribute of GABA / BDZ /A
PCT-BB receptors complex, as for NMDA receptors?
*Corresponding author. Tel.:17-95-334-7789; fax:17-95-338-8500.
E-mail address: bazyan@mail.ru (A.S. Bazyan). Similarly, little is known of the GABA / BDZ / PCT-BBA
mechanisms in kindling seizures retrieval after very long 2.2.1. Three to four-months-old rats
times of acute administration of subconvulsive doses of The first series comprised the rats, in which a convulsive
convulsants. dose of PTZ (50 mg / kg) resulted in seizure scores of 4–5
3
Previously, we have found [29] that changes in [ H]- points. They were decapitated 60 min (30 min after the diazepam binding are maximal in the cerebellar cortex, termination of seizures) or 48 h later and on day 7 after the rather than in the neocortex, of the kindled rats. It has been injection (n536, including control rats).
shown that 81% of the total BDZ receptors in cerebellar The second series comprised the rats, in which a cortex contain high-affinity sites for PTZ [6]. In the subconvulsive dose of PTZ (20 mg / kg) elicited kindled forebrain, only 34% of the total BDZ receptor population seizures scoring 4–5 points, followed by decapitation 60 bear high-affinity sites for PTZ. At the same time, the min (30 min after the termination of seizures) or 48 h later cerebellar cortex actualizes movement control. The cere- and on day 7 after the last injection (n548, including bellar cortex exerts its influence by inhibiting and dis- control rats).
inhibiting motor control action, in the premotor networks The third series comprised the rats endogenously sensi-of the brainstem, sensomotor cortex, and spinal cord [19– tive and insensitive to the convulsant. One group of 22,24]. On the other hand, the cerebellum and its associ- animals (n525) was treated with 25 mg / kg PTZ and the ated brainstem circuitry does appear to be essential (neces- second group with 30 mg / kg (n525). Then, the rats were sary and sufficient) for learning and memory of the selected according to their sensitivity: the first subgroup conditioned response [32,33,62–64,68]. This indicates that (n56) with seizures (2 points) provoked by 25 mg / kg long-term reactions are inherent to the neurons of cere- PTZ; and the second subgroup (n56) with no seizures bellum; for instance, in long-term depression [23]. provoked by 30 mg / kg PTZ. Both groups were decapitated
The goal of the present investigation was to study on day 7 after the injection.
3
long-term characteristics of [ H]-diazepam binding in
cerebellar cortex after an acute injection of PTZ in 2.2.2. After 6 months (about 10-months-old rats)
convulsive doses, after kindling development; peculiarities For the period of 6 months both the control and kindled
3
of the retrieval of kindled seizures and of [ H]-diazepam rats were kept in the breeding facility. Their weights at the binding 6 months after the termination of kindling, follow- second stage of experiments reached 360–450 g.
ing a challenge with a subconvulsive dose of PTZ. At the second stage, the persistence of kindling was studied. Two control groups and two groups of kindled rats were treated with 20 and 30 mg / kg PTZ (n525 in each
2. Materials and methods control and kindled group).
3
For the study of [ H]-diazepam binding four groups of Male Wistar rats (150–200 g, about 3 months old) were animals were used (n56 in each group): (1) control rats, used, a total of 266 animals. Five series of experiments no PTZ challenge; (2) acute seizures control rats, 30 min were conducted in two stages. Pentylenetetrazole (Sigma, after the termination of acute seizures (2–3 points) induced USA) or saline (control rats) were injected intraperitoneal- by a PTZ (30 mg / kg) challenge; (3) kindled control rats, ly. The seizures were scored by the following scale: (1) a no PTZ challenge, but with a history of seizures (4–5 twitch of the head; (2) occasional clonic seizures; (3) a points) 6 months ago; (4) kindling1PTZ challenge, 30 min series of clonic seizures, or clonus of the forepaws; (4) after the termination of seizures (3–4 points) induced by a clonic–tonic seizures while standing on the hind paws PTZ (30 mg / kg) challenge.
(kangaroo posture); (5) clonic–tonic seizures and falling
3
on one side. 2.3. [ H]-diazepam binding
2.1. Kindling procedure Cerebellar cortex was removed and used to isolate a
crude fraction of synaptic membranes. The cerebellar A subconvulsive dose of PTZ (20 mg / kg) was injected cortex was homogenized and centrifuged for 10 min at in rats (n5100) once daily for 24 days. The daily 10003g in the medium: 0.32 M sucrose, 1 mM EDTA and injections gradually increased the sensitivity to the convul- 5 mM Tris–HCl, pH 7.4 at 48C. The supernatant was sant. The dose of PTZ (20 mg / kg), which was subconvul- centrifuged for 20 min at 20,0003g. The pellet (P ) was2
sive in itself, evoked seizures that intensified and finalized frozen at 2408C. After washing by centrifugation, the
in a clonic–tonic paroxysm. pellet was resuspended in 4 ml of 50 mM Tris–HCl buffer,
pH 7.4. A portion of the suspension of synaptic
mem-2.2. Preselection of animals branes (400 ml) was added to 50 ml of an aqueous
3
sam-ples were run in duplicate. The samsam-ples were incubated for 40 min at 48C and filtered through a GF / B filter (What-man, UK), followed by four successive washes with ice-cold Tris-buffer. Radioactivity of the filters was measured in Bray’s scintillation cocktail using a Rackbeta (LKB, Sweden) counter. Protein content was determined by the method of Lowry et al. [39]. The density (Bmax) and the dissociation constant (K ) were calculated from the Scat-d
chard plots (M6S.E.M.). For statistical analysis, Student’s unpaired t-test (Tables), two-way (Fig. 2A) and one-way (Fig. 2B) ANOVA test were used.
3. Results
Fig. 1. Time course of kindling by daily injections of a subconvulsive dose of pentylenetetrazole (20 mg / kg) for 24 days (n5100).
3.1. Three to four-months-old rats
3.1.1. Acute PTZ (50 mg /kg) injection 3.1.3. Study of sensitivity
Acute treatment with PTZ (50 mg / kg) resulted in Kindled rats were treated as a PTZ-sensitive group, and
3
seizures scoring 4–5 points. The density (Bmax) of [ H]- control rats as a PTZ-resistant group. A lack of response diazepam binding sites was significantly reduced by 15% on day 7 after the seizures induced either by kindling or by at 30 min after the termination of seizures and by 13% at acute PTZ (50 mg / kg) was used as a criterion to group the
3
48 h with no change in the K (Table 1). No significantd rats according to their sensitivity. No differences in [
H]-changes were found on day 7. diazepam binding between the sensitive and insensitive
rats were found on day 7 after the selection (Table 2).
3
3.1.2. [ H]-diazepam binding after the kindling
Initially, a subconvulsive dose of PTZ (20 mg / kg) did 3.2. Ten-months-old rats, 6 months after the termination
3
not induce seizures and [ H]-diazepam binding was un- of kindling changed (Table 1). Daily injections of this dose resulted in
the development of kindling (Fig. 1) and a significant 3.2.1. Seizures retrieval
3
decrease in the Bmax of [ H]-diazepam binding by 19% at The kindling response of high sensitivity to low doses of 30 min and by 16% at 48 h after the termination of PTZ was preserved through the 6-months rest period after seizures. The K of the binding was unchanged. On day 7,d the kindling treatment, but not completely and with some no significant changes were observed (Table 1). These attenuation. A subconvulsive dose of PTZ (20 mg / kg) findings in the kindled rats are similar to the results found induced no seizures in the control rats (Fig. 2A), but this after an acute injection of the convulsive dose. dose elicited seizures in 60% of the kindled rats (1–2
Table 1 3
[ H]-Diazepam binding by cerebellar cortex membranes of 4 months old rats at 30 min, 48 h and 7 days after the termination of seizures at the acute pentylenetetrazole administration and kindling development
30 min 48 h Day 7
Control (n56) PTZ (n56) Control (n56) PTZ (n56) Control (n56) PTZ (n56) Pentylenetetrazole acute administration, 50 mg / kg
Bmax 1386697 1123676* 1360695 1142679* 1328690 1302689
(fmol / mg protein)
K (nM)d 9.0261.08 8.8561.06 9.1060.82 8.9260.80 9.1560.85 9.0960.78
Pentylenetetrazole acute administration, 20 mg / kg
Bmax 1142697 1127694 – – – –
(fmol / mg protein)
K (nM)d 8.2960.84 8.3560.79 – – – –
Kindling, 20 mg / kg
Bmax 1109689 898663* 1156672 971668* 1130667 1160678
(fmol / mg protein)
K (nM)d 8.0360.88 7.9660.80 7.8860.79 7.9460.87 8.1260.74 8.4160.92
*
Table 2 before, neither in control with acute seizures nor in kindled 3
[ H]-Diazepam binding by cerebellar cortex membranes of rats with rats. different sensibility to pentylenetetrazole (day 7 after test)
3
Animals Bmax(fmol / mg protein) K (nM)d 3.2.2. [ H]-diazepam binding at seizures retrieval
3
Insensitive (n56) 923684 7.7860.81 [ H]-diazepam binding was unaltered in the ten months
Sensitive (n56) 958637 7.6560.35 old control rats, but an acute PTZ challenge (30 mg / kg)
resulted in seizures (scores 2–3 points) that were
accom-3
points). At a higher dose of PTZ (30 mg / kg) seizures were panied by a decrease in both indices of [ H]-diazepam observed in 56% of the control rats (maximal scores of binding: the Bmax to 66%, and the K to 73% (Table 3).d
2–3 points) and in 100% of the kindled rats (maximal In the kindled control rats with a history of seizures
3
scores of 3–4 points). These observations suggest that the (4–5 points) 6 months before, the Bmax of [ H]-diazepam high sensitivity of rats to PTZ, induced by kindling, was binding was reduced to 54% without a PTZ challenge.
3
attenuated after 6 months, but at the same time the kindled After a rechallenge with PTZ, the Bmax of [ H]-diazepam rats were more sensitive than the control rats. binding was found to be enhanced to 78%, still being The pattern of seizures in the control and kindled rats significantly lower than in the control rats, with no change challenged with 30 mg / kg of PTZ was different (Fig. 2B). in the Kd (Table 3). This paradoxical finding can be In the control rats, the first peak of seizures occurred at 5 logically explained as shown in Fig. 3. At the time of min and the second peak was observed at 20 min after the termination of kindling the BDZ receptor density is PTZ challenge. About 40% of the kindled rats displayed a reduced to 81% versus the control 4-months-old rats. After period of low-level seizures scoring 1–3 points, which a rest period of 6 months, there was a decrease in BDZ lasted for 10 min. The first peak of seizures with maximal receptor density to 54% in the kindled rats without a PTZ scores from 3 to 4 points was observed at 20 min after the challenge. Acute PTZ administration to the kindled rats PTZ challenge, and the second peak occurred at 35 min. induced seizures and partially restored the BDZ receptor About 60% of the kindled rats displayed only the low-level density, just to the level of BDZ density found in the seizures scoring from 1 to 3 points, which lasted for 45 kindled 4-months-old rats (78%), which was established 6 min. This type of latency was not observed 6 months months before. At the same tame, the K of BDZ receptord
Fig. 2. Pattern of seizures in 10-months-old control and kindled rats. A. Seizures induced by 20 (ncont525; nkind525) and 30 (ncont525; nkind525) mg / kg x
pentylenetetrazole, averaged by maximal scores. *PF,0.05 versus own control, PF,0.05 versus own control and kindled rats at 20 mg / kg PTZ. B. Time course of pentylenetetrazole-induced seizures at 30 mg / kg (ncont514; nkind525). *PF,0.05.
Table 3
3
Effects of acute pentylenetetrazole (30 mg / kg) administration on [ H]-diazepam binding by cerebellar cortex membranes of 10 months old control and a
kindled rats
Control Control1PTZ Kindled control Kindled1PTZ
(n56) (acute seizures, n56) (n56) (rechallenge, n56)
?
Bmax 967679 6416106* 518681** 752680*
(fmol / mg protein)
K (nM)d 10.361.02 7.5660.99* 9.5460.78 10.761.21
a ?
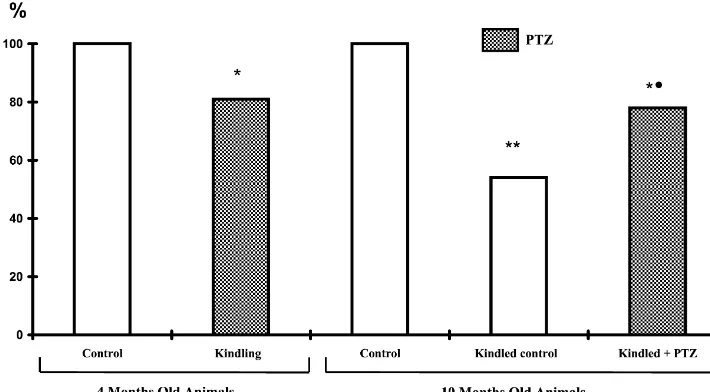
Fig. 3. Depression of benzodiazepine receptor density (Bmax) in 4- and 10-months-old kindled rats. The results are expressed as % versus control rats. 3
[ H]-Diazepam binding was measured using synaptic membranes isolated at the following time points. 4-months-old rats. At 30 min after the last kindled seizures induced by daily pentylenetetrazole (20 mg / kg) injections. 10-months-old rats. Kindled control: at 6 months after kindling with no pentylenetetrazole challenge; kindled1pentylenetetrazole: at 30 min after seizures induced by a pentylenetetrazole challenge (30 mg / kg). Vide statistics in
?
Table 1: *Pt,0.05 versus control; **Pt,0.01 versus control; Pt,0.05 versus kindled control.
binding was unchanged in the kindled rats, whereas in the role is played by intracellular reactions that follow the control 10-months-old rats that had seizures after an acute receptor occupation. It is known that the activity of BDZ PTZ challenge the K was significantly altered (Table 3).d receptors is dependent on protein kinase C [27,44]. Then, a feedback mechanism is suggested: phosphorylation or rephosphorylation of BDZ receptors by a BDZ
receptor-4. Discussion specific protein kinase C. This reaction can be termed
autocontrol feedback reaction. It is possible that autocon-We found that seizures of the same severity, but induced trol feedback reaction is modulated by the BDZ receptor by two different procedures, a single convulsive dose of and modifies its density. Furthermore, BDZ receptor-de-PTZ (50 mg / kg) and kindling with a subconvulsive dose pendent protein kinase C mechanism is induced by gene (20 mg / kg), resulted in similar modifications of BDZ expression [27,44]. It is possible that gene expression can receptors of GABA supramolecullar receptor complex: aA explain the long-term (up to 48 h) decrease of BDZ long-term (48 h) decrease in the Bmax, with no change in receptor density in the cerebellum by acute seizures. the K , and subsequent normalization on day 7. This resultd Extending this logical way, we can suggest that the shows that kindling is an elaboration of high sensitivity to establishment of kindling is based on the consecutive subconvulsive doses of PTZ. This result also shows that enhancement of protein kinase C-dependent reaction and
3
the characteristics of [ H]-diazepam binding to the BDZ modification of BDZ receptors. It is possible that on day 7 cerebellar receptors of animals with control levels of after the establishment of kindling the BDZ receptor sensitivity was not different from the characteristics of density was normalized, but intracellular processes still kindled animals with high level of sensitivity 7 days after continued. We may suggest, therefore, that the decrease in the termination of seizures. Normalization of the reaction BDZ receptor density 6 months after kindling is the track on day 7 makes it possible to suggest initial binding of continuing intracellular processes.
constants of cerebellum BDZ receptors. A special in- The high sensitivity induced by kindling was somewhat vestigation of the two types of control animals, endoge- attenuated after 6 months. This is a typical process for nously sensitive and resistant to PTZ, showed that they elaborated or learned reactions that attenuate with time also were similar 7 days after the termination of seizures. without reminder. It is known [59–61] that reminder
3
autocontrol feedback mechanisms which in this situation is Our results show that kindling is nevertheless preserved manifested as autoenhancement feedback mechanisms, and at a high sensitivity for as long as 6 months. At the same would retrieve seizures and BDZ receptor constants. time, the BDZ receptor response was qualitatively different But, the theoretical view that seizures are determined by from the seizures induced by acute PTZ treatment of the relationship between excitatory and inhibitory pro- young and old control animals; in that in young rats only cesses is supported by a number of experimental data the density of BDZ receptors was reduced, while in older [7,8,14,18,42,58]. Our primary interest in pursuing this rats both the density and affinity of BDZ receptors was work was initially directed at BDZ receptors, on the altered.
assumption that long-term responses are more likely to At 7 days after the termination of kindling the BDZ develop at allosteric modulatory sites of the GABA -A receptor density was normalized, but 6 months later the receptor complex. This assumption appears to be justified, BDZ receptor density was about two-fold lower. These
2
considering that GABA , BDZ receptors and ClA channels results correspond with the literature. A normalization of are allosterically inhibited by PTZ and that 81% of the NMDA receptor activity on day 7 was found in amygdala total BDZ receptors in the cerebellum contain high-affinity kindling [2], and normalization of GABAA [30] and sites for PTZ [6]. Accordingly, we feel that the changes in NMDA [48] receptor subunit mRNA level was observed BDZ binding can be interpreted as representative for the on day 5 after hippocampal kindling. Some time after the whole GABAA supramolecular complex in the cerebellar normalization, the receptor response is spontaneously cortex during seizures and kindling. altered again: a decrease in GABAA receptor sensitivity We found that seizures of the same severity, but induced was reported at 3–4 weeks after amygdala kindling by two different procedures, a single convulsive dose of [15,16]. An enhanced hippocampal NMDA receptor activi-PTZ (50 mg / kg) and kindling with a subconvulsive dose ty was observed 1 month after kindling [71], and a novel (20 mg / kg), resulted in similar modifications of BDZ population of NMDA receptors was expressed 28 days receptors of GABA supramolecullar receptor complex: aA after hippocampal kindling [31]. In addition, 6 weeks after
21
long-term (48 h) decrease in the Bmax, with no change in kindling an enhancement of the Ca current was observed the K , and subsequent normalization on day 7. It is knownd in the hippocampal CA area of the rat [70].1
[15,16,66] that kindling can lead to a long-term decrease of However, these data are not always confirmed in some the GABAA receptor complex activity, due to changes in reports. No changes of NMDA receptor function were the synthesis of the respective proteins [30,36]. A series of found on day 28 [2] and 3 and 6 weeks [26] after the 40 kindling-induced seizures (by rapid hippocampal stimu- kindling, as well as of GABAA receptor activity [28] 4 lation) led to biphasic alterations of GABAA receptor weeks after the kindling.
subunit mRNA levels in dentate gyrus with only minor A review of the literature allows us to suggest that the changes in CA –CA [30]. Up to 4 h after the last seizure1 3 decrease in BDZ binding observed 6 months after the mRNA expression for alpha 1 subunit was slightly de- kindling in our study may be unrelated to the mechanism creased in dentate gyrus, whereas marked reductions were of kindling storage and seizure retrieval. It is possible that observed for beta 3 and gamma 2 subunits (by 41 and the decrease in GABA / BDZ / PCT-BB receptor complexA
48%, respectively). Between 12 and 48 h there were major activity represents a compensatory reaction to a reduced increases of alpha 1 (by 59%) and gamma 2 (by 35%) excitatory system activity, since a decrease in NMDA subunits mRNA levels but no significant changes of beta 3 receptor activity has been reported 1 and 1.5 months after subunit mRNA expression. The subunits mRNA levels had kindling [43,45]. We think that only a compensatory returned to control values after 5 days [30]. These results reaction of excitatory processes is able to explain the are similar to ours: the biphasic changes of GABAA absence of seizures against the background of a one-half receptor subunits may be related to the onset of normaliza- decrease of the inhibitory system in our kindled control tion of the receptor density. 10-months-old rats. It is possible that spontaneous changes The long-term response induced by acute PTZ treatment of the receptor activity can arise after the termination of is presumably maintained by gene expression. Various long-term convulsant treatment or electrostimulation. But types of long-term responses are known to be maintained the latency of these spontaneous changes in the receptor by gene expression [4,17,56,57]. Therefore, we think that activity seems to depend on experimental conditions. kindling develops because of a long-term response of Our results suggest other mechanisms of seizure retriev-GABA / BDZ / PCT-BB receptor complex. It is possibleA al 6 months after the kindling. An acute administration of that small (by acute subconvulsive dose PTZ injection and PTZ (30 mg / kg) to 10-months-old kindled control rats not elicited experimentally) long-term decreases of enhances the activity of GABA / BDZ / PCT-BB receptorA
GABAA receptor complex activity are cumulated to a complex, increasing it to the level of 4-months-old kindled convulsive threshold in the course of the daily injections rats as well as producing seizures. In this case, seizures and are consolidated by gene expression. The presumed might be initiated by a simultaneous increase of excitatory time course of this cumulative process is reflected in Fig. system activity, bringing it to the level of 4-months-old
excitatory and inhibitory systems of the brain, establishing References
a new balance between them and consolidating it via gene
[1] K. Akiyama, A. Daigen, N. Yamada, T. Itoh, I. Kohira, H. Ujike, S.
expression (basic mechanism of memory consolidation).
Otsuki, Long-lasting enhancement of metabotropic excitatory amino
At 6 months after the kindling, a subconvulsive PTZ
acid receptor-mediated polyphosphoinositide hydrolysis in the
challenge induces seizures by recovering a new consoli- amygdala / pyriform cortex of deep prepiriform cortical kindled rats, dated receptor activity. It is a newly consolidated activity, Brain Res. 569 (1992) 71–77.
because in 10-months-old intact control rats the seizures [2] K. Akiyama, Y. Yoneda, K. Ogita, T. Itoh, A. Daigen, I. Sora, I. Kohira, H. Ujike, S. Otsuki, Ionotropic excitatory amino acid
induced by a convulsive doze of PTZ (30 mg / kg) are
receptors in discrete brain regions of kindled rats, Brain Res. 587
accompanied by a qualitatively different type of BDZ
(1992) 73–82.
binding response, a decrease in both density and affinity of [3] A. Becker, G. Grecksch, H. Matthies, The influence of diazepam on the BDZ receptor binding. Simultaneous modification of learning processes impaired by pentylenetetrazole kindling, Naunyn
Schmiedebergs Arch. Pharmacol. 349 (1994) 492–496.
excitatory and inhibitory systems by kindling has been
[4] J.E. Biewenga, L.H. Schrama, W.H. Gispen, Presynaptic
phos-consistently found in a number of studies [7–
phoprotein B-50 / GAP-43 in neuronal and synaptic plasticity, Acta
9,14,18,42,50,51,58]. Biochim. Pol. 43 (1996) 327–338.
When a new consolidated receptor density is attained by [5] Y.F. Chang, X.M. Gao,L-lysine is a barbiturate-like anticonvulsant
acute PTZ administration to 10-months-old kindled rats, it and modulator of the benzodiazepine receptor, Neurochem. Res. 20 (1995) 931–937.
is accompanied by an autoenhancement of seizures (Fig. 2
[6] A.Y. Chwen, E.A. Swinyard, H.H. Wolf, Pentylenetetrazole may
B). In the first 5 min, no differences in seizure activity are
discriminate between different types of benzodiazepine receptors, J.
observed between the control and kindled rats. Then, the Neurochem. 41 (1983) 830–833.
mechanism of intercellular autoenhancement is switched [7] M.G. Corda, M. Orlandi, D. Lecca, O. Giorgi, Decrease in
GABA-on and the seizures are intensified. The functiGABA-on of ergic function induced by pentylenetetrazole kindling in rats: antagonism by MK-801, J. Pharmacol. Exp. Ther. 262 (1992)
autoenhancement is to accomplish autocontrol or
au-792–800.
tomodification of the GABA / BDZ / PCT-BB receptorA [8] M.G. Corda, M. Orlandi, D. Lecca, G. Carboni, V. Frau, O. Giorgi,
complex activity, initially suppressed by the long-term Pentylenetetrazole-induced kindling in rats: effect of GABA func-depression mechanism (Table 1). The alteration of BDZ tion inhibitors, Pharmacol. Biochem. Behav. 40 (1991) 329–333.
receptor conformation / density induced by PTZ binding to [9] S.I. Deutsch, C.H. Park, L.G. Lukacs, C. Morn, L. Koetzner, J. Mastropaolo, MK-801 alters the GABA receptor complex and
PCT-BB receptors is maintained for 48 h, when PCT-BB A
potentiates flurazepam’s antiseizure efficacy, Pharmacol. Biochem.
receptors are no longer occupied. These processes can
Behav. 51 (1995) 909–915.
proceed in three stages, the two last stages from it are [10] N. Durmuller, M. Craggs, B.S. Meldrum, The effect of the
non-intercellular: NMDA receptor antagonist GYKI 52466 and NBQX and the
(1) occupation and allosteric depression of the GABA /A competitive NMDA receptor antagonist D-CPPene on the develop-ment of amygdala kindling and on amygdala-kindled seizures,
BDZ / PCT-BB receptors density; (2) modification of
sec-Epilepsy Res. 17 (1994) 167–174.
ond messengers and protein kinase C-induced
phosphoryl-[11] F.J. Ehlert, ‘Invers agonists’, cooperativity and drug action at
ation / dephosphorylation of the GABA / BDZ / PCT-BBA benzodiazepine receptors, Trends Pharmacol. Sci. 7 (1986) 28–32.
receptor complex, thus maintaining the depression of [12] L.K. Friedman, T.T. Gibbs, D.H. Farb, Gamma-aminobutyric acid A
GABA / BDZ / PCT-BB receptor density; (3) modificationA receptor regulation: heterologous uncoupling of modulatory site interactions induced by chronic steroid, barbiturate, benzodiazepine,
of gene expression and inhibition of GABA / BDZ / PCT-A
or GABA treatment in culture, Brain Res. 707 (1996) 100–109.
BB receptor protein synthesis. A newly established level of
[13] M. Gasior, R.B. Carter, S.R. Goldberg, J.M. Witkin, Anticonvulsant
depression can replace the genetically determined level. and behavioral effects of neuroactive steroids alone and in conjunc-Normally, seizures would be observed at stages 2 and 3 of tion with diazepam, J. Pharmacol Exp. Ther. 282 (1997) 543–553.
the process. For instance, NMDA receptor activity was [14] O. Giorgi, M. Orlandi, M. Geic, M.G. Corda, MK-801 prevents the decrease in 35S-TBPS binding in the rat cerebral cortex induced by
found to be normalized at 7–28 days after amygdala and
pentylenetetrazole kindling, Brain Res. Bull. 27 (1991) 835–837.
hippocampal kindling [2]. At the same time, metabotropic
[15] T.D. Hernandez, D.W. Gallager, Development of long-term
subsen-excitatory amino acid receptor-stimulated polyphos- sitivity to GABA in dorsal raphe neurons of amygdala-kindled rats, phoinositide hydrolysis was enhanced for a long period of Brain Res. 582 (1992) 221–225.
time in amygdala- and hippocampal-kindled rats [1]. We [16] T.D. Hernandez, J.B. Rosen, D.W. Gallager, Long-term changes in sensitivity to GABA in dorsal raphe neurons following amygdala
suggest that in our experiments a new consolidated state of
kindling, Brain Res. 517 (1990) 294–300.
receptors is attained independently of the current state of
[17] A. Hicks, S. Davis, J. Rodger, A. Helme-Guizon, S. Laroche, J.
receptors. Mallet, Synapsin I and syntaxin 1B: key elements in the control of
neurotransmitter release are regulated by neuronal activation and long-term potentiation in vivo, Neuroscience 79 (1997) 329–340. [18] K. Hirayama, R. Murata, S. Matsuura, Effects of an N-methyl-D
-aspartate antagonist and a GABAergic antagonist on entorhinal Acknowledgements
tetanic responses during the early stages of amygdala kindling in rats, Neurosci. Res. 19 (1994) 397–405.
This work was supported by the Russian-Netherlands [19] J.C. Houk, J.T. Buckingham, A.G. Barto, Models of the cerebellum
[20] M. Ito, Neurons of Cerebellar Nuclei, in: M.A.B. Brazier (Ed.), The Ferrendelli, Physiological comparison of alpha-ethyl-alpha-methyl-gamma-thiobutyrolactone with benzodiazepine and barbiturate Interneuron, UCLA Forum, 1969, pp. 309–327.
modulators of GABA receptors, Neuropharmacology 35 (1996) [21] M. Ito, Neurophysiological aspects of the cerebellar motor control A
123–136. system, Int. J. Neurobiol. 7 (1970) 162–176.
[42] L.R. Merlin, R.K. Wong, Synaptic modifications accompanying [22] M. Ito, in: The Cerebellum and Neuronal Control, Raven Press, New
epileptogenesis in vitro: long-term depression of GABA-mediated York, 1984, 580 pp.
inhibition, Brain Res. 627 (1993) 330–340. [23] M. Ito, Long-term depression, Ann. Rev. Neurosci. 12 (1989)
[43] J.V. Nadler, M.A. Thompson, J.O. McNamara, Kindling reduces 85–102.
sensitivity of CA3 hippocampal pyramidal cells to competitive [24] M. Ito, Movement and thought: identical control mechanisms by the
NMDA receptor antagonists, Neuropharmacology 33 (1994) 147– cerebellum, Trends Neurosci. 16 (1993) 448–450.
153. [25] T. Ito, T. Suzuki, S.E. Wellman, I.K. Ho, Chronic pentobarbital
[44] Y. Nomura, Y. Kitamura, T. Ohnuki, T. Arima, Y. Yamanaka, K. administration alters gamma-aminobutyric acid A receptor alpha
Sasaki, Y. Oomura, Alterations in acetylcholine, NMDA, benzo-3
6-subunit mRNA levels and diazepam-insensitive [ H]Ro15-4513
diazepine receptors and protein kinase C in the brain of the binding, Synapse 22 (1996) 106–113.
senescence-accelerated mouse: an animal model useful for studies [26] S.M. Jones, K.M. Johnson, Effects of amygdaloid kindling on
on cognitive enhances, Behav. Brain Res. 83 (1997) 51–55. NMDA receptor function and regulation, Exp. Neurol. 106 (1989)
[45] M.M. Okazaki, J.O. McNamara, J.V. Nadler, N-methyl-D-aspartate 52–60. receptor autoradiography in rat brain after angular bundle kindling, [27] J.D. Johnston, S.A. Price, D.R. Bristow, Flunitrazepam rapidly Brain Res. 482 (1989) 359–364.
reduces GABA(A) receptor subunit protein expression via a protein [46] W.R. Olsen, GABA–benzodiazepine–barbiturate receptor interac-kinase C-dependent mechanism, Br. J. Pharmacol. 124 (1998) tions, J. Neurochem. 35 (1981) 1–13.
1338–1340. [47] R.W. Olsen, J. Yang, R.G. King, A. Dilber, G.B. Stauber, R.W. [28] W. Kamphuis, T.C. De Rijk, F.H. Lopes da Silva, GABA receptorA Ransom, Barbiturate and benzodiazepine modulation of GABA
beta 1–3 subunit gene expression in the hippocampus of kindled receptor binding and function, Life Sci. 39 (1986) 1969–1976. rats, Neurosci. Lett. 174 (1994) 5–8. [48] G.D. Pratt, M. Kokaia, J. Bengzon, Z. Kokaia, J.M. Fritschy, H. [29] M.N. Karpova, V.V. Zhulin, R.N. Glebov, O.Yu. Pankov, R.I. Mohler, O. Lindvall, Differential regulation of N-methyl-D-aspartate
3
Kruglikov, Binding of H-diazepam in rats brain 11 months after receptor subunit messenger RNAs in kindling-induced epilep-korazol kindling, Bull. Exp. Biol. Med. 117 (1994) 135–157. togenesis, Neuroscience 57 (1993) 307–318.
[30] M. Kokaia, G.D. Pratt, E. Elmer, J. Bengzon, J.M. Fritschy, Z. [49] C. Psarropoulou, N. Matsokis, F. Angelatou, G. Kostopoulos, Kokaia, O. Lindvall, H. Mohler, Biphasic differential changes of Pentylenetetrazole-induced seizures decrease gamma-aminobutyric GABAA receptor subunit mRNA levels in dentate gyrus granule acid-mediated recurrent inhibition and enhance adenosine-mediated cells following recurrent kindling-induced seizures, Mol. Brain Res. depression, Epilepsia 35 (1994) 12–19.
23 (1994) 323–332. [50] L. Rocha, R.F. Ackermann, J. Engel Jr., Chronic and single [31] J.E. Kraus, G.C. Yeh, D.W. Bonhaus, J.V. Nadler, J.O. McNamara, administration of pentylenetetrazole modifies benzodiazepine re-Kindling induces the long-lasting expression of a novel population ceptor-binding: an autoradiographic study, Epilepsy Res. 24 (1996) of NMDA receptors in hippocampal region CA3, J. Neurosci. 14 65–72.
(1994) 4196–4205. [51] L. Rocha, M. Briones, R.F. Ackermann, B. Anton, N.T. Maidment, [32] D.J. Krupa, J.K. Thompson, R.F. Thompson, Localization of a C.J. Evans, J. Engel Jr., Pentylenetetrazole-induced kindling: early memory trace in the mammalian brain, Science 260 (1993) 989– involvement of excitatory and inhibitory systems, Epilepsy Res. 26
991. (1996) 105–113.
[33] D.G. Lavond, J.J. Kim, R.F. Thompson, Mammalian brain substrates [52] M.A. Rogawski, R.G. Porter, Antiepileptic drugs: pharmacological of aversive classical conditioning, Annu. Rev. Psychol. 44 (1993) mechanisms and clinical efficacy with consideration of promising
317–342. developmental stage compounds, Pharmacol. Rev. 42 (1990) 223–
[34] M. Lazarova-Bakarova, V.V. Petkov, M. Genkova-Papasova, Effect 286.
of diazepam and medazepam on pentylenetetrazole kindling in [53] H.C. Rosenberg, Differential expression of benzodiazepine anticon-albino rats, Acta Physiol. Pharmacol. Bulg. 16 (1990) 37–41. vulsant cross-tolerance according to time following flurazepam or [35] E. Lewin, J. Peris, V. Bleck, N.R. Zahniser, R.A. Harris, Diazepam diazepam treatment, Pharmacol. Biochem. Behav. 51 (1995) 363–
36 2
sensitizes mice to FG 7142 and reduces muscimol-stimulated Cl 368.
flux, Pharmacol. Biochem. Behav. 33 (1989) 465–468. [54] C. Rundfeldt, P. Wlaz, D. Honack, W. Loscher, Anticonvulsant [36] E. Lewin, V. Bleck, J.E. Dildy-Mayfield, R.A. Harris, GABA andA tolerance and withdrawal characteristics of benzodiazepine receptor glutamate receptor subunit mRNAs in cortex of mice chemically ligands in different seizure models in mice. Comparison of kindled with FG 7142, Mol. Brain Res. 22 (1994) 320–322. diazepam, bretazenil and abecarnil, J. Pharmacol. Exp. Ther. 275 [37] W. Loscher, D. Honack, Effects of the competitive NMDA receptor (1995) 693–702.
antagonist, CGP 37849, on anticonvulsant activity and adverse [55] L. Schmid, M. Bottlaender, E. Brouillet, C. Fuseau, M. Maziere, effects of valproate in amygdala-kindled rats, Eur. J. Pharmacol. 234 Vigabatrin modulates benzodiazepine receptor activity in vivo: a (1993) 237–245. positron emission tomography study in baboon, J. Pharmacol. Exp. [38] W. Loscher, C. Rundfeldt, D. Honack, Low doses of NMDA Ther. 276 (1996) 977–983.
receptor antagonists synergistically increase the anticonvulsant [56] T. Smirnova, S. Laroche, M.L. Errington, A.A. Hicks, T.V. Bliss, J. effect of the AMPA receptor antagonist NBQX in the kindling Mallet, Transsynaptic expression of a presynaptic glutamate receptor model of epilepsy, Eur. J. Neurosci. 5 (1993) 1545–1550. during hippocampal long-term potentiation, Science 262 (1993) [39] O.H. Lowry, N.J. Rosebrough, A.L. Farr, R.J. Randall, Protein 433–436.
measurement with the Folin phenol reagent, J. Biol. Chem. 193 [57] P.K. Stanton, J.M. Sarvey, Blockade of norepinephrine-induced (1951) 256–274. long-lasting potentiation in the hippocampal dentate gyrus by an [40] R.J. Marley, C. Heninger, T.D. Hernandez, D.W. Gallager, Chronic inhibitor of protein synthesis, Brain Res. 361 (1985) 276–283.
3
[59] M.J. Summers, S.F. Crowe, K.T. Ng, Administration of glutamate specific changes in the [ H]-muscimol binding in the rat hippocam-following a reminder induces transient memory loss in day-old pus: a quantitative autoradiographic study, Neuroscience 59 (1994) chicks, Cogn. Brain Res. 3 (1995) 1–8. 817–826.
[60] M.J. Summers, S.F. Crowe, K.T. Ng, Administration of lanthanum [67] M.N. Titulaer, W. Kamphuis, F.H. Lopes da Silva, Autoradiographic 35
cloride following a reminder induces a transient loss of memory analysis of [ S]t-butylbicyclophosphorothionate binding in kindled retrieval in day-old chicks, Cogn. Brain Res. 4 (1996) 109–119. rat hippocampus shows different changes in CA1 area and fascia [61] M.J. Summers, S.F. Crowe, K.T. Ng, Administration ofDL-2-amino- dentata, Neuroscience 66 (1995) 547–554.
5-phosphonovaleric acid (AP5) induces transient inhibition of [68] J.A. Tracy, J.K. Thompson, D.J. Krupa, R.F. Thompson, Evidence reminder-activated memory retrieval in day-old chicks, Cogn. Brain of plasticity in the pontocerebellar conditioned stimulus pathway Res. 5 (1997) 311–321. during classical conditioning of the eyeblink response in the rabbit, [62] R.F. Thompson, Neural mechanisms of classical conditioning in Behav. Neurosci. 112 (1998) 267–285.
mammals, Philos. Trans. R. Soc. Lond. B. Biol. Sci. 329 (1990) [69] C. Vale, A. Pomes, E. Rodriguez-Farre, C. Sunol, Allosteric
interac-161–170. tions between gamma-aminobutyric acid, benzodiazepine and
pic-[63] R.F. Thompson, S. Bao, L. Chen, B.D. Cipriano, J.S. Grethe, J.J. rotoxinin binding sites in primary cultures of cerebellar granule Kim, J.K. Thompson, J.A. Tracy, M.S. Weninger, D.J. Krupa, cells. Differential effects induced by gamma- and delta-hexachloro-Associative Learning, Int. Rev. Neurobiol. 41 (1997) 151–189. cyclohexane, Eur. J. Pharmacol. 319 (1997) 343–353.
[64] R.F. Thompson, J.K. Thompson, J.J. Kim, D.J. Krupa, P.G. Shin- [70] M. Vreugdenhil, W.J. Wadman, Kindling-induced long-lasting en-kman, The nature of reinforcement in cerebellar learning, Neurobiol. hancement of calcium current in hippocampal CA1 area of the rat: Learn. Mem. 70 (1998) 150–176. relation to calcium-dependent inactivation, Neuroscience 59 (1994) [65] M.K. Ticku, G. Maksay, Convulsant / depressant site of action at the 105–114.
allosteric benzodiazepine-GABA receptor-ionophore complex, Life [71] G.C. Yeh, D.W. Bonhaus, J.V. Nadler, J.O. McNamara, N-methyl-D -Sci. 33 (1983) 2363–2375. aspartate receptor plasticity in kindling: quantitative and qualitative [66] M.N. Titulaer, W. Kamphuis, C.W. Pool, J.J. van Heerikhuize, F.H. alterations in the N-methyl-D-aspartate receptor-channel complex,
![Table 2[ H]-Diazepam binding by cerebellar cortex membranes of rats with3](https://thumb-ap.123doks.com/thumbv2/123dok/3136664.1382210/4.612.49.549.659.720/table-diazepam-binding-cerebellar-cortex-membranes-rats-with.webp)