DOKUMENTASI
FIKSASI NITROGEN DAN ASOSIASI TANAMAN LEGUM
Oleh :
Anak Agung Istri Kesumadewi
NIP. 19681223 199303 2001
PROGRAM STUDI AGROEKOTEKNOLOGI
FAKULTAS PERTANIAN
UNIVERSITAS UDAYANA
2016
ii
DAFTAR ISI
Nomer Judul Halaman
DAFTAR TABEL ... iii
DAFTAR GAMBAR ... iv
I. PENDAHULUAN I ... 1
II. KEANEKARAGAMAN HAYATI MIKROBA DALAM SIKLUS NITROGEN ... 4
2.1. Fiksasi N2 dan Suplai Nitrogen oleh Tanaman Legum ... 9
2.2. Mineralisasi Nitrogen ...12
III. TRANSFER NITROGEN DARI TANAMAN LEGUM KE TANAMAN NON LEGUM .14 3.1. Mekanisme Transfer N Jangka Pendek ...15
3.2. Mekanisme Transfer N Jangka Panjang ...17
IV. ASOSIASI TANAMAN LEGUMINOSE ...20
4.1. Definisi dan Sistem Klasifikasi ...20
4.2. Aspek Positif Asosiasi Tanaman ...21
4.3. Interaksi dalam Asosiasi Tanaman ...22
4.4. Asosiasi Tanaman Legum ...23
V. KESIMPULAN ...25
iii
DAFTAR TABEL
Nomer Judul Halaman
1. Siklus Nitrogen : Alur Proses dan Mikroba yang Terlibat ... 6 2. Genus dan Spesies Rhizobium serta Tanaman Inang ...11 3. Kadar N di Bagian Akar dan Atas Tanaman Legum ...17
iv
DAFTAR GAMBAR
Nomer Judul Halaman
1. Siklus Nitrogen ... 5
2. Beberapa Contoh Organisme Perombak Bahan Organik ... 8
3. Contoh Komponen Sistem Penambatan N2 oleh Rhizobium ...10
4. Transfer N dari Tanaman Legum ke Tanaman Non Legum ...16
Tanaman leguminose merupakan salah satu komponen fungsional ekosistem daratan. Jenis tanaman tersebut merupakan bagian dari 3 besar kelompok tanaman berbunga. Leguminose mampu menghasilkan bahan organik dalam jumlah tinggi, mencegah bahaya erosi dan meningkatkan kesuburan tanah. Kemampuan tanaman leguminose memfiksasi nitrogen dari udara dapat meningkatkan kandungan N di dalam tanah. Leguminose dapat digunakan sebagai tanaman penutup tanah dan mempunyai fungsi konservasi tanah dan air serta menekan pertumbuhan gulma dan serangga hama. Budidaya campuran tanaman leguminose dengan tanaman pangan lainnya berpotensi untuk meningkatkan jumlah dan kualitas bahan kering.
Tanaman leguminose sangat menentukan produktivitas dan proses sinergis yang terjadi dalam suatu asosiasi tanaman karena leguminose umumnya tidak berkompetisi dalam memperoleh nitrogen. Sebaliknya, N2 yang
difiksasi oleh tanaman leguminose juga disumbangkan kepada tanaman lainnya yang berada di sekitarnya (Paynel et al., 2001; Trannin et al., 2000). Transfer Nitrogen yang dapat dilakukan oleh tanaman legum secara langsung maupun tidak langsung kepada tanaman lain di sekitarnya memungkinkan pertumbuhan tanaman yang berdekatan dengan legum untuk tumbuh dengan lebih baik. Hal tersebut dapat mendorong terjadinya asosiasi antar tanaman legum dan non legum. Asosiasi tersebut juga mempengaruhi keragaman mikroba di dalam tanah terutama yang berhubungan dengan siklus nitrogen karena keragaman
eksudat akar keragaman jenis serta kualitas serasah bahan organik yang dihasilkan dari kelompok tanaman yang berbeda.
Tanaman leguminose terdiri dari beragam spesies yang memiliki peranan dan nilai ekonomis berbeda. Beberapa jenis leguminose dibudidayakan untuk tujuan konservasi hutan, atau rehabilitasi lahan kritis, misalnya tanaman sengon dan akasia. Kedua tanaman tersebut dipilih karena mampu menghasilkan biomasssa kayu dengan cepat serta bernilai ekonomis. Beberapa kelompok lainnya digunakan sebagai tanaman peneduh, misalnya flamboyan. Tanaman leguminose yang tergolong perdu atau semak dapat digunakan sebagai penutup tanah atau bahan pupuk hijau, misalnya clover dan lucerne. Leguminose yang dapat digunakan sebagai bahan pangan memiliki nilai ekonomis yang lebih tinggi dibandingkan jenis lainnya. Beberapa jenis leguminose tersebut antara lain adalah kedelai, kacang hijau, kacang tanah, kacang polong, buncis, dan bangkuang.
Budidaya leguminose secara monokultur maupun tumpangsari dapat memberikan keuntungan ekologis dan ekonomis. Keuntungan ekologis terbesar adalah kemampuannya menambat N2 dan menyuplainya kepada tanaman yang
berada di sekitarnya. Tanaman leguminose pada kondisi tertentu dapat menyetimulir terbentuknya keanekaragaman hayati yang lebih tinggi melalui asosiasi antar tingkatan tropik. Fungsi ekologis tersebut akan sangat mendukung keberlanjutan suatu fungsi ekosistem.
Keuntungan ekonomis yang seringkali dapat diupayakan dari budidaya tanaman legum tersedianya komoditas yang bernilai ekonomis sebagai bahan baku industri kerajinan, pangan, dan pakan. Beberapa jenis akasia adalah
sumber utama industri kerajinan patung tradisional karena memiliki pertumbuhan yang cepat dan dapat dibudidayakan pada berbagai jenis tanah dan iklim. Tanaman kacang-kacangan seperti kedelai, kacang tanah, kacang bogor, buncis, ercis dan bengkuang merupakan sumber pendapatan penting bagi petani lahan kering dan sebagian di antaranya adalah sumber utama bahan baku industri, misalnya kecap, kedelai dan tahu. Disisi lain, gamal dan lamtoro adalah jenis tanaman legum yang potensial digunakan sebagai pakan ternak.
Makalah ini merupakan suatu review pustaka yang mengulas peranan tanaman leguminose dalam menyuplai nitrogen bagi pertumbuhan tanaman non leguminose di sekitarnya. Berbagai mekanisme yang berhubungan dengan topik tersebut dibahas secara relatif detail.
II. KEANEKARAGAMAN HAYATI MIKROBA
DALAM SIKLUS NITROGEN
Keanekaragaman hayati yang meliputi keragaman genetik, spesies dan fungsional sangat menentukan fungsi ekosistem (Tilman et al., 1996). Fungsi ekosistem dipengaruhi oleh keberadaan dan aktivitas kelompok organisme fungsional. Kelompok fungsional merupakan suatu kesatuan spesies yang menunjukkan respons (respons grup) atau pengaruh (efek grup) yang sama terhadap proses utama dalam ekosistem (Hooper et al., 2005). Setiap tingkatan tropik dalam suatu ekosistem memiliki kelompok fungsional spesifik. Salah satu contoh kelompok fungsional yang berpengaruh dalam proses biogeokimia adalah mikroba yang terlibat di dalam siklus unsur nitrogen, yang meliputi mikroba heterotrof, nitrifier, denitrifer dan penambat N2. Salah satu contoh
produser yang berperan dalam menentukan siklus N adalah tanaman leguminose.
Perubahan ketersediaan nitrogen sangat menentukan keanekaragaman hayati dalam suatu ekosistem. Nitrogen adalah unsur hara makro yang diperlukan dalam jumlah terbesar oleh tanaman. Nitrogen memiliki beberapa bentuk, yaitu gas bebas (N2), nitrogen organik (asam-asam amino, protein,
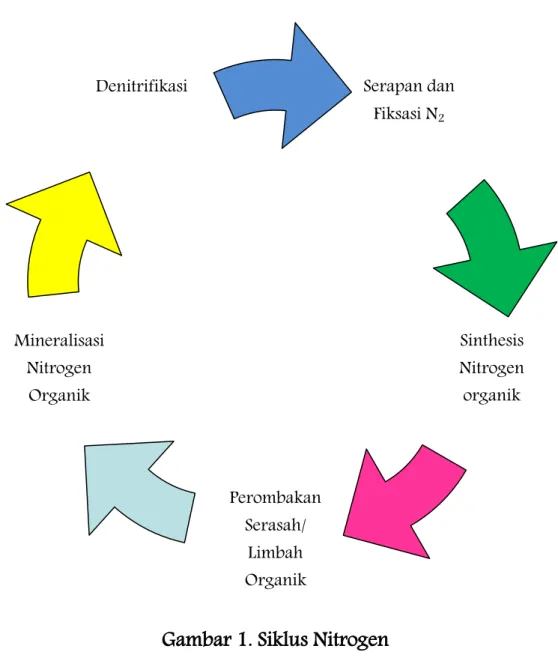
peptide), dan anorganik (ammonia, ammonium, nitrat, nitrit, dll). Perubahan bentuk unsur nitrogen terjadi melalui proses berbeda dalam siklus nitrogen (Gambar 1). Siklus nitrogen secara umum meliputi penambatan N2, sinthesis
nitrogen organik, perombakan serasah organik, mineralisasi nitrogen dan denitrifikasi.
Gambar 1. Siklus Nitrogen
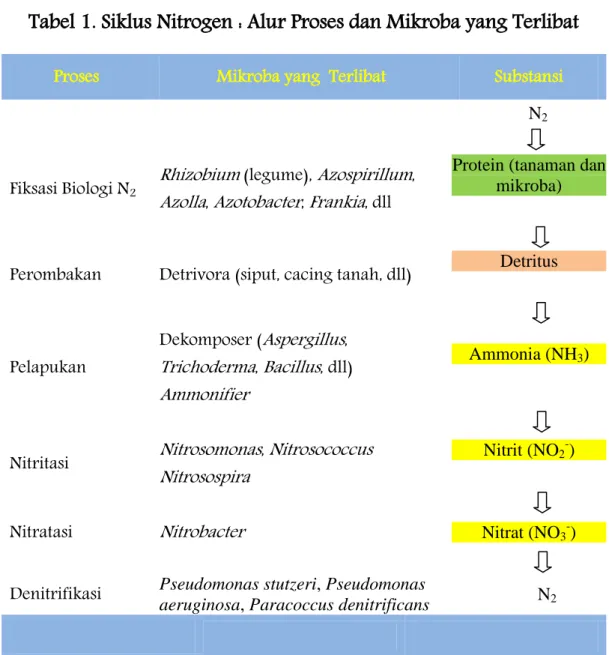
Perubahan bentuk nitrogen dilakukan oleh beberapa organisme dari tingkatan tropik yang berbeda. Berbagai bentuk nitrogen, proses transformasinya dan organisme yang terlibat di dalam siklus nitrogen ditampilkan dalam Tabel 1.Gas nitrogen merupakan komponen terbesar gas yang ada di atmosfer (72 %). Akan tetapi, nitrogen dalam bentuk gas N2 tidak
dapat dimanfaatkan secara langsung dalam proses biokimia sebagian besar mahluk hidup. Beberapa kelompok organisme fungsional berperan dalam transformasi bentuk N2 sehingga dapat dimanfaatkan dalam metabolisme
Serapan dan Fiksasi N2 Perombakan Serasah/ Limbah Organik Denitrifikasi Sinthesis Nitrogen organik Mineralisasi Nitrogen Organik
mahluk hidup. Oleh karena itu, kehilangan salah satu atau beberapa kelompok fungsional dalam siklus N akan mengubah atau menghentikan proses yang terlibat di dalam siklus N.
Tabel 1. Siklus Nitrogen : Alur Proses dan Mikroba yang Terlibat
Proses Mikroba yang Terlibat Substansi
N2
Fiksasi Biologi N2 Rhizobium (legume), Azospirillum,
Azolla, Azotobacter, Frankia, dll
Protein (tanaman dan mikroba)
Perombakan Detrivora (siput, cacing tanah, dll) Detritus
Pelapukan
Dekomposer (Aspergillus, Trichoderma, Bacillus, dll) Ammonifier
Ammonia (NH3)
Nitritasi Nitrosomonas, Nitrosococcus
Nitrosospira
Nitrit (NO2-)
Nitratasi Nitrobacter Nitrat (NO3-)
Denitrifikasi Pseudomonas stutzeri, Pseudomonas aeruginosa, Paracoccus denitrificans N2
Gas nitrogen merupakan komponen terbesar gas yang ada di atmosfer (72 %). Akan tetapi, nitrogen dalam bentuk gas N2 tidak dapat dimanfaatkan
secara langsung dalam proses biokimia sebagian besar mahluk hidup. Beberapa kelompok organisme fungsional berperan dalam transformasi bentuk N2
sehingga dapat dimanfaatkan dalam metabolisme mahluk hidup. Oleh karena itu, kehilangan salah satu atau beberapa kelompok fungsional dalam siklus N akan mengubah atau menghentikan proses yang terlibat di dalam siklus N.
Bakteri penambat N2 dapat mengkonversi N2 menjadi ammonia (NH3).
Ammonia selanjutnya dimetabolisme oleh mikroba dan tanaman menjadi protein sebagai salah satu penyusun tubuhnya. Nitrogen organik yang terikat dalam tubuh mahluk hidup yang sudah mati akan dirombak oleh detritivora (Gambar 2). Perombakan serasah organik dilanjutkan dengan pelapukan oleh mikroba sehingga terjadi perubahan N-organik menjadi N-anorganik melalui proses ammonifikasi dan mineralisasi N oleh bakteri nitrifikasi membentuk senyawa nitrat. Nitrat kemudian diabsobsi oleh mikroba maupun tanaman dan diubah menjadi N-organik, atau nitrat juga dapat mengalami denitrifikasi pada suasana reduktif menjadi gas N2.
Keanekaragaman hayati tanaman sangat menentukan siklus N (Oelman dan Wilcke, 2004). Keragaman hayati tanaman dapat mempengaruhi serapan total N karena pemanfaatan sumberdaya secara sinergis atau kompetitif. Apabila nutrisi diperoleh dari sumber tersedia yang berbeda (ruang, waktu dan bentuk yang berbeda) maka serapan N total oleh tanaman akan meningkat sehingga peluang terjadinya leaching lebih kecil (Hooper dan Vitousek, 1997). Spesies tanaman yang tumbuh lambat tidak terlalu banyak membutuhkan N. Tanaman yang berbeda kandungan N-nya akan memiliki serasah dengan kandungan N yang berbeda sehingga waktu dan jenis organisme yang diperlukan untuk proses dekomposisinya juga akan berbeda sehingga pada akhirnya mempengaruhi siklus N (Wedin dan Pastor, 1993).
Gambar 2. Beberapa Contoh Organisme Perombak Bahan Organik
Keanekaragaman hayati yang lebih besar juga dapat menimbulkan efek kompetitif. Efek kompetitif dapat terjadi apabila tanaman memanfaatkan sumber nutrisi, ruang habitat pada waktu yang bersamaan atau efek sinergis/komplementer akan terjadi apabila spesies tanaman memanfaatkan nutrisi, sinar matahari, habitat dan air dengan cara berbeda.keluwing
Cacing tanah Lipan
Siput Rayap
Kumban g
Keberadaan tanaman leguminose dalam suatu ekosistem umumnya menyebabkan terjadinya peningkatan kadar N (nitrat dan total) tanah. Peningkatan kadar nitrat tanah disebabkan karena adanya suplai N dari N2 yang
difiksasi oleh tanaman leguminose (Marquard et al., 2009; Olemann dan Wilcke, 2004; Zak et al., 2003). Peningkatan kadar nitrat tanah dapat berpengaruh positif ataupun negatif bagi lingkungannya. Suplai N oleh tanaman leguminose dapat meningkatkan produksi dan biomassa tumbuhan di sekitarnya (Spehn, et al., 2002; Weigelt et al., 2009) atau deposisi N yang terlalu besar dapat menurunkan produksi sporokap cendawan mikorhiza (Lileskov dan Bruns, 2001). Berkurangnya produksi sporokap akan menurunkan populasi mikorhiza di dalam tanah.
2.1. Fiksasi N
2dan Suplai Nitrogen oleh Tanaman Legum
Leguminose tergolong kelompok Fabaceae atau leguminoceae. Tanaman leguminose terdiri dari hampir 20.000 spesies yang tergabung dalam 750 genus. Hanya sekitar 3.500 spesies tanaman leguminose yang diketahui menambat N2
(Moreira, 2007) dan hanya 15 % spesies yang sudah diuji kemampuannya untuk bersiombiosis dengan rhizobia (Allen dan Allen, 1981). Leguminose memiliki fungsi ekologis dan ekonomis yang penting karena merupakan salah satu kelompok fungsional dalam memelihara kesuburan tanah suatu ekosistem dan sumber protein bagi manusia dan ternak serta bahan kayu yang berkualitas.
Fiksasi N2 dari atmosfer merupakan proses biologi terpenting kedua
setelah fotosintesis. Dalam proses tersebut terjadi reduksi gas N2 menjadi 2
molekul ammonia yang dilakukan oleh mikroba yang memiliki enzim nitrogenase. Fiksasi N2 dapat terjadi secara simbiosis antara tanaman legum
dengan rhizobia penambat N2. Proses tersebut dapat menyumbangkan lebih
dari 100 juta m3 ton N per tahun dan memenuhi 66 % kebutuhan nitrogen
untuk lahan pertanian.
Penambatan N2 melalui tanaman leguminose hanya terjadi di dalam
bintil akar efektif yang mengandung bakteroid rhizobium. Tanaman leguminose yang tidak membentuk bintil akar atau memiliki bintil akar yang tidak efektif tidak dapat menambat N2. Bintil akar efektif terbentuk apabila perakaran
tanaman leguminose diinfeksi oleh spesies rhizobium yang sesuai secara genetik. Jumlah spesies Rhizobium yang telah dikenal selama ini terdiri dari 16 spesies yang tergolong ke dalam 4 genus (Tabel 2). Pembentukan bintil akar dikendalikan oleh gen nod A, B, C, dan D yang terdapat pada seluruh Rhizobia (Gambar 3), sedangkan gen yang menyandi kesesuaian rhizobia dengan inang adalah nod E, F, G, H, I, J, K, L, M, P, Q.
Tabel 2. Genus dan Spesies Rhizobium serta Tanaman Inang
Spesies Tanaman Inang
Rhizobium
R. leguminosorum Pea, field bean, lentil, phaseoli
R. loti Lotus, trefoil
R. tropici Leucaena, ipil-ipil
R. etli Phaseolus
R. galegae Galega, Leucaena
R. buakuii Astragalus R. ciceri Cicer R. medditerraneum Cicer Sinorhizobium. S. meliloti Alfalfa S. fredii Kedelai S. saheli Sesbania
S. teranga Sesbania, acacia
Bradyrhizobium B. japonicum Kedelai B. elkanii Glycine B. liaoningense Glycine Azorhizobium A. caulinodans Sesbania Sumber : Graham (1998)
Penambatan N2 oleh rhizobia terjadi melalui reduksi molekul N2 menjadi
ammonia dengan reaksi berikut :
Ammonia yang terbentuk kemudian ditransfer dari bakteroid ke dalam sel akar inang dan dikonversi menjadi aspargin. Aspargin selanjutnya ditranslokasikan ke bagian atas tanaman, sedangkan tanaman menyuplai nutrisi dalam bentuk fotosintat untuk mendukung aktivitas Rhizobia. Minchin dan Pate (1973 dalam Bergensen, 1977) menyatakan bahwa 32 % fotosintat dialirkan ke dalam bintil akar yang digunakan untuk pertumbuhan dan pemeliharaan bintil (5 %), untuk respirasi (12 %) serta dikembalikan kepada tanaman dalam bentuk kombinasi dengan nitrogen (15 %).
Nitrogen yang difiksasi melalui tanaman leguminose dapat secara langsung dan tidak langsung ditransfer kepada tanaman lainnya yang tumbuh di sekitar tanaman leguminose. Proses transfer nitrogen oleh tanaman leguminose dapat melalui beberapa mekanisme. Transfer N terbesar dapat dilakukan setelah proses mineralisasi N organik menjadi N anorganik. Dengan adanya proses transfer N tersebut merupakan salah satu faktor pendukung terbentuknya asosiasi tanaman leguminose dengan jenis tanaman lainnya.
2.2. Mineralisasi Nitrogen
Tanaman pada umumnya menyerap N dalam bentuk N anorganik (ammonium dan nitrat) untuk mendukung pertumbuhannya. N anorganik terbentuk dari proses mineralisasi senyawa-senyawa organik yang mengandung N. Proses mineralisasi N organik terjadi melalui 2 tahapan reaksi, yaitu ammonifikasi dan nitrifikasi. Ammonifikasi adalah proses perombakan senyawa N organik secara enzimatik menjadi ammonium. Beberapa enzim yang terlibat adalah proteinase, protease, peptidase, khitinase, ketobiase. lisozim, endonuklease, eksonuklease dan urease. Ammonia yang terbentuk selanjutnya
dikonversi menjadi nitrat melalui proses nitrifikasi dengan reaksi sebagai berikut :
NH3 1,5O2 + NO2- + H+ + H2O
NO2- + ½ O2 NO3
-Beberapa jenis mikroba yang terlibat dalam proses nitrifikasi antara lain adalah Nitrosomonas, Nitrosococcus, Nitrobacter, dan Nitrospira.
III. TRANSFER NITROGEN DARI TANAMAN LEGUM
KE TANAMAN NON LEGUM
Transfer nitrogen (N) adalah pemindahan N dari tanaman legum ke tanaman non legum yang umumnya adalah rumput (Bropy et al., 1987). Istilah tersebut juga digunakan untuk menggambarkan pengaruh menguntungkan residu N tanaman legum yang sudah mati (Ofori dan Stern, 1987). Proporsi N pada tanaman rumput yang berasal dari leguminose yang ditanam secara tumpangsari sangat bervariasi tergantung kepada lama waktu pengamatan, spesies tanaman, umur tanaman, metodologi yang digunakan, serta kondisi lingkungan dan penelitian. Bropy et al (1987) menemukan bahwa 68 % nitrogen yang terkandung di dalam rumput kanari (Phalaris arundinacea L.) berasal dari alfalfa (Medicago sativa L.) dan 79% nitrogen dari tanaman Lotus corniculata L. Jumlah tersebut merupakan 17 dan 13 % dari total N yang difiksasi berturut-turut oleh alfalfa dan Lotus corniculata L. Penelitian Haystead dan Marriot (1979) membuktikan terjadinya transfer N sebesar 6 – 12 % dari tanaman white clover ke tanaman ryegrass (Lolium perens L.). Jumlah nitrogen yang ditransfer oleh tanaman alfalfa ke bromegrass adalah 14 kg/ha/th dengan proporsi 5 kg/ha/th berasal dari tanah dan 9 kg/ha/th berasal dari fiksasi N2.
Pada hamparan rumput di New Zealand, 50 % kebutuhan N rumput disuplai oleh tanaman white clover (Ledgard, 1991 dalam Tomm, 1993).
Suplai N oleh tanaman leguminose kepada tanaman non leguminose menyebabkan terjadinya peningkatan pertumbuhan dan produksi tanaman non leguminose. Salah satu contohnya adalah jumlah produksi pakan ternak dari campuran rumput dan tanaman leguminose sama besarnya dengan produksi
rumput monokultur dengan pemupukan lebih dari 100 kg N/ha (Knight, 1984 dalam Tomm, 1993).
Berdasarkan kajian terhadap beberapa hasil penelitian, mekanisme transfer nitrogen dapat digolongkan ke dalam 2 kelompok , yaitu mekanisme transfer jangka pendek (short term) dan jangka panjang (long term). Transfer N jangka pendek tidak melibatkan proses dekomposisi jaringan tanaman, sedangkan transfer N jangka panjang melibatkan proses dekomposisi bahan organik.
3.1. Mekanisme Transfer N Jangka Pendek
Mekanisme transfer N jangka pendek terdiri dari beberapa proses, yaitu (1) ekskresi senyawa nitrogen oleh tanaman yang kemudian diasimilasi oleh tanaman lainnya yang berasosiasi dengan tanaman tersebut (Gambar 4), (2) transfer N yang difasilitasi oleh mikoriza, dan (3) proses leaching N terlarut dari daun tanaman. Beberapa tanaman leguminose yang membentuk nodul mengeksresikan senyawa N yang kemudian digunakan oleh tanaman lainnya yang tidak mampu menambat N2 (Ruschel et al., 1979). Whitney dan Kanehiro
(1967) menyatakan, sejumlah besar proporsi N dilepaskan dari akar tanaman leguminose tropis dalam waktu 1 minggu setelah rontoknya daun. Laju dan jumlah N terfiksasi yang dilepaskan oleh perakaran tanaman leguminose semakin besar pada tanaman yang merana karena proses perontokan daun dan akibat perlakuan pemupukan (Wilman, 1970).
Interaksi interspesifik asosiasi tanaman leguminose dan rumput dipengaruhi oleh mikorhiza vesicular arbuscular (MVA) (Hetrick et al., 1989). Asosiasi tersebut meningkatkan zone serapan akar sehingga memperbesar
penyerapan unsur hara. Jaringan hifa yang terbentuk pada tanaman yang membentuk MVA dapat juga menginfeksi dan membentuk MVA pada tanaman lain yang berada didekatnya sehingga memungkinkan terjadinya transfer unsur hara termasuk N. Selain itu, keberadaan MVA dapat menambah jumlah ketersediaan P sehingga meningkatkan laju fiksasi N2 (Barea dan Azcon Aguilar,
1983) yang pada akhirnya mungkin meningkatkan ketersediaan dan transfer N. Van Kessel et al. (1985) menemukan terjadinya peningkatan transfer N dari tanaman kedelai ke tanaman jagung apabila kedua tanaman tersebut dikolonisasi oleh cendawan mikorhiza.
Sebagian besar kandungan N tanaman pakan ternak terakumulasi di dalam daun. Nitrogen tersebut dapat mengalami leaching dari daun karena pengaruh hujan terutama pada saat pertumbuhan atau penuaan daun (Whitney dan Kanehiro, 1967). Senyawa nitrogen yang terkandung di dalam air presipitasi dapat diserap secara langsung oleh kanopi tanaman (Harper et al., 1987) atau diserap langsung oleh akar tanaman.
Sumber : Tarui et al. (2013)
Gambar 4. Transfer N dari Tanaman Legum ke Tanaman Non Legum Transfer N dari tanaman legum ke rumput
3.2. Mekanisme Transfer N Jangka Panjang
Mekanisme transfer N jangka panjang meliputi proses dekomposisi akar , bintil akar, batang daun dan bunga, dan serasah tanaman leguminose (Gambar 5), serta pelepasan N dari kotoran ataupun urin ternak pemakan leguminose. Jumlah N yang dilepaskan dari proses tersebut sangat tergantung kepada kandungan N tanaman leguminose. Setiap jenis tanaman leguminose memiliki kandungan N yang berbeda (Tabel 3). Tidak seluruh N tersebut diperoleh dari hasil fiksasi N2. Tanaman leguminose pakan ternak dapat memenuhi 90 %
kebutuhan N-nya dari fiksasi N2, sedangkan tanaman leguminose penghasil biji
hanya mampu memenuhi 50 % dari kebutuhan N-nya dari fiksasi N2 (Paul dan
Clark, 1996). Tanaman leguminose pakan ternak umumnya dibudidayakan dalam waktu yang lebih lama, sedangkan leguminose penghasil biji dipanen lebih cepat sehingga waktu untuk memfiksasi N2 sepanjang siklus hidupnya
lebih lama pada tanaman leguminose pakan ternak.
Tabel 3. Kadar N di Bagian Akar dan Atas Tanaman Legum
Tanaman Kadar N di Bagian Atas Tanaman (kg/ha) Tanaman (kg/ha) Kadar N di Akar Total N (kg/ha) Legum Biji
Lupin putih 448 93 541
Faba bean 320 57 377
Filed Pea 291 40 331
Spring vetch 238 36 274
Legum Pakan Ternak
Red clover 381 118 499
White clover 322 131 453
Lucerne 469 157 626
Sainfoin 184 140 324
Setelah tanaman leguminose penghasil biji dipanen, tanah tempat tumbuhnya mungkin memiliki kandungan N anorganik yang lebih tinggi dibandingkan dengan tanaman yang tidak memfiksasi N2. Terdapat
kemungkinan leguminose tidak memanfaatkan N anorganik tanah dalam jumlah besar. Hal tersebut disebabkan oleh 2 hal yang berbeda. Pertama, sistem perakaran tanaman legume penghasil biji yang lunak dan serabut tidak memungkinkan pemanfaatan ammonium dan nitrat di lapisan bawah tanah secara efisien. Kedua, rendahnya rasio C/N akar tanaman legumniosa dan deposisi nutrisi di akar menyebabkan total mineralisasi tanaman leguminose lebih besar.
Proses pengguguran atau pemangkasan daun dapat menyebabkan terjadinya peluruhan akar dan bintil akar tanaman leguminose yang diikuti dengan pertumbuhan kembali jaringan akar dan bintil akar yang baru (Herriot dan Wells, 1960). Akar dan bintil akar yang mati mengandung sebagian besar N yang dapat ditransfer. Daun tanaman leguminose yang sudah tua dan kemudian gugur dapat mengalami proses dekomposisi dan merupakan sumber N bagi tanah dan tanaman di sekitarnya (Whitney dan Kanehiro, 1967). Sebagai contoh adalah nitrogen yang dikembalikan ke dalam tanah melalui daun tanaman alfalfa dan bromegrass yang jatuh berturut-turut adalah 13 dan 14 kg/ha/th (Tomm, 1993).
Beberapa jenis tanaman leguminose yang berperan sebagai mulsa atau penutup tanah hidup sering difungsikan ganda sebagai sumber pupuk hijau. Pada periode tertentu tanaman tersebut dipanen dan dibenamkan ke dalam tanah. Dekomposisi serasah tanaman leguminose berlangsung lebih cepat
karena memiliki rasio C/N yang rendah. Eason dan Newman (1990) melaporkan bahwa 60 % N dan 70 % P dari ryegrass dilepaskan ke dalam tanah dalam waktu 3 minggu setelah dekomposisi. Peningkatan jumlah N dan P yang dilepaskan dalam proses dekomposisi tersebut meningkat nyata dalam kurun waktu 30 hari.
Pembudidayaan tanaman leguminose semusim yang dipanen untuk pakan ternak akan menyumbangkan residu N dalam jumlah lebih kecil dibandingkan tanaman pakan ternak yang dibudidayakan dalam sistem penggembalaan. Pada sistem penggembalaan, jumlah bahan organik yang dipanen dan keluar dari sistem tersebut lebih kecil karena adanya pengembalian dalam bentuk kotoran dan urin ternak. Dekomposisi kotoran dan ekskresi urin ternak mengandung sejumlah N yang dikembalikan ke dalam tanah sehingga dapat digunakan kembali oleh tanaman lain yang berasosiasi dengan tanaman leguminose.
Diadopsi dari : http://www.biopowerlanka.com/super-seed.html (diunduh pada tanggal 22 Juli 2016)
Gambar 5. Transfer N Jangka Panjang dari Tanaman Legum ke Tanaman Non Legum Residu Tanaman Nitrogen Anorganik Fiksasi Biologi Nitrogen Dekomposisi
IV. ASOSIASI TANAMAN LEGUMINOSE
4.1. Definisi dan Sistem Klasifikasi
Salah satu cara untuk memahami ekosistem dan menerapkan prinsip-prinsip pengelolaan ekosistem adalah berdasarkan sistem klasifikasi ekologis. Klasifikasi memungkinkan pengelompokan, pembandingan, sintesis, pemetaan dan inventarisasi informasi serta menyediakan bahan dan media komunikasi mengenai hasil klasifikasi. Berbagai sistem klasifikasi telah dikembangkan dengan tujuan dan cara berbeda, misalnya Biogeoclimatic Ecosystem Classification (MacKenzie, 2004) di Kanada dan National Vegetation Classification System (NVCS) di Amerika (Christy, 2004). Akan tetapi, pendekatan yang umumnya digunakan adalah berdasarkan konsep asosiasi tanaman.
Asosiasi menggambarkan munculnya kembali komunitas tanaman yang memiliki diagnose spesies yang spesifik, yaitu suatu karakteristik yang bervariasi dalam komposisi spesies dan skala yang tegas mengenai kondisi atau struktur habitatnya. Asosiasi tanaman merupakan kumpulan satu spesies relatif tanaman dalam satu hamparan lahan tertentu. Misalnya, asosiasi tanaman pada lahan basah di daerah barat daya Oregon. Berdasarkan kunci dan deskripsi khusus (National Vegetation Classification System) ditemukan 122 jenis asosiasi tanaman yang meliputi 14 jenis hutan dan tanaman berkayu, 28 semak, 78 tanaman herba, dan 2 kelompok tanaman nonvaskular. Suatu jenis tanaman monotipik dinyatakan sebagai asosiasi apabila menempati areal minimal seluas 100 m2. Untuk membedakan asosiasi pohon dan semak dengan asosiasi herba,
maka hamparan asosiasi yang dinyatakan sebagai asosiasi herba adalah hamparan dengan minimal 20 % tutupan oleh tanaman herba. Asosiasi hutan yang ditemukan di Kanada terdiri dari 236 asosiasi. Sebagian besar (190 atau 80%) memiliki paling tidak 1 asosiasi yang serupa (tingkat kesamaan > 60 %) dan 60 (25 %) dari bagian tersebut tergolong sangat mirip (tingkat kesamaan > 70 %).
4.2. Aspek Positif Asosiasi Tanaman
Setiap jenis tanaman secara genetik dan fenotip memiliki kemampuan untuk berasosiasi dengan tanaman lainnya. Seperti tanaman lainnya, tanaman leguminose dapat berasosiasi dengan tanaman lainnya. Salah satu bentuk prinsip asosiasi tanaman legum yang diterapkan secara luas adalah dalam sistem agroforestry. Dalam sistem tersebut, tanaman legum sebagai tanaman utama ataupun sela dikombinasikan dengan jenis tanaman lainnya dalam suatu hamparan tertentu.
Beberapa keuntungan yang dapat diperoleh oleh tanaman dalam suatu asosiasi adalah : penambahan N dari proses fiksasi N2, penekanan pertumbuhan
serangga hama secara biokimia, interaksi spasial fisik, dan habitat yang menguntungkan. Fiksasi N2 yang dilakukan oleh rhizobia akan meningkatkan
ketersediaan N bagi tanaman leguminose maupun tanaman lainnya.
Beberapa jenis tanaman tertentu dapat menghasilkan eksudat kimia dari akar maupun bagian atas tanaman yang dapat menekan atau mengundang serangga hama sehingga tanaman yang berada di sekitarnya terlindung dari serangan hama tersebut. Salah satu contohnya adalah thiopene yang dihasilkan
oleh tanaman marigold Afrika merupakan suatu repelen bagi nemathoda patogen.
Tanaman yang tumbuh tinggi, memerlukan lebih banyak matahari dan dapat berbagi ruang tumbuh dengan tanaman yang lebih pendek serta tidak terlalu banyak memerlukan pencahayaan sehingga kedua tanaman tersebut dapat memberikan total hasil yang lebih tinggi dibandingkan dengan tanaman yang sama yang tumbuh secara homogen. Interaksi spasial tersebut juga dapat melindungi tanaman dari serangan hama tanaman. Tanaman yang memiliki tegakan tinggi atau kanopi rapat dapat melindungi tanaman yang lebih lemah yang berada di bawahnya dengan efek naungan atau melindunginya dari hembusan angin yang kuat.
4.3. Interaksi dalam Asosiasi Tanaman
Setiap jenis tanaman memiliki kanopi. Kanopi merupakan arsitektur komunitas yang mengandung komposisi spesies dan tempat terjadinya siklus unsur hara, transfer energi, interaksi tanaman-binatang, dan mewadahi aspek konservasi dari permukaan tanah sampai dengan perbatasan komunitas dengan atmosfer (Rinker dan Lowman, 2001). Keberadaan kanopi merupakan tempat interaksi tanaman dengan organisme lainnya. Komponen kanopi antara lain meliputi bagian atas tanaman, organisme setempat (epifit dan epifil), organsime dengan mobilitas tinggi (misalnya burung, mamalia dan serangga) dan proses yang terjadi di dalam kanopi (misalnya siklus unsur hara).
Habitat yang lebih menguntungkan secara biologis dapat terbentuk dalam suatu asosiasi tanaman. Beberapa jenis tanaman dapat menyediakan
tempat hidup yang baik bagi beberapa serangga dan arthropoda musuh alam sehingga dapat menekan serangan hama pada tanaman di sekitarnya. Asosiasi tanaman menyediakan habitat tempat tumbuhnya beragam organisme dari berbagai tingkatan tropik. Interaksi nyata terdapat antara herbivora di bagian kanopi tanaman dengan fauna tanah dan proses dekomposisi bahan organik sehingga mempengaruhi ketersediaan unsur hara di dalam tanah. Beberapa kelompok herbivora dapat mempengaruhi laju dekomposisi dan siklus unsur hara karena mengkonsumsi bagian-bagian tertentu tanaman (Pastor dan Cohen, 1997). Penurunan jumlah daun karena serangan serangga herbivora menyebabkan terjadinya translokasi unsur hara terutama nitrogen dari tegakan tanaman (Reynolds et al., 2000).
Akar setiap jenis tanaman akan menghasilkan eksudat akar yang memiliki komposisi kimia berbeda. Senyawa kimia tersebut dapat dimanfaatkan sebagai nutrisi oleh beberapa jenis mikroba tertentu sehingga terbentuk asosiasi tanaman dengan mikroba tanah. Beberapa kelompok dekomposer dapat memanfaatkan serasah atau bahan organik tanah sebagai sumber karbonnya. Beberapa organisme pathogen dapat memangsa organisme tertentu di dalam tanah.
4.4. Asosiasi Tanaman Legum
Tanaman leguminose dapat berasosiasi dengan beberapa jenis tanaman lainnya, seperti kelompok rumput-rumputan, biji-bijian (Alvey et al., 2003), semak, dan ditumpangsarikan dengan beberapa jenis tanaman pangan (Alvey et al., 2003; Mustafa et al., 2004) dan perkebunan (Liphadzi dan Reinhardt, 2004;
Pound et al., 1980). Leguminose berinteraksi secara spesifik dengan mikroba fungsional penambat N2 dan kadangkala dengan cendawan mikorhiza.
Setiap jenis tanaman leguminose memiliki kandungan N berbeda satu sama lain sehingga rasio C/N-nya juga berbeda. Perbedaan rasio C/N serasah tanaman tersebut menyebabkan proses dekomposisi yang berbeda dengan melibatkan populasi dekomposer yang berbeda. Hasil penelitian Oyun et al (2006) menunjukkan, bahwa populasi terbesar dekomposer ditemukan pada kombinasi serasah Acacia yang memiliki rasio C/N tinggi dengan Gliricida yang memiliki rasio C/N lebih rendah. Populasi dekomposer pada serasah murni gliricida maupun acacia lebih rendah daripada populasi dekomposer pada serasah campuran.
Tanaman leguminose penutup tanah juga mampu menyediakan habitat yang sesuai untuk perkembangan musuh alam bagi hama arthropoda (Hokkanen, 1991). Penelitian lain menyebutkan bahwa pemilihan spesies tanaman leguminose untuk penutup tanah perlu dikaji supaya tidak menjadi sumber hama bagi tanaman lainnya. Lapointe (2003) menemukan bahwa tanaman Cajanus cajan tidak sesuai digunakan sebagai tanaman penutup tanah di sela-sela pertanaman jeruk citrun karena berdampak positif terhadap pertumbuhan larva hama Diaprepes abbreviatus (Coleoptera: curculionidae) yang menyerang akar tanaman citrun..
V. KESIMPULAN
Tanaman leguminose merupakan tanaman fungsional yang mampu menambat N2 dan mensuplai nitrogen kepada tanaman non leguminose yang
ada di sekitarnya. Penambatan N2 merupakan salah satu bagian dari siklus
nitrogen. Suplai nitrogen yang dilakukan oleh tanaman leguminose kepada tanaman non leguminose dapat melalui beberapa mekanisme, yaitu mekanisme jangka pendek tanpa melalui proses dekomposisi (ekskresi senyawa nitrogen oleh tanaman yang kemudian diasimilasi oleh tanaman lainnya yang berasosiasi dengan tanaman tersebut, transfer N yang difasilitasi oleh mikoriza, dan proses leaching N terlarut dari daun tanaman) dan mekanisme jangka panjang (dekomposisi akar dan bintil akar tanaman leguminose, dekomposisi batang daun dan bunga tanaman leguminose, serasah tanaman leguminose, dan pelepasan N dari kotoran ataupun urin ternak pemakan leguminose).
Suplai nitrogen yang dilakukan oleh tanaman leguminose kepada tanaman non leguminose dapat membantu memenuhi sebagian kebutuhan N tanaman non leguminose. Suplai nitrogen oleh tanaman leguminose kepada lingkungannya merupakan salah satu faktor pendukung terbentuknya asosiasi tanaman leguminose dengan tanaman lainnya atau dengan organisme dari tingkatan tropik lainnya..
DAFTAR PUSTAKA
Alvey, S., C.H. Yang., A. Buerkert, D.E. Crowley. 2003. Cereal/legume rotation effects on rhizosphere bacterial community structure in west African soils. Biol Fertil Soils. 37:72-82.
Barea, J.M., F. El-Atrach, and R. Azcon. 1989. Mycorrhiza and phosphate interactions as affecting plant development, N2-fixation, N-transfer and
N-uptake from soil in legume-grass mixtures by using a 15N dilution
technique. Soil Biol. Biochem. 21:581-589.
Brophy, L.S., G.H. Heichel, and M.P. Risselle. 1987. Nitrogen transfer from forage legumes to grass in a systematic planting design. Crop Sci. 27:753-758.
Christy, J.A. 2004. Native Freshwater Wetland Plant Associations of Northwestern Oregon. Natural Heritage Information Center, Oregon State University. USA.
Eason, W.R. and E.I. Newman. 1990. Rapid cycling of nitrogen and phosphorous from dying roots of Lolium perenne. Oecologia. 82:432-436.
Graham, P.H. 1998. Biological Dinitrogen Fixation : Symbiotic. In. Principles and Apllications of Soil Microbiology. D.M. Sylvia, J.J. Fuhrman, P.G. Hartel and. D.A. Zuberer (Eds.). Prentice Hall. UK. Pp:322-345.
Harper, L.A., R.R. Shape, G.W. Langdale, and J.E. Giddens. 1987. Nitrogen cycling in awheat crop: soil, plant, and aerial transport. Agron. J. 79:965-973.
Hetrick, B.A.D., G.W.T. Wilson, and D.C. Harnett. 1989. Relationship between mycorrhizal dependence and competitive ability of two tallgrass prairie grasses. Can. J. Bot. 67:2608-2615.
Hokkanen, H.M.T. 1991. Trap cropping in pest management. Annu. Rev. Entomol. 36:119-138.
Hooper, D.U. and Vitousek, P.M. 1997. The effects of plant composition and diversity on ecosystem processes. Science 277:1302-1305.
Hooper DU, FS Chapin III, JJ Ewel, A Hector, P Inchausti, S Lavorel, JH Lawton, DM Lodge, M Loreau, S Naeem, B Schmid, H Setälä, AJ Symstad, J Vandermeer and DA Wardle. 2005. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol Monog75:3-35.
Lapointe, S. 2003. Leguminose cover crops and their interactions with citrus and Diaprepes abbreviatus (Coleoptera: Curculionidae).
Lilleskov, E.A. and T.D. Bruns. 2001. Nitrogen and ectomycorrhizal fungal communities: what we know, what we need to know. New. Phytologist. 149:154-158.
Liphadzi, K.B. and C.F. Reinhardt. 2006. Using companion plants to assist Pinus patula establishment on former agricultural lands. openUP.
MacKenzie, W.H. 2004. Plant associations as ecosystem : Issues in application at-risk status. In Hooper, T.D. (Ed) Proceedings of The Species at Risk. Victoria, B.C.
Marquard, E., Weigelt, A. Temperton, V.M., Roscher, C., Schumacher, J,m Buchmann, N, Fischer, M., Weisser, W.W. and Schmid, B. 2009. Plant species richness and functional composition drive overyielding in a 6-year grassland experiment. J. Ecol. Accepted.
Moreira, F.M. D.S. 2007. Soil biodiversity: genetic resources to enhance nitrogen fixation in agriculture and forestry. Lotus Newsletter. Vol : 37(3): 112-113.
Mustafa, TAN, Y. Serin, H. Ibrahim Erkovan. 2004. Effects of Barley as a companion crop on the hay yield and plant density of red clover and the botanical composition of Hay. Turk J. Agric. For. 28 :35-41.
Oelman, Y. dan Wilcke, W. 2004. The effect of biodiversity on nitrogen in the soil : species number versus presence of legumes. Geophysical Research Abstracts, Vol. 6, 05406. European Geosciences Union.
Ofori, F. and W.R. Stern. 1987. Cereal-legume intercropping systems. Adv. Agron. 41:41-90.
Oyun, M.B., Akharayi, F.C. and Adetuyi, F.C. 2006. Microbial population in decomposing legume litter of differing quality. American Journal of Agric. And Biol. Sci. 1(1):22-26.
Paynel F., Murray, P.J., and Cliquet, J.B. 2001. Root exudates: a pathway for short-term N transfer from clover and ryegrass. Plant and Soil, 229:235-243.
Pound, B. A. Santana, and G. Ruiz. 1980. Effect of companion crops on establishment and subsequent yield of Leucaena leucocephala. Trop. Anim. Prod. 5:3.
Rinker, H.B. and M. D. Lowman. 2001. Literature Review: canopy herbivory and soil ecology, the top-down impact of forest processes. Selbyana, 22(2):225-231.
Ruschel, A.P., E. Salati and P.B. Vose. 1979. Nitrogen enrichment symbiosis. Plant Soil. 51:425-429.
Tarui, A., A. Matsumura, S. Asakura, K. Yamawaki, R. Hattori and H. Daimon. 2013. Enhancement of nitrogen uptake in oat by cutting hairy vetch grown as an associated crop. www.plant.org: 83-91
Tilman D. Wedin D., and Knops J. 1996. Productivity and Sustainability influenced by biodiverfsity in grassland ecosystems. Nature. 379:718-720.
Tomm, G.O. 1993. Nitrogen Transfer in An Alfalfa-Bromegrass Mixture. Thesis. Dept. of Crop Science. And Plant Ecology University Saskatchewan Saskatoon, Saskatchewan, Canada.
Trannin, W.S. Urquisa, S., Guerra, G. Ibijbijen, J. and Cadisch, G. 2000. Interspecies competition an d N transfer in a tropical grass-legume mixture. Biol. Fertil. Soils. 32:441-448.
Van Kessel, C., J.P. Roskoski, and K. Keane. 1988. Ureide production by N2
-fixing and non-N2-fixing leguminous trees. Soil Biol. Biochem.
20:891-897.
Wedin, D. and Pastor J. 1993. Species effects on nitrogen cycling : a test with perennial grasses. Oecologia 84:433-441.
Weigelt, A., W.W. Weisser, N. Buchmann, and M. Scherer-Lorenzen. 2009. Biodiversity for multifunctional grasslands: equal productivity in high-diversity low-input and low-high-diversity high-input systems. Biogeosciences Discuss., Vol. 6, 3187-3214.
Whitney, A.S., and Y. Kanehiro. 1967. Pathways of nitrogen transfer in some tropical legume-grass associations. Agron. J. 59:585-588.
Zak, D.R., W.E. Holmes, D.C. White, A.D. Peacock, and D. Tilman. 2003. Plant diversity, soil microbial communities and ecosystem function: are there any links. Ecology:84(8):2042-2050.