Can plant palatability trials be used to predict the effect of
rabbit grazing on the flora of ex-arable land?
A. Diaz
∗Farmland Ecology Unit, Scottish Agricultural College, Mill of Craibstone, Bucksburn, Aberdeen, AB21 9TS, UK
Received 10 May 1999; received in revised form 10 May 1999; accepted 28 September 1999
Abstract
Surplus food production has led to increased interest in taking land out of arable farming, notably under set-aside regulations, and converting it into grasslands that resemble semi-natural plant communities. Despite the value of being able to predict the long-term success of such habitat creation projects, there is little ecological data available on which to base such predictions. This study tests whether plant palatability trials can be a useful method to predict the effect of grazing by rabbits on the success of individual species growing in grasslands. The palatability of 22 common, grassland species was tested by offering wild rabbits pot-grown plants of each species in a variety of grassland vegetation backgrounds in NE Scotland. Plants were inserted into plots consisting of a regular grid of holes cut into rabbit-grazed grasslands. The palatability of each species was tested in 10 such plots, on 20 occasions each. Plots varied in their vegetation composition and were each grazed by rabbits from a different warren. Plant species were found to differ greatly in their palatability to rabbits. Palatability was found to be unaffected by the composition of the surrounding vegetation. A survey was also carried out to determine the floristic composition of ungrazed and rabbit grazed swards of 8-year old set-aside. A strong relationship was found between the palatability of most plant species and their comparative abundance in ungrazed and grazed swards. Possible reasons for the few exceptions are discussed. In general, these findings indicate that palatability trials can be a useful and robust method of predicting the effect of rabbits on the success of individual plant species growing in grasslands. A method such as this, that allows the impact of rabbits on floristic composition to be predicted, can be used to inform grassland recreation and management decisions. ©2000 Published by Elsevier Science B.V. All rights reserved.
Keywords: Palatability; Rabbits; Grazing; Set-aside; Herbivory
1. Introduction
There is much current interest in the potential for converting arable land into grasslands that resemble long established, semi-natural vegetation types.
How-∗Present address: School of Conservation Sciences, Bournemouth
University, Dorset House, Talbot Campus, Fern Barrow, Poole, Dorset, BH12 5BB, UK; Tel.: +44-1202-595178; fax: +44-1202-595255.
ever, arable systems and semi-natural vegetation types characteristically have a very different soil chem-istry, flora and fauna. Consequently, many ecological changes need to occur during the transition from an arable system to a re-created semi-natural vegetation and these need to be well understood. Rabbits, which can be common on farmland, are known to influence the floristic composition of many types of vegetation (Farrow, 1917; Fenton, 1940; Gillham, 1955; Ran-well, 1960; Thomas, 1960, 1963; Watt, 1981; Bishop
and Davy, 1984; Foran, 1986). Rabbits have a range of direct and indirect effects on plant community composition. Direct effects can be caused by factors such as herbivory, scrapes, middens, trampling and seed dispersal in faeces. All of these factors may also produce indirect effects such as changes in the activity of other herbivores or changes in plant–plant competitive interactions. The effects of rabbit activity occur at different scales — for example, scrapes and middens can result in dramatic but localised impacts on vegetation composition (Thomas, 1963; Dixon and Hambler, 1993), while the effects of herbivory may be more diffuse and so affect a greater proportion of the vegetation at a locality. Recent investigations into the influence of rabbits on vegetation have sought to distinguish the impact of the various effects (Crawley, 1990; Prins and Nell, 1990; Hambler et al., 1995). Achieving this is difficult as many of the effects may be impossible to separate experimentally (Hambler et al., 1995) or may significantly interact, so impeding interpretation. Direct measurement of the extent of herbivory on different plant species is very difficult to obtain for free-living rabbits with open access to feeding areas. An index of relative herbivory can be obtained by scoring the proportion of stems and leaves that have been nibbled (Zeevalking and Fresco, 1977), but this gives no information on the amount of each stem or leaf that has been consumed unless fenced controls are used. However, the use of fenced controls has two important weaknesses; (i) the cages may cause micro-environmental changes (Williams, 1951; Watt, 1957, 1960, 1962), and (ii) they may not produce an accurate measure of herbivory as the rel-ative growth of different species may not be the same under grazed and ungrazed conditions. Alternative techniques such as the analysis of rabbit stomach con-tents (Myers and Bults, 1977) or rabbit faeces (e.g., Williams et al., 1974; Bhadresa, 1977) can establish the proportion of different species consumed but not where they were consumed. Direct observation of grazing can establish where species were consumed but not the proportion, unless combined with an anal-ysis of gut contents immediately after the feeding event. Such data would be difficult to obtain for wild rabbits in field conditions.
This study examines the potential for a non-invasive and non-destructive approach to estimating relative herbivory by wild rabbits in the field. It investigates
the relative palatability of a range of grasses and forbs to wild rabbits and then examines the extent to which such data can be used to predict the difference in abundance (% cover) of these species in grazed and ungrazed swards. Included is an examination of whether relative palatabilities are consistent when tested against a range of vegetation types.
2. Methods
2.1. Assessment of relative plant palatability
In the early summer of 1996, seeds of plant species commonly found on set-aside grassland in NE Scot-land were germinated on filter paper in Petri dishes and then transplanted into top soil from ex-arable land. Two hundred plants of each of 22 species were grown in seed trays in a glasshouse for 2 weeks. Each plant was then potted into an individual 100 ml pot, filled with top soil, and moved outside to a gravel bed. Plants were kept well watered and grown to a simi-lar above-ground size (ca. 0.3 g dry weight). This took between 5 and 7 weeks, depending on the species. No plants reached flowering stage during this time.
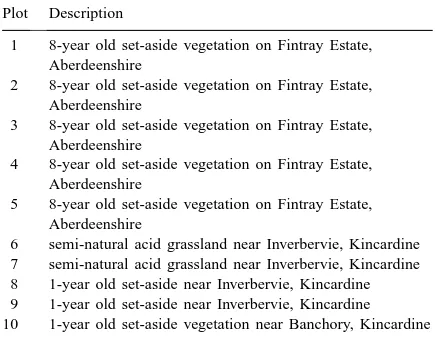
The palatability of these plants to rabbits was com-pared at 10 plots. These measured 5 m×3 m and were situated on homogeneous stands of self-sown vegeta-tion (Table 1). Each plot was grazed by rabbits from
Table 1
Location and description of the plots into which grassland plant species were inserted to test their palatability to wild rabbits
Plot Description
1 8-year old set-aside vegetation on Fintray Estate, Aberdeenshire
2 8-year old set-aside vegetation on Fintray Estate, Aberdeenshire
3 8-year old set-aside vegetation on Fintray Estate, Aberdeenshire
4 8-year old set-aside vegetation on Fintray Estate, Aberdeenshire
5 8-year old set-aside vegetation on Fintray Estate, Aberdeenshire
Table 2
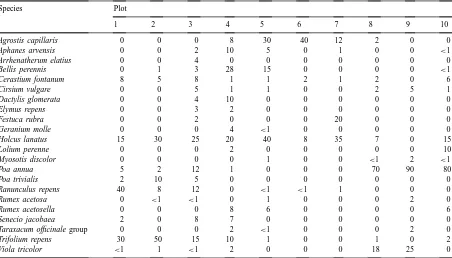
The abundance (% cover) of palatability-study species in the existing ‘background’ vegetation on each plot
Species Plot
a different warren. The plots chosen all had a high incidence of rabbits but varied in their floristic com-position. This allowed a test of whether background vegetation had an impact on relative plant palatabil-ity (Table 2). Test plants were inserted into a regu-lar grid of 22 holes, cut into the turf at each of the plots. The holes were spaced 1 m apart. This produced a grid of 24 potential holes, but as only 22 species were available, at random, two corner spaces were ex-cluded from the grid of holes each time. A spacing of 1 m meant that test plants were placed sufficiently far apart for rabbits to chose between test plants and the background vegetation rather than between test plants directly (pers. obs.).
The palatability of individual plants of each of the 22 species was tested simultaneously. At each plot, one plant from each species was inserted into the grid of 22 holes. Plants were assigned to holes at random and left for 24 h. At the end of this time, they were scored as either eaten (if any part of the plant was taken) or not eaten. This procedure was repeated on 19 further days per plot between the second week of August 1996 and the first week of September 1996. A count was made
of the number of times out of the 20 replicate times that each species was eaten at each plot. This was converted into a percentage palatability for each plant species at each plot. All plots were sampled on the same day; very wet days were avoided in case heavy rain sub-stantially reduced above-ground rabbit activity.
Table 3
The total number of plants of each species that were nibbled by rabbits (max=20 per plot)
Species Plot
1 2 3 4 5 6 7 8 9 10 Mean±SD
Elymus rpens 19 20 20 18 17 20 19 18 19 19 18.9±0.99
Arrhenatherum elatius 18 20 20 19 17 17 19 19 20 20 18.9±1.20
Dactylis glomerata 18 18 20 16 20 18 18 16 20 20 18.4±1.58
Taraxacum officinale group 19 19 20 18 17 16 19 18 17 17 18.0±1.25
Poa trivialis 20 15 17 18 19 16 15 20 20 19 17.9±2.02
2.2. Survey of sites to determine floristic composition and incidence of rabbits
To test the extent to which estimates of relative plant species’ palatability, obtained using the above method, could be related to the relative herbivory of species in grasslands, an examination was made of the correlation between the incidence of rabbits and plant species’ abundance in grasslands across NE Scotland. Fields surveyed were set-aside land that had been out of agricultural production for 8 years and had been al-lowed to revegetate from naturally available seed and propagules. These fields were located in three sepa-rate regions of north east Scotland: (i) near Dalcross, Inverness-shire, (ii) near Kirkton of Oyne, Aberdeen-shire and (iii) Fintray Estate, Fintray, AberdeenAberdeen-shire. All fields contained a mosaic of vegetation types. Most vegetation types were dominated by grasses, but some were composed mainly of low-growing forbs. Rabbits had formed burrows and variously-sized warrens in or around many of the fields studied.
In June 1996, a survey was carried out of the floris-tic composition and levels of rabbit incidence on sites
in these fields. Each site measured 25 m×25 m. Thirty sites were located at Dalcross, 40 at Kirkton of Oyne, and 40 at Fintray Estate. A total of 10 fields were sampled at Dalcross, 21 at Kirkton of Oyne and 22 at Fintray Estate. Sites were selected so as to sample all of the vegetation types found, except for vegeta-tion types on the few marshy areas present. No site contained rabbit burrows or warrens. All sites were in homogeneous stands of vegetation, but no stand con-tained more than one site. At each site, the total percent cover of each vascular plant species present was es-timated by eye. Plant nomenclature follows Clapham et al. (1987).
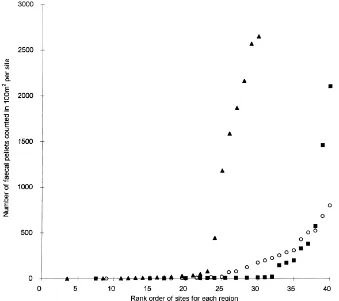
Fig. 1. The range in faecal pellet counts per site in the three regions studied. Closed squares represent Fintray sites. Open circles represent Oyne sites. Closed triangles represent Dalcross sites.
adjustments to quadrat positions. The quadrats were revisited 7 days after having been cleared and a count was made of all faecal pellets present. The differ-ences between the sites found by this method were in agreement with those found using two other indices of rabbit incidence, night counts and the removal of carrot bait (Diaz, 1998).
3. Results
3.1. Assessment of relative plant palatability
Plant species varied greatly in their observed palata-bility to rabbits (Table 3) (Chi square for species grouped across plots=2016.73, d.f.=21, P<0.001). Most palatable were many of the grass species such as Elymus repens and Arrhenatherum elatius, while Senecio jacobaea and Cirsium vulgare were least
palatable. Species’ differences in palatability were generally consistent across plots even for many species of intermediate palatability, such as Poa annua and Holcus lanatus (Chi square for plot×species’ ef-fects (Mather, 1966)=100.63, d.f.=168, P > 0.995). However, all of the most variable palatability scores were recorded for species of intermediate palatabil-ity; results for Festuca rubra were by far the most variable (Table 3).
3.2. Survey of sites to determine floristic composition and incidence of rabbits
Table 5
Spearman Rank correlations (r) between species’ abundance (% cover) and levels of rabbit incidence (number of faecal pellets) in 625 m2
sites in each regiona
Fintray (n=40 sites) Oyne (n=40 sites) Dalcross (n=30 sites)
Species r P Species r P Species r P
Aphanes arvensis +0.66 <0.001 Myosotis discolor +0.73 <0.001 Poa annua +0.80 <0.001 Bellis perennis +0.66 <0.001 Holcus lanatus +0.67 <0.001 Veronica officinalis +0.76 <0.001 Poa annua +0.66 <0.001 Cerastium fontanum +0.67 <0.001 Myosotis discolor +0.75 <0.001 Agrostis capillaris +0.54 <0.001 Trifolium repens +0.62 <0.001 Erophila verna +0.70 <0.001 Myosotis discolor +0.54 <0.001 Lolium perenne +0.58 <0.001 Aphanes arvensis +0.69 <0.001 Cerastium fontanum +0.50 0.001 Cirsium arvense +0.50 0.001 Capsella bursa-pastoris +0.64 <0.001 Trifolium dubium +0.50 0.001 Veronica serpyllifolia +0.47 0.002 Myosotis arvensis +0.64 <0.001 Senecio jacobaea +0.48 0.002 Ranunculus repens +0.45 0.004 Viola arvensis +0.58 0.001 Sagina procumbens +0.48 0.002 Bellis perennis +0.36 0.024 Agrostis capillaris +0.53 0.003 Veronica officinalis +0.48 0.002 Cirsium vulgare +0.33 0.037 Vicia sativa +0.52 0.003
Viola tricolor +0.41 0.009 Papaver rhoeas +0.50 0.005
Trifolium repens +0.38 0.015 Trifolium dubium +0.49 0.005
Cirsium vulgare +0.35 0.025 Geranium molle +0.44 0.015
Rumex acetosa +0.37 0.044 Senecio vulgaris +0.36 0.049
Dactylis glomerata −0.54 <0.001 Elymus repens −0.52 0.001 Dacylis glomerata −0.76 <0.001 Poa trivialis −0.48 0.002 Holcus mollis −0.48 0.002 Taraxacum officinale group −0.72 <0.001
Arrhenatherum elatius −0.42 0.007 Poa trivialis −0.56 0.001
Epilobium obscurum −0.32 0.048 Chamaenerion angustifolium −0.53 0.003 Ranunculus repens −0.47 0.009 Geranium dissectum −0.46 0.010
aOnly species that occurred in at least 20% of the sites per region are shown. Only correlations of P
<0.05 are shown.
correlated to levels of rabbit incidence (Table 5). Species that were significantly more abundant where rabbit incidence was higher tended to be low-growing forbs, e.g., Bellis perennis and Myoso-tis discolor, or species of disturbed ground, e.g., Papaver rhoeas and P. annua. By contrast, species whose abundance was negatively related to the inci-dence of rabbits were often taller-growing grasses e.g., Dactylis glomerata and Poa trivialis.
3.3. The relationship between plant palatability and sward composition
The extent to which differences in sward compo-sition between grazed and ungrazed sites could be related to differences in plant palatability was inves-tigated for the 22 species in the palatability trials. Although the sites showed a range of incidence of rabbits and plant abundance (Fig. 2), they were for the purpose of this analysis categorised as ‘grazed’ or ‘ungrazed’ based on whether they had more than 125
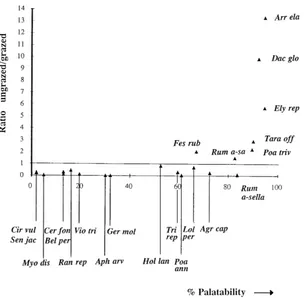
faecal pellets per 100 m2per week recorded on the site. The mean relative abundance of each plant species on ‘ungrazed’ and ‘grazed’ sites was calculated. A clear relationship emerged when the percent palatability of each plant species were plotted against the ratio of its mean relative abundance on ungrazed and grazed sites (Fig. 3). Most very palatable species were seen to be far more abundant on ungrazed sites and most un-palatable species were more abundant on grazed sites.
4. Discussion
Fig. 2. The relationship between the incidence of rabbits on each of the 110 sites studied (measured as number of faecal pellets found) and the abundance (measured as % cover) of a palatable plant species (Poa trivialis); an unpalatable species (Myosotis discolor); a species of intermediate palatability (Poa annua). Open triangles represent Poa trivialis. Crosses represent Myosotis discolor. Solid circles represent Poa annua.
competitive ability and palatability for species in the Silwood grassland (Crawley, 1990), this finding is con-sistent with the prediction of competitor release of unpalatable species when exposed to rabbit grazing (Pacala and Crawley, 1992). The quite low levels of palatability found for P. annua and H. lanatus agree with previous work which suggests that, compared to other grasses, these are avoided by rabbits (Fenton (1940) and Gillham (1955) for H. lanatus and Ham-bler et al. (1995) for P. annua). For other species with intermediate palatabilities, the relationship was weaker, perhaps because the direct effect of grazing is less important than other direct and indirect effects of rabbits. For example, P. annua is far more abun-dant in heavily grazed areas despite its intermediate palatability to rabbits. P. annua germinates more read-ily in light conditions on disturbed ground (Grime
Fig. 3. The relationship between a plant species’ palatability to rabbits and the ratio of its abundance (% cover) in grazed and ungrazed sites.
study is less reliable than for other species as the vari-ance of these data was relatively high.
The precise interaction of factors which determines feeding preference for different plant species by rab-bits is unknown and may vary over time (Chapuis and Forgeard, 1982). Roles have been suggested for tis-sue protein content (Beaumont et al., 1933; Burton et al., 1964; Myers and Bults, 1977) and for chemical and physical defences (Burton et al., 1964). Such de-fences may become more pronounced as the plant ages (Beaumont et al., 1933; Burton et al., 1964). Also, it is known that tissue damage caused by rabbit grazing may induce the production of chemical defences, such as glucosinolates, in some species (Macfarlane Smith et al., 1991). In this study, the plants in the field var-ied in age and included flowering adults. By contrast, those in the palatability trial were at a pre-flowering stage of development and had not been previously sub-jected to herbivore attack. However, despite all of these possible sources of variation, results from this study
range of background vegetation types. There is con-siderable anecdotal evidence that suggests that some species are consistently ignored by rabbits if there is alternative food, e.g., Urtica dioica, Cirsium spp. and S. jacobaea, whilst others, including many grasses, appear to be consistently palatable (reviewed in Tans-ley, 1949; Thompson and Worden, 1956; Thompson, 1994). Despite this, the degree of consistency found in this study could be considered as surprising as in-fluences on relative palatability have been detected in captive rabbits. These include pre-exposure to the plant species either directly or through maternal trans-mission (Bilkó et al., 1994) and effects of specific plant species’ combinations when the palatability of a number of species is measured by offering only pairs of species at any one time (Soane, 1980). It may be that pre-exposure also affects palatability in field con-ditions but that the range of plants eaten by the wild rabbits was so large that this effect was not sufficient to be detected in the present field study. Likewise, it may be that the palatability of a species was not greatly affected by the nature of its adjacent ‘back-ground’ species in this study because the rabbits were free ranging and had ample food sources.
5. Conclusions
This study indicates that palatability data can be used as a robust measure of relative herbivory of plant species by wild rabbits across a range of vegetation growing on ex-arable land. The extent to which rel-ative herbivory determines total floristic composition depends on the degree of influence of other factors, particularly colonisation events and successional pro-cesses. Consequently, estimates of relative herbivory may be more useful for evaluating the effect of rabbit grazing on the floristic composition of established grassland than for predicting the effect of rabbits on the floristic composition of early successional stage, self-sown vegetation. Such information on the extent to which different grassland species are eaten by rab-bits may be valuable in deciding whether a change in the incidence of rabbits will produce desirable changes in the vegetation. Estimates of relative her-bivory produced from relative palatability data may also be of considerable value in predicting the effect of rabbits on the plagioclimax floristic composition
of grasslands sown with known mixes of seed. Palata-bility trials could be used to assess the palataPalata-bility of both sown and potentially invasive species and this information could then be used to inform grazing management decisions.
Acknowledgements
I thank the land owners, estate managers and ten-ant farmers of Fintray Estate, Harthill Farms and Moray Estates for granting me access to the set-aside land surveyed. I also thank Dr M.A. Ford for useful discussion and Dr J. Warren, Mr F. Wilson and two anonymous referees for their helpful comments on a draft of this manuscript. This research was funded by a research grant from the Leverhulme Trust, receipt of which is gratefully acknowledged. The Scottish Agricultural College receives financial support from the Scottish Office Agriculture, Environment and Fisheries Department.
References
Beaumont, A.B., Stitt, R.E., Snell, R.S., 1933. Some factors affecting the palatability of pasture plants. J. Am. Soc. Agron. 25, 123–128.
Bhadresa, R., 1977. Food preferences of rabbits Oryctolagus cuniculus L. at Holkham sand dunes, Norfolk. J. Appl. Ecol. 14, 287–291.
Bilkó, Á., Altbäcker, V., Hudson, R., 1994. Transmission of food preference in the rabbit: the means of information transfer. Physiol. Behav. 56, 907–912.
Bishop, G.F., Davy, A.J., 1984. Significance of rabbits for the population regulation of Hieracium pilosella in Breckland. J. Ecol. 72, 273–284.
Burton, G.W., Knox, F.E., Beardsley, D.W., 1964. Effect of age on the chemical composition, palatability and digestibility of grass leaves. Agron. J. 56, 160–161.
Chapuis, J.L., Forgeard, F., 1982. Liaisons entre les choix alimentaires du lapin de garenne, Oryctolagus cuniculus (L.), et la phénologie de la végétation sur des pelouses et des landes incendiées. Acta Œcol. 3, 495–510.
Clapham, A.R., Tutin, T.G., Moore, D.M., 1987. Flora of the British Isles, 3rd ed. Cambridge University Press, Cambridge. Crawley, M.J., 1990. Rabbit grazing, plant competition and seedling recruitment in acid grassland. J. Appl. Ecol. 27, 803– 820.
Dixon, J.M., Hambler, D.J., 1993. Wildlife and reclamation ecology: rabbit middens on seeded limestone quarry-spoil. Environ. Cons. 20, 65–73.
Farrow, E.P., 1917. On the ecology of the vegetation of Breckland III. General effects of rabbits on the vegetation. J. Ecol. 5 1–18.
Fenton, E.W., 1940. The influence of rabbits on the vegetation of certain hill-grazing districts of Scotland. J. Ecol. 28 438–449.
Foran, B.D., 1986. The impact of rabbits and cattle on an arid calcareous shrubby grassland in central Australia. Vegetatio 66, 49–59.
Gillham, M.E., 1955. Ecology of the Pembrokeshire islands III. The effect of grazing on the vegetation. J. Ecol. 43, 172–206. Grime, J.P., Hodgson, J.G., Hunt, R., 1988. Comparative Plant Ecology. A Functional Approach to Common British Species. Unwin Hyman, London.
Hambler, D.J., Dixon, J.M., Hale, W.H.G., 1995. Ten years in rehabilitation of spoil: appearance, plant colonists, plant colonists and the dominant herbivore. Environ. Cons. 22, 323– 334.
Macfarlane Smith, W.H., Griffiths, D.W., Boag, B., 1991. Overwinter variation in glucosinolate content of green tissue of rape (Brassica napus) in response to grazing by wild rabbit (Oryctolagus cuniculus). J. Sci. Food Agric. 56, 511–521. Mather, K., 1966. Statistical Analysis in Biology. Methuen,
London, pp. 201–202.
Myers, K., Bults, H.G., 1977. Observations on changes in the quality of food eaten by the wild rabbit. Austr. J. Ecol. 2, 215–229.
Pacala, S.W., Crawley, M.J., 1992. Herbivores and plant diversity. Am. Nat. 40, 243–260.
Prins, A.H., Nell, H.W., 1990. Positive and negative effects of herbivory on the population dynamics of Senecio jacobaea L. and Cynoglossum officinale L. Oecologia 83, 325–332.
Ranwell, D.S., 1960. Newborough Warren, Anglesey III. Changes in the vegetation on parts of the dune system after the loss of rabbits by Myxomatosis. J. Ecol. 48, 385–395.
Soane, G.A., 1980. Food selection by the rabbit. Ph.D. Thesis, University College of North Wales, Bangor.
Southern, H.N., 1940. Ecology and population dynamics of the wild rabbit (Oryctolagus cuniculus). Ann. Appl. Biol. 27, 509– 527.
Tansley, A.G., 1949. The British Isles and Their Vegetation. Cambridge University Press, Cambridge.
Thomas, A.S., 1960. Changes in vegetation since the advent of myxomatosis. J. Ecol. 48, 287–306.
Thomas, A.S., 1963. Further changes in vegetation since the advent of myxomatois. J. Ecol. 51, 151–183.
Thompson, H.V., 1994. The rabbit in Britain. In: Thompson, H.V., King, C. (Eds.), The European Rabbit: the History and Biology of a Successful Colonizer, Oxford University Press, Oxford, pp. 64–107.
Thompson, H.V., Worden, A.N., 1956. The Rabbit. Collins, London, p. 98.
Watt, A.S., 1957. The effect of excluding rabbits from grassland B (Mesobrometum) in Breckland. J. Ecol. 45, 861–878. Watt, A.S., 1960. The effect of excluding rabbits from acidiphilous
grassland in Breckland. J. Ecol. 48, 601–604.
Watt, A.S., 1962. The effect of excluding rabbits from grassland A (Xerobrometum) in Breckland, 1936–60. J. Ecol. 50, 181–198. Watt, A.S., 1981. A comparison of grazed and ungrazed grassland
A in East Anglican Breckland. J. Ecol. 69, 499–508. Williams, S.S., 1951. Microenvironment in relation to experimental
techniques. J. Br. Grass. Soc. 6, 207–217.
Williams, O.B., Wells, T.C.E., Wells, D.A., 1974. Grazing management of Woodwalton Fen: seasonal changes in the diet of cattle and rabbits. J. Appl. Ecol. 11, 499–516.