Oseana, Volume XXV, Nomor 2, 2000 : 1-11 ISSN 0216-1877 EPIFIT PADA LAMUN
oleh Muhammad Husni Azkab 1)
ABSTRACT
EPIPHYTES ON SEAGRASS. Epiphytes is any organism that lives upon a
plant, whether or not a nutritional relationship exists. Epiphytes of seagrasses consist of flora (macro and micro algae), fauna, bacteria and detritus. This paper will describe flora and fauna epiphytes on seagrasses in different parts of the world.
PENDAHULUAN
Secara umum epifit dapat diartikan adalah setiap organisme yang hidup di sebuah tanaman, ada atau tidak ada hubungannya dengan nutrisi yang ada. Pada beberapa literatur sejumlah tumbuhan dan hewan epifit pada lamun telah disusun dan diketengahkan yang berkaitan dengan assosiasi epifit dengan inangnya (host). Beberapa hasil percobaan telah menunjukkan adanya hubungan antara epifit dengan inangnya tersebut (HARLIN 1980).
Tulisan ini merupakan resume flora dan fauna epifit lamun dari belahan dunia.
EPIFIT-EPIFIT PADA LAMUN I. Flora
I.I. Makroalgae
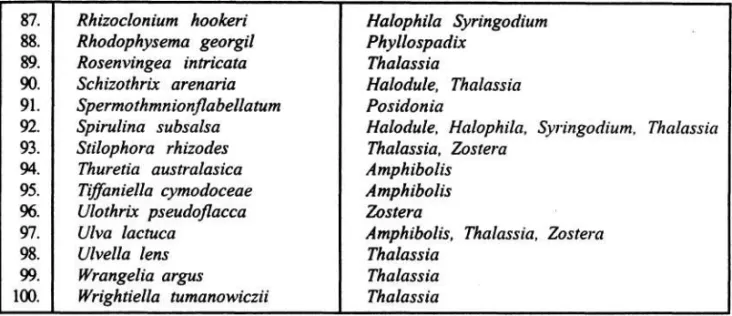
Pada Tabel 1 ditampilkan epifit makroalgae pada lamun. Pada awal abad ke-20 Davis tahun 1913 (HARLIN 1980) telah
mempublikasikan sebuah daftar makroalgae yang tumbuh pada Zostera marina pada suatu area di Massachusetts, Amerika Serikat. Ada sekitar 48 jenis epifit telah dikoleksi dari permukaan daun lamun tersebut pada saat surut rendah dari tujuh lokasi air payau. Walaupun ada perbedaan salinitas dan pergerakan air pada dua habitat yang berbeda, tetapi tidak ada jenis epifit yang tumpang- tindih. Algae hijau merupakan jenis yang dominan, dimana algae coklat dan merah sangat umum pada perairan terbuka. HUMM (1964) telah mencatat epifit dari Thalassia testudinum di Florida. Dari kurang lebih 100 jenis epifit yang ditemukan, hanya beberapa jenis epifit yang terdapat pada lamun, yang umumnya adalah algae coklat. Jenis epifit yang dikoleksi sangat umum terdapat pada inang tanaman lamun dan pada substrat. Lebih lanjut HUMM (1964) melaporkan bahwa ada 19 epifit adalah algae coklat, sedangkan EARLE (1969) melaporkan ada 27 jenis algae cokiat hidup pada T. testudinum, Halodule wrightii di bagian timur Teluk Mexico. Pada
bagian barat pantai Florida, BALANTINE & HUMM (1975) memperoleh 66 jenis epifit algae bentik pada T. testudinum, H. wrightii, Syringodium filiforme dan Halophila engelmannii. Distirbusi epifit tersebut adalah Rhodophyta 30 jenis, Phaephyta 8 jenis dan Chlorophyta dan Cyanophyta masing-masing 14 jenis.
Posisi epifit makroalgae pada daun lamun telah diteliti oleh Van Den ENDE & HAAGE (1963). Sejumlah epifit makroalgae terdapat pada daun muda dari Zostera ma- rina. Distribusi epifit terutama ditemukan dekat dasar pada daun muda. Pada daun tua, epifit banyak pada daerah daun antara 20- 30cm. Pada rimpang (rhizome) tidak ditemukan epifit. Lebih lanjut penelitian Van Der BEN (1969) dari daun Posidonia oceanica pada dua lokasi di pantai Prancis, Laut Mediterranian telah terindentifikasi 90 jenis epifit yang terdiri dari Rhodophyta 60 jenis, Phaephyta 22 jenis dan Chlorophyta 8 jenis.
DUCKER et al. (1977) telah menemukan 105 epifit algae pada lamun Amphibolis antartica dan A. griffithii yang merupakan jenis lamun endemik di perairan Australia (Table 1). Banyak dari epifit alge tersebut ditemukan pada rhizome yang hadir sepanjang tahun, dimana banyak dari epifit tersebut yang hidup musiman di daun.
1.2. Mikroalgae
Pada Tabel 2 disajikan epifit mikroalgae. Pada suatu goba di daerah payau, WOOD (1959) membandingkan komunitas mikroalgae dari lamun dan makroalgae dengan permukaan lumpur dan sekitar kolom air. Jumlah jenis yang dikumpulkan terdiri dari 39 epifit, 45 fitoplankton dan 63 algae lumpur. Jumlah ini menunjukkan bahwa dengan hanya pada satu lokasi, jenis lebih sedikit dari yang hidup kolom air atau daerah dasar. Kerapatan diatomea pada lamun lebih besar dari sekitar dinding karang. Jenis diatomea ini ditemukan pada usus ikan yang merupakan jenis epifit
yang biasa ditemukan pada substrat lumpur. Hal ini menunjukkan bahwa algae ini penting sebagai makanan hewan tersebut.
KITA & HARADA (1962) memban-dingkan komposisi jenis fitoplankton pada padang lamun Zostera dengan mikroalgae pada helai daun kira-kira 70 cm dari dasar pada inang pada tiga sampling stasiun dekat Seto, Jepang. Walaupun ada variasi dari satu lokasi ke lokasi lain dan dari satu tanaman ke tanaman lain, jenis dari bentuk fitoplankton merupakan suatu yang umum dan mikroalgaea epifit memisahkan diri. Hanya sedikit yang tumpang-tindih, sedangkan jenis epifit yang ditampilkan pada Tabel 1 dan Tabel 2 dapat dikatakan sebagai anggota epifit sementara. Lebih lanjut, algae dilaporkan lebih melimpah pada bagian atas dari pada bagian permukaan bawah pada lamun yang dapat melayang dekat horizontal pada waktu mengoleksi epifit tersebut. Seperti pada studi WOOD (1959), jumlah algae permeter luas permukaan (1,1 -3,7 x 108) lebih kecil dari bawah substrat lumpur (9,8 x 109) (MOUL & MASON 1957). Mayoritas epifit adalah diatomea, umumnya adalah
Cocconeis scutellum dan Nitscheia
longissima. Biomassa akan meningkat yang menuju ke puncak helai daun, rata-rata 0,1 mg berat kering/cm2.
Bertolak belakang antara mikroalgae dari daun Zostera dan algae besar seperti yang dilaporkan KITA & HARADA (1962), yang diamati oleh MAIN & MclNTIRE (1974) di Muara Yaquina, Oregon, dimana flora pada lamun dan batu karang sebenarnya tidak dapat dibedakan. Lebih lanjut, komposisi jenis dari Z. marina tidak ada perbedan yang berarti dari Entervmorpha, Poliyshiponia dan
Ulva, makroalgae yang didapatkan pada area yang sama. Hal ini perbedaannya lebih besar dari individu ke individu dari jenis inang ke jenis inang. Pada perbedaan individu ditemukan adanya perbedaan antara epifit helai daun dengan kolom air, sedimen, pengawetan, dan bagian yang ternaungi. Hal ini menunjukkan bahwa inang makrofita
sebagai dasar terjadinya interaksi. MAIN & McINTIRE (1974) telah mengidentifikasi 221
taxa diatomea, dimana ada 36 taxa yang sangat melimpah (Tabel 2).
BROWN (1962) melaporkan bahwa pada satu musim dari tambak di Inggris mengestimasi produksi bersih epifit 0,24 g / m2/hari dengan membandingkan 2,2 g/m2/hari pada inang, Zostera marina, Produktivitas ini lebih erat hubungannya dengan temperatur dibandingkan dengan faktor lain yang diukur (cahaya, nutrien atau salinitas). Lebih lanjut BROWN (1962) menemukan jenis-jenis epifit juga hidup pada sedimen dan kolom air.
II. BAKTERI DAN DETRITUS
Detritus sering dilaporkan pada lamun, dan kehadiran epifit flora dan fauna. Penelitian terhadap permukaan daun Zostera dengan elektron mikroskopis (scan electron microgra-phy) menunjukkan adanya detritus, bakteri kokos dan filamen, diatomea (SIEBURTH & THOMAS 1973). Kolonisasi dari mikroorganisme tidak selektif, dan bentuk bakteri lebih kecil bila dibandingkan dengan diatomea.
Tidak seperti bakteri yang mengurai mated dengan baik, dan hanya 2% dari fotosintesis yang diserap sebagai karbon organik (BRYLINSKY 1971). Tetapi sekali proses penguraian berjalan, materi organik yang dikeluarkan pada bakteri dapat menghasilkan tepung (karbohidrat) (BELL & MITCHELL 1972).
III. FAUNA
Pada Tabel 3 ditampilkan epifit fauna. Grup dominan dari invertebrata yang hidup pada Zostera di muara sungai York, Virginia telah diteliti oleh MARSH (1973). Dari seluruh fauna yang diseleksi, pada empat taxon utama adalah: Gastropoda, 23 jenis dengan populasi 43,2%; Amphipoda, 23 jenis dengan populasi 18,5%; Isopoda, 4 jenis dengan populasi 16,7%; dan Polikhaeta, 18 jenis dengan populasi 15%. Pada penelitian tersebut
ditemukan 5 jenis epifit yang dominan yaitu;
Bittium varium, Paracerceis caudata, Crepidula convexa, Ampithoe longimana,
dan Erichwnella attenuata. Lebih lanjut pada penelitian yang berdasarkan kedalaman, maka diversitas jenis lebih besar pada lokasi yang lebih dalam. Jumlah jenis lebih sedikit yang ditemukan pada musim dingin (winter) jika organisme tersebut masuk ke dalam sedimen, tetapi tidak ada perbedaan dalam distribusi- nya. Jika lebih 30% kelimpahan pada komunitas
Zostera dibandingkan dengan yang ditemukan di sekitarnya, hanya Odostomia bisuturalis
yang umum ditemukan pada kedua habitat tersebut. Jadi populasi fauna jelas dan homogen. Kesimpulan ini juga berlaku pada padang Thalassia testudinum di Florida yang diteliti oleh MARSH (1973) dan padang lamun di Inggeris dan Prancis yang diteliti oleh LEDOYER (1962). Sumber makanan primer dari epifit fauna adalah epifit flora. Jika lamun mengandung banyak epifit, maka beratnya akan naik dan cahaya akan menurun. Hal ini akan memudahkan patahnya helai daun dan terlepas sehingga hanyut ke lain tempat.
Pada Tanjung Cod, Massachusetts, NAGLE (1968) memisahkan epifit fauna dari daun Zostera. Secara umum, fauna yang dekat pada dasar tanaman akan menghuni di dasar dan menghabiskan sebagian waktu hidupnya sebagai epifit, sedangkan yang dekat ujung merupakan pemakan suspensi yang menangkap makanannnya dari kolom air. Lebih lanjut NAGLE (1968) menunjukkan adanya mutualisme antara flora epifit dan satu elemen dari epifit fauna. Amphipoda yang memakan algae akan dimakan oleh ikan predator (Fun- dulus) dan amphipoda akan membersihkan detritus dari algae. Karena kedua komponen tersebut saling membutuhkan, maka NAGLE (1968) mengatakan bahwa assosiasi ini adalah "protocooperation".
NISHIHIRA (1968) mengamati distribusi hydrozoa pada daun Zostera ma-rina dan Phyllospadix iwatensisi di Jepang.
Hasil penelitiannya menunjukkan bahwa Clytia edwardsi mulai hidup pada bagian daun tua dan kerapatan epifit meningkat dan membuat habitatnya meluas, kemudian populasi menyebar pada bagian daun muda (tengah dan ujung daun). Orientasi pertumbuhan pada bagian daun muda ini telah diamati pada perkapuran algae merah, Fosliella zostericola
dan invertebrata, Spirorbis and Hydroides.
Kedua jenis inang ini dikoleksi pada lokasi yang berbeda.
Perbedaan distribusi dari epifit invertebrata juga telah dilaporkan oleh KITA & HARADA (1962). Pada ujung Z. marina
telah ditemukan melimpahnya Diala vitrea
dan Boloceroides mcmurrichi, dan pada bagian dasar ditemukan Spirorbisforminosus
dan Caprella kroyeri. Berdasarkan pengamatan di daerah perairan dangkal (sub- littoral), pada Thalassia dan Halodule di Florida, O'GOWER & WACASEY (1967) menenemukan adanya perbedaan distribusi epifit invertebrata yang berbeda pada perairan yang bergerak (tidal flow).
KONTRIBUSI EPIFIT ALGAE
Epifit algae hijau telah dilaporkan untuk mengfiksasi nitrogen dengan kecepatan dari 2,4-16,5 ug N/mg tanaman-N/h dalam cahaya (GOERING & PARKER 1972). Pengkuran ini telah dilakukan pada algae hijau pada lamun
T. testudinum, S. filiforme, H. wrightii dan
Ruppia maritima di Teluk Redfish, Texas. M c R O Y e t a l . ( 1 9 7 3 ) t e l a h m e n g u j i
T. tesdtudinum dan S. filiforme dari Floruida dan Z marina dari Carolina Utara dan Alaska. Mereka melaporkan hasil yang bervariasi, tetapi hasilnya semuanya rendah (0,5%), dan menyimpulkan bahwa sumber nitrogen pada lamun masih dalam pertanyaan besar.
Kontribusi epifit algae untuk total produktivitas pada komunitas lamun telah diteliti oleh JONES (1986) dan PENHALE (1977) dengan perbedaan metoda. Jones menggunakan "differential manometry" untuk mengukur volume gas basil fotosintesa dari komponen epifit. Dan data menunjukkan perkiraan kecepatan 700ml O2/hari atau 18 kg
tanaman berpembuluh/m2/tahun, pada komunitas yang padat. Perbedaan kecepatan ditemukan karena adanya perbedaan irradiasi dan biomassa. Tetapi secara umum 1/4 - 1/3 dari metabolisme komunitas berasal dari epifit flora yang juga dapat dijadikan sumber makanan bagi hewan herbivora.
PENHALE (1977) telah meneliti produktivitas epifit dari tanaman inang pada periode lebih 12 bulan dengan menggunakan
14
C-CO2. Kontribusi epifit algae sekitar 24%
dari biomassa (24,7 g berat kering/m2). Pada setiap satu gram, respon fotosintesa dari epifit algae (0,88mg C/g/jam) lebih setara dengan tanaman inang (0,65 mg C/gram/jam). Tetapi hal ini ada perbedaan dalam biomaassa. Total kontrobusi epifit diperkirakan 18%/tahun.
SAND-JENSEN (1977) telah meneliti efek adanya epifit dalam proses fotosintesa pada tanaman inang dengan mengetahui konsentrasi CO2 dan intensitas cahaya. Dia
telah menemukan setengah jenuh untuk penyerapan 14C-CO2 oleh Zostera marina dan
tanpa adanya epifit. Algae yang utama adalah
diatomea, Cocconeis scutellum yang
menggunakan karbon konsentrasi rendah dari CO2 dan intensitas cahaya yang rendah dari
tanaman inang. (SAND-JENSEN (1977) menemukan adanya reduksi fotosintesa 31%, sehingga dia menyimpulkan bahwa adanya efek lebih besar karena adanya organisme epifit algae tersebut.
DAFTAR PUSTAKA
BALLANTINE, DL. and H.J. HUMM 1977. Benthic algae of the Anclote estuary. I. Epiphytes of seagrass leaves. Florida Sci. 38: 144-149.
BEUU W. and R. MITCHELL 1972. Chemotac- tic and growth responses of marine bacterial to algal extracellular products.
Biol. Bull. 143 : 265-277.
BROWN, C.L. 1962. On the ecology of aufwuchs of Zostera marina in Charlestown Pond, Rhode Island. Master's thesis, University of Georgia. Athena, Ga, 90 pp.
DUCKER, S.C., NJ.F000RD and R.B. KNOX 1977. Biology of Australian seagrasses; the genus Amphibolis C. Agardh (Cymodoceaeae). Aust. J. Bot. 25:67- 95.
EARLE, S.A. 1969. Phaeophyta of the eastern Gulf of Mexico. Phycologia 7:71-254. GOERING, J.J. and P.L. PARKER 1972.
Nitro-gen fixation by epiphytes on seagrasses. Limnol Oceanogr. 17: 320- 323.
HARLIN, M.M. 1980. Seagrass epiphytes. In : Handbook of seagrass biology: an ecosystem perspective (RC. Phillips & C.P. McROY, eds.). Garland STPM Press. New York 117-151.
HUMM, H.J. 1964. Epiphytes of the seagrass,
Thalassia testudinum, in Florida. Bull. Mar. Sci. Gulf Carib. 14 : 306-341. JONES, J.A. 1968. Primary productivity by the
tropical marine turtle grass Thalassia testudinum Konig, and its epiphytes. Doctoral dissertation, University of Miami, Fla. 196 pp.
KITA, T. and E. HARADA 1962. Studies on the epiphytes communities. L Abun-dance and distribution of microalgae and small animals on the Zostera
blades. Publ. Seto Mar. Biol Lab. 10 : 245-257.
McROY, C.P., J.J. GOERING and B. CHANEY 1973. Nitrogen fixation associated with seagrasses. Limnol. Ocenogr. 18 : 998- 1002.
MAIN, S.P. and CD. McINTIRE 1974. The distribution of epiphytic diatoms in Yaquina estuary, Oregon (USA). Bot. Mar. 17: 88-99.
MARSH, GA. 1973. The Zostera epifaunal community in the York River, Virginia.
Chesapeake Sci 14: 87-97.
MOUL, E.T. and D. MASON 1957. Study on diatom population on sand and mud flats in the Woods Hole area. Biol. Bull. 113:351.
NAGLE, J.S. 1968. Distribution of the epibiota of macroepibenthic plants. Univ. Texas. Contri. Mar. Sci. 13: 105-1 14.
NISHIHIRA, M. 1968. Distribution pattern of Hydrozoa on the broad-leaved eel- grass and narrow-leaved eelgrass. Bull Biol. Sm. Asamushi 13: 125-138. O'GOWER, A.K., and J.W. WACASEY 1967.
Animal communities associated with
Thalassia (Diplanthera) and sand beds in Biscayne Bay. I. Analysis of com-munities in relation to water move-ments. Bull. Mar.Sci. 17: 175-219. PENHALE, PA. 1977. Macrophyte-epiphyte
biomass and productivity in an eel- grass (Zostera marina L). community. J. Exp. Mar. Biol. Ecol. 26:211-224.
SAND-JENSEN, K. 1975. Effect of epiphytes on eelgrass photosynthesis. Aquat. Bot 3: 55-63.
SIEBURTH, J. McN and CD. THOMAS 1973. Fouling on eelgrass (Zostera marina
L.) J. Phycol 9 : 46-50.
Van Den ENDE, G. and P. HAAGE 1963. Der epiphytenbewuch von Zostera marina
und der bretonischen Kuste. Bot. Mar.
5:105-110.
Van Der BEN, D. 1969. Les epiphytes des feuilles de Posidonia oceanica sur les cotes francaises de la Mediterranee.
Proc. Internal Seweed Symp. (Madrid) 6: 79-84.
WOOD, E.J.F. 1959. Some aspects of the ecology of Lake Macquarie, N.S.W., with regard to an alleged depletion of fish. VI. Plant communities and their significance. Auctr. J. Mar. Freshwater Res. 10: 322-344.
WOOD, E.J.F. 1972. Substratum : unicellular plants. In : Marine ecology, Vol. 1 (0. Kinne ed). Biologische Anstalt Helgoland, Hamburg. 127 1-1276.