PRODUKSI DAN KARAKTERISASI BAKTERIOSIN ASAL
Lactobacillus fermentum 2B2 SERTA AKTIVITAS
ANTIMIKROBANYA TERHADAP
BAKTERI PATOGEN
SKRIPSI
LAMRIA MAGDALENA
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR 2009
RINGKASAN
LAMRIA MAGDALENA. D14053309. 2009. Produksi dan Karakterisasi Bakteriosin Asal Lactobacillus fermentum 2B2 dan Aktivitas Antimikrobanya terhadap Bakteri Patogen. Skripsi. Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Pembimbing Utama : Irma Isnafia Arief, S.Pt., M.Si
Pembimbing Anggota : Dr. Ir. Rarah Ratih Adjie Maheswari, DEA
Bakteri asam laktat (BAL) yang terdapat secara alami di dalam daging atau susu memiliki kemampuan sebagai bahan pengawet alami karena menghasilkan senyawa antimikroba. Salah satu senyawa antimikroba yang dihasilkan yaitu bakteriosin. Penggunaan bakteriosin sebagai biopreservative sangat menjanjikan karena bersifat bakterisidal atau bakteristatik terhadap bakteri lain khususnya bakteri yang berkerabat dekat dengan bakteri penghasilnya (Vuyst and Vandamme, 1994).
Potensi bakteriosin ini meningkatkan penelitian dalam mempelajari dan mendapatkan informasi yang lebih dalam mengenai produksi bakteriosin, purifikasi parsial bakteriosin, konsentrasi penghambatan minimum dalam minimum inhibitor concentration (MIC) dan minimum bactericidal concentration (MBC) terhadap bakteri patogen, serta karakterisasi bakteriosin melalui uji kepekaan bakteriosin terhadap enzim katalase dan proteolitik.
Penelitian ini dilaksanakan di Laboratorium Mikrobiologi Bagian Teknologi Hasil Ternak Perah dan Laboratorium Ruminansia Besar, Fakultas Peternakan, Institut Pertanian Bogor. Penelitian dilaksanakan selama satu tahun dari bulan September 2008 sampai September 2009. Penelitian ini menggunakan satu isolat BAL yang berasal dari daging sapi segar yaitu Lactobacillus fermentum 2B2 serta empat bakteri uji (Staphylococcus aureus ATCC 25928, Salmonella typhimurium ATCC 14028, Escherichia coli ATCC 25922, dan enteropatogenik Escherichia coli lokal). Peubah yang diamati adalah diameter zona hambat yang terbentuk yang dilakukan dengan metode sumur difusi agar, serta konsentrasi minimum dalam menghambat bakteri indikator.
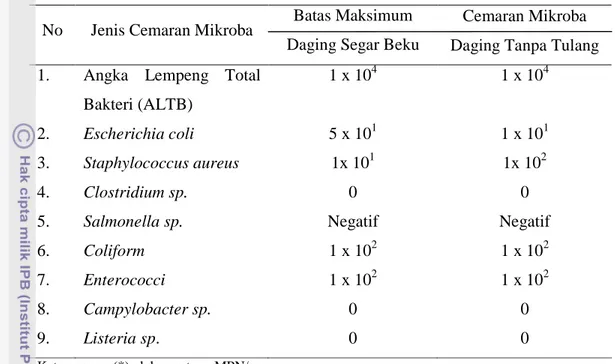
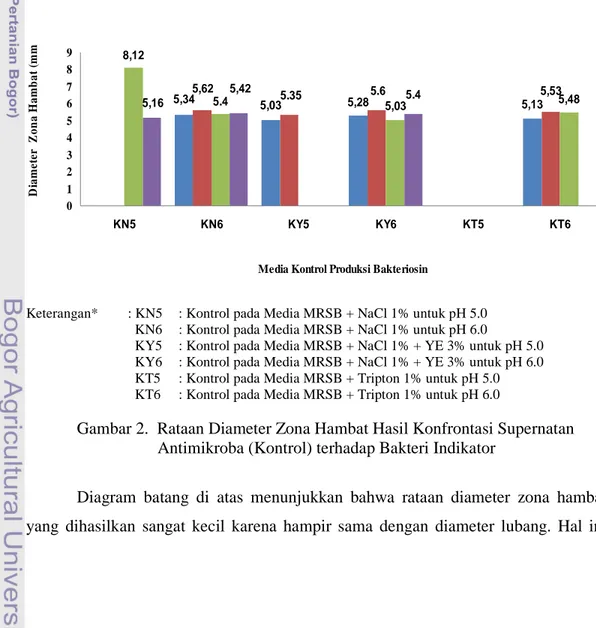
Hasil penelitian ini terdiri dari lima tahap. Hasil tahap produksi bakteriosin didapatkan bahwa respon bakteri indikator terhadap substrat kasar bakteriosin pada media yang berbeda tidak berpengaruh nyata (P>0,05), tetapi ditinjau dari rataannya media tripton paling baik diantara media yang lain. Hasil tahap purifikasi parsial bakteriosin juga didapatkan hasil yang tidak berbeda (P>0,05). Namun, rataan tertinggi diperoleh saat menggunakan media tripton. Pada tahap kedua ini juga didapatkan hasil bahwa zona hambat terbesar terjadi pada bakteri indikator S. aureus. Hasil pada tahap ke tiga didapatkan bahwa MIC terjadi pada konsentrasi 70%, sedangkan MBC terjadi pada konsentrasi 90%. Hasil pada tahap uji bakteriosin terhadap enzim katalase menunjukkan bahwa bakteriosin tanpa pengaruh hidrogen peroksida masih dapat membentuk zona hambat. Pada tahap kepekaan bakteriosin terhadap enzim proteolitik menunjukkan bahwa ikatan peptida bakteriosin terhidrolisis oleh enzim tripsin, tetapi tidak oleh enzim pepsin.
ABSTRACT
Production and Characterization of Bacteriocin from Lactobacillus fermentum 2B2 and Its Antimicrobial Activities Against Pathogen Bacteria
Magdalena, L., I. I. Arief, and R. R. A. Maheswari
Lactic acid bacteria (LAB) occur naturally in several raw materials like beef and milk. LAB produce antimicrobial substance, such as bacteriocin. Bacteriocin is proteinaceous in nature and bactericidal or bacteristatic against other, mostly closely related bacteria. Lactobacillus fermentum 2B2 isolated from fresh beef. The objectives of the research were to study the best media for Lactobacillus fermentum 2B2 to produce bacteriocin and to know antimicrobial activity of crude bacteriocin against Staphylococcus aureus ATCC 25923, Salmonella typhimurium ATCC 14028, Escherichia coli ATCC 25922, and enteropathogenic Esherichia coli as indicator strains, to know minimum concentration of crude bacteriocin inhibit indicator bacteria in minimum inhibitor concentration (MIC) and minimum bactericidal concentration (MBC) methode and its sensitivity compound with catalase and proteolitic enzyms. The first step showed no significant result happenned from different media (P>0,05), but based on means tripton was the best media. Second step, tripton was the best media based on means and the best antimicrobial activity with the biggest inhibitor zone was against S. aureus. MIC showed in 70% concentration of crude bacteriocin, while in MBC showed in 90% concentration of crude bacteriocin to inhibit S. aureus. Bacteriocin of Lactobacillus fermentum 2B2 hidrolated with tripsyn, but no with pepsin.
PRODUKSI DAN KARAKTERISASI BAKTERIOSIN ASAL
Lactobacillus fermentum 2B2 SERTA AKTIVITAS
ANTIMIKROBANYA TERHADAP
BAKTERI PATOGEN
LAMRIA MAGDALENA D14053309
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR 2009
PRODUKSI DAN KARAKTERISASI BAKTERIOSIN ASAL
Lactobacillus fermentum 2B2 SERTA AKTIVITAS
ANTIMIKROBANYA TERHADAP
BAKTERI PATOGEN
Oleh:
LAMRIA MAGDALENA D14053309
Skripsi ini telah disetujui dan disidangkan di hadapan Komisi Ujian Lisan pada tanggal 23 Oktober 2009
Pembimbing Utama Pembimbing Anggota
Irma Isnafia Arief, S.Pt, M. Si Dr. Ir. Rarah R. A. Maheswari, DEA
Dekan Ketua Departemen
Fakultas Peternakan Ilmu Produksi dan Teknologi Peternakan Institut Pertanian Bogor Fakultas Peternakan
Institut Pertanian Bogor
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 6 Mei 1987 di Bogor. Penulis adalah anak tunggal dari pasangan M. Limbong dan Dinaria Simamora. Riwayat pendidikan Penulis dimulai dari Taman Kanak-Kanak Candra Kirana Bogor (1992-1993), Sekolah Dasar Negeri Lawanggintung I Bogor (1993-1999), Sekolah Menengah Pertama Negeri 7 Bogor (1999-2001), Sekolah Menengah Pertama Negeri 1 Tanjung Morawa (2001-2002), Sekolah Menengah Umum Negeri 5 Medan (2002-2005). Penulis kemudian masuk Institut Pertanian Bogor melalui jalur SPMB (Seleksi Penerimaan Mahasiswa Baru) pada tahun 2005 dan terdaftar sebagai mahasiswa pada Program Studi Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan pada tahun berikutnya.
Selama menjadi mahasiswa IPB, Penulis aktif dan menjadi pengurus periode 2007-2008 dalam Unit Kegiatan Mahasiswa (UKM) Paduan Suara Agria Swara. Penulis aktif dan menjadi pengurus periode 2007-2008 dalam UKM Persekutuan Mahasiswa Kristen (PMK). Penulis juga berpartisipasi dalam berbagai jenis kepanitiaan yang diadakan di kampus.
Sebagai salah satu syarat untuk memperoleh gelar sarjana pada Fakultas Peternakan, Institut Pertanian Bogor, Penulis melakukan penelitian selama 1 tahun dengan judul “Produksi dan Karakterisasi Bakteriosin Asal Lactobacillus fermentum 2B2 serta Aktivitas Antimikrobanya terhadap Bakteri Patogen”.
KATA PENGANTAR
Puji, hormat, dan syukur Penulis persembahkan kepada Tuhan Yesus atas segala berkat, kasih, serta tuntunanNya, khususnya dalam menyelesaikan penelitian dan penulisan karya ilmiah ini. Penulis juga mengucapkan terima kasih kepada semua pihak yang telah mendukung dan membantu dari segi materi, tenaga, serta doa sehingga skripsi dengan judul “Produksi dan Karakterisasi Bakteriosin Asal Lactobacillus fermentum 2B2 serta Aktivitas Antimikrobanya terhadap Bakteri Patogen” dapat diselesaikan.
Perkembangan pasar yang mengikuti permintaan konsumen terhadap pangan sehat dan tidak mengandung bahan pengawet sintetis meningkatkan perhatian penelitian terhadap bahan pengawet alami, salah satunya penggunaan bakteriosin sebagai biopreservatif. Bakteriosin yang berasal dari bakteri asam laktat (BAL) telah terbukti aman untuk dikonsumsi dan dapat memperpanjang masa simpan produk.
Penelitian ini bertujuan untuk mempelajari dan mendapatkan informasi yang lebih, khususnya dalam produksi, purifikasi parsial bakteriosin, konsentrasi penghambatan minimum (MIC dan MBC) terhadap bakteri patogen, dan karakterisasi bakteriosin kasar melalui uji kepekaan terhadap enzim katalase dan proteolitik dari BAL yang belum diteliti seperti Lactobacillus fermentum 2B2 yang berasal dari daging.
Bogor, November 2009
DAFTAR ISI
Halaman
RINGKASAN ... i
ABSTRACT... ii
RIWAYAT HIDUP ... iii
KATA PENGANTAR ... iv
DAFTAR ISI... v
DAFTAR TABEL ... vii
DAFTAR GAMBAR... viii
DAFTAR LAMPIRAN... ix PENDAHULUAN ... 1 Latar Belakang ... 1 Tujuan... 2 TINJAUAN PUSTAKA... 3 Mikrobiologi Daging ... 3
Bakteri Asam Laktat ... 4
Lactobacillus... 5 Antimikroba ... 5 Asam Organik ... 6 Hidrogen Peroksida... 6 Bakteriosin ... 7 Bakteri Patogen... 8 Staphylococcus aureus... 8 Escherichia coli ... 9 Salmonella typhimurium ... 10 Enzim ... 10 Enzim Katalase... 12 Enzim Proteolitik... 12
Minimum Inhibitory Concentration (MIC) ... 13
METODE... 14
Waktu dan Lokasi ... 14
Materi... 14
Bahan ... 14
Alat... 14
Rancangan ... 14
Prosedur... 15
Produksi Bakteriosin pada Media yang Berbeda... 15
Purifikasi Parsial Bakteriosin... 16
Uji Kepekaan Substrat Kasar Bakteriosin terhadap Enzim
Katalase... 20
Uji Kepekaan Substrat Kasar Bakteriosin terhadap Enzim Proteolitik... 22
HASIL DAN PEMBAHASAN ... 24
Produksi Bakteriosin... 24
Penghambatan Berbagai Bakteri Indikator oleh Substrat Kasar Bakteriosin yang Diproduksi dalam Media yang Berbeda ... 26
Penghambatan Bakteri Staphyloccocus aureus ATCC 25923 oleh Substrat Kasar Bakteriosin yang Diproduksi dalam Media yang Berbeda... 29
Penghambatan Bakteri Salmonella typhimurium ATCC 14028 oleh Substrat Kasar Bakteriosin yang Diproduksi dalam Media yang Berbeda... 30
Penghambatan Bakteri Escherichia coli ATCC 25922 (ETEC) oleh Substrat Kasar B akteriosin yang Diproduksi dalam Media yang Berbeda... 31
Penghambatan Bakteri enteropatogenik Escherichia coli (EPEC) lokal oleh Substrat Kasar Bakteriosin yang Diproduksi dalam Media yang Berbeda ... 32
Purifikasi Parsial Bakteriosin... 33
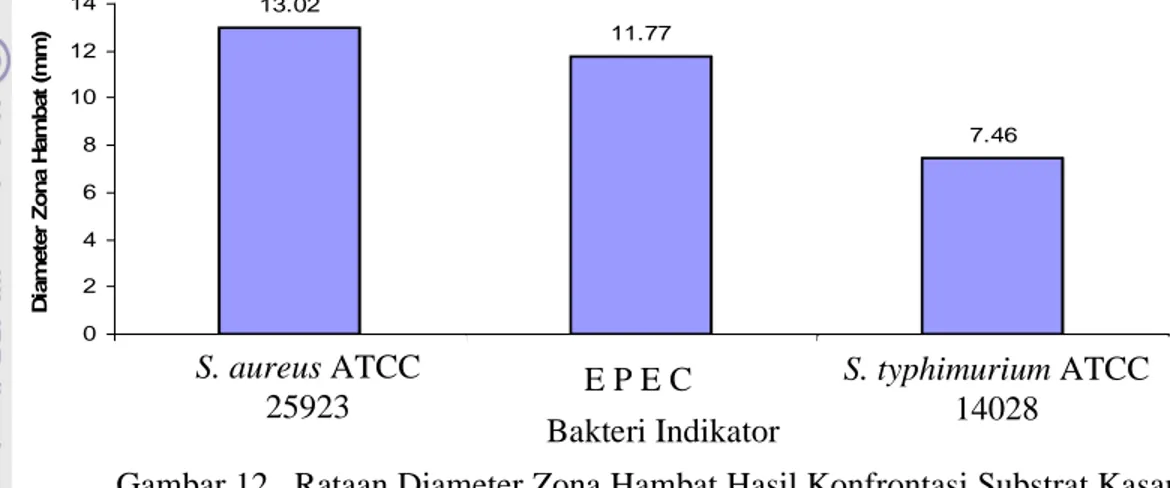
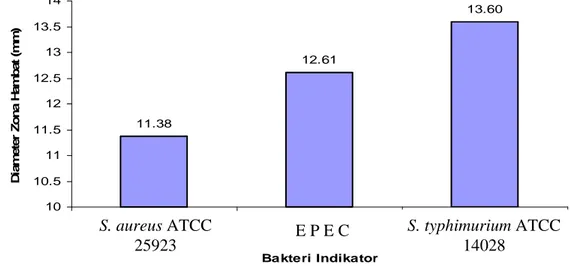
Penghambatan Bakteri Staphyloccocus aureus ATCC 25923 (Bakteri Gram Positif) oleh Substrat Kasar Bakteriosin... 37
Penghambatan Bakteri Salmonella typhimurium ATCC 14028 dan Escherichia coli (EPEC) (Bakteri Gram Negatif) oleh Substrat Kasar Bakteriosin... 38
Konsentrasi Penghambatan Minimum Bakteri Indikator (MIC dan MBC)... 39
Uji Kepekaan Substrat Kasar Bakteriosin terhadap Enzim Katalase... 40
Uji Kepekaan Substrat Kasar Bakteriosin terhadap Enzim Proteolitik .... 41
KESIMPULAN DAN SARAN ... 45
Kesimpulan... 45
Saran…... 45
UCAPAN TERIMA KASIH ... 46
DAFTAR PUSTAKA ... 47
DAFTAR TABEL
Nomor Halaman 1. Batas Maksimum Cemaran Mikroba pada Daging (cfu/g) ... 4
DAFTAR GAMBAR
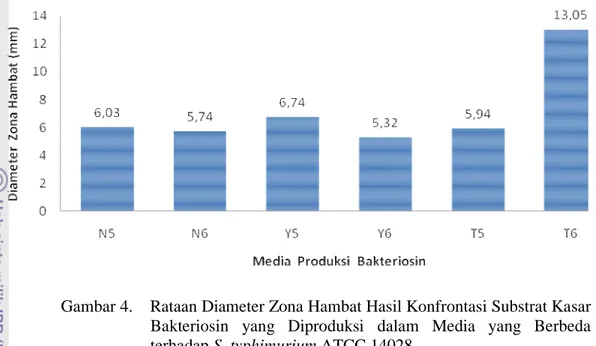
Nomor Halaman 1. Rataan Diameter Zona Hambat Hasil Konfrontasi Substrat Kasar Bakteriosin
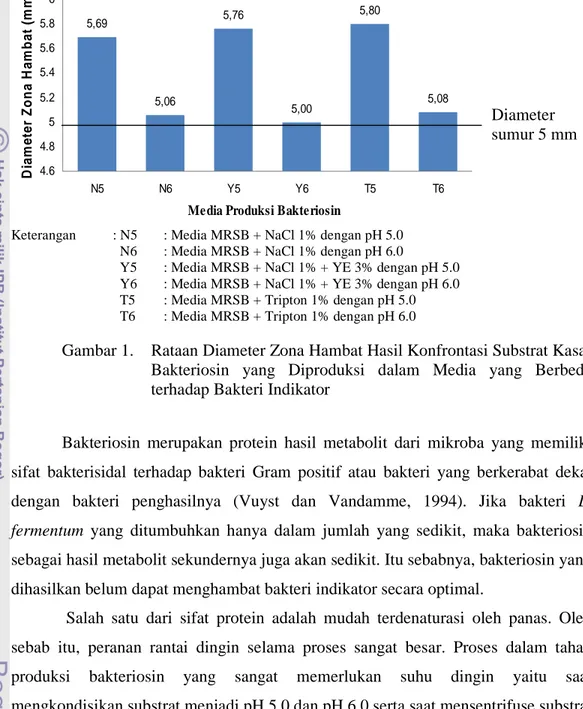
yang Diproduksi dalam Media yang Berbeda terhadap Bakteri Indikator ... . 26 2. Rataan Diameter Zona Hambat Hasil Konrontasi Supernatan Antimikroba
(Media Kontrol) terhadap Bakteri Indikator... 28 3. Rataan Diameter Zona Hambat Hasil Konfrontasi Substrat Kasar Bakteriosin yang Diproduksi dalam Media yang Berbeda terhadap S.aureus ATCC
25923 ... 30 4. Rataan Diameter Zona Hambat Hasil Konfrontasi Substrat Kasar Bakteriosin
yang Diproduksi dalam Media yang Berbeda terhadap S. typhimurium ATCC 14028 ... . 31 5. Rataan Diameter Zona Hambat Hasil Konfrontasi Substrat Kasar Bakteriosin yang Diproduksi dalam Media yang Berbeda terhadap E. coli (ETEC) ATCC 25922 ... . 32 6. Rataan Diameter Zona Hambat Hasil Konfrontasi Substrat Kasar Bakteriosin yang Diproduksi dalam Media yang Berbeda terhadap Enteropatogenik E. coli (EPEC) Lokal... 33
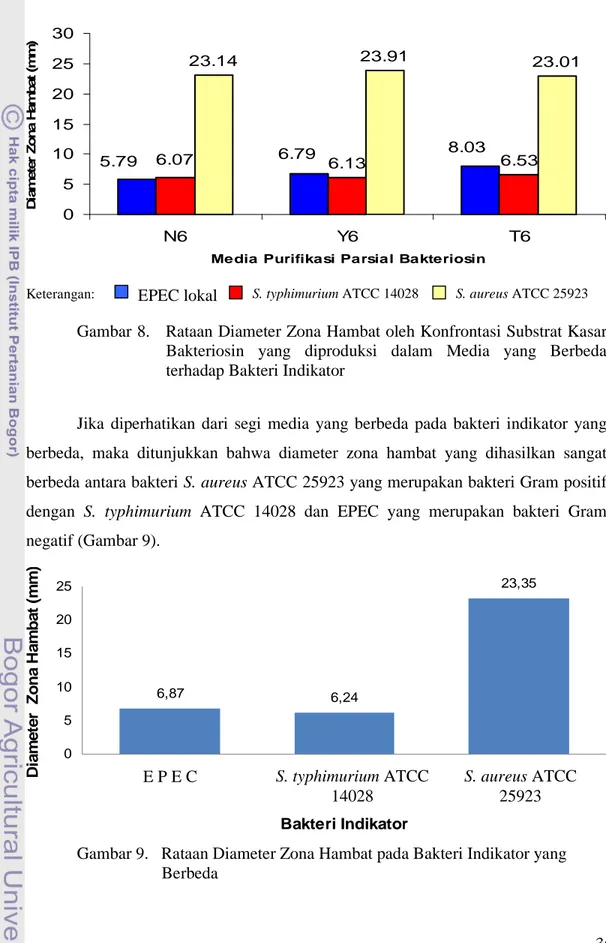
7. Protein Hasil Presipitasi Amonium Sulfat ... . 35 8. Rataan Diameter Zona Hambat oleh Konfrontasi Substrat Kasar Bakteriosin
pada Media yang Berbeda terhadap Bakteri Indikator... . 36 9. Rataan Diameter Zona Hambat pada Bakteri Indikator yang Berbeda... 37 10. Perbedaan Diameter Zona Hambat EPEC (a) dan S. aureus ATCC 25923 (b) pada Media dengan Penambahan Tripton 1%... 37 11. Penghambatan S.aureus ATCC 25923 oleh Substrat Kasar Bakteriosin... 41 12. Rataan Diameter Zona Hambat Hasil Konfrontasi Substrat Kasar Bakteriosin pada Media Tripton dengan Penambahan Enzim Katalase terhadap Bakteri Indikator... 41 13. Rataan Diameter Zona Hambat Hasil Konfrontasi Substrat Kasar Bakteriosin pada Media Tripton dengan Penambahan Enzim Pepsin terhadap Bakteri Indikator... 44 14. Rataan Diameter Zona Hambat Hasil Konfrontasi Substrat Kasar Bakteriosin pada Media Tripton dengan Penambahan Enzim Tripsin terhadap S. aureus ATCC 25923... 45
DAFTAR LAMPIRAN
Nomor Halaman 1. Analisis Ragam Rataan Diameter Zona Hambat Substrat Kasar Bakteriosin
pada Media yang Berbeda terhadap Staphylococcus aureus ATCC 25923
pada Tahap Produksi Bakteriosin... 53 2. Analisis Ragam Rataan Diameter Zona Hambat Substrat Kasar Bakteriosin
pada Media yang Berbeda terhadap Salmonella typhimurium ATCC 14028
pada Tahap Produksi Bakteriosin... 53 3. Analisis Ragam Rataan Diameter Zona Hambat Substrat Kasar Bakteriosin
pada Media yang Berbeda terhadap Escherichia coli (ETEC) ATCC 25922
pada Tahap Produksi Bakteriosin... 53 4. Analisis Ragam Rataan Diameter Zona Hambat Substrat Kasar Bakteriosin
pada Media yang Berbeda terhadap Enteropatogenik Escherichia coli (EPEC) Lokal pada Tahap Produksi Bakteriosin... 54 5. Analisis Ragam Rataan Diameter Zona Hambat Substrat Kasar Bakteriosin
pada Media yang Berbeda terhadap Staphylococcus aureus ATCC 25923
pada Tahap Purifikasi Parsial Bakteriosin... 54 6. Analisis Ragam Rataan Diameter Zona Hambat Substrat Kasar Bakteriosin
pada Media yang Berbeda terhadap Salmonella typhimurium ATCC 14028 pada Tahap Purifikasi Parsial Bakteriosin... 54 7. Analisis Ragam Rataan Diameter Zona Hambat Substrat Kasar Bakteriosin pada Media yang Berbeda terhadap Enteropatogenik Escherichia coli (EPEC) Lokal pada Tahap Purifikasi Parsial Bakteriosin... 55 8. Rataan dan Standar Deviasi Diameter Zona Hambat Substrat Kasar
Bakteriosin pada Media yang Berbeda terhadap Bakteri Indikator pada Tahap Produksi Bakteriosin... 55 9. Rataan dan Standar Deviasi Diameter Zona Hambat Substrat Kasar
Bakteriosin pada Media yang Berbeda terhadap Bakteri Indikator pada
Tahap Purifikasi Parsial Bakteriosin... 55 10. Rataan dan Standar Deviasi Substrat Kasar Bakteriosin pada Media MRSB
dengan Penambahan Tripton 1% terhadap Staphylococcus aureus ATCC
25923 pada Tahap Konsentrasi Penghambatan Minimum (MIC dan MBC).. 55 11. Rataan dan Standar Deviasi Substrat Kasar Bakteriosin pada Media MRSB
dengan Penambahan Tripton 1% terhadap Bakteri Indikator pada Tahap Uji
Kepekaan Enzim Katalase... 56 12. Rataan dan Standar Deviasi Substrat Kasar Bakteriosin pada Media MRSB dengan Penambahan Tripton 1% terhadap Bakteri Indikator pada Tahap Uji Kepekaan Enzim Proteolitik... 56 13. Fermentasi Gula-Gula Sederhana dari Lactobacillus fermentum 2B2... 57
14. Total Asam Tertitrasi Substrat Kasar Bakteriosin pada Tahap Produksi Bakteriosin... 57 15. Kondisi pH Awal dan pH Akhir Substrat Kasar Bakteriosin pada Tahap Purifikasi Parsial Bakteriosin... 57 16. Morfologi Isolat Bakteri Asam Laktat dan Bakteri Indikator... 57 17. Alat sentrifuge 10.000 rpm... 57 18. Zona Hambat Substrat Kasar Bakteriosin terhadap S. aureus ATCC 25923 pada Tahap Produksi Bakteriosin... 58 19. Zona Hambat Substrat Kasar Bakteriosin terhadap enteropatogenik E. coli lokal (Bakteri Gram Negatif) pada Tahap Purifikasi Parsial Bakteriosin... 58 20. Zona Hambat Substrat Kasar Bakteriosin terhadap S. aureus ATCC 25923 (Bakteri Gram Positif) pada Tahap Purifikasi Parsial Bakteriosin... 58 21. Tahap Konsentrasi Penghambatan Minimum (MIC dan MBC)... 59 22. Zona Hambat Substrat Kasar Bakteriosin terhadap Bakteri Indikator pada Tahap Uji Kepekaan Enzim Katalase... 59 23. Zona Hambat Substrat Kasar Bakteriosin terhadap S. aureus ATCC 25923 (a dan b) S. typhimurium ATCC 14028 (c) pada Tahap Uji Kepekaan Enzim Proteolitik... 60
PENDAHULUAN
Latar Belakang
Produk pangan hasil ternak khususnya daging merupakan bahan pangan sumber protein yang cukup digemari oleh masyarakat. Namun, produk pangan ini merupakan pangan yang mudah rusak sehingga memerlukan perlakuan khusus untuk memperpanjang daya simpannya. Alternatif dalam mengatasi masalah pembusukan tersebut adalah dengan pengawetan atau preservasi. Salah satu metode preservasi yaitu penambahan bahan pengawet.
Bahan pengawet dapat terbuat dari bahan kimia atau alami. Bahan pengawet yang bersifat kimia dan antibiotik memiliki beberapa kekurangan yaitu: (1) bahaya mikroorganisme yang resisten terhadap antibiotik dan dapat menimbulkan infeksi pada konsumen, (2) konsumen menjadi sensitif atau resisten terhadap antibiotik tertentu sehingga dapat mengganggu pemakaian untuk tujuan kesehatan, (3) kemungkinan toksisitas karena residu yang masih aktif dalam daging (Soeparno, 1994), (4) beberapa pengawet kimia diserap oleh bahan organik (darah, feses, dan jaringan) sehingga mengurangi efektivitas antimikroba (McKane dan Kandel, 1985).
Bahan pengawet yang bersifat alami yaitu bahan pengawet yang berasal dari bahan-bahan alami, salah satunya adalah penggunaan senyawa antimikroba yang berasal dari bakteri asam laktat seperti diasetil, hidrogen peroksida, asam-asam organik, dan bakteriosin (Schved et al., 1993). Penggunaan senyawa antimikroba asal bakteri asam laktat tidak menghasilkan efek samping yang merugikan karena tidak mengandung toksin yang membahayakan saluran pencernaan serta dapat didegradasi oleh enzim-enzim pencernaan. Salah satu senyawa antimikroba yang dikembangkan sebagai biopreservatif adalah bakteriosin.
Bakteriosin merupakan suatu senyawa protein yang memiliki sifat bakterisidal dan bakteristatik terhadap bakteri Gram positif atau bakteri yang dekat kekerabatannya dengan bakteri penghasil (Vuyst and Vandamme, 1994). Bakteriosin yang dihasilkan oleh bakteri asam laktat sangat menguntungkan bagi industri pangan karena aktivitasnya mampu menghambat pertumbuhan bakteri patogen maupun bakteri perusak yang biasanya terdapat dalam makanan (Gonzales et al., 1996). Penggunaan Lactobacillus fermentum 2B2 ini didasari oleh hasil penelitian Widiasih (2008) yang menyatakan bahwa Lactobacillus fermentum 2B2 merupakan salah satu
isolat bakteri asam laktat asal daging yang mempunyai aktivitas penghambatan (didominasi asam organik) yang paling baik terhadap ketiga bakteri uji (Staphylococcus aureus ATCC 25928, Salmonella typhimurium ATCC 14028, Escherichia coli ATCC 25922) yang dibuktikan dengan pembentukan rataan diameter zona hambat terbesar.
Tujuan
Penelitian ini bertujuan untuk mencari media terbaik dalam mengoptimalkan produksi bakteriosin, purifikasi parsial bakteriosin, mencari konsentrasi penghambatan minimum bakteriosin terhadap bakteri patogen (MIC dan MBC), dan karakterisasi bakteriosin kasar melalui uji kepekaan terhadap enzim katalase dan proteolitik.
TINJAUAN PUSTAKA
Mikrobiologi Daging
Buckle et al. (1987) menyatakan bahwa mutu mikrobiologi suatu produk makanan ditentukan oleh jumlah dan jenis mikroorganisme yang terdapat dalam bahan pangan. Hal ini akan menentukan ketahanan simpan dari produk tersebut ditinjau dari kerusakan oleh mikroorganisme. Keamanan produk ditentukan oleh jumlah miroorganisme patogenik yang terdapat di dalamnya. Populasi mikroorganisme yang berada pada suatu bahan pangan umumnya bersifat sangat spesifik dan tergantung pada jenis bahan pangan dan kondisi tertentu dari penyimpanannya.
Daging sangat memenuhi persyaratan untuk perkembagan mikroorganisme, termasuk mikroorganisme perusak atau pembusuk. Hal ini disebabkan daging mempunyai kadar air tinggi antara 68%-75%, kaya akan zat yang mengandung nitrogen dengan kompleksitas yang berbeda, mengandung sejumlah karbohidrat yang dapat difermentasikan, kaya akan mineral dan kelengkapan faktor untuk pertumbuhan mikroorganisme, mempunyai pH yang menguntungkan bagi sejumlah mikroorganisme sekitar 5,3–6,5 (Soeparno, 1994).
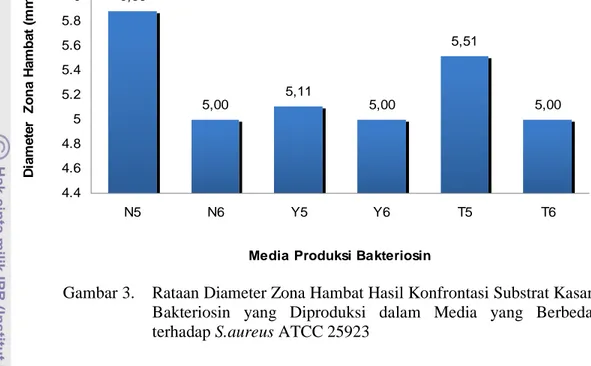
Kebanyakan bakteri tumbuh dipermukaan daging, namun tidak tertutup kemungkinan ditemukan bakteri dalam daging. Bakteri yang dapat mencapai jaringan dalam karkas dengan berbagai cara, diantaranya melalui mekanisme berikut: (1) jaringan ternak sehat dapat mengandung sebuah populasi kecil bakteri namun dinamis bila bakteri secara terus-menerus memperoleh akses ke dalam jaringan ternak hidup dengan penetrasi membran mukosa saluran respirasi dan pencernaan untuk mengganti yang telah dibasmi oleh mekanisme ketahanan tubuh ternak, (2) bakteri dari usus dapat menyerang jaringan karkas, baik selama pemotongan (agonal invasion) maupun setelah pemotongan (post mortem invasion), (3) bakteri dapat terbawa ke jaringan oleh luka sebelum pemotongan, (4) bakteri yang mengkontaminasi permukaan karkas dapat mempenetrasi ke lapisan jaringan otot yang lebih dalam. Tipe bakteri yang umum dijumpai pada daging adalah strain dari Pseudomonas, Moraxella, Acinetobacter, Lactobacillus, Brochtrix thermophacta dan beberapa genera dari famili Enterobacteriaceae (Gill, 1982). SNI mensyaratkan batas maksimum cemaran mikroba seperti tercantum dalam Tabel 1.
Tabel 1. Batas Maksimum Cemaran Mikroba pada Daging (cfu/g) SNI No. 01-6366-2000
No Jenis Cemaran Mikroba Batas Maksimum Daging Segar Beku
Cemaran Mikroba Daging Tanpa Tulang 1. 2. 3. 4. 5. 6. 7. 8. 9.
Angka Lempeng Total Bakteri (ALTB) Escherichia coli Staphylococcus aureus Clostridium sp. Salmonella sp. Coliform Enterococci Campylobacter sp. Listeria sp. 1 x 104 5 x 101 1x 101 0 Negatif 1 x 102 1 x 102 0 0 1 x 104 1 x 101 1x 102 0 Negatif 1 x 102 1 x 102 0 0
Keterangan : (*) dalam satuan MPN/gram (**) dalam satuan kualitatif
Bakteri Asam Laktat
Bakteri asam laktat merupakan bakteri Gram positif, non spora, berbentuk kokus atau batang, memiliki komposisi dasar DNA kurang dari 50 mol% G + C. Bakteri asam laktat pada umumnya mengandung katalase, dan memfermentasi karbohidrat untuk hidupnya. Glukosa dikonversi menjadi asam laktat (bakteri homofermentatif) atau menjadi asam laktat, karbon dioksida, etanol, dan asam asetat (bakteri heterofermentatif) (Vuyst dan Vandamme, 1994).
Genus bakteri asam laktat yaitu Aerococcus, Alloiococcus, Carnobacterium, Enterococcus, Lactobacillus, Lactococcus, Leuconostoc, Pediococcus, Streptococcus, Tetragenococcus, dan Vagococcus. Bakteri asam laktat dengan kelompok katalase negatif dan berbentuk kokus dapat dibagi menjadi dua grup yaitu bakteri fakultatif anaerob atau bakteri mikroaerofilik dan bakteri anaerob sempurna. Streptococcus merupakan genus terbesar pada grup pertama (Vuyst dan Vandamme, 1994).
Vuyst dan Vandamme (1994) menyatakan bahwa bakteri asam laktat pada umumnya didapatkan pada makanan termasuk pada daging fermentasi, sayuran, buah-buahan, minuman, dan produk susu. Bakteri asam laktat memiliki peranan yang
sangat penting pada makanan dan teknologi makanan, yang mana peranan utamanya dalam menghambat pertumbuhan bakteri perusak makanan. Bakteri asam laktat menghasilkan atau memproduksi senyawa penghambat pertumbuhan dan juga asam laktat yang memiliki dampak positif bagi kesehatan manusia dan hewan.
Lactobacillus
Lactobacillus merupakan genus terbesar dari bakteri asam laktat yaitu hampir 80 spesies. Genus ini tidak berspora, berbentuk batang dan beberapa cocobasil, katalase negatif, umumnya anaerob fakultatif atau mikroaerofilik, menghasilkan asam laktat, dan membutuhkan nutrisi yang kompleks. Bakteri asam laktat ini tumbuh optimum pada kondisi sedikit asam, yaitu pada pH diantara 4,5 sampai 6,4. Bakteri asam laktat umumnya tidak bersifat patogen (Prescott, 2003).
Lactobacillus ini dapat dibagi lagi menjadi tiga grup, yaitu (1) Grup I. Lactobacillus homofermentatif obligat; (2) Grup II. Lactobacillus heterofermentatif fakultatif; (3) Group III. Lactobacillus heterofermentatif obligat pembentuk gas. Lactobacillus pada grup III ini memfermentasi heksosa menjadi asam laktat, asam asetat, etanol, dan karbon dioksida. Pentosa difermentasi menjadi asam laktat dan asetat. Berdasarkan asam nukleat data biokimia, strain yang jelas ditemukan pada Lactobacillus brevis dan Lactobacillus fermentum (Vuyst dan Vandamme, 1994). Lactobacillus fermentum merupakan bakteri yang dapat memfermentasi beberapa gula-gula sederhana yaitu galaktosa, glukosa, laktosa, rafinosa, sukrosa, dan xylosa (Arief, 2005).
Antimikroba
Bakteri asam laktat dapat memproduksi senyawa antimikroba selain asam laktat dan asam asetat. Senyawa ini bersifat antagonistik terhadap mikroorganime dalam spektrum yang luas, sehingga dapat berkontribusi sebagai pengawet. Senyawa ini dihasilkan dalam jumlah yang lebih sedikit dibanding asam laktat atau asam asetat yaitu asam format, asam lemak bebas, amonia, etanol, hidrogen peroksida, diasetil, asetoin, 2,3 butanediol, asetaldehid, benzoat, enzim bakteriolitik, antibiotik, dan bakteriosin. Beberapa dari senyawa tersebut menunjukkan aktivitas antagonistik terhadap banyak jenis mikroorganisme patogen dan perusak makanan (Vuyst dan Vandamme, 1994).
Asam Organik
Asam organik (asetat, laktat, malat, sitrat) adalah unsur pokok alami dalam makanan. Asam organik dipergunakan secara luas sebagai bahan tambahan dalam pengawetan makanan. Aktivitas mikrobial asam organik terutama didasarkan pada kemampuannya dalam menurunkan pH makanan dalam bentuk cair atau larutan. Saat nilai pH lebih rendah dari 4,0, asam akan menghambat pertumbuhan bakteri, sedangkan khamir dan kapang berkompetisi melawan bakteri pada nilai pH dibawah 5,0 (Samelis dan Sofos, 2001c).
Asam organik lipofilik lemah (asetat, propionat, sorbat, dan benzoat) melakukan penetrasi terhadap membran sel dalam bentuk nondisosiasi untuk menghambat pertumbuhan atau menyebabkan kematian dengan cara menguraikan atau mengasamkan sitoplasma. Mekanisme aktivitas antimikroba didasarkan pada penghambatan enzim, fungsi membran, transpor nutrien, dan aktivitas metabolisme keseluruhan (Samelis dan Sofos, 2001c).
Hidrogen Peroksida
Vuyst dan Vandamme (1994) mengungkapkan bahwa produksi hidrogen peroksida membuat bakteri asam laktat bersifat antagonistik terhadap bakteri lain. Bakteri asam laktat sanggup membentuk hidrogen peroksida dan mengakumulasikannya pada media pertumbuhan asalkan bakteri ini katalase negatif. Akumulasi ini akan meningkatkan tingkat autoinhibitor.
Mekanisme penghambatan oleh hidrogen peroksida yaitu dengan meningkatkan permeabilitas membran dengan O2- dan OH. Hasilnya efek bakterisidal dari metabolit oksigen tidak hanya terjadi sebatas sel bakteri tetapi juga terjadi sampai pada struktur molekul dasar dari asam nukleat dan sel protein (Vuyst dan Vandamme, 1994).
Antagonistik hidrogen peroksida tidak hanya berasal dari diri sendiri jika pada lingkungan alami (susu, daging, dan saliva atau air liur), tetapi melalui reaksi sekunder produk. Misalnya, Lactobacillus asal daging menghasilkan hidrogen peroksida yang bereaksi dengan senyawa protein dalam daging yang membentuk senyawa penghambat (Vuyst dan Vandamme, 1994).
Bakteriosin
Bakteriosin menurut (Klaenhammer, 1990) adalah protein atau peptida yang disintesa melalui ribosom yang dapat menghambat atau membunuh bakteri lain. Tagg et al., (1976) menyatakan bahwa semua anggota dari Eubacteria dan Archaea yang diambil dari ekosistem alamiahnya pasti menghasilkan bakteriosin. Namun, jika tidak ditemukan bakteriosin disebabkan karena penelitinya yang belum menemukan kondisi yang tepat yang menunjukkan bacteriosinogenicity in vitro.
Kondisi bakteri di laboratorium ditumbuhkan pada media monokultur (tidak ada persaingan) dan kondisi rendah stres karena kelebihan nutrisi sehingga akan mempermudah pelepasan bakteriosin. Namun, kultur pada laboratorium sering kali ditinggikan suhu pertumbuhannya yang justru akan mengeliminasi plasmid bakteriosin (Riley dan Chavan, 2007).
Metode dengan menggunakan media agar merupakan metode yang paling umum untuk mendeteksi adanya bakteriosin secara in vitro, tetapi beberapa bakteriosin harus didalam media cair. Kondisi optimum pada bakteri indikator tidak perlu tepat dengan kondisi produksi bakteriosin. Namun tentu saja, produksi bakteriosin dapat ditinggikan ketika bakteri dalam keadaan stress (nutrisi dan lingkungan). Adapun media suplemen khusus yang berpengaruh nyata dalam memproduksi bakteriosin yaitu penambahan yeast extract (YE) (meningkatkan produksi mutacin), glukosa (mempengaruhi penahanan katabolit dari bakteriosin streptococal), dan ion magnesium (menahan ekspresi lantibiotik) (Riley dan Chavan, 2007).
Produksi bakteriosin menurut Matsuaki et al. (1996) dipengaruhi oleh level sumber karbon, nitrogen, dan phosfat yang bisa didapat melalui media. Sumber karbohidrat yang berbeda menghasilkan bakteriosin yang berbeda. Nisin sebagai contoh dapat diproduksi dari glukosa, sukrosa, dan xylosa oleh Lactococcus lactis 10-1 (Matsuaki et al., 1996). Biswas et al. (1991) menyatakan bahwa glukosa, sukrosa, xylosa, dan galaktosa adalah sumber karbon terbaik dalam menghasilkan Pediocin AcH dalam media tanpa buffer. Holo et al., (1991) mengungkapkan bahwa semua bakteriosin disintesa dengan sekuen terminal N yang fungsinya mencegah bakteriosin bersifat aktif saat masih ada didalam bakteri penghasilnya.
Bakteriosin dihasilkan sebagai pre-propeptida yang diperoses dan dieksternalisasi atau dikeluarkan melalui perlengkapan transpor (Nes et al., 1996). Parente et al. (1997) menyatakan bahwa produksi bakteriosin berbanding lurus dengan pertumbuhan bakteri. Produksi bakteriosin pada umumnya terjadi pada fase pertumbuhan dan berhenti pada akhir fase eksponensial atau terkadang sebelum berakhir fase log. Jika ingin menghasilkan bakteriosin yang optimal, maka perlu disesuaikan waktu inkubasi yang sesuai pada fase pertumbuhan dari bakteri L. fermentum 2B2.
Klasifikasi bakteriosin didasarkan pada ada atau tidak adanya asam amino yang tidak umum, lanthionin, dan β-lanthionin (Jack et al., 1995). Menurut (Klaenhammer, 1990) bakteriosin asal bakteri asam laktat dibagi menjadi empat kelas, yaitu (1) Kelas I. Bakteriosin yang telah dikenal atau lantibiotik; (2) Kelas II. Kecil (< 10 kDa), membran aktif bakteriosin stabil kondisi panas; (3) Kelas III. Lebih besar (> 30 kDa) bakteriosin labil terhadap panas; (4) Kelas IV. Bakteriosin kompleks yang tergabung dengan lipid atau karbohidrat.
Mekanisme bakteriosin cenderung melawan bakteri Gram positif dengan sedikit pengaruh dari molekul reseptor atau pelepasan bakteriosin dari sel penghasilnya ditingkatkan oleh pengaruh lisin atau bakteriosin yang dilepas berupa protein. Ketiadaan membran terluar pada bakteri Gram positif meniadakan kemungkinan pengaruh molekul reseptor yang terjadi pada bakteri Gram negatif dengan bakteri yang sensitif. Potensi letal bakteriosin asal bakteri Gram positif terhadap sel yang sensitif tergantung pada kecocokan atau kesesuaian antara permukaan serangan dan interaksi molekul hidrofobik. Perbedaan lain yaitu tingkat imunitas bakteriosin yang dihasilkan dari bakteri Gram positif kurang kuat dibanding colicin (Tagg et al., 1976).
Bakteri Patogen
Banyaknya jenis organisme yang dapat menginitasi dan menyebabkan penyakit karena kerusakan makanan salah satunya kelompok bakteri patogen perusak makanan. Bakteri patogen penyebab kerusakan makanan dibagi menjadi dua grup yaitu bakteri Gram positif dan bakteri Gram negatif. Bakteri yang termasuk pada grup pertama yaitu Staphylococcus, Bacillus aureus, B. anthracis, Clostridium botulinum, C. perfringens, dan Listeria monocytogenes. Bakteri yang masuk dalam
grup ke dua yaitu Salmonella, Shigella, Escherichia, Yersinia, Vibrio, Campylobacter, dan Brucella (Jay, 2000).
Staphylococcus aureus
Bakteri S. aureus merupakan bakteri Gram positif yang dikenal sebagai bakteri jahat yang menghasilkan eksotoksik yang tidak ada pada bakteri baik. Gastroenteritis staphylococcal disebabkan terinfeksi oleh satu atau lebih enterotoksin yang dihasilkan beberapa spesies Staphylococcus. Spesies dari Staphylococcus ini banyak didapatkan dalam jumlah besar pada permukaan tubuh yang terbuka. Bakteri ini juga banyak ditemukan pada makanan komersial (Jay, 2000).
Jay (2000) menyatakan bahwa Staphylococcus membutuhkan asam amino sebagai sumber nitrogennya, dan thiamin serta asam nikotinat yang adalah vitamin B. Bakteri ini tumbuh dalam suasana anaerob maka dibutuhkan urasil, sedangkan untuk tumbuh secara aerob dan menghasilkan enterotoksin dibutuhkan monosodium glutamat sebagai sumber C, N, dan energi. S. aureus memiliki toleransi yang tinggi pada senyawa seperti tellurit, merkuri klorida, neomycin, polymyxin, dan sodium azide, serta semua agen selektif dalam kultur media.
Temperatur tumbuh bakteri S. aureus pada umumnya berkisar 7–47,8oC, dan untuk menghasilkan enterotoksin berkisar 10oC–46oC, sedangkan suhu optimumnya pada 40oC dan 45oC. S. aureus tumbuh baik pada media tanpa NaCl, namun tetap dapat tumbuh baik pada konsentrasi NaCl 7-10%, bahkan beberapa dapat tumbuh pada konsentrasi 20%. S. aureus tumbuh pada kondisi pH 4,0-9,8, sedangkan suhu optimumnya pada pH 6,0-7,0 (Jay, 2000).
Escherichia coli
Bakteri Escherichia coli telah dikenal sebagai penyebab penyakit pada manusia sejak ditemukan pada tahun 1985. Penyakit yang disebabkan oleh bakteri ini yaitu diare, disentri, pneumonia, meningitis, infeksi ginjal dan kandung kemih. E. coli telah menjadi indikator pencemaran kotoran pada air dan susu bahkan telah menjadi indikator higienis dalam berbagai jenis makanan (Bell dan Kyriakides, 1998).
Enteropatogenik E. coli tidak menghasilkan enterotoksin, walaupun demikian tetap dapat menyebabkan diare tetapi yang hanya menyerang anak-anak dibawah umur 1 tahun. ETEC sekali menyerang mengeluarkan satu sampai dua enterotoksin.
Enterotoksigenik E. coli menyerang dan hidup berkoloni pada usus halus melalui faktor antigen fimbrial kolonisasi (CFAs). Diare yang disebabkan oleh ETEC tidak hanya menyerang anak-anak tetapi juga orang dewasa. Diare yang disebabkan oleh ETEC ini biasanya menyebar dan terjadi tiba-tiba yang disertai demam. Jumlah bekteri ETEC yang diperlukan untuk mengakibatkan diare pada orang dewasa berkisar 108-1010 (Jay, 2000).
Salmonella typhimurium
Salmonella merupakan bakteri Gram negatif, non-spora, berbentuk batang yang tak dapat dibedakan dengan E. coli dibawah mikroskop atau pada media biasa. Bakteri ini tersebar luas pada manusia dan hewan. Berdasarkan tujuan epidemilogi, Salmonella dibagi menjadi tiga grup yaitu (1) Salmonella yang menyerang manusia yaitu S. Typhi, S. Paratyphi A, S. Paratyphi C. Grup ini termasuk demam tipoid dan paratipoid, yang menjadi penyebab hampir semua penyakit yang disebabkan oleh Salmonella; (2) Serovar yang telah adaptasi pada inang (beberapa patogen pada manusia yang berasal dari makanan) yaitu S. Galinarum (unggas), S. Dublin (sapi), S. Abortus-equi (kuda), S. Abortus-ovis (domba), dan S. Choleraesuis (angsa); (3) Nonadaptasi serovar yaitu patogenik pada manusia dan hewan lain dan serovar perusak makanan (Jay, 2000).
Jay (2000) menyatakan bahwa habitat utama dari Salmonella ada pada saluran pencernaan dari hewan seperti burung, reptil, hewan ternak, manusia. dan terkadang serangga. Bakteri ini dapat tumbuh dalam jumlah besar pada media dan menghasilkan koloni yang tampak jelas dalam 24 jam dengan suhu 37oC. S. typhimurium membutuhkan nitrat, nitrit, dan NH3 sebagai sumber nitrogen. Kondisi
pH optimum untuk tumbuhnya adalah pH netral, dengan nilai diatas 9,0 dan dibawah pH akan mati, sedangkan untuk pertumbuhan terbaiknya dibutuhkan pH antara 6,6 dan 8,2. Temperatur terendah untuk S. typhimurium terjadi pada 6,2oC.
Enzim
Enzim adalah katalisator organik (biokatalisator) yang dihasilkan oleh sel. Enzim berfungsi seperti katalisator anorganik, yaitu untuk mempercepat reaksi kimia. Setelah reaksi berlangsung, enzim tidak mengalami perubahan jumlah, sehingga jumlah enzim sebelum dan setelah reaksi adalah tetap. Enzim mempunyai
selektivitas dan spesifitas yang tinggi terhadap reaktan yang direaksikan dan jenis reaksi yang dikatalisasi (Sumarsih, 2007).
Saat berlangsungnya reaksi enzimatik terjadi ikatan sementara antara enzim dengan substratnya (reaktan). Ikatan sementara ini bersifat labil dan hanya untuk waktu yang singkat saja. Selanjutnya ikatan enzim-substrat akan pecah menjadi enzim dan hasil akhir. Enzim yang terlepas kembali setelah reaksi dapat berfungsi lagi sebagai biokatalisator untuk reaksi yang sama. Pada umumnya enzim tersusun dari protein. Protein penyusun enzim dapat berupa protein sederhana atau protein yang terikat pada gugusan non-protein. Banyak enzim yang hanya terdiri protein saja, misal tripsin (Sumarsih, 2007).
Sumarsih (2007) menyatakan bahwa protein adalah bagian utama enzim yang dihasilkan sel, maka semua hal yang dapat mempengaruhi protein dan sel akan berpengaruh terhadap reaksi enzimatik. Faktor-faktor yang mempengaruhi reaksi enzimatik yaitu:
1. Substrat (reaktan)
Kecepatan reaksi enzimatik umumnya dipengaruhi kadar substrat. Penambahann kadar substrat sampai jumlah tertentu dengan jumlah enzim yang tetap, akan mempercepat reaksi enzimatik sampai mencapai maksimum. Penambahan substrat selanjutnya tidak akan menambah kecepatan reaksi. Kecepatan reaksi enzimatik juga dipengaruhi kadar enzim.
2. Suhu
Sama seperti reaksi kimia pada umumnya, maka reaksi enzimatik dipengaruhi oleh suhu. Kenaikan suhu sampai optimum akan diikuti pula oleh kenaikan kecepatan reaksi enzimatik. Kepekaan enzim terhadap keadaan suhu melebihi optimum menyebabkan terjadinya perubahan fisikokimia protein penyusun enzim. Umumnya enzim mengalami kerusakan (denaturasi) pada suhu diatas 50oC. Walaupun demikian ada beberapa enzim yang tahan terhadap suhu tinggi, misalnya taka-diastase dan tripsin.
3. Kemasaman (pH)
Kondisi pH dapat mempengaruhi aktivitas enzim. Daya katalisis enzim menjadi rendah pada pH rendah maupun tinggi, karena terjadinya denaturasi protein enzim. Enzim mempunyai gugus aktif yang bermuatan positif (+) dan negatif (-).
Aktivitas enzim akan optimum kalau terdapat keseimbangan antara kedua muatannya. Pada keadaan masam muatannya cenderung positif, dan pada keadaan basa muatannya cenderung negatif sehingga aktivitas enzimnya menjadi berkurang atau bahkan menjadi tidak aktif. Kondisi pH optimum untuk masing-masing enzim tidak selalu sama. Sebagai contoh amylase jamur mempunyai pH optimum 5,0, arginase mempunyai pH optimum 10.
4. Penghambat enzim (inhibitor)
Inhibitor enzim adalah zat atau senyawa yang dapat menghambat enzim dengan beberapa cara penghambatan sebagai berikut: (a) Penghambat bersaing (kompetitif) yaitu daya penghambatannya dipengaruhi oleh kadar penghambat, kadar substrat dan aktivitas relatif antara penghambat dan substrat; (b) Penghambat tidak bersaing (non-kompetitif) yaitu daya penghambatannya dipengaruhi oleh kadar penghambat dan afinitas penghambat terhadap enzim.
Enzim Katalase
Moss (1987) menyatakan bahwa katalase ada dimana-mana secara alami. Katalase didapatkan pada semua mikroorganisme aerob baik dalam sel tumbuhan maupun sel hewan. Katalase adalah enzim tetramerik haemin yang terdiri dari empat identik tertra hedral disusun oleh setiap subunit 60000 g/mol. Oleh sebab itu, enzim ini terdiri dari empat ferriproliporphyrin grup/molekul sekitar 240000.
a. Dekomposisi H2O2 menjadi H2O dan O2 2H2O2 H2O + O2
b. Oksigen donor H menjadi methanol, athanol, asam format,phenol, dengan konsumsi 1 mol peroksida
ROOH + AH2 H2O + ROH
Pencegahan enzim katalase menjadi inaktif selama percobaan (umumnya 30 detik), diperlukan konsentrasi H2O2 yang relatif rendah (10 mmol/l). Konsentrasi
H2O2 merupakan hal yang kritis sehingga harus proporsional antara konsentrasi
substrat dengan rata-rata dekomposisi. Pengaruh suhu dalam dekomposisi H2O2 kecil
sehingga dapat dilakukan pada kisaran 0oC–37oC, tetapi suhu 20oC (temperatur ruang) yang paling direkomendasikan. Pada suhu ini dapat mempercepat reaksi katalase walaupun durasi inkubasi pendek (≤ 30 detik ). Aktivitas antara pH 6,8–7,5, tetapi pH optimumnya pada pH 7,0 (Moss, 1987).
Enzim Proteolitik
Proteinase memiliki siklus hidup yang kompleks. Enzim ini disekresikan dan ditransportasikan didalam sel sebagai pre-proenzim. Enzim ini disimpan dalam bentuk aktif, bentuk laten atau bentuk inaktif proenzim dalam bagian khusus sel sebelum digunakan dalam reaksi biologi atau dilepaskan untuk fungsi fisiologi (Geiger dan Fritz, 1982).
Tripsin pertama kali didapatkan dalam bentuk kristal oleh Northrop dan Kunitz tahun 1934. Tripsin merupakan endopeptidase yang terutama memecah protein dan peptida pada ikatan carboxamid dari residu lisin dan arginin. Enzim ini disintesa dalam bagian eksokrin pada pankreas sebagai prekursor atau zymogen (tripsinogen) dan disimpan dalam granul zymogen. Enzim ini disekresikan kedalam saluran pankreas yang kemudian masuk ke dalam usus setelah stimulus yang sesuai (Geiger dan Fritz, 1982).
Pepsin labil pada nilai pH diatas 6,0 Pepsin dapat mengkatalisasi transpeptidasi dari kedua amino-transfer dan karboksil-transfer pada substrat yang sesuai. Reaksi ini tidak hanya penting untuk mekanisme enzim, tetapi turut juga pada percobaan yang mengunakan substrat peptida dengan amino atau karboksil yang tidak terlindung atau percobaan dengan menggunakan beberapa substrat peptida jika peptida lain juga ada (Ryle, 1970).
Minimum Inhibitory Concentration (MIC)
Penghambatan mikroba oleh suatu senyawa antimikroba dapat dinyatakan dalam nilai MIC. Cosentino (1999) mendefinisikan MBC sebagai konsentrasi terendah yang dapat menghambat pertumbuhan mikroba sebanyak 90% dari inokulum asal selama inkubasi 24 jam. Nilai MIC senyawa antimikroba yang lebih rendah menunjukkan bakteri lebih sensitif terhadap senyawa tersebut (Naufalin, 2005). Fase pertumbuhan berpengaruh terhadap sensitifitas bakteri terhadap senyawa antimikroba. Bakteri pada fase stasioner lebih sensitif terhadap antimikroba asam lemak rantai pendek dari bakteri fase pertumbuhan. Hal ini disebabkan karena penambahan asam rantai pendek seperti asam propionat pada fase pertumbuhan (Thompson dan Hinton, 1996).
METODE
Lokasi dan Waktu
Penelitian ini dilaksanakan dari bulan September 2008 sampai bulan September 2009. Penelitian dilakukan di Laboratorium Mikrobiologi Bagian Teknologi Hasil Ternak Perah dan Laboratorium Ruminansia Besar, Fakultas Peternakan, Institut Pertanian Bogor.
Materi
Bahan-bahan utama yang digunakan adalah isolat bakteri asam laktat asal daging yaitu Lactobacillus fermentum 2B2 dan bakteri indikator (patogen) (Staphylococcus aureus ATCC 25923, Eschericia coli ATCC 25922 , Salmonella typhimuriumATCC 14028, dan enteropatogenik Escherichia coli lokal) untuk tahap produksi bakteriosin. Tahap purifikasi parsial bakteriosin, serta tahap kepekaan enzim katalase dan proteolitik tanpa menggunakan Escherichia coli ATCC 25922. Pada tahap MIC dan MBC hanya menggunakan bakteri uji S. aureus. Media yang digunakan yaitu de Man Ragosa Sharp Broth (MRSB), Nutrient Agar (NA), Nutrient Broth (NB), Buffer Pepton Water (BPW), Muller Hilton Agar (MHA), Yeast Extract (YE), NaCl, tripton, NaOH 1 N, amonium sulfat, enzim katalase, tripsin, pepsin, KH2PO4 0,05 M Tris Hydrochloride (pH 8,0), 0,2 M sitrat (pH 6,0), larutan Mc.
Farland No. 0,5, es batu, dan aquadest.
Peralatan yang digunakan dalam penelitian ini antara lain : Ose, cawan Petri, tabung reaksi, sentrifuse, kertas saring, alumunium foil, karton coklat, plastik wrap, timbangan, autoclaf, alat titrasi, gelas ukur, pipet, pipet pasteur, pipet mikro, tip, tabung ependorf, millipore 0,22 µm, tabung Erlenmeyer, tabung Scott, spuit volume 10 ml, dan jangka sorong.
Rancangan
Rancangan penelitian yang digunakan adalah rancangan acak lengkap (RAL) dengan 3 ulangan untuk tahap produksi bakteriosin, karakterisasi bakteriosin, dan konsentrasi minimum inhibitor. Perlakuannya yaitu penggunaan media yang berbeda pada tahap produksi dan karakterisasi bakteriosin, perbedaan konsentrasi substrat kasar bakteriosin pada tahap MIC dan MBC. Model matematikanya sebagai berikut:
Keterangan :
Yij : Nilai respon perlakuan media yang berbeda pada substrat kasar bakteriosin
terhadap bakteri indikator µ : Nilai tengah populasi
Bi : Pengaruh media yang berbeda (tahap produksi dan purifikasi parsial
bakteriosin) dan perbedaan konsentrasi substrat kasar bakteriosin (tahap MIC dan MBC)
εij : Pengaruh galat percobaan
Rancangan percobaan lainnya yang digunakan adalah metode statistik deskritif untuk optimasi produksi bakteriosin, purifikasi parsial bakteriosin, konsentrasi minimum inhibitor, uji enzim katalase dan proteolitik.
Prosedur
Produksi Bakteriosin pada Media yang Berbeda
Penyegaran Bakteri Asam Laktat Lactobacillus fermentum 2B2. Kultur L. fermentum 2B2 dari media MRSB dipipet sebanyak 1 ml dan dimasukkan ke dalam media MRSB yang ditambah YE 3% (9 ml), kemudian diinkubasi selama 24 jam pada suhu 37oC.
Produksi Bakteriosin dari Lactobacillus fermentum 2B2. Kultur L. fermentum 2B2 hasil penyegaran berumur 24 jam dipipet sebanyak 2 ml dan dimasukkan ke dalam 3 media (masing-masing media sebanyak 18 ml) yaitu (1) MRSB dan NaCl 1% ; (2) MRSB, YE 3 %, dan NaCl 1%; serta (3) MRSB dan tripton 1%, masing-masing disiapkan untuk pH 5,0 dan pH 6,0 (ada 6 tabung). Kultur yang dihasilkan untuk setiap media sebanyak 20 ml. Kultur L. fermentum kemudian diinkubasi selama 20 jam pada suhu 370C. Kultur L. fermentum 2B2 umur 20 jam tersebut dimasukkan ke dalam ependorf sesuai dengan medianya masing-masing, kemudian disentrifuse selama 20 menit dengan kecepatan 10.000 rpm pada suhu 40C. Supernatan bebas sel yang dihasilkan dari setiap media dipisahkan dengan endapannya. Supernatan bebas sel dimasukkan dalam tabung untuk diukur pH awalnya, sedangkan sebagian kecil substrat disisihkan untuk diukur total asam tertitrasi (TAT). Supernatan pada setiap media dikondisikan menjadi pH 5,0 dan pH
6,0 dengan penambahan NaOH 1 N. Supernatan yang telah dikondisikan pHnya diukur kembali TATnya. Supernatan bebas sel pada pH 5,0 dan pH 6,0 disterilkan dengan cara disaring menggunakan filter 0,22 µm (millipore). Supernatan bebas sel tersebut digunakan dalam uji antagonistik (konfrontasi) dengan bakteri indikator. Produksi Bakteri Indikator sebagai Kultur Kerja. Masing-masing bakteri indikator (Staphylococcus aureus ATCC 25923, Eschericia coli ATCC 25922, Salmonella typhimuriumATCC 14028, dan enteropatogenik Escherichia coli lokal) diinkubasi selama 24 jam pada suhu 37oC. Bakteri indikator umur 24 jam distandarisasi jumlahnya dengan cara disetarakan kekeruhannya sesuai standar Mc Farland no. 0,5 didalam media NaCl sehingga dihasilkan populasi bakteri setara 1,5 x 108 sel bakteri/ml. Konfrontasi bakteri indikator dengan substrat bakteriosin pada tahap produksi bakteriosin menggunakan konsentrasi bakteri 1,5 x 106 sel bakteri/ml yang diperoleh dengan cara mengencerkannya 100 kali dalam buffer pepton water (BPW) steril.
Konfrontasi Substrat Kasar Bakteriosin dengan Bakteri Indikator. Supernatan bebas sel (substrat kasar bakteriosin) pada setiap media dengan masing-masing pH 5,0 dan pH 6,0 dikonfrontasi dengan keempat bakteri indikator dengan metode difusi agar. Bakteri indikator dimasukkan ke dalam cawan sebanyak 1 ml kemudian dituang media MHA sebanyak 25 ml. Cawan digerakkan membentuk angka delapan untuk menghomogenkan media dengan bakteri. Agar yang telah mengeras dilubangi dengan menggunakan pipet Pasteur (diameter 5 mm) dan agarnya dibuang dengan menggunakan Ose. Supernatan bebas sel dimasukkan ke dalam lubang tersebut sebanyak 50 µl. Cawan ditutup dengan kertas saring kemudian ditutup dengan penutup cawan. Cawan dimasukkan ke dalam lemari pendingin selama 2 jam kemudian diinkubasi selama 20 jam. Zona bening yang dihasilkan diukur diameternya.
Purifikasi Parsial Bakteriosin
Penyegaran Bakteri Asam Laktat Lactobacillus fermentum 2B2. Kultur L. fermentum 2B2 dari media MRSB dipipet sebanyak 1 ml dan dimasukkan ke dalam media MRSB yang ditambah YE 3% (9 ml), kemudian diinkubasi selama 24 jam pada suhu 37oC.
Produksi Bakteriosin dari Lactobacillus fermentum 2B2. Kultur L. fermentum 2B2 hasil penyegaran berumur 24 jam dipipet sebanyak 7 ml dan dimasukkan ke dalam 3 media (masing-masing media sebanyak 63 ml) yaitu (1) MRSB dan NaCl 1% ; (2) MRSB, YE 3 %, dan NaCl 1%; serta (3) MRSB dan tripton 1%. Kultur yang dihasilkan untuk setiap media sebanyak 70 ml. Kultur L. fermentum kemudian diinkubasi selama 20 jam pada suhu 370C. Kultur L. fermentum 2B2 umur 20 jam tersebut dimasukkan ke dalam ependorf kemudian disentrifuse selama 20 menit dengan kecepatan 10.000 rpm pada suhu 40C. Supernatan bebas sel yang dihasilkan dari setiap media dipisahkan dengan endapannya. Supernatan bebas sel dimasukkan ke dalam tabung untuk diukur pH awalnya. Supernatan pada setiap media dikondisikan menjadi pH 6,0 dengan penambahan NaOH 1 N. Supernatan bebas sel dengan pH 6,0 disterilkan dengan cara disaring menggunakan filter 0,22 µm (millipore) kemudian ditambahkan amonium sulfat sebanyak 40% pada setiap media. Gerakkan wadah substrat tersebut secara perlahan hingga seluruh amonium sulfat larut didalam substrat. Substrat didiamkan selama 24 jam dalam lemari pendingin. Substrat yang telah didiamkan semalaman tersebut akan menghasilkan protein yang mengapung diatas substrat. Protein dipisahkan dengan sebagian besar substrat dengan mengeluarkan substrat yang berada di bawah protein yang mengapung. Protein (substrat kasar bakteriosin) tersebut ditambahkan dengan buffer KH2PO4.
Substrat kasar bakteriosin tersebut digunakan dalam uji antagonistik (konfrontasi) dengan bakteri indikator.
Produksi Bakteri Indikator sebagai Kultur Kerja. Masing-masing bakteri indikator (Staphylococcus aureus ATCC 25923, Salmonella typhimurium ATCC 14028, dan enteropatogenik Escherichia coli lokal) diinkubasi selama 24 jam pada suhu 37oC. Bakteri indikator umur 24 jam distandarisasi jumlahnya dengan cara disetarakan kekeruhannya sesuai standar Mc Farland no. 0,5 dalam media NaCl sehingga dihasilkan populasi bakteri setara 1,5 x 108 sel bakteri/ml.
Konfrontasi Substrat Kasar Bakteriosin dengan Bakteri Indikator. Substrat kasar bakteriosin pada pH 6,0 dikonfrontasi dengan ketiga bakteri indikator dengan metode difusi agar. Bakteri indikator dimasukkan ke dalam cawan sebanyak 1 ml kemudian dituang media MHA sebanyak 25 ml. Cawan digerakkan membentuk angka delapan untuk menghomogenkan media dengan bakteri. Agar yang telah
mengeras dilubangi dengan menggunakan pipet Pasteur (diameter 5 mm) dan agarnya dibuang dengan menggunakan Ose. Substrat kasar bakteriosin dimasukkan ke dalam lubang tersebut sebanyak 50 µl. Cawan ditutup dengan kertas saring kemudian ditutup dengan penutup cawan. Cawan dimasukkan ke dalam lemari pendingin selama 2 jam kemudian diinkubasi selama 20 jam. Zona bening yang dihasilkan diukur diameternya.
Konsentrasi Penghambatan Minimum (MIC dan MBC)
Penyegaran Bakteri Asam Laktat Lactobacillus fermentum 2B2. Kultur L. fermentum 2B2 dari media MRSB dipipet sebanyak 1 ml dan dimasukkan ke dalam media MRSB yang ditambah YE 3% (9 ml), kemudian diinkubasi selama 24 jam pada suhu 37oC.
Produksi Bakteriosin dari Lactobacillus fermentum 2B2. Kultur L. fermentum 2B2 hasil penyegaran berumur 24 jam dipipet sebanyak 7 ml dan dimasukkan ke dalam media MRSB ditambah tripton 1% (63 ml). Kultur yang dihasilkan sebanyak 70 ml. Kultur L. fermentum kemudian diinkubasi selama 20 jam pada suhu 370C. Kultur L. fermentum 2B2 umur 20 jam tersebut dimasukkan ke dalam ependorf kemudian disentrifuse selama 20 menit dengan kecepatan 10.000 rpm pada suhu 40C. Supernatan bebas sel yang dihasilkan dari setiap media dipisahkan dengan endapannya dengan dimasukkan dalam tabung untuk diukur pH awal. Supernatan pada setiap media dikondisikan menjadi pH 6,0 dengan penambahan NaOH 1 N. Supernatan bebas sel dengan pH 6,0 disterilkan dengan cara disaring menggunakan filter 0,22 µm (millipore) kemudian ditambahkan amonium sulfat sebanyak 40% dalam setiap media. Gerakkan wadah substrat tersebut secara perlahan hingga seluruh amonium sulfat larut didalam substrat. Substrat didiamkan selama 24 jam dalam lemari pendingin. Substrat yang telah didiamkan semalaman tersebut akan menghasilkan protein yang mengapung diatas substrat. Protein dipisahkan dengan sebagian besar substrat dengan mengeluarkan substrat yang ada di bawahnya, kemudian ditambahkan dengan buffer KH2PO4. Substrat kasar bakteriosin tersebut
digunakan dalam uji antagonistik (konfrontasi) dengan bakteri indikator.
Produksi Bakteri Indikator sebagai Kultur Kerja. Bakteri indikator S. aureus ATCC 25923 diinkubasi selama 24 jam pada suhu 370C. Bakteri S. aureus ATCC
25923 umur 24 jam yang akan digunakan distandarisasi jumlahnya dengan cara disetarakan kekeruhannya sesuai standar Mc Farland no. 0,5 dalam media NaCl sehingga dihasilkan populasi bakteri setara 1,5 x 108 sel bakteri/ml.
Media NB, substrat kasar bakteriosin, dan bakteri indikator dimasukkan ke dalam tabung reaksi dengan perbandingan volume pada (Tabel 2.), kemudian divortex agar menyatu. Kultur tersebut kemudian dimasukkan ke dalam cawan ditambahkan dengan MHA kemudian diinkubasi selama 24 jam pada suhu 370C.
Tabel 2. Volume Media NB, Substrat Kasar Bakteriosin, dan Bakteri Indikator
Media NB Substrat Kasar Bakteriosin Bakteri Indikator
--- ml --- 4,5 0 0,5 4,0 0,5 0,5 3,5 1,0 0,5 3,0 1,5 0,5 2,5 2,0 0,5 2,0 2,5 0,5 1,5 3,0 0,5 1,0 3,5 0,5 0,5 4,0 0,5 0 4,5 0,5
Pertumbuhan bakteri indikator pada setiap cawan dihitung secara manual. Perhitungan jumlah koloni meggunakan metode Aerobic Plate Count pada Bacteriological Analytical Manual (BAM, 2001).
Rumus Hitung APC:
Formula: N = C x d [(1x N1) + (0,1x N2)]
Keterangan: N : Jumlah koloni per ml atau g produk
C : Jumlah semua koloni pada cawan yang dapat dihitung
N1 : Jumlah koloni pada cawan pengenceran pertama yang dapat dihitung N2 : Jumlah koloni pada cawan pengenceran kedua yang dapat dihitung d : Faktor pengencer dari perhitungan pertama dihasilkan
2. Cawan dengan jumlah koloni kurang dari 25
Hitung jumlah yang ada pada cawan dari setiap pengenceran. Rerata jumlah koloni per cawan dan kalikan dengan faktor pengencer dari jumlah perhitungan pertama dihasilkan.
2. Cawan dengan jumlah koloni lebih dari 250
Hitung koloni pada cawan kemudian dikali dengan faktor pengencer. Uji Kepekaan Substrat Kasar Bakteriosin terhadap Enzim Katalase
Penyegaran Bakteri Asam Laktat Lactobacillus fermentum 2B2. Kultur L. fermentum 2B2 dari media MRSB dipipet sebanyak 1 ml dan dimasukkan ke dalam media MRSB yang ditambah YE 3% (9 ml), kemudian diinkubasi selama 24 jam pada suhu 37oC.
Produksi Bakteriosin dari Lactobacillus fermentum 2B2. Kultur L. fermentum 2B2 hasil penyegaran berumur 24 jam dipipet sebanyak 7 ml dan dimasukkan ke dalam media MRSB ditambah tripton 1% (63 ml). Kultur yang dihasilkan sebanyak 70 ml. Kultur L. fermentum kemudian diinkubasi selama 20 jam pada suhu 370C. Kultur L. fermentum 2B2 umur 20 jam tersebut dimasukkan ke dalam ependorf kemudian disentrifuse selama 20 menit dengan kecepatan 10.000 rpm pada suhu 40C. Supernatan bebas sel yang dihasilkan dari setiap media dipisahkan dengan endapannya dengan dimasukkan dalam tabung untuk diukur pH awal. Supernatan pada setiap media dikondisikan menjadi pH 6,0 dengan penambahan NaOH 1 N. Supernatan bebas sel dengan pH 6,0 disterilkan dengan cara disaring menggunakan filter 0,22 µm (millipore) kemudian ditambahkan amonium sulfat sebanyak 40% dalam setiap media. Gerakkan wadah substrat tersebut secara perlahan hingga seluruh amonium sulfat larut didalam substrat. Substrat didiamkan selama 24 jam dalam lemari pendingin. Substrat yang telah didiamkan semalaman tersebut akan menghasilkan protein yang mengapung diatas substrat. Protein (substrat kasar
bakteriosin) dipisahkan dengan sebagian besar substrat dengan mengeluarkan substrat yang ada di bawahnya, kemudian ditambahkan dengan buffer KH2PO4.
Substrat kasar bakteriosin tersebut digunakan dalam uji antagonistik dengan bakteri indikator.
Produksi Bakteri Indikator sebagai Kultur Kerja. Bakteri indikator S. aureus ATCC 25923 diinkubasi selama 24 jam pada suhu 370C. Bakteri S. aureus ATCC 25923 umur 24 jam yang akan digunakan distandarisasi jumlahnya dengan cara disetarakan kekeruhannya sesuai standar Mc Farland no. 0,5 sehingga dihasilkan populasi bakteri setara 1,5 x 108 sel bakteri/ml.
Penggabungan Substrat Kasar Bakteriosin dengan Enzim Katalase. Enzim katalase (2,0 U/mg) distabilkan dengan buffer 10 mM potasium fosfat yang dikondisikan pada pH 7.0 dengan penambahan NaOH 1 N. Sampel substrat kasar bakteriosin sebanyak 1 ml dicampur dengan 1 mg/ml enzim katalase (1 : 1), kemudian diinkubasi selama 60 menit pada suhu 37oC (Savadogo et al., 2004). Konfrontasi Substrat Kasar Bakteriosin dengan Bakteri Indikator. Substrat kasar bakteriosin yang telah ditambahkan enzim katalase dikonfrontasi dengan bakteri indikator S. aureus ATCC 25923 dengan metode difusi agar. Bakteri indikator S. aureus ATCC 25923 dimasukkan ke dalam cawan sebanyak 1 ml kemudian dituang media MHA sebanyak 25 ml. Cawan digerakkan membentuk angka delapan untuk menghomogenkan media dengan bakteri. Agar yang telah mengeras dilubangi dengan menggunakan pipet Pasteur (diameter 5 mm) dan agarnya dibuang dengan menggunakan ose. Substrat kasar bakteriosin dimasukkan ke dalam lubang tersebut sebanyak 50 µl. Cawan ditutup dengan kertas saring kemudian ditutup dengan penutup cawan. Cawan dimasukkan ke dalam lemari pendingin selama 2 jam kemudian diinkubasi selama 20 jam. Zona bening yang dihasilkan diukur diameternya. Jika terdapat zona hambat pada uji antagonistik anatara bakteri indikator dengan substrat kasar bakteriosin yang diberi enzim katalase, maka menunjukkan bahwa substrat tersebut merupakan bakteriosin tanpa pengaruh dari sifat antimikroba hidrogen peroksida.
Uji Kepekaan Substrat Kasar Bakteriosin terhadap Enzim Proteolitik
Penyegaran Bakteri Asam Laktat Lactobacillus fermentum 2B2. Kultur L. fermentum 2B2 dari media MRSB dipipet sebanyak 1 ml dan dimasukkan ke dalam media MRSB yang ditambah YE 3% (9 ml), kemudian diinkubasi selama 24 jam pada suhu 37oC.
Produksi Bakteriosin dari Lactobacillus fermentum 2B2. Kultur L. fermentum 2B2 hasil penyegaran berumur 24 jam dipipet sebanyak 7 ml dan dimasukkan ke dalam media MRSB ditambah tripton 1% (63 ml). Kultur yang dihasilkan sebanyak 70 ml. Kultur L. fermentum kemudian diinkubasi selama 20 jam pada suhu 370C. Kultur L. fermentum 2B2 umur 20 jam tersebut dimasukkan ke dalam ependorf kemudian disentrifuse selama 20 menit dengan kecepatan 10.000 rpm pada suhu 40C. Supernatan bebas sel yang dihasilkan dari setiap media dipisahkan dengan endapannya dengan dimasukkan dalam tabung untuk diukur pH awal. Supernatan bebas sel dikondisikan menjadi pH 6,0 dengan penambahan NaOH 1 N. Supernatan bebas sel dengan pH 6,0 disterilkan dengan cara disaring menggunakan filter 0,22 µm (millipore) kemudian ditambahkan amonium sulfat sebanyak 40% dalam setiap media. Gerakkan wadah substrat tersebut secara perlahan hingga seluruh amonium sulfat larut didalam substrat. Substrat didiamkan selama 24 jam dalam lemari pendingin. Substrat yang telah didiamkan semalaman tersebut akan menghasilkan protein yang mengapung diatas substrat. Protein dipisahkan dengan sebagian besar substrat dengan mengeluarkan substrat yang ada di bawahnya, kemudian ditambahkan dengan buffer KH2PO4. Substrat kasar bakteriosin tersebut digunakan
dalam uji antagonistik (konfrontasi) dengan bakteri indikator.
Produksi Bakteri Indikator sebagai Kultur Kerja. Bakteri indikator S. aureus ATCC 25923 diinkubasi selama 24 jam pada suhu 370C. Bakteri S. aureus umur 24 jam yang akan digunakan distandarisasi jumlahnya dengan cara disetarakan kekeruhannya sesuai standar Mc Farland no. 0,5 sehingga dihasilkan populasi bakteri setara 1,5 x 108 sel bakteri/ml.
Penggabungan Substrat Kasar Bakteriosin dengan Enzim Proteolitik. Enzim-enzim dan buffernya yang digunakan secara berturut-turut yaitu tripsin (15000 U/mg) dalam 0,05 M buffer Tris Hidroklorid (pH 8,0), dan pepsin (3,2 U/ml) dalam
0,2 M buffer sitrat (pH 3,0). Sampel substrat kasar bakteriosin sebanyak 1 ml dicampur dengan 1 mg/ml masing-masing enzim proteolitik (1 : 1). Substrat kasar bakteriosin yang ditambahkan enzim pepsin diinkubasi selama 60 menit pada suhu 37oC, sedangkan yang ditambahkan dengan enzim tripsin pada suhu 250C (suhu kamar) (Savadogo et al., 2004).
Konfrontasi Substrat Kasar Bakteriosin dengan Bakteri Indikator. Substrat kasar bakteriosin yang telah ditambahkan enzim proteolitik dikonfrontasi dengan bakteri indikator S. aureus ATCC 25923 dengan metode difusi agar. Bakteri indikator S. aureus ATCC 25923 dimasukkan ke dalam cawan sebanyak 1 ml kemudian dituang media MHA sebanyak 25 ml. Cawan digerakkan membentuk angka delapan untuk menghomogenkan media dengan bakteri. Agar yang telah mengeras dilubangi dengan menggunakan pipet Pasteur (diameter 5 mm) dan agarnya dibuang dengan menggunakan ose. Substrat kasar bakteriosin yang telah ditambahkan enzim proteolitik dimasukkan ke dalam lubang tersebut sebanyak 50 µl. Cawan ditutup dengan kertas saring kemudian ditutup dengan penutup cawan. Cawan dimasukkan ke dalam lemari pendingin selama 2 jam kemudian diinkubasi selama 20 jam. Zona bening yang dihasilkan diukur diameternya. Jika tidak terdapat zona hambat pada uji antagonistik antara bakteri indikator dengan substrat kasar bakteriosin yang diberi enzim proteolitik, maka menunjukkan bahwa substrat tersebut merupakan protein.
HASIL DAN PEMBAHASAN
Produksi Bakteriosin
Isolat bakteri asam laktat (BAL) yang digunakan adalah Lactobacillus fermentum 2B2 yang berasal dari daging sapi. Bakteri L. fermentum 2B2 ini berdasarkan penelitian Widiasih (2008) merupakan bakteri Gram positif dengan katalase negatif dan berbentuk batang dengan susunan tunggal maupun rantai pendek. Bakteri ini menghasilkan supernatan antimikroba yang didominasi oleh asam organik yang memiliki zona hambat yang cukup baik terhadap bakteri Gram positif maupun Gram negatif.
Bakteri indikator yang digunakan pada tahap produksi bakteriosin ada 4 spesies berbeda yaitu Staphylococcus aureus ATCC 25923, Eschericia coli ATCC 25922 , Salmonella typhimuriumATCC 14028, dan enteropatogenik Escherichia coli lokal. Pemilihan keempat bakteri indikator disebabkan bakteri-bakteri ini yang sering mencemari makanan bahkan menyebabkan penyakit pada manusia. Bell dan Kyriakides (1998) mengungkapkan bahwa E. coli telah menjadi bakteri indikator pencemaran air dan susu pada awalnya dan berkembang menjadi indikator pada berbagai jenis makanan.
Mikroorganisme seperti halnya makhluk yang lain juga mengalami pertumbuhan. Beberapa faktor utama yang mempengaruhi pertumbuhan mikroorganisme meliputi nutrisi, waktu, suhu, air, pH dan tersedianya oksigen (Buckle et al., 1985). Pertumbuhan yang baik terjadi bila kebutuhan nutrisi tercukupi, demikian pula produksi bakteriosin akan terjadi secara optimum bila nutrisi tersedia dan sesuai dengan kebutuhan untuk mensekresikan zat aktif tersebut oleh L. fermentum 2B2.
Bakteriosin merupakan protein sehingga perlu diujikan sumber protein yang berbeda dalam media, yaitu (1) MRSB dan NaCl 1% ; (2) MRSB, YE 3 %, dan NaCl 1%; (3) MRSB dan tripton 1% yang masing-masing media dikondisikan pada pH 5,0 dan pH 6,0. Pemilihan media tersebut berdasarkan tingkat aktivitas penghambatan senyawa antimikroba yang tinggi pada bakteri indikator yang diuji (Ogunbawo et al., 2003). Media yang digunakan seluruhnya menggunakan MRSB sebagai media pokok. MRSB merupakan media yang mengandung nitrogen dan karbon sebagai sumber nutrisi untuk bakteri tumbuh. Hasil penelitian (Makara, 2002) membuktikan