www.elsevier.com / locate / livprodsci
Neither native nor popped cornmeal in the ration of dry cows
affects magnesium absorption
a ,
*
a b aJ.Th. Schonewille
, A.Th. Van ‘t Klooster , J.W. Cone , H.J. Kalsbeek-Van der Valk ,
a a
H. Wouterse , A.C. Beynen
a
Department of Nutrition, Faculty of Veterinary Medicine, Utrecht University, P.O. Box 80.152, 3508 TD Utrecht, The Netherlands
b
DLO-Institute for Animal Science and Health, Department of Ruminant Nutrition, P.O. Box 65, 8200 AB Lelystad, The Netherlands Received 16 November 1998; received in revised form 19 May 1999; accepted 1 June 1999
Abstract
The hypothesis tested was that addition of starch to the ration of cows would stimulate magnesium (Mg) absorption because of a lowering of ruminal pH, which renders Mg more soluble and thus more available for transport across the epithelium of the rumen, which is the major site of Mg absorption in ruminants. The trial had a 535 Latin-square design in which five non-pregnant, non-lactating multiparous cows were fed rations containing either a mix of cellulose and maizeglutenfeed or native or popped cornmeal each at two levels (equivalent to 11 or 20% starch in the dry matter). The dietary periods lasted 28 days. The amount and type of dietary starch did not significantly affect total gastro-intestinal tract Mg absorption, post-prandial ruminal pH, rumen fluid concentrations of Mg, K and total volatile fatty acids; for all five treatments combined, the measured values were 5.660.45% of intake, 6.560.04, 0.760.12 mmol / l, 41.060.63 mmol / l, and
10562.3 mmol / l (means6S.E., n55), respectively. For all cows and treatments combined, there was a significant, negative relationship between ruminal pH and Mg concentration in rumen fluid. The ruminal Mg concentrations were low when compared with earlier work. On the basis of published in-vitro studies showing that concentrations of soluble Mg fall to more or less constant, low values when pH values are above 6, it is suggested that the high baseline values of ruminal pH in
this study, had prevented a statistically significant effect of starch intake, if any, on Mg absorption. 2000 Elsevier
Science B.V. All rights reserved.
Keywords: Magnesium; Starch; Rumen; Fermentation; Absorption
1. Introduction (Schonewille et al., 1997a). In ruminants, high K
intakes inhibit magnesium (Mg) absorption which Grass silages from areas with intensive livestock enhances the risk of hypomagnesaemia (Fontenot et production are generally high in potassium (K) al., 1989). To safeguard the Mg status of dairy cows fed grass silages rich in K, it seems more appropriate to improve Mg absorption from the diet rather than to supplement the ration with Mg because high Mg *Corresponding author. Tel.: 131-30-2531581; fax: 1
31-30-intakes by ruminants may cause a depressed nutrient 2531817.
E-mail address: [email protected] (J.Th. Schonewille) utilisation (Chester-Jones et al., 1989). 0301-6226 / 00 / $ – see front matter 2000 Elsevier Science B.V. All rights reserved.
In a study with dry goats the replacement of 2. Materials and methods dietary cellulose, at a concentration of 342 g / kg dry
matter, by an equal amount of native corn starch had 2.1. Animals and experimental design a marked stimulatory effect on Mg absorption
(Schonewille et al., 1997b). It is not known whether Five non-pregnant, non-lactating multiparous cows this observation extends to dairy cattle. In any event, (age 10.360.8 y; mean6S.E.), fitted with rumen
the use of high amounts of starch in the ration of cannulas, were used. The cows were of a Friesian– dairy cows entails the risk of ruminal acidosis and Holstein3Holstein–Friesian cross. During the
ex-increases feed costs to a practically unacceptable periment, they were housed in a stanchion barn. level. Thus, to test whether consumption of corn The trial had a 535 Latin-square design and was
starch raises Mg absorption in cows, we used in the preceded by a 14-day pre-experimental period that present study diets that contained either 100 or 200 g allowed the cows to become adapted to rations based starch / kg dry matter in the form of cornmeal. primarily on artificially dried grass. Each experimen-A possible mechanism by which starch could tal period lasted 28 days. The animals were random-increase Mg absorption is a lowering of ruminal pH ly assigned to each sequence of feeding on the five which renders Mg more soluble and thus more experimental rations.
available for transport across the epithelium of the
rumen, which is the major site of Mg absorption in 2.2. Rations ruminants (Rogers and Van ‘t Klooster, 1969; Tomas
and Potter, 1976). Native corn starch is slowly During the pre-experimental period the daily degraded by ruminal bacteria but popping of corn ration of all cows consisted of 4.5 kg artificially starch increases the rate of ruminal fermentation dried grass, 0.2 kg barley straw and 2.2 kg of a (Cone et al., 1989; Cone and Vlot, 1990) and may commercially available concentrate containing 9.0 g further lower ruminal pH (Malestein et al., 1988). Ca, 4.9 g P and 6.3 g Mg / kg. During the experimen-Thus, it could be suggested that popped cornmeal tal periods, the commercial concentrate was replaced enhances Mg absorption more than would native by 0.1 kg of a pelletted experimental concentrate and cornmeal. This suggestion was tested also in this 2.1 kg powdered feed containing the dietary vari-feeding trial with dry cows. ables. The ingredient composition of the
experimen-Table 1
Ingredient composition of the experimental rations
Control Native cornmeal Popped cornmeal
10% starch of dm 20% starch of dm 10% starch of dm 20% starch of dm
a
Constant components (kg / d) 4.8 4.8 4.8 4.8 4.8
b
The constant components consisted of 4.5 kg of artificially dried grass (924 dry matter / kg), 0.2 kg of barley straw (930 g of dry matter / kg), and 0.1 kg of experimental concentrate (910 g of dry matter / kg). The experimental concentrate consisted of 5.0 g of beet vinasses, 92.6 g of beetpulp, 1.4 g of MgO, and 1.0 g premix. The premix consisted of 609.4 mg of CaCO , 3.0 mg of CoSO3 4?7H O, 2.12
mg of Na SeO2 3?5H O, 5.3 mg of KIO , 30.5 mg of MnSO2 3 4?H O, 27.5 mg of ZnSO2 4?H O, 150.0 mg of CuSO2 4?5H O, 17.2 mg of2
retinylacetate (50,000 IU), 5.0 mg of cholecalciferol (200 000 IU), and 150 mgDL-a-tocopherylacetate (150 IU).
b
The control mix (917 g of dry matter / kg) consisted of 63 g of distilled water / kg, 639 g of cellulose / kg, and 298 g of maizeglutenfeed / kg.
c
tal rations is shown in Table 1. The control ration During the last eight days of each experimental contained cellulose and maizeglutenfeed which was period, urine and faeces were collected quantitatively replaced by either native or popped cornmeal so as to from each cow. Urine was collected by using urinals formulate the experimental rations. In essence, this attached to the cows with the use of leather harnas-replacement involved an exchange of cellulose ses. Urine coursed down into two vessels so that
(Arbocel , Internatio B.V., Rotterdam, The Nether- approximately 80% was collected in one vessel. The lands) and corn starch. The native and popped remaining urine was collected in the other vessel cornmeal were from the same batch and were which contained Na-azide as a preservative. Total supplied by Presco International, Weert, The Nether- urine collections from each 24-h period were lands. The analysed composition of the whole rations weighed, and 0.5% of the total urine was sampled is shown in Table 2. from the preserved urine and stored at 2188C in a
The animals were fed individually. On an energy plastic bottle which contained 100 ml 6 M HCl. The basis, the rations provided approximately the require- daily faeces production of each cow was mixed ment for maintenance. The rations were offered thoroughly, and 3% of the wet weight was stored at twice daily in two equal portions at 08:00 and 17:00 2188C. At the end of each collection period, the
h. Feed refusals, if any, were recorded. stored faeces fractions of each cow were combined, mixed thoroughly and sampled. The samples were 2.3. Collection of samples dried at 608C for four days, ground, and stored in
air-dry form in sealed jars at room temperature until During the last week of each experimental period, analysis.
the experimental feedstuffs were sampled daily and On day 19, approximately 10 min prior to the then pooled, ground and stored in sealed jars at room morning meal, 350 ml of Cr-EDTA solution (100 g temperature. Cr-EDTA / l) was injected into the rumen as a marker Blood samples were taken on day 21 of each for estimating the rumen volume and passage rates of experimental period. Between 16:00 and 16:30 h, the liquid phase from the rumen. Rumen liquid before the afternoon meal, blood was sampled from samples (approximately 30 ml) were taken at 07:45, the jugular vein into evacuated heparinized tubes. 09:00, 10:00, 11:00, 13:00, 15:00 and 17:00 h. The blood samples were centrifuged for 10 to 15 min Immediately after collection, pH of ruminal fluid was at approximately 2700 g, and the plasma was col- recorded and the rumen liquid samples were cen-lected and stored at 2188C. trifuged at room temperature at 2700 g for 15 min,
Table 2
Analysed composition of the experimental rations (g / kg dry matter)
Control Native cornmeal Popped cornmeal
10% starch of dm 20% starch of dm 10% starch of dm 20% starch of dm
Crude protein 179 185 191 184 188
Crude fat 45 49 53 50 55
Crude ash 107 106 105 106 105
Crude fibre 337 260 181 260 180
Neutral detergent fibre 643 551 459 551 458
Acid detergent fibre 377 283 187 281 184
Acid detergent lignine 19 19 19 18 17
Starch 23 112 201 114 205
Mg 1.97 1.95 1.92 1.95 1.93
K 33.1 33.3 33.5 33.3 33.5
Na 1.17 1.10 1.04 1.10 1.03
and the supernatant was stored in plastic tubes at stuffs and faeces was estimated by atomic absorption
2188C. An aliquot of the supernatant from the spectroscopy (Perkin-Elmer 3110; Perkin-Elmer,
rumen liquid samples taken at 07:45, 09:00, 11:00, Norwalk, CT, USA). Calcium and K in feedstuffs 13:00, 15:00 and 17:00 h was centrifuged at 208C at and K in supernatant of rumen fluid were estimated
30,000 g for 30 min and the supernatant was stored by atomic absorption spectroscopy and Na in feed-in plastic tubes at 2188C. From the rumen liquid stuffs by atomic emission spectroscopy (Perkin
samples taken at 07:45, 09:00, 11:00, 13:00, 15:00 Elmer 3110). Magnesium in plasma, urine, superna-and 17:00 h, 2 ml non-centrifuged fluid was de- tant of rumen fluid, and chromium (III) in rumen proteinised according to the method of Bergmeyer fluid were measured directly by atomic absorption (1970). After 10 min, the de-proteinised rumen spectroscopy. The accuracy of each assay run was liquid samples were centrifuged at room temperature monitored using a commercial reference sample (hay at 2700 g for 15 min and the supernatant was powder, CRM 129; Community Bureau of Refer-collected and stored in plastic tubes at 2188C until ence, Brussels, Belgium) and laboratory reference
analysis of volatile fatty acids (VFAs). samples, and was found to be within 5% deviation from the target values. The combined within and 2.4. Chemical analyses between-run precision of the determinations was
#3.0% (coefficient of variation).
Samples of the feedstuffs were subjected to the VFAs in rumen fluid were determined gas-Weende analysis. Nitrogen contents were determined chromatographically (Perkin Elmer Autoscan) with a by the macro-Kjeldahl method (IDF, 1986); a factor 2 m glass column (Chromosorb WAW DMCS; 17% of 6.25 was used to convert g of N into crude neopentyl glycol adipate, 1% H PO ). Pivalic acid3 4 protein. Ether extracts of the feedstuffs were pre- (0.045%, w / v) was added to the de-proteinised pared according to the AOAC (1984); the solvent ruminal fluid as an internal standard.
was evaporated and the crude-fat residue weighed.
The crude fibre contents of the feedstuffs were 2.5. Statistical analyses estimated using the Fibertec System M2 (Tecator,
Stockholm, Sweden). The NDF, ADF and ADL All data were checked for normal distribution content of the feedstuffs were estimated according to using the Kolmogorov–Smirnov test and then were the methods described by Goering and Van Soest subjected to analysis of variance (ANOVA) with (1970). To determine the starch content of the animal, experimental period and dietary treatment as feedstuffs, they were enzymatically treated with factors (Wilkinson, 1990). Rumen-liquid mineral amyloglucosidase from Aspergilles niger (EC data were subjected to repeated measurement analy-3.2.1.3) to hydrolyse all starch to glucose (Keppler ses with animal, experimental period and dietary and Decker, 1974). Subsequently, glucose was mea- treatment as factors (Wilkinson, 1990). When a sured enzymatically with a test combination (Boeh- dietary factor had a statistically significant influence, ringer Mannheim Diagnostica, Mannheim, Germany) Bonferroni’s test was used to identify the diets that and a computerised autoanalyser (Beckman had different effects on the variable involved.
Synchron CX Systems; Beckman, Mijdrecht, The Throughout, the level of statistical significance was Netherlands). The free glucose content of the feed- pre-set at P,0.05.
stuffs was measured directly. Starch was calculated as total glucose minus free glucose. For the
ex-perimental feedstuffs, i.e., the control mix and the 3. Results two types of cornmeal, the in-vitro rate of
fermen-tation was determined according to the method 3.1. In-vitro fermentation of the dietary variables described by Cone et al. (1996). Prior to the
de-termination of the selected minerals in feedstuffs and To check the fermentation profiles of the dietary faeces, the samples were ashed (4808C for 6 h) and variables, the control mix, native and popped
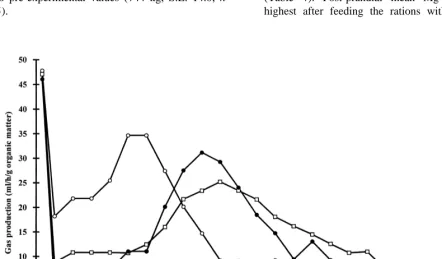
gas production was measured (Cone et al., 1996). 3.3. Magnesium balance and plasma Mg The high rate and subsequent decline in the rate of
gas production during the first hour of the incuba- Magnesium intake was similar for all five dietary tions (Fig. 1) probably represents the degradation of treatments (Table 3). Upon ANOVA, faecal excre-the water-soluble fraction of excre-the experimental feed- tion of Mg was found to be significantly affected by stuffs (Cone et al., 1997). After 5 h of incubation, the factor diet (P50.013), but significant differences
the incubation containing the popped cornmeal had between specific rations could not be identified by the highest rate of gas production. After 10 h, the Bonferroni’s t-test. Consequently, Mg absorption and rate had fallen to approximately 20% of the peak urinary excretion were similar for the treatments. value, but now the control mix and native cornmeal The dietary treatments did not affect (P50.205)
produced their peak values. After 20 h the rate of plasma Mg. For all treatments combined, mean total gas production had returned to baseline values plasma Mg concentration was 0.80 mmol / l (S.E. for the three feedstuffs. 0.014; n55).
3.2. Feed intake and body weight 3.4. Rumen liquid: Mg and K concentration, pH
and VFAs
Experimental rations were consumed completely
throughout the experiment. Mean body weight at the Mg and K concentrations in rumen liquid, before end of the experiment was 746 kg (S.E. 16.0; n55), the morning feed and also after feeding, were not
which was almost identical (P50.803; paired t-test) significantly influenced by the dietary treatments
to pre-experimental values (744 kg, S.E. 14.0; n5 (Table 4). Post-prandial mean Mg values were
5). highest after feeding the rations with either 20%
Table 3
Magnesium balance after the feeding of the experimental rations
Control Native cornmeal Popped cornmeal Pooled S.E.M. P
10% starch 20% starch 10% starch 20% starch
of dm of dm of dm of dm
nd5Not determined because the cows were offered a restricted amount of feed.
Table 4
Concentrations of Mg and K in rumen liquid (mmol / l), total volatile fatty acids (VFAs in mmol / l) and ruminal pH in cows fed the experimental rations
Control Native cornmeal Popped cornmeal Pooled S.E.M. P
10% starch 20% starch 10% starch 20% starch
of dm of dm of dm of dm
Ruminal Mg concentration
07.45 0.32 0.28 0.45 0.56 0.35 0.169 0.789
a
Post prandial 0.49 0.60 0.96 0.49 1.03 0.194 0.191
Ruminal K concentration
07.45 23.3 23.5 25.3 23.9 25.0 0.771 0.309
Post prandial 42.0 41.1 42.0 38.3 41.6 0.993 0.102
Ruminal VFA concentration
07.45 93.7 97.6 95.5 95.8 97.1 2.980 0.898
Post prandial 105.0 100.9 102.1 102.3 113.8 3.594 0.141
Ruminal pH
07.45 6.73 6.67 6.71 6.71 6.65 0.045 0.716
b b c
Post prandial 6.49 6.52 6.52 6.49 6.34 0.040 0.038
a
Post-prandial values are geometrical means for five values; i.e., these values can be considered as an estimation of the area under the curve (Wolever and Jenkins, 1986). Cows were given a restricted amount of feed at 08:00 and 17:00 h.
b,c
Values in the same row with different superscript were borderline significantly different (P,0.10), Bonferroni test).
native or popped cornmeal. The ruminal pH de- mean post-prandial concentrations of total VFAs creased significantly (P,0.05) after feeding, irre- were highest after feeding the ration with 20%
spective of dietary treatment. Post-prandial ruminal popped cornmeal, but the increase versus that for the pH was borderline significantly lower after feeding other treatments was not statistically significant. of the ration with 20% popped cornmeal instead of Post-prandial concentrations (mM) of total VFAs, native cornmeal. acetate, and propionate, and the acetate-to-propionate Before the morning feed, concentrations of total ratio were similar (P.0.100) for the five rations; for
VFAs were similar for all dietary treatments. After all treatments combined (n55), the mean values
feeding, total VFA concentrations were significantly were 104.8 (S.E. 2.341), 72.5 (S.E. 1.648), 15.1 (P,0.05) increased when either the control ration or (S.E. 0.356) and 4.8 (S.E. 0.047), respectively. Upon
ruminal fluid were found to be significantly affected 1987) is not yet fully understood. A possibility is by the factor diet (P50.044), but Bonferroni’s t-test that the addition of starch to the ration may stimulate
did not identify specific rations that produced differ- Mg absorption by increasing the rate of ruminal ent values. The butyrate concentration for all treat- fermentation and decreasing ruminal pH. A lowering ments combined was 13.8 mM (S.E. 0.432) of ruminal pH renders Mg more soluble and thus more available for transport across the epithelium of 3.5. Rumen volume and passage rate the rumen which is the major site of Mg absorption in ruminants (Rogers and Van ‘t Klooster, 1969; Rumen volume, absolute and fractional outflow of Tomas and Potter, 1976). The discrepancy between the liquid phase were not affected (P.0.807) by the outcome of our study and that of Wilson et al.
any dietary treatment, the combined values for all (1969) might be related to a difference in starch-treatments (n55) being 70 l (S.E. 1.4), 5.9 l / h (S.E. induced change of ruminal pH. Feed intake in our
0.07) and 8.7% / h (S.E. 0.215), respectively. study was approximately three-times lower than that in the study of Wilson et al. (1969). The level of feed intake is negatively associated with the
buffer-4. Discussion ing capacity of ruminal fluid (Robinson et al., 1986).
Thus, the pH-lowering effect of supplemental starch In this feeding trial, Mg absorption was not might have been greater in the study of Wilson et al. significantly affected by the starch content of the (1969) so that Mg absorption was effectively in-ration. In studies with goats (Schonewille et al., creased which in turn allowed the cows to maintain 1997b) and sheep (Pfeffer et al., 1970; Giduck and their plasma Mg values.
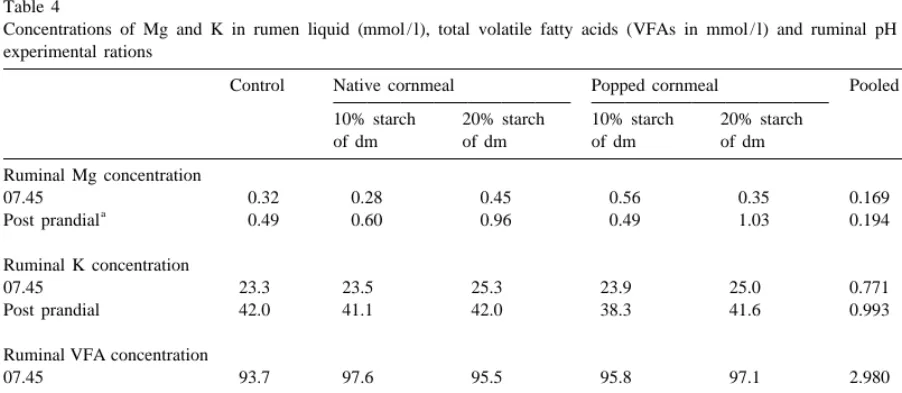
Fontenot, 1987) rations with a starch content of at For all dietary treatments combined, a significant least 298 g / kg dry matter significantly increased Mg (P50.009), negative relationship was found
Fig. 2. Relationship between pH of ruminal fluid and Mg concentration in ultracentrifuged rumen fluid [j control,nnative cornmeal
(10%),mnative cornmeal (20%), spopped cornmeal (10%), dpopped cornmeal (20%)]. Data correspond with ante- and postfeeding 2
rumen samples (6 time points / dietary treatment) The solid line represents the regression line; y5 21.3766x19.6215 (R 519.5%; adj
P 50.009, P 50.005). model,pH constant
fermentation kinetics measured with a fully automated time short-chain fatty acids produced during ruminal
related gas production apparatus. Anim. Feed Sci. Technol. 61, fermentation of dietary carbohydrates may stimulate
113–128.
Mg absorption by the delivery of protons to the CVB (Centraal Veevoeder Bureau), 1997. Voedernormen Land-21 1
Mg / 2 H exchangers located in the apical mem- bouwhuisdieren en Voederwaarde Veevoeders, CVB, Lelystad. Dalley, D.E., Isherwood, P., Sykes, A.R., Robson, A.B., 1997. brane of the epithelium. However, total VFA
con-Effect of in vitro manipulation of pH on magnesium solubility centrations were not significantly altered by the
in ruminal and caecal digesta in sheep. J. Agric. Sci. Camb. feeding of starch. Since ruminal Mg concentrations 129, 107–111.
were very low in our study, both the K-sensitive and Fontenot, J.P., Allen, V.G., Bunce, G.E., Goff, J.P., 1989. Factors influencing magnesium absorption and metabolism in rumin-the K-insensitive transport component facilitated
ants. J. Anim. Sci. 67, 3445–3455. only low rates of ruminal Mg absorption. These
Giduck, S.A., Fontenot, J.P., 1987. Utilization of magnesium and considerations explain why the observed percentages other macrominerals in sheep supplemented with different of apparent Mg absorption were relatively low and readily-fermentable carbohydrates. J. Anim. Sci. 65, 1667– why no effect of starch feeding was observed. 1673.
Goering, H.K., Van Soest, P.J., 1970. Forage fiber analyses In conclusion, this study shows that the addition of
(apparatus, procedures and some applications). In: Agric. cornmeal to the ration of dry cows did not
sig-Handbook No. 379, ARS-USDA, Washington, DC.
nificantly influence Mg absorption. It is suggested IDF (International Dairy Federation), 1986. International IDF that the high baseline values of ruminal pH and high Standard 20A, International Dairy Federation, Brussels. buffering capacity of ruminal fluid had prevented an Kemp, A., Deijs, W.B., Hemkes, O.J., Van Es, A.J.H., 1961.
Hypomagnesaemia in milking cows:intake and utilization of effect of starch intake, if any, on Mg absorption.
magnesium from herbage by lactating cows. Neth. J. Agric. Sci. 9, 134–149.
Keppler, D., Decker, K., 1974. In: Bergmeyer, H.U. (Ed.), Acknowledgements Methods of Enzymatic Analysis, 2nd ed, Academic Press, New
York.
¨
Leonhard, S., Martens, H., Gabel, G., 1989. New aspects of This study was supported by the Product Board
magnesium transport in ruminants. Acta Vet. Scand. 86 Animal Feed (Produktschap Diervoeder), The Hague,
(Suppl.), 146–151.
The Netherlands. Wim Lensink is thanked for his Leonhard-Marek, S., Martens, H., 1996. Effects of potassium on biotechnical assistance as is Jan Van Der Kuilen for magnesium transport across rumen epithelium. Am. J. Physiol. chemical analysis. 271, G1034–G1038.
Malestein, A., Van ‘t Klooster, A.Th., Cone, J.W., 1988. De-gradability of various types of starch by incubation with rumen fluid or bacteriala-amylase. J. Anim. Physiol. Anim. Nutr. 59,
References 225–232.
¨
Martens, H., 1979. In vivo untersuchungen uber die absorption ¨ Association of Official Analytical Chemists, 1984. Official Meth- von magnesium aus dem pansen von schafen. Eine abschatzung
¨ ¨
ods of Analysis, 14th ed., AOAC, Arlington, VA. der maximalen absorptionskapazitat des pansens. Berl. Munch. ¨
Care, A.D., Brown, R.C., Farrar, A.R., Pickard, D.W., 1984. Tierarzl. Wchr. 92, 152–155.
Magnesium absorption from the digestive tract of sheep. Q. J. Martens, H., Blume, I., 1986. Effect of intraruminal sodium and Exp. Physiol. 69, 577–587. potassium concentrations and of the transmural potential Chester-Jones, H., Fontenot, J.P., Veit, H.P., Webb, Jr. K.E., 1989. difference on magnesium absorption from the temporarily Physiological effects of feeding high levels of magnesium to isolated rumen of sheep. Quart. J. Exp. Physiol. 71, 409–415.
¨
sheep. J. Anim. Sci. 67, 1070–1081. Martens, H., Gabel, G., Strozyk, H., 1987. The effect of potassium Cone, J.W., Vlot, M., 1990. Comparison of degradability of starch and the transmural potential difference on magnesium transport in concentrates by enzymes and rumen fluid. J. Anim. Physiol. across an isolated preparation of sheep rumen epithelium. Anim. Nutr. 63, 142–148. Quart. J. Exp. Physiol. 72, 181–188.
´
Cone, J.W., Cline-Theil, W., Malestein, A., Van ‘t Klooster, A.Th., Martens, H., Harmeyer, J., 1978. Magnesium transport by isolated 1989. Degradation of starch with rumen fluid. A comparison of rumen epithelium of sheep. Res. Vet. Sci. 24, 161–168.
¨
different starch sources. J. Sci. Food Agric. 49, 173–183. Martens, H., Leonard, S., Gabel, G., 1991. Minerals and digestion: Cone, J.W., Van Gelder, A.H., Driehuis, F., 1997. Description of exchanges in the digestive tract. In: Jouany, J.P. (Ed.), Rumen gas production profiles with a three-phasic model. Anim. Feed Microbial Metabolism and Ruminant Digestion, Inst. Natl. Sci. Technol. 66, 31–45. Rech. Agron, Paris, pp. 199–226.
inorganic elements in the digestive tract on rations containing or glucose in the diet and the effects on apparent magnesium different proportions of hay and rolled barley. Br. J. Nutr. 24, absorption in goats. J. Dairy Sci. 80, 1738–1743.
197–204. Schonewille, J.Th., Van ‘t Klooster, A.Th., Van Mosel, M., 1992. A Ram, L., Schonewille, J.T., Martens, H., Van ‘t Klooster, A.T., comparative study of the in vitro solubility and availability of Beynen, A.C., 1998. Magnesium absorption by whethers fed magnesium from different sources. Tijdschr. Diergeneesk. 117, potassium bicarbonate in combination with different dietary 105–108.
magnesium concentrations. J. Dairy Sci. 81, 2485–2492. Schonewille, J.Th., Van ‘t Klooster, A.Th., Beynen, A.C., 1994. Robinson, P.H., Tamminga, S., Van Vuuren, A.M., 1986. Influence High phosphorus intake depresses apparent magnesium absorp-of declining level absorp-of feed intake and varying proportion absorp-of tion in pregnant heifers. J. Anim. Physiol. Anim. Nutr. 71, starch in the concentrate on rumen fermentation in dairy cows. 15–21.
Livest. Prod. Sci. 15, 173–189. Tomas, F.M., Potter, B.J., 1976. The site of magnesium absorption Rogers, P.A.M., Van ‘t Klooster, A.Th., 1969. Observations on from the ruminant stomach. Br. J. Nutr. 36, 37–45.
digestion and absorption of food along the gastrointestinal tract Van ‘t Klooster, A.Th., 1967. De toestand van calcium, mag-of fistulaled cows. 3 The fate mag-of Na, K, Ca, Mg and P in the nesium en enkele andere mineralen in darminhoud en mest van digesta. Meded. LandbHogesch. Wageningen 69–11, 26–39. herkauwers in verband met hun resorptie. Thesis, Meded. Scharrer, E., Lutz, T., 1990. Effects of short chain fatty acids and LandbHogesch. Wageningen, p. 67-5.
K on absorption of Mg and other cations by the colon and Wilkinson, L., 1990. Systat: the System For Statistics, SYSTAT caecum. Z. Ernaehrwiss. 29, 162–168. Inc, Evanston.
Schonewille, J.Th., Ram, L., Van ‘t Klooster, A.Th., Wouterse, H., Wilson, G.F., Reid, C.S.W., Metson, A.J., Butler, G.W., 1969. Beynen, A.C., 1997a. Intrinsic potassium in grass silage and Influence of starch and peanut oil on plasma magnesium in magnesium absorption in dry cows. Livest. Prod. Sci. 48, grazing dairy cows. NZ J. Agric. Res. 12, 467–488. 99–110. Wolever, T.M.S., Jenkins, D.J.A., 1986. The use of the glycemic Schonewille, J.Th., Ram, L., Van ‘t Klooster, A.Th., Wouterse, H., index in predicting the blood glucose response to mixed meals.

![Fig. 2. Relationship between pH of ruminal fluid and Mg concentration in ultracentrifuged rumen fluid [j(10%), control, n native cornmeal m native cornmeal (20%), s popped cornmeal (10%), d popped cornmeal (20%)]](https://thumb-ap.123doks.com/thumbv2/123dok/1051848.933259/8.612.50.499.77.330/relationship-ruminal-concentration-ultracentrifuged-cornmeal-cornmeal-cornmeal-cornmeal.webp)
