RINGKASAN SKRIPSI
PROGRAM STUDI SARJANA BIOLOGI SITH – ITB Semester I 2012/13 (Oktober 2012)
Judul : Analisis ekspresi gen Ma-ACS5 selama proses pematangan buah pisang Cavendish (Musa sp. AAA Group) dalam Fruit Storage Chamber (FSC) dengan menggunakan metode Real-Time PCR (qPCR)
Nama : Indra Chandra Paramaputra (10606043) Pembimbing : Dr. Fenny Martha Dwivany
Kelompok Keilmuan : Genetika dan Bioteknologi Molekuler
I. Pendahuluan
Indonesia telah dikenal sebagai produsen buah untuk konsumsi masyarakat lokal maupun global. Salah satu jenis buah yang banyak diekspor oleh Indonesia adalah buah pisang. Produksi buah pisang dari tahun ke tahun selalu terjadi peningkatan. Walaupun terjadi peningkatan produksi, jumlah ekspor pisang sendiri mengalami penurunan setiap tahunnya. Hal ini dikarenakan pada saat ini sistem penyimpanan dan pengiriman pasca-panen yang ada di Indonesia masih kurang baik, sehingga banyak hasil panen yang busuk sebelum sampai pada konsumen. Proses pematangan buah pisang sangat dipengaruhi oleh keberadaan etilen yang disintesis buah secara alami dari prekursor S-AdoMet (S-adenosil-methionin/ SAM) dan ACC (1-aminosiklopropan-1-asam karboksilat). Proses tersebut dikatalisis oleh ACC sintase (ACS) dan ACC oksidase (ACO) yang secara berurutan dikode oleh gen ACS dan Ma-ACO (Liu et al., 1999; Inaba et al., 2007). Keberadaan etilen kemudian menginduksi perubahan metabolisme dengan mengaktivasi beberapa gen yang berhubungan dengan pematangan dan respons penuaan. Teraktivasinya gen-gen tersebut akan menginduksi beberapa proses yang terjadi di dalam buah, diantaranya produksi autokatalitik etilen, modifikasi dinding sel buah, produksi senyawa volatil, perubahan metabolisme lemak dan karbohidrat (Bierhals et al., 2004). Akibatnya, ketika produksi etilen melonjak maka buah akan cepat matang dan kemudian membusuk. Upaya yang dapat dilakukan untuk memperpanjang masa penyimpanan adalah dengan memperlambat laju metabolisme yang berhubungan dengan pematangan buah. Dalam penelitian ini akan dilakukan pengembangan studi fruit storage chamber (FSC) sebagai alternatif metode penyimpanan buah pasca-panen, agar kualitas buah pasca-panen tetap baik dan segar. Pengembangan studi FSC yang dilakukan pada penelitian ini adalah dengan mengkondisikan buah pisang berada di dalam chamber yang diberi perlakuan pemberian gas dengan komposisi gas N2:CO2:O2 = 90:5:5.
II. Bahan dan Metode
Bahan dan perlakuan
Dalam penelitian ini digunakan buah pisang cavendish merk Sunpride yang dipesan dari PT. Sewu Segar Nusantara, Bandung, Jawa Barat. Pisang yang digunakan merupakan pisang yang dipanen pada usia 12 minggu sehingga masih berwarna hijau muda. Selain itu, pisang yang dipilih untuk penelitian ini tidak memiliki luka pada kulit pisang nya. Buah kemudian dipisahkan satu-persatu kemudian secara acak dimasukkan ke dalam FSC yang terdiri dari lima chamber kaca saling terhubung dan disegel. Komposisi gas yang digunakan untuk eksperimen adalah gas nitrogen : karbondioksida : oksigen = 90:5:5. Untuk perlakuan kontrol, pisang disimpan di dalam chamber yang terbuka sehingga udara atmosfer dapat masuk. Sampel pisang yang telah diberi perlakuan akan dianalisis ekspresi gen nya dengan mengisolasi RNA pisang tersebut.
Isolasi RNA total dan analisis RT-PCR
Sampel jaringan yang telah disimpan sebelumnya kemudian digunakan untuk isolasi RNA dengan metode Zhuang et al (2006) yang telah dimodifikasi. Setelah isolasi dilakukan proses DNAse untuk menghilangkan DNA genomik dengan menggunakan kit Fermentas (No. katalog #EN0521). Setelah itu sintesis untai pertama cDNA disintesis dengan mengggunakan iScript cDNA synthesis KIT dari BioRad (No. katalog 170-8890). Gen terkait biosintesis etilen kemudian diamplifikasi dengan primer spesifik, yaitu Ma-ACS5 (TTAATTAACGTCGGATATGT dan TATCCTCGTTTCCTATAACA) dengan Ma-GAPDH sebagai pembanding (TCAACGACCCCTTCATCAC dan AGCAGCCTTG-TCCTTGTCA). Analisis kurva leleh dan optimasi primer yang digunakan telah dilakukan pada penelitian sebelumnya (Karmawan 2009; Handayani, 2010). PCR diamplifikasi sebanyak 35 kali dengan 94oC selama 30 detik, 47.6 oC dan 72 oC selama 30 detik.
Analisis level mRNA dengan metode Real-Time PCR (qPCR)
Kuantifikasi level mRNA dilakukan dengan instrumen Bio-Rad iCycler® CFX 96™ Thermal Cycler yang terhubung dengan iQ™5 Real-Time PCR Detection Systems. Pewarna yang digunakan adalah SsoFast™ EvaGreen® Supermix (No. katalog #172-5200) dengan volume akhir 20µl. Filter tips Xcluda™ digunakan untuk mencampurkan seluruh komponen reaksi ke dalam pelat Microseal 96-Well Skirted PCR Plates Low-Profile™. Pada saat eksperimen, sampel direplikasi sebanyak tiga kali, kontrol negatif (non template control / NTC dan air bebas-nuklease) juga disertakan dalam setiap kali analisis. Reaksi qPCR diamplifikasi sebanyak 40 kali dengan 95 oC selama 10 detik dan 47.6 oC selama 30 detik.
Analisis kuantifikasi relatif dari setiap gen terkait biosintesis etilen ditentukan dengan rumus 2-ΔΔCt (Livak et al., 2001) dengan menggunakan Ma-GAPDH sebagai gen pembanding.
III. Hasil dan Pembahasan
Isolasi RNA total dan analisis RT-PCR
absorbansi pada panjang gelombang 280/260 nm. Nilai rasio hasil isolasi RNA yang tergolong murni berkisar antara 1,7-2,2 (Lassois et al., 2008). Berdasarkan hasil penelitian yang dilakukan oleh Robertlee (2012), isolasi RNA tidak terdegradasi dan memiliki rasio antara 1,85-2,0. Nilai tersebut menunjukkan bahwa sampel memiliki kemurnian yang baik. Hasil elektroforesis menunjukkan RNA yang diperoleh tidak mengalami degradasi sehingga dapat dilanjutkan dengan perlakuan DNase kemudian sintesis cDNA.
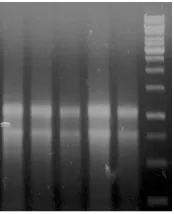
Gambar III.1. Elektroferogram sampel RNA total yang diisolasi dari jaringan daging buah pisang. Diperoleh isolat RNA total dengan rasio λ260/280 berkisar antara 1,85-2,1. Digunakan
ladder 1 kb fermentas sebagai pembanding.
Setelah mendapatkan isolat RNA total, tahap selanjutnya adalah DNAse. Proses ini dilakukan untuk menghilangkan kontaminasi DNA genomik. Setelah DNAse, tahap selanjutnya adalah sintesis cDNA dengan menggunakan cDNA synthesis kit (iScript cDNA synthesis KIT dari BioRad). Setelah sintesis cDNA, dilakukan amplifikasi gen target (Ma-ACS5) dan gen kontrol (Ma-GAPDH) dengan metode RT-PCR. Fragmen gen Ma-ACS5 berhasil diamplifikasi dari cDNA dengan metode RT-PCR. Hasil yang diperoleh pada amplifikasi Ma-ACS5 memiliki ukuran fragmen yang sesuai bila dibandingkan dengan ladder yang digunakan, yaitu 143 bp. Hasil tersebut sesuai dengan penelitian sebelumnya bahwa gen Ma-ACS5 terekspresi selama proses pematangan (Handayani, 2010). Sampel cDNA yang sama juga digunakan sebagai cetakan pada RT-PCR gen Ma-GAPDH sebagai gen referensi. Elektroferogram yang diperoleh menunjukkan ukuran fragmen yang sesuai, yaitu sekitar 236 bp.
Analisis ekspresi gen Ma-ACS5 dengan metode Real-Time PCR (qPCR)
Setelah mengkonfirmasi bahwa seluruh sampel cDNA dapat diamplifikasi dengan menggunakan gen terkait sintesis etilen Ma-ACS5 dan Ma-GAPDH sebagai gen referensi, tahap selanjutnya adalah analisis level ekspresi gen dengan metode qPCR. Pasangan primer yang digunakan untuk menganalisis GOI (gen of interest / gen target)dan GR (gen referensi) telah diuji dengan analisis kurva leleh pada penelitian sebelumnya (Karmawan et al., 2009 dan Handayani, 2010). Kuantifikasi ekspresi gen dari semua sampel dilakukan dengan menggunakan Eva Green sebagai pewarna fluoresens. Eva Green tidak berpendar ketika berada bebas di dalam larutan, tetapi pendarannya akan sangat meningkat cepat ketika berikatan dengan DNA rantai ganda (double stranded DNA). Kemudian peningkatan sinyal Eva Green itulah yang berkorelasi dengan jumlah amplikon selama PCR.
Metode yang digunakan adalah kuantifikasi relatif dengan rumus 2-ΔΔCt (Livak et al.,
hari ke-7. Hal ini sesuai dengan literatur yang menyebutkan bahwa saat buah memasuki tahapan klimaterik maka akan terjadi lonjakan produksi etilen selama pematangan kemudian tiba-tiba menurun secara cepat. Peningkatan konsentrasi etilen ini diketahui menjadi faktor utama dalam proses pematangan buah seperti pisang, alpukat, tomat dan melon (Bower et al., 2002). Setelah tahapan tersebut, produksi etilen kemudian menurun dan disebut dengan tahapan post-klimaterik (Hoffman dan Yang, 1980). Yang dan Hoffman (1984) menyebutkan bahwa pada beberapa spesies tumbuhan, produksi etilen meningkat secara dramatis selama germinasi, penuaan daun dan bunga serta absisi dan pematangan buah. Dalam hal ini, etilen direspon oleh protein reseptor dan sinyal tersebut ditransduksikan melalui kaskade fosforilasi yang kemudian menyebabkan terjadinya aktivasi faktor transkripsi dari ekspresi gen-gen yang berhubungan dengan etilen (Chang et al, 1996). Burg dan Burg (1965) juga menyebutkan bahwa pisang memperlihatkan adanya lonjakan produksi etilen secara tiba-tiba dan kemudian menurun dengan sangat cepat.
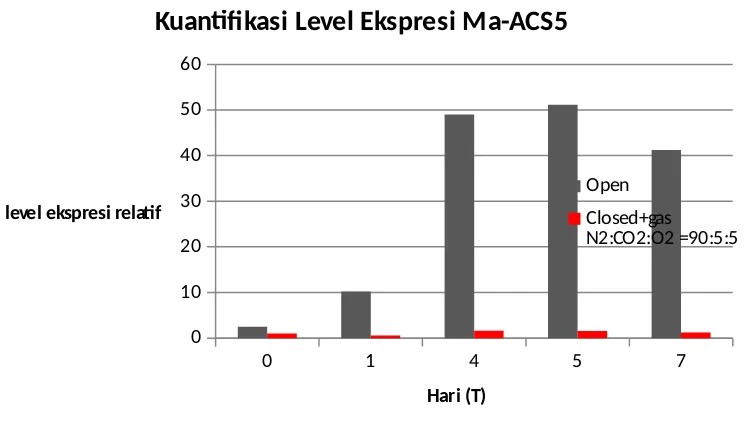
Pada sampel buah pisang dengan perlakuan closed chamber dengan pemberian gas nitrogen : karbondioksida : oksigen = 90:5:5, pola ekspresi Ma-ACS5 mengalami peningkatan hanya 1,5 kali lebih besar pada hari ke-5 dibandingkan hari ke-0, kemudian kembali menurun di hari ke-7. Hal ini berbeda jauh apabila dibandingkan dengan perlakuan kontrol. Level ekspresi gen dari hari ke-0 hingga hari ke-7 hanya mengalami sedikit peningkatan. Rendahnya ekspresi Ma-ACS5 diduga berpengaruh terhadap penurunan jumlah etilen akibat produksi enzim ACC oksidase yang berkurang, sehingga mengakibatkan penundaan pematangan buah yang disimpan pada kondisi tersebut. Penundaan proses pematangan pada buah pisang ini salah satunya diakibatkan karena buah disimpan dalam komposisi gas oksigen yang rendah sehingga menghambat kerja enzim ACC oksidase di dalam biosintesis etilen.
0 1 4 5 7
0 10 20 30 40 50 60
Kuantifikasi Level Ekspresi Ma-ACS5
Open Closed+gas N2:CO2:O2 =90:5:5
Hari (T) level ekspresi relatif
IV. Kesimpulan
Pada penelitian ini, pemberian komposisi gas dalam FSC diuji coba untuk menentukan profil ekspresi gen Ma-ACS5. Hasil RT-PCR menunjukkan bahwa Ma-ACS5 terdeteksi pada seluruh kondisi penyimpanan. Ekspresi gen Ma-ACS5 dianalisis menggunakan metode kuantifikasi relatif qPCR dengan GAPDH sebagai gen pembanding. Pola ekspresi Ma-ACS5 pada open chamber mengalami peningkatan 10 kali lebih besar setelah 1 hari penyimpanan dan meningkat hingga 50 kali lebih besar pada hari ke-5, kemudian menurun sekitar 20% pada hari ke-7. Pola ekspresi Ma-ACS5 pada closed chamber dengan komposisi gas N2 : CO2 : O2 = 90 : 5 : 5 mengalami peningkatan hanya 1,5 kali lebih besar pada hari ke-5
dibandingkan hari ke-0, kemudian kembali menurun di hari ke-7. Profil level ekspresi Ma-ACS5 cenderung meningkat pada buah pisang yang matang secara alami, sedangkan type starch-debranching enzyme from banana fruit J. Agric. Food Chem. 52: 7412 – 7418
Bower, J. Holford, P., Latché, A., Pech, J.C. 2002. Culture conditions and detachment of the fruit influence the effect of ethylene on the climacteric respiration of melon. Postharvest Biol. Technol., 26 : 135 - 146.
Burg SP, Burg EA. 1965. Relationship between ethylene production and ripening in bananas. Botanical Gazette. 126, 200– 204.
Chang SJ, Puryear JD, Dias MADL, Funkhouser EA, Newton RJ, Cairney J. 1996. Gene expression under water deficit in loblolly pine (Pinus taeda) : Isolation and characterization of cDNA clones.
Physiologia Plantarum 95 : 139 – 148. Handayani R. U. 2010. Studi pola ekspresi
Hoffman, N.E. and SF. Yang. 1980. Changes of 1-Aminocyclopropane-l-carboxilic acid content in ripening fruits in relation to their ethylene production rates. J. Am. Sot. Hort. Sci., 105 (1980) 492-495.
Inaba A., Liu X., Yokotani N., Yamane M., Lu W. J., Nakano R., Kubo Y. 2007. Studies on the internal feedback regulation of ethylene biosynthesis and signal transduction during fruit ripening, and the improvement of fruit quality. J. Japan Soc. Horticulrutal Sci.76 :1-12
Karmawan L. U., Suhandono S., Dwivany F. M.
2009. Isolation of MA-ACS gene family and expression study of MA-ACS1 gene in preserving RNA expression in situ with an effective RNA extraction method makes it possible to study gene expression in any banana fruit tissue. J. Fruits 64:127-137 Liu X., S. Shiomi A., Nakatsuka Y., Kubo R.,
Nakamura, dan Inaba A. 1999. Characterization of Ethylene Biosynthesis Associated with Ripening in Banana Fruit. J. Plant Physio., 121: 1257–1265
Livak KJ, Schmittgen TD. 2001. Analysis of plants. Annual Review of Plant Physiology
35: 155-189.
Diperiksa dan disetujui oleh: Pembimbing: