Studi Genetik Ukuran Biji Padi dan Ketahanan
Kedelai terhadap Virus Kerdil
Tiur S. Silitonga, Asadi, dan Hadis Siregar Balai Penelitian Bioteknologi dan Sumberdaya Genetik Pertanian
ABSTRAK
Studi ini dilakukan dari bulan April sampai Desember 2001 di rumah kaca Balitbio dan Instalasi Penelitian Sukamandi. Sebagai bahan studi ukuran biji pada padi, telah dibuat persilangan antara tetua berbiji besar, sedang, dan kecil, serta telah diperoleh biji F1 dan F2. Dari sepuluh kombinasi persilangan kedelai bersama resiprokalnya antara tetua tahan dan peka juga telah diperoleh biji F1 dan F2. Seluruh biji tetua, F1 dan F2 telah ditanam untuk mempelajari sifat ukuran biji pada padi dan ketahanan kedelai terhadap virus kerdil. Hasil studi menunjukkan bahwa tidak terdapat pengaruh tetua betina (maternal effect) ter-hadap pewarisan sifat tahan SSV pada kedelai. Ketahanan SSV pada genotipe Mlg 2521 dikendalikan oleh satu gen mayor yang bersifat dominan, namun pada genotipe B3570 dan Taichung sekurang-kurangnya dikendalikan oleh dua gen mayor dengan interaksi duplikat resesif epistasis.
Kata kunci: Pewarisan, SSV, padi, kedelai
ABSTRACT
This study was conducted from April to December 2001 at green house, RIFCB Bogor and Sukamandi Research Station. The crossing between big grain with medium and small size were conducted to study the mode of inheritance in grain size. Ten cross combinations along with their reciprocals were also made in soybean which resistant, intermediate and susceptible to SSV. All the seeds which consist of parents, F1 and F2 seeds were already grown to study the inheritance of the grain size in rice and resistance to SSV in soybean. There was no maternal effect found in the soybean crossing between resistant and susceptible. Resistance to SSV in Mlg 2521 was caused by a dominant gene, while in B3570 and Taichung were caused by 2 major genes with interaction of duplicate recessive efistatic.
Key words: Inheritance, SSV, rice, soybean
PENDAHULUAN
Kebutuhan padi (beras) masih terus meningkat dari tahun ke tahun. Untuk memenuhi kebutuhan tersebut dilakukan berbagai upaya untuk menghasilkan varietas unggul berdaya hasil tinggi dan berumur genjah (pendek). Beberapa varie-tas yang saat ini telah dihasilkan mempunyai malai panjang dan gabah agak besar tetapi jumlah biji per malai kurang atau kehampaan masih tinggi. Jumlah biji per malai berhubungan erat dengan ukuran biji. Biji yang besar biasanya jumlahnya sedikit per malai, sebaliknya biji yang kecil pada umumnya jumlahnya banyak per malai. Potensi hasil tanaman padi ditentukan oleh komponen hasil yang terdiri dari jumlah malai per m2, panjang malai, banyaknya gabah isi per malai, dan bobot 1000 butir gabah. Komponen hasil ini memiliki hubungan yang sangat erat satu de-ngan lainnya
(Matsushima, 1980). Untuk itu, usaha perakitan varietas unggul perlu memperhatikan sifat komponen hasil karena sifat ini ditentukan oleh jenis varietas (Vergara, 1970).
Bank gen tanaman pangan yang terdiri dari sumber daya genetik padi dan palawija mengandung sumber genetik yang berguna bagi penelitian pemuliaan dan bioteknologi. Program pemuliaan dan bioteknologi diselaraskan dengan kebu-tuhan konsumen dan kondisi atau masalah yang terdapat di lapang. Penelitian da-sar seperti studi genetik dengan memanfaatkan sumber gen yang tersedia di bank gen sangat diperlukan untuk mendapatkan informasi tentang model pewarisan sifat-sifat tertentu. Informasi ini akan membantu para pemulia di dalam melakukan seleksi ke arah yang lebih efektif dan efisien.
Perbaikan terhadap komponen hasil padi seperti ukuran biji (bobot 1000 biji) melalui persilangan sangat mendukung perakitan varietas padi yang berdaya hasil tinggi. Pengetahuan di dalam pemilihan tetua untuk persilangan sangat diperlukan. Penggabungan sifat antara tetua berbiji kecil dengan tetua berbiji besar melalui persilangan diharapkan akan memperbaiki ukuran biji di samping dapat memper-tahankan sifat lain seperti jumlah biji per malai banyak dan bobot 1000 butir berat.
Produksi kedelai dalam negeri saat ini masih belum mencukupi kebutuhan konsumsi nasional, sehingga masih diperlukan impor yang jumlahnya terus me-ningkat dari tahun ke tahun (Sumarno, 1999). Faktor biotik yakni serangan penyakit virus merupakan kendala peningkatan produksi (Sumarno et al., 1983). Di antara beberapa penyakit virus kedelai yang telah dilaporkan, virus kerdil kedelai atau soybean stunt virus (SSV) merupakan penyakit yang dominan di Indonesia. Penye-barannya cukup luas, ditemukan di setiap sentra produksi kedelai di Sumatera, Jawa, dan Nusa Tenggara Barat (Martoatmodjo, 1992; Nakano et al., 1998). Serang-an virus ini dapat menurunkan produksi kedelai sampai 50% (Honda et al., 1988). Oleh sebab itu, untuk mengetahui permasalahan tersebut diperlukan penelitian mengenai genetika untuk mengetahui model pewarisan sifat gen dari bentuk/ukur-an biji pada padi dan ketahanan terhadap SSV pada kedelai dan mengetahui jumlah gen yang mengendalikan sifat-sifat biji tersebut.
Jumlah gen pengendali dan cara pewarisan sifat dari persilangan padi berbiji besar dan kecil dan persilangan kedelai yang tahan dan peka SSV sangat diperlu-kan dan merupakan informasi yang berguna bagi pemulia di dalam menentukan prosedur pemuliaan untuk menciptakan varietas unggul berbiji besar dan berpo-tensi hasil tinggi, serta varietas unggul kedelai yang tahan SSV.
BAHAN DAN METODE
Bahan yang digunakan pada studi genetik padi adalah tetua berbiji kecil (Sibuyung Pendek dan Mayas Putih), tetua berbiji sedang (IR64 dan IR42), dan tetua berbiji besar (Cibodas dan Cirata). Persilangan tetua berbiji besar dengan berbiji sedang dan kecil serta resiproknya dilakukan di rumah kaca Balitbio pada MK dan MH 2000. Perbanyakan biji F2 dilakukan di rumah kaca Balitbio pada MK 2001 dan evaluasi daya waris dilakukan di KP Balai Penelitian Tanaman Padi, Sukamandi pada MH 2001. Rancangan lingkungan yang digunakan adalah acak kelompok dengan tiga ulangan. Ukuran populasi P1, P2, F1, dan F2 untuk setiap ulangan berturut-turut adalah 20, 20, 20, dan 200 tanaman sehingga untuk setiap kombinasi
persilangan diperlukan sebanyak 60 biji P1, 60 biji P2, 60 biji F1, dan 600 biji F2. Pemupukan dengan takaran 200 kg urea, diberikan tiga tahap, yaitu masing-masing sepertiga pada waktu tanam, kemudian pada umur 4 dan 7 minggu setelah tanam. Pupuk P dan K dengan takaran 100 kg TSP + 100 kg KCl/ha diberikan seluruhnya pada waktu tanam. Setelah biji dipanen, kemudian diukur panjang dan lebar biji, bobot 1000 butir, dan dihitung nisbah genetiknya.
Studi genetik kedelai menggunakan tetua persilangan terdiri dari tiga tetua tahan SSV (B3570, Mlg 2521, dan Taichung), dan dua tetua rentan (Wilis dan Orba) (Tabel 2). Persilangan dilakukan secara diallel tanpa mengikutkan penyerbukan sendiri tetuanya, yaitu berdasarkan rumus P (P-1). Sebagai bahan studi digunakan tetua 1(P1), tetua 2(P2), masing-masing kombinasi persilangan, biji F1 dan F2. Pengaruh induk betina (maternal effect) diketahui dengan melihat reaksi ketahan-an terhadap SSV pada tanaman F1 dan resiprokalnya melalui uji t. Masing-masing biji F1, F2 dengan populasi masing-masing 20 biji tetua, 20 biji F1, dan 200-240 biji F2 ditanam (1 biji/pot). Seminggu setelah tanam seluruh tanaman uji diinokulasi dengan SSV strain Jakarta dan 5 hari setelah inokulasi setiap daun tanaman uji di-ambil untuk dideteksi dengan metode Dot-ELISA. Skoring tingkat ketahanan ber-dasarkan tampilan warna setiap sampel pada permukaan membran dari hasil analisis Dot-ELISA (skor 0-3) menurut Nakano (1998) (diskusi pribadi).
Nisbah genetik (genetic ratio) diketahui berdasarkan perbandingan jumlah tanaman berbiji besar dan kecil pada padi dan yang tahan dan rentan pada kede-lai. Model pewarisan dan banyaknya gen pengendali sifat-sifat ukuran biji padi dan ketahanan terhadap SSV diduga berdasarkan prosedur yang dikemukakan oleh Weber dan Moorthy (1952), yaitu dengan menggambarkan sebaran frekuensi ta-naman F2 berdasarkan skala yang telah ditetapkan. Kesesuaian nisbah genetik di-ketahui melalui uji goodness of fit dengan Chi square (Gomez dan Gomez, 1984). Dari uji kesuaian nisbah genetik ini akan dapat diduga jumlah dan sifat gen pengendali ketahanan penyakit SSV dan ukuran biji padi (Griffing, 1950).
Untuk mengetahui derajat dominasi gen ketahanan dilakukan analisis nisbah potensi (potence ratio) menurut Petr dan Frey (1966).
1. 0,0 < Hp < 0,25 = linear aditif (no dominance)
2. 0,26 < Hp < 0,75 = dominan sebagian (partial dominance) 3. 0,76 < Hp < 1,25 = dominan sempurna (complete dominance) 4. Hp > 1,25 = dominan lebih (over dominance)
Nilai heritabilitas dalam arti luas dihitung dari sifat ketahanan diduga dengan menggunakan rumus berikut (Weber dan Moorthy, 1952):
dimana, H2 = heritabilitas; s2F2 = ragam F2; s2E = ragam lingkungan: s2E = Vs2P1.s2P2.s2F1; s2P1 = ragam P1; s2P2 = ragam P2; dan s2F1 = ragam F1.
Nilai heritabilitas tersebut dikelompokkan sebagai berikut: H2 >0,50 = tinggi, 0,20 < H2 < 0,50 = sedang, H2 < 0,20 = rendah. 100% x F2 s E s F2 s H 2 2 2 2=
HASIL DAN PEMBAHASAN
Produktivitas varietas unggul ditentukan oleh sifat morfologi dan agronomi yang dimiliki. Batang pendek dan kokoh mempunyai anakan produktif sedang sampai banyak, daun tegak dan sempit, malai panjang, banyaknya gabah isi per malai banyak, bobot 1000 butir berat, dan kehampaan rendah serta tahan terhadap hama dan penyakit. Di samping itu, pengaruh lingkungan sangat besar terhadap pertumbuhan dan daya hasil tanaman padi. Hal lain yang dapat mempengaruhi pertumbuhan adalah pemupukan. Pupuk nitrogen dapat meningkatkan perkem-bangan pertumbuhan pada fase vegetatif dan generatif (Vergara, 1970). Hal ini akan mendukung terbentuknya komponen hasil yang baik. Ketidakseimbangan di antara komponen hasil akan sangat mempengaruhi potensi hasil yang diperoleh.
Beberapa penelitian menunjukkan bahwa jumlah malai per rumpun tidak berpengaruh terhadap hasil (Silitonga, 1989; Sutaryo et al., 1988). Selanjutnya di-katakan bahwa persentase kehampaan merupakan penduga yang efektif terhadap hasil. Banyaknya butir isi per malai berhubungan nyata dengan hasil, demikian pula bobot 1000 butir, namun tidak berpengaruh banyak.
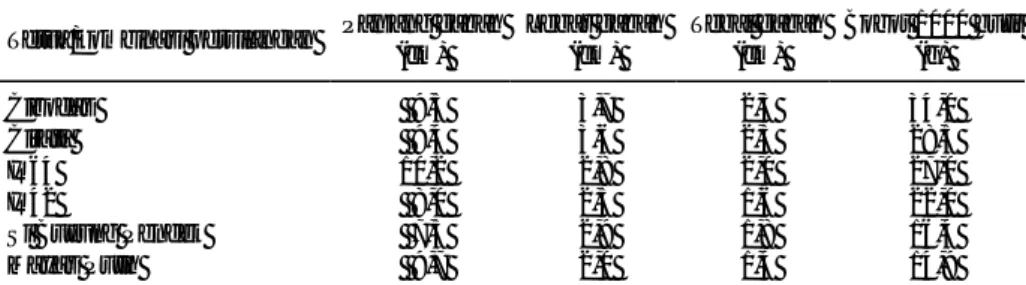
Persilangan varietas berbiji besar yang mempunyai bobot 1000 butir 34 g telah dilakukan dengan varietas yang berbobot 1000 butir sedang dan kecil, dan telah menghasilkan biji F1 (Tabel 1). Jumlah F1 yang dihasilkan bervariasi dari 2-382 biji. Sebagian biji F1 ditanam kembali untuk memperoleh biji F2 dan sebagian untuk dilakukan silang balik (Back cross).
Pada MH 2001 seluruh hasil persilangan beserta tetua dievaluasi secara simultan dan dihitung nilai pewarisannya. Hasil penelitian padi menunjukkan bah-wa ukuran gabah F1 hasil persilangan padi biji besar dengan biji kecil seperti Cirata/Sibuyung mempunyai panjang, lebar, dan tebal yang berada di antara kedua tetuanya, sedangkan pada F2 terlihat bahwa biji lebih panjang dari tetua yang ter-panjang, sementara lebar dan tebal gabah mempunyai rata-rata ukuran di antara kedua tetuanya (Tabel 2).
Jika dibandingkan persilangan Cirata/Sibuyung dengan resiprokalnya (Sibuyung/Cirata) terlihat perbedaan panjang gabah. Hal ini menunjukkan terdapat pengaruh tetua betina dalam persilangan.
Pada persilangan antarpadi berbiji besar seperti Cibodas terlihat bahwa biji F2 Cirata/Cibodas sebagian besar (73,4%) berukuran lebih panjang atau sama dengan kedua tetuanya dan sebanyak 26,6% lebih pendek ukurannya.
Tabel 1. Karakteristik tetua yang digunakan dalam persilangan padi, rumah kaca RPI, MK 2001
Tetua/kombinasi persilangan Panjang gabah (cm) Lebar gabah (cm) Tebal gabah (cm) Bobot 1000 butir (g) Cibodas 9,5 3,7 2,3 34,0 Cirata 9,4 3,6 2,3 28,5 IR64 10,2 2,8 2,0 27,0 IR42 8,0 2,5 1,6 22,0 Si Buyung Pendek 7,5 2,9 1,8 16,4 Mayas Putih 9,7 2,0 1,4 14,9 Ratio P/L gabah: >3 = ramping; 2,1-3 = sedang; 1,1-2 = lonjong; < 1,1 = bulat
Sebaliknya dengan lebar biji terlihat bahwa 26,2% mempunyai lebar sama atau lebih lebar dari kedua tetua dan 73,8% mempunyai lebar lebih kecil dari tetuanya. Sama halnya dengan tebal biji hanya 14 rumpun (8,33) mempunyai tebal sama atau lebih besar dengan kedua tetuanya, dan 91,67% lebih ramping (tipis) dari kedua tetuanya. Dari pengamatan ini diketahui bahwa jika benih bertambah panjang, maka lebar dan tebal biji berkurang.
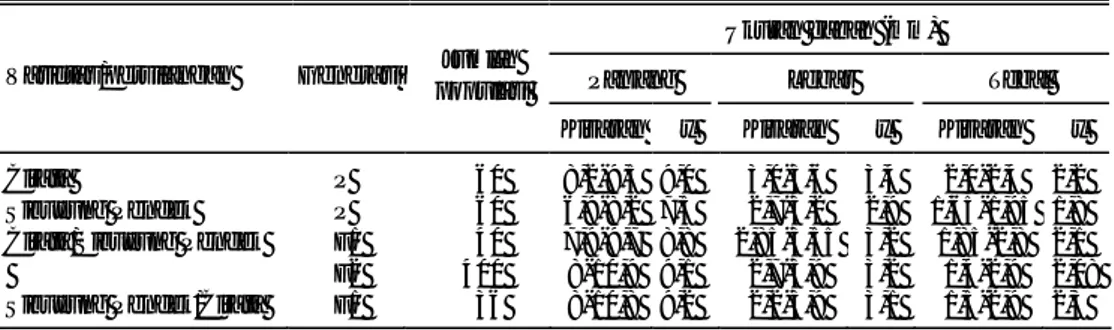
Tetua kedelai yang tahan dan peka terhadap SSV telah disilangkan dan menghasilkan biji F1. Sekitar 20 biji F1 masing-masing dari 20 kombinasi persilang-an (asal lima tetua) telah ditpersilang-anam di kurung kawat Balitbio pada bulpersilang-an November 2000. Penelitian ini dilanjutkan pada MK dan MH 2001. Selama pertanaman F1 ini telah diamati sifat-sifat morfologi dan agronomi, terutama sifat atau markah yang mengindikasikan persilangan berhasil atau tidak, seperti warna bunga (ungu domi-nan terhadap putih), warna hipokotil (ungu domidomi-nan terhadap hijau), warna bulu (coklat dominan terhadap abu). Untuk sifat agronomi seperti umur berbunga, umur masak, tinggi tanaman, banyaknya polong per tanaman dan hasil pada tanaman F1 lebih sulit digunakan sebagai markah (penanda). Akan tetapi bila sifat morfologinya sulit digunakan sebagai penanda berhubung kedua tetuanya memi-liki sifat morfologi yang hampir sama, maka sifat agronominya dapat dijadikan pedoman, terutama bila kedua tetua memiliki perbedaan agronomi yang tegas seperti umur tanaman, tinggi tanaman, banyaknya polong, dan ukuran biji. Dari pertanaman F1 menunjukkan bahwa semua tanaman F1 memiliki warna bunga ungu, hal ini disebabkan kelima tetua yang digunakan berwarna ungu. Tanaman F1 yang memiliki warna bulu abu ditemukan pada persilangan Taichung x B3570 dan B3570 x Taichung, ini disebabkan warna bulu kedua tetua Taichung dan B3570 adalah abu. Karena warna bulu coklat dominan terhadap abu kendati tetua jantan berbulu abu dan betina berbulu coklat, warna bulu tanaman F1 akan tetap coklat, seperti pada persilangan Orba x B3570, Mlg x Taichung, Mlg x B3570 dan Wilis x B3570.
Tabel 2. Ukuran gabah dari persilangan biji besar dengan biji kecil dan sedang Ukuran gabah (mm)
Panjang Lebar Tebal Varietas/persilangan Generasi Jumlah
populasi
Kisaran x Kisaran x Kisaran x Cirata P 60 8,2-9,5 9,0 3,0-3,6 3,4 2,0-2,4 2,2 Sibuyung Pendek P 60 6,9-8,2 7,5 2,7-3,2 2,9 1,65-1,95 1,8 Cirata/Sibuyung Pendek F1 40 7,9-9,7 8,8 2,85-3,55 3,2 1,85-2,8 2,1 F2 400 8-10,9 9,1 2,7-3,9 3,2 1,4-2,9 2,08 Sibuyung Pendek/Cirata F1 36 8-10,8 9,2 2,2-3,9 3,1 1,5-2,9 2,3
Di samping sifat morfologi seperti bentuk dan warna daun, ketebalan daun, sifat agronomi seperti umur berbunga, umur panen, banyaknya polong, dan tinggi tanaman (Tabel 3) dapat juga digunakan untuk mengkonfirmasikan apakah persi-langan jadi atau tidak. Pada persipersi-langan Taichung (umur genjah) x Mlg 2521 (umur dalam) atau resiprokalnya, umur tanaman F1-nya berkisar antara 83-90 hari yakni lebih panjang dari umur tetua genjah (Taichung) dan lebih pendek dari umur tetua dalam (Mlg2521).
Dari penanaman 20 kombinasi persilangan telah diperoleh biji F2 sesuai dengan yang dibutuhkan (200-250 biji/kombinasi). Biji F2 ini bersama biji tetua (P1 dan P2) ditanam sekaligus untuk menguji ketahanan terhadap SSV dan mengeta-hui genetika ketahanannya serta model pewarisannya.
Di dalam tulisan ini hasil yang dibahas adalah kombinasi persilangan antara tiga tetua tahan (Mlg 2521, B3570, dan Taichung) dengan dua tetua rentan (Wilis dan Orba). Kombinasi persilangannya adalah Mlg 2521 x Orba, B3570 x Wilis, B3570 x Orba dan Taichung x Wilis. Gejala infeksi SSV baru diamati secara visual (skor gejala pada tanaman), seleksi secara serologis dengan metode ELISA belum selesai dilakukan karena anti serumnya belum tersedia. Dari keempat persilangan tersebut berdasarkan skor gejala SSV pada tanaman, uji t menunjukkan F1 dan F1 resiprokalnya tidak memperlihatkan perbedaan yang nyata (Tabel 4). Hal ini menunjukkan bahwa tidak Tabel 3. Sifat morfologi dan agronomi tanaman tetua kedelai dan F1 dari 20 kombinasi persilangan,
rumah kaca, Balitbio, 2001
Warna Umur (hari) Persilangan & x %
Bunga Bulu Berbunga Panen
Banyaknya polong/tanaman
Tinggi tanaman (cm)
Taichung x Orba Ungu Coklat 36-43 83-86 9-68 37-81 Taichung x B3570 Ungu Abu 36-47 87-89 13-114 46-78 Taichung x Wilis Ungu Coklat 36-42 82-85 17-70 40-85 Taichung x Mlg 2521 Ungu Coklat 41-51 84-97 7-91 46-120 Orba x Taichung Ungu Coklat 37-44 83-87 14-71 52-70 Orba x Wilis Ungu Coklat 40-46 87-92 41-65 78-98 Orba x B3570 Ungu Coklat 39-51 89-97 31-79 62-82 Orba x Mlg 2521 Ungu Coklat 45-50 85-93 70-81 77-101 B3570 x Taichung Ungu Abu 39-50 83-89 19-100 46-75 B3570 x Orba Ungu Coklat 39-49 89-100 31-87 61-90 B3570 x Wilis Ungu Coklat 42-57 88-100 21-105 38-81 B3570 x Mlg 2521 Ungu Coklat 40-49 90-100 20-154 71-108 Mlg 2521 x Taichung Ungu Coklat 42-50 83-90 16-98 55-100 Mlg 2521 x Orba Ungu Coklat 42-51 87-90 25-108 71-104 Mlg 2521 x Wilis Ungu Coklat 42-47 87-94 37-94 82-109 Mlg 2521 x B3570 Ungu Coklat 42-51 90-100 13-140 37-105 Wilis x Taichung Ungu Coklat 39-45 84-85 16-87 35-71 Wilis & Orba Ungu Coklat 42-47 85-100 22-78 70-98 Wilis x B3570 Ungu Coklat 42-47 88-95 26-99 62-90 Wilis x Mlg 2521 Ungu Coklat 45-46 90-95 33-110 72-122 Taichung Ungu Abu 35 77 23 49,5 Mlg 2521 Ungu Coklat 50 95 80 74,2 B3570 Ungu Abu 45 90 36 65,0 Orba Ungu Coklat 36 80 24 60,0 Wilis Ungu Coklat 37 80 23 88,2
terdapat pengaruh tetua betina dalam pewarisan sifat tahan SSV. Pengaruh tetua betina merupakan faktor lain yang mempengaruhi pewarisan sifat di luar khromosom yakni diturunkan melalui sitoplasma yang banyak mengandung sel telur, tidak seperti sel sperma (serbuk sari) yang tidak mempunyai sitoplasma atau hanya mempunyai inti sel. Soybean stunt virus adalah termasuk kelompok Cucumber Mosaic Virus (CMV). Rustikawati (2000) juga tidak menemukan adanya pengaruh tetua betina terhadap sifat tahan CMV pada tanaman cabai merah.
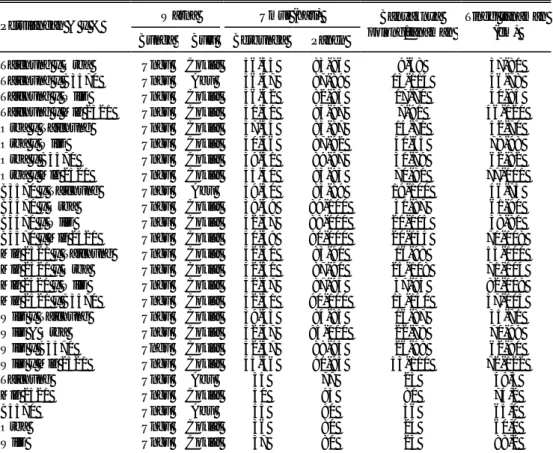
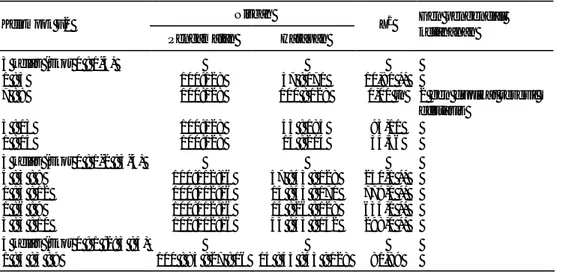
Berdasarkan pengelompokan frekuensi nilai pengamatan skor gejala pada populasi F2, pengelompokan dengan dua kelas (tahan dan rentan), nisbah yang cocok adalah 3 (tahan) : 1 (rentan) pada persilangan Mlg 2521 x Orba (Tabel 5), 7 (tahan) : 9 (rentan), 9 (tahan) : 7 (rentan) pada persilangan B3570 x Wilis, Taichung x Wilis, dan B3570 x Orba (Tabel 6, 7, dan 8). Populasi F2 persilangan Mlg 2521 x Orba juga cocok untuk pengelompokan dengan 3 kelas (12 : 3 : 1) (Tabel 5). Namun jika dilihat dari nilai probabilitasnya, dibandingkan dengan dua kelas, pengelompokan, tiga kelas kurang sesuai karena memiliki nilai probabilitas lebih kecil (0,10-0,005) sementara nilai probabilitas pada pengelompokan dua kelas adalah 0,50-0,30.
Pada persilangan Mlg 2521 x Orba dengan kecocokan nisbah F2 3 (tahan) : 1 (rentan), memberikan informasi bahwa ketahanan genotipe Mlg 2521 dikendalikan oleh satu gen dominan. Hal ini didukung oleh pengamatan F1-nya di mana 82%
Tabel 4. Uji t pada F1 dan F1 resiprokal (F1R) untuk
sifat ketahanan terhadap SSV Rata-rata skor gejala Persilangan F1 F1S Uji t (0,05) Mlg 2521 x Orba 0,18 0,09 0,863 tn B3570 x Wilis 1,17 1,09 0,000 tn B3570 x Orba 0,58 0,75 0,592 tn Taichung x Wilis 3,00 2,40 1,961 tn tn = tidak nyata
Tabel 5. Analisis segregasi populasi F2 persilangan Mlg 2521 x Orba Nisbah Kelompok F2 Pengamatan Harapan X2 Gen pengendali ketahanan 3 kelas (skor 0 : 1-4) 3 : 1 169 : 54 167:56 0,95 tn 1 gen dominan 9 : 7 169 : 54 125:98 35 : 24 ** 13 : 3 169 : 54 181:42 4 : 22 ** 15 : 1 169 : 54 209:14 114,28 ** 3 kelas (skor 0 : 1-2 : 3-4) 9 : 3 : 4 169 : 48 : 6 125 : 42 : 56 60,99 ** 12 : 3 : 1 169 : 48 : 6 167 : 42 : 14 4,45 tn 9 : 6 : 1 169 : 48 : 6 125 : 84 : 14 35,49 ** 10 : 3 : 3 169 : 48 : 6 139 : 42 : 42 38,19 ** 4 kelas (skor 0 : 1 : 3 : 4) 9 : 3 : 3 : 1 169: 33:15:6 125 : 42: 42 : 14 33,34 ** tn = tidak nyata
individu F1 memiliki skor 0 (tahan) dan 18% skor 1 (agak tahan). Persilangan B3570 x Wilis dan Taichung x Wilis dengan kecocokan nisbah F2-nya 7 (tahan) : 9 (rentan), dan B3570 x Orba dengan nisbah F2-nya 9 (tahan) : 7 (rentan) meng-indikasikan bahwa ketahanan genotipe B3570 dan Taichung sama-sama dikendali-kan oleh dua gen dengan interaksi duplikat resesif efistasis. Artinya gen homozigot resesif pada satu lokus bersifat epistatik terhadap gen dominan. Fenotipe yang dihasilkan pada penelitian ini adalah gejala, sedangkan karakter ketahanan bersifat resesif. Pada nisbah 7 (tahan) : 9 (rentan), jika gen tersebut adalah A dan B dengan asumsi sebagai berikut:
A = dominan untuk pemunculan gejala
a = menimbulkan gejala apabila dalam keadaan homozigot B = dominan untuk pemunculan gejala
b = menimbulkan gejala apabila dalam keadaan homozigot Maka kemungkinan genotipe F2-nya adalah
A-B = 9 (muncul gejala) aa-B = 3 (tanpa gejala) A-bb = 3 (tanpa gejala) aabb = 1 (tanpa gejala)
Sehingga kemungkinan genotipe tetua tahan adalah aa-B, A-bb, dan aabb. Tabel 6. Analisis segregasi populasi F2 persilangan B3570 x Wilis
Nisbah Kelompok F2 Pengamatan Harapan X2 (0,05) Gen pengendali ketahanan 3 kelas (skor 0 : 1-4) 1 : 3 105 : 131 5 : 177 47,82 **
7 : 9 105 : 131 103 : 133 0,069 tn 2 gen duplikat resesif efistasis 3 : 13 105 : 131 192 : 44 211,44 ** 1 : 15 103 : 131 29 : 207 227,07 ** 3 kelas (skor 0 : 1-2 : 3-4) 4 : 3 : 9 105 :62 : 69 19 : 49 : 133 189,7 ** 1 : 3: 12 105 : 62 : 69 15 : 44 : 177 612,2 ** 1 : 6 : 9 105 : 62 : 69 15 : 80 : 133 574,8 ** 3 : 3 : 10 105 : 62 : 69 44 : 44 : 148 134,0 ** 4 kelas (skor 0 : 1 : 3 : 4) 1 : 3 : 3 : 9 105 : 12 : 50 : 69 15 : 44 : 44 : 133 594,9 ** tn = tidak nyata
Tabel 7. Analisis segregasi populasi F2 persilangan B3570 x Orba Nisbah
Kelompok F2
Pengamatan Harapan
X2 Gen pengendali ketahanan
3 kelas (skor 0 : 1-4)
3 : 1 131 : 101 174 : 58 42,5 **
9 : 7 131 : 101 130 : 102 0,017 tn 2 gen duplikat resesif efistasis 13 : 3 131 : 101 188 : 44 91,12 ** 15 : 1 131 : 102 218 : 14 575,36 ** 3 kelas (0 : 1-2 : 3-4) 9 : 3 : 4 131 : 74 : 27 130 : 44 : 58 37,03 ** 12 : 3 : 1 131 : 74 : 27 17 : 44 : 14 43,15 ** 9 : 6 : 1 131 : 74 : 27 130 : 88 : 14 14,30 ** 10 : 3 : 3 131 : 74 : 27 144 : 44 : 44 28,2 ** 4 kelas (0 : 1 :2: 3 : 4) 9 : 3 : 3 : 1 131 : 159 : 15 : 27 130 : 44 : 44 : 14 36,3 ** tn = tidak nyata
Tabel 8. Analisis segregasi populasi F2 persilangan Taichung x Wilis Nisbah Kelompok F2 Pengamatan Harapan X2 Gen pengendali ketahanan 3 kelas (skor 0 : 1-4) 1 : 3 100:128 57 : 171 10,81 **
7 : 9 100:128 100 : 128 0,00 tn 2 gen duplikat resesif efistasis 3 : 13 100:128 43 : 185 93,11 1 : 15 100:128 14 : 214 34,56 3 kelas (skor 0 : 1-2 : 3-4) 4 : 3 : 9 100:112:16 57 : 43 : 128 241,1 ** 1 : 3 : 12 100:112:16 14 : 43 : 171 779,1 ** 1 : 6 : 9 100:112:16 14 : 26 : 128 634,1 ** 3 : 3 : 10 100:112:16 43 : 43 : 142 298,0 ** 4 kelas (skor 0 : 1 :2: 3 : 4) 1 : 3 : 3 : 9 100 : 85 : 27 : 16 14 : 43 : 43 : 128 81,89 tn = tidak nyata KESIMPULAN
1. Studi genetik ukuran biji pada padi, tetua berbiji kecil (Sibuyung Pendek dan Mayas Putih) memiliki biji yang jauh lebih kecil dari tetua berbiji besar (Cibodas dan Cirata). Ukuran biji Cibodas 48-128%, jauh lebih besar dibanding-kan dengan Sibuyung dan Mayas Putih, sehingga memenuhi syarat untuk peng-ujian studi genetik pewarisan sifat ukuran biji.
2. Dari uji F1 dan F1 resiprokalnya terdapat pengaruh tetua betina terhadap ukur-an biji padi.
3. Hasil uji F1 dan F1 resiprokalnya tidak memperlihatkan adanya pengaruh tetua betina (maternal effect) terhadap pewarisan sifat tahan SSV pada kedelai.
4. Apabila ketahanan terhadap SSV pada kedelai diukur berdasarkan pengham-batan gejala mosaik, keriput pada daun, dan kerdil pada tanaman maka keta-hanan SSV pada genotipe Mlg 2521 diduga dikendalikan oleh 1 gen mayor yang bersifat dominan, namun pada genotipe B3570 dan Taichung sekurang-kurang-nya dikendalikan oleh 2 gen mayor dengan interaksi duplikat resesif efistasis.
DAFTAR PUSTAKA
Gomez, K.A., and A.A. Gomez. 1984. Statistical procedures for agricultural research. John Wiley and Son. New York, Chishesters, Brisbane, Toronto, Singapore. 680 p.
Griffing. B. 1950. Analysis of quantitative gene action by constant parent regression and related techniques. Genetics 35(303):300-321.
Honda, Y., M. Muhsin, N. Iizuka, and K. Yoshida. 1988. Comparisons among Indonesian isolates and Japanese strains of soybean stunt virus. Reprinted from JARQ Vol. 22, No.1, 1988. Tropical Agriculture Research Center, MAFF, Japan. Martoatmodjo, R. 1992. Viruses of soybean (Glycine max (L.) Merril) in Java and
Lampung. Identification, distribution, and control. UNPAD, Bandung, Indonesia. Matsushima, S. 1980. Rice cultivation for the million diagnosis of rice cultivation on
techniques of yield increase. Japan Scientific Press.
Nakano, M., H. Sawahata, Asadi, Roechan, M. Muhsin, D.M. Arsyad, N. Saleh, and Jumanto. 1998. Distribution of soybean virus diseases in Indonesia. Tentative report of JICA-RIFCB Project on Breeding of Virus Resistant on Soybean.
Petr, F.C. and K.J. Frey. 1966. Genotipe correlation, dominance, and heritability cucumber mosaic virus sympton development in pepper. Hort. Science 19(4):586-587.
Rustikawati. 2000. Identifikasi genotipe tahan dan pewarisan sifat ketahanan terhadap cucumber mosaic virus (CMV) pada cabai merah (Capsicum annuum L.). Program Pasca Sarjana Institut Pertanian Bogor.
Silitonga, T.S. 1989. Analisis koefisien lintasan dari komponen hasil galur-galur padi hibrida. Penelitian Pertanian 9(2):68-70.
Sumarno. 1999. Strategi pengembangan produksi kedelai nasional mendukung gema palagung 2001. Lokakarya Kedelai. Puslitbangtan. Bogor, 16 Maret 1999. Sumarno, D.M. Arsyad, dan I. Manwan. 1983. Teknologi usahatani kedelai.
Lokakarya Pengembangan Kedelai. Pusat Penelitian dan Pengembangan Tanaman Pangan. Bogor, 13 Desember 1983.
Sutaryo, B., B. Suprihatno, dan Z. Harahap. 1988. Analisis komponen lintasan dari komponen hasil perbanyakan benih padi hibrida. Penelitian Pertanian 8(1):46-48.
Vergara, B.S. 1970. Plant growth development. In Rice Production Manual. International Rice Research Institute, Los Banos, Laguna, Phillippines.
Weber, C.R. and D.R. Moorthy. 1952. Heritable and non heritable relationships and variability of content and agronomic characters in the F2 generation on soybean crosses. Agron. J. 55(2):202-209.