MUTAGENESIS DENGAN TRANSPOSON PADA
Xanthomonas
oryzae
pv
. oryzae
PENYEBAB HAWAR DAUN PADI
YANI MULYANI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
BOGOR 2013
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Mutagenesis dengan Transposon pada Xanthomonas oryzae pv. oryzae Penyebab Hawar Daun Padi adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, November 2013 Yani Mulyani NIM G34090039
2
ABSTRAK
YANI MULYANI. Mutagenesis dengan Transposon pada Xanthomonas oryzaepv. oryzae Penyebab Hawar Daun Padi. Dibimbing oleh ARIS TRI WAHYUDI dan ALINA AKHDIYA
Xanthomonas oryzae pv. oryzae (Xoo) merupakan bakteri yang menyebabkan penyakit hawar daun bakteri (HDB) pada padi sehingga dapat menurunkan produktivitasnya. Penggunaan agens biokontrol dianggap sebagai solusi yang efektif. Penelitian ini bertujuan untuk mendapatkan mutan Xanthomonas oryzae pv. oryzae melalui mutagenesis dengan transposon. Introduksi transposon mini-Tn5Km1 yang dibawa oleh E.coli S-17-1(λ pir) ke dalam isolat Xoo STG21 dilakukan dengan teknik konjugasi diparental mating. Mutagenesis dengan transposon dilakukan dengan tiga perbandingan donor dan resipien 1:1, 1:10, dan 10:1. Frekuensi konjugasi rata-rata secara berurutan, yaitu 2.7×10-6; 1.1×10-6; 2.4×10-6. Lima mutan (M93, M99, M107, M112, dan M120) dari 300 mutan yang diujikan tidak menyebabkan reaksi hipersensitif pada tanaman tembakau. Uji yang dilakukan pada tanaman padi IR64 menunjukkan kelima mutan tersebut mampu menurunkan gejala HDB dengan presentase secara berturut-turut 96%; 94%; 94%; 91%; dan 92%.
Kata kunci: HDB, mutagenesis, transposon, Xanthomonas
ABSTRACT
YANI MULYANI. Transposon Mutagenesis of Xanthomonas oryzae pv. oryzae Caused Agent of Bacterial Leaf Blight of Rice. Supervised by ARIS TRI WAHYUDI and ALINA AKHDIYA
Xanthomonas oryzae pv. oryzae (Xoo) causes bacterial leaf blight (BLB) disease of rice that can reduce rice productivity. The use of biocontrol agents is an effective solution to solve this problem. The aim of this study was to obtain mutants of Xanthomonas oryzae pv oryzae through transposon mutagenesis. Transposon mini-Tn5Km1 carried by E.coli S-17-1 (λ pir) was introduced into Xanthomonas oryzae pv oryzae STG21 by conjugation diparental mating. Frequency of tranconjugation obtained from this study was about 2.7×10-6, 1.1×10-6, dan 2.4×10-6 at the ratios of 1:1, 1:10, and 10:1 respectively. Five mutants (M93, M99, M107, M112, and M120) out of 300 mutants tested did not cause a hypersensitive reaction on tobacco plants. However, on rice IR64 plants showed that those mutants could reduce the symptoms of BLB with presentage of 96%, 94%, 94%, 91% and 92% respectively.
3
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biologi
MUTAGENESIS DENGAN TRANSPOSON PADA
Xanthomonas
oryzae
pv
. oryzae
PENYEBAB HAWAR DAUN PADI
YANI MULYANI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
BOGOR 2013
Judul Skripsi : Mutagenesis dengan Transposon pada Xanthomonas oryzae pv. oryzae Penyebab Hawar Daun Padi
Nama : Yani Mulyani NIM : G34090039
Disetujui oleh
Prof Dr Aris Tri Wahyudi, MSi Pembimbing I
Alina Akhdiya, MSi Pembimbing II
Diketahui oleh
Dr Iman Rusmana, MSi Ketua Departemen
2
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas kasih sayang-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Februari sampai Juli 2013 ini ialah agens biokontrol, dengan judul Mutagensesis dengan Transposon pada Xanthomonas oryzae pv. oryzae Penyebab Hawar Daun Bakteri.
Terima kasih penulis ucapkan kepada Bapak Prof Dr Aris Tri Wahyudi dan Ibu Alina Akhdiya MSi selaku pembimbing serta Prof Dr Alex Hartana selaku anggota tim penguji skripsi. Seluruh biaya penelitian ini didanai oleh pembimbing utama, untuk itu saya mengucapkan terima kasih. Terima kasih juga diucapkan pada Bapak Jaka, Ibu Heni, dan Mas Aldian selaku teknisi laboratorium Mikrobologi yang telah banyak membantu. Ucapan terima kasih secara khusus penulis juga sampaikan pada Siti Meliah, SSi MSi yang telah memberikan saran dan bantuan pada penelitian ini.
Semoga karya ilmiah ini bermanfaat.
Bogor, November 2013 Yani Mulyani
3
DAFTAR ISI
DAFTAR TABEL vi DAFTAR GAMBAR vi DAFTAR LAMPIRAN vi DAFTAR TABEL 4 DAFTAR GAMBAR 4 PENDAHULUAN 1 Latar Belakang 1 Tujuan 2BAHAN DAN METODE 2
Waktu dan Tempat 2
Galur Bakteri, Kondisi Pertumbuhan, Media dan Antibiotik 2
Mutagenesis Xoo dengan Transposon 3
Uji Hipersensitivitas 4
Uji Patogenisitas 4
HASIL DAN PEMBAHASAN 4
SIMPULAN 10
DAFTAR PUSTAKA 10
4
DAFTAR TABEL
1 Galur bakteri dan plasmid yang digunakan dalam penelitian 2 2 Karakteristik mutan Xoo hasil mutagenesis dengan transposon 5 3 Frekuensi konjugasi pada mutagenesis dengan transposon pada
berbagai perbandingan konsentrasi sel donor dan resipien 6 4 Hasil reaksi hipersensitivitas yang dilakukan pada daun tembakau
dari mutan hasil mutagenesis dengan transposon 7 5 Perbandingan panjang gejala HDB tipe liar terhadap mutan 8
DAFTAR GAMBAR
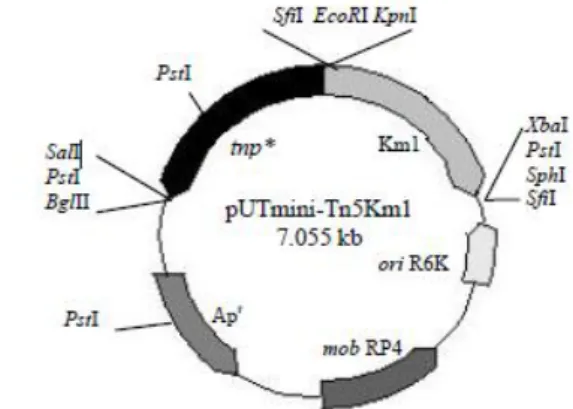
1 Peta plasmid pUTmini-Tn5Km1 (7.055 kb) 3
2 Koloni mutan hasil mutagenesis transposon pada media LA + Rif (50µg/ml) + Km (50 µg/ml ) setelah diinkubasi 24-120 jam pada suhu
ruang 5
3 Respon pada daun tembakau yang dinokulasikan dengan isolat E.coli DH5α (A), akuades steril (B), STG 21(C), mutan hasil transposon hipersensitif positif (D) dan mutan hipersensitif negatif (E) . Bagian
yang disuntik ditunjukan dengan anak panah 7
4 Koloni isolat M93 dan M99 pada media LA+Kan Rif (50 µg mL-1)
setelah diinkubasi 48 jam pada suhu ruang 8
5 Penampilan daun padi yang diinfeksi mutan M93, M99 dan M107, M112, M120, akuades steril (K-) dan STG 21 (K+) 9
PENDAHULUAN
Latar Belakang
Xanthomonas oryzae pv.oryzae (Xoo) merupakan bakteri Gram negatif dengan ciri-ciri morfologi sel berbentuk batang, serta memiliki panjang sekitar 0.7-2.0 µm dan lebar 0.4-0.7 µm. Selnya motil dengan flagela kutub tunggal. Koloni pada media padat yang mengandung glukosa berbentuk bulat, cembung, berlendir, dan berwarna kuning disebabkan pigmen xanthomonadin (Muneer 2007).
Infeksi Xoo pada tanaman padi menimbulkan penyakit hawar daun bakteri (HDB). Menurut Gnamanickam et al. (1999), hawar daun bakteri merupakan penyakit yang menginfeksi secara sistemik dengan gejala berupa bercak berwarna abu-abu putih di sepanjang tulang daun. Gejala ini tampak jelas pada stadia pembentukan anakan, dan kejadian penyakit meningkat seiring dengan pertumbuhan tanaman yang memuncak pada stadia pembungaan. HDB merupakan salah satu penyakit padi yang penting di Indonesia. Serangan HDB di Indonesia pada periode bulan Oktober 2011 – Maret 2012 mencapai 98% (DITJEN TP 2012).
Saat ini, kebutuhan akan pengendalian organisme pengganggu tanaman (OPT) yang ramah lingkungan dan efisien menjadi suatu tantangan. Pengendalian penyakit HDB yang umum dilakukan adalah dengan menggunakan varietas padi tahan penyakit (Herlina dan Silitonga 2011). Namun, penggunaan varietas ini kurang efektif karena Xanthomonas oryzae pv. oryzae mudah membentuk patotipe baru. Sebagaimana yang dilaporkan Hoang et al. (2008) bahwa masing-masing varietas padi hanya tahan terhadap gen tertentu, contohnya varietas IRBB 51 hanya tahan terhadap gen Xa-4 + xa-13, varietas IRBB54 tahan terhadap gen xa-5 + Xa-21, dan varietas IRBB 63 tahan terhadap gen xa-5 +Xa-7 + xa-13. Pengendalian secara kimia yang tidak proporsional dikhawatirkan akan merusak lingkungan dan memerlukan biaya cukup mahal (BB PADI 2012). Oleh karena itu, pengendalian biologis dengan agens biokontrol menjadi solusi alternatif yang menjanjikan untuk mengendalikan HDB.
Beberapa spesies bakteri yang umum hidup pada tanaman berpotensi sebagai agens biokontrol. Namun, bakteri antagonis ini dikhawatirkan akan kehilangan kemampuan sebagai agens biokontrol jika lingkungan berubah. Menurut Rukayadi et al. (2000) bakteri isogenik nonpatogen dapat digunakan sebagai agens biokontrol. Galur bakteri ini dapat dihasilkan melalui mutagenesis dengan transposon. Transposon merupakan elemen genetik yang dapat berpindah dari satu lokasi ke lokasi lain dalam genom. Transposon akan menyisip pada genom secara random.
Mutan Xoo dapat dikonstruksi dengan transposon mini-Tn5Km1 yang dibawa E.coli S-17-1(λ pir). Mutan Xoo tersebut dilaporkan tidak menginduksi reaksi hipersensitivitas dan dapat mengurangi gejala HDB sebesar 80% (Meliah 2010). Oleh karena itu, mutan isogenik yang nonpatogen berpotensi sebagai agens biokontrol. Mutan yang isogenik diperkirakan dapat mereduksi populasi tipe liar Xanthomonas oryzae pv. oryzae dalam relung yang sama dengan cara
2
berkompetisi. Berdasarkan pada penelitian Lindow (1987) bahwa dua strain bakteri berbeda yang terdapat pada permukaan daun yang sama mempunyai kebutuhan nutrisi dan habitat yang sama.
Tujuan
Tujuan penelitian ini adalah untuk mendapatkan mutan Xanthomonas oryzae pv.oryzae melalui mutagenesis dengan transposon.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilakukan pada bulan Februari-Juli 2013 di Laboratorium Mikrobiologi dan rumah kaca mikrobiologi Departemen Biologi IPB.
Galur Bakteri, Kondisi Pertumbuhan, Media dan Antibiotik
Plasmid dan bakteri yang digunakan dalam penelitian ini dicantumkan pada Tabel 1. Xanthomonas oryzae pv. oryzae ditumbuhkan pada media Xanthomonas Agar (XA: CaCO3 30 g/L, glukosa 10 g/L, yeast extract 5 g/L, dan agar 15 g/L ) pada suhu ruang, Escherichia coli S-17 (λ pir) dan DH5α ditumbuhkan pada media Luria Agar (LA: tryptone 10 g/L, NaCl 10 g/L, yeast extract 5 g/L, dan agar 15 g/L), dan Luria Bertani Broth (LB) pada suhu 37˚ C. Antibiotik yang digunakan terdiri dari Rifampisin (Rif), Ampisilin (Amp) dan Kanamisin (Km) (Sigma) dengan konsentrasi yang digunakan sebesar 50 µg/mL. Tanaman tembakau dan benih padi IR64 berturut-turut digunakan untuk uji hipersensitivitas dan patogenitas. Alat yang digunakan adalah syringe, cawan, Laminar Air Flow Cabinet (LAFC) dan alat laboratorium umumnya.
Tabel 1 Galur bakteri dan plasmid yang digunakan dalam penelitian
Galur bakteri atau plasmid Karakteristik Sumber atau referensi Galur Xanthomonas oryzae pv. oryzae
STG 21 Hrp+ Meliah (2010)
MSTG 21 Rifr, Hrp+ Meliah (2010)
M93 Rifr, Kmr, Hr- Penelitian ini
M99 Rifr, Kmr, Hr- Penelitian ini
M107 Rifr, Kmr, Hr- Penelitian ini
M112 Rifr, Kmr, Hr- Penelitian ini
M120 Rifr, Kmr, Hr- Penelitian ini
Galur Escherichia coli
S-17 (λ pir) Ampr, Kmr Herrero et al. 1990
DH5α Hrp- Koleksi Lab
Plasmid Mikrobiologi IPB
pUTmini-Tn5Km1 mini-Tn5Km1:
Ampr, Kanr
Herrero et al. 1990
3
Mutagenesis Xoo dengan Transposon
Mutagenesis transposon dilakukan dengan teknik konjugasi diparental mating antara E.coli S17-1 (λ pir) pembawa plasmid pUTmini-Tn5Km1 (donor) dengan Xanthomonas oryzae pv. oryzae (resipien). Resipien ditumbuhkan pada 50 mL media Xanthomonas Agar (XA) cair + Rif 50 µg mL-1. Kultur diinkubasi pada mesin bergoyang dengan kecepatan 120 rpm pada suhu ruang sampai mencapai populasi sel 108 sel mL-1 (± 20 jam). Donor ditumbuhkan pada 50 mL media LB + Kanamisin (Km) 50 µg mL-1 dalam erlenmeyer 250 mL lalu diinkubasi pada mesin bergoyang dengan kecepatan 120 rpm pada suhu 37°C sampai diperoleh populasi sel yang sama dengan resipien (± 20 jam). Konjugasi dilakukan dengan perbandingan jumlah sel donor : resipien, yaitu 1:1, 10:1, dan 1:10. Jumlah sel yang digunakan pada penelitian ini adalah 108:108, 107:108, dan 108:107 untuk masing-masing perbandingan donor dengan resipien secara berurut. Sebelum dilakukan konjugasi kultur sel dihitung dengan menggunakan hemasitometer. Selanjutnya, untuk mendapatkan sel 107 sel mL-1 dilakukan pengenceran dari kultur sel berjumlah 108 sel mL-1 dengan menggunakan NaCl 0.85 % dan dipatikan kembali jumlah selnya dengan dihitung menggunakan hemasitometer. Kultur sel donor sebanyak 1 mL disentrifugasi pada kecepatan 10.000 rpm selama 5 menit sedangkan resipien sebanyak 1 mL disentrifugasikan pada kecepatan 10.000 rpm selama 10 menit. Pelet sel yang terbentuk dicuci sebanyak 2-3 kali lalu diresuspensi dalam larutan NaCl 0.85 %. Pelet sel resipien kemudian diresuspensikan dalam 40 µL LB. Selanjutnya suspensi sel resipien dipindahkan ke dalam tabung mikro berisi pelet sel donor, kemudian dicampurkan menggunakan pipet mikro secara perlahan. Suspensi campuran dipindahkan ke membran filter milipore steril yang berada di atas media LA. Sebagai kontrol negatif, ke atas membran filter juga diteteskan suspensi sel donor dan resipien secara terpisah. Setelah diinkubasi selama 24 jam pada suhu ruang, masing-masing membran filter diangkat lalu dimasukkan ke dalam 1 mL larutan NaCl 0.85% steril pada tabung mikro, kemudian divorteks untuk melepaskan sel yang terdapat pada membran filter. Selanjutnya sebanyak 100 µL masing-masing suspensi disebar pada media selektif LA + Km 50 µg mL-1 + Rif 50 µg mL-1 lalu diinkubasi selama 24 jam pada suhu ruang. Berikut ini peta plasmid pUTmini-Tn5Km1 yang disisipkan pada genom Xanthomonas oryzae pv. oryzae ( Gambar 1).
4
Uji Hipersensitivitas
Uji hipersensitivitas mutan Xoo pada tanaman tembakau dilakukan berdasarkan metode yang dipaparkan Zou et al.(2006). Suspensi sel mutan (±108 sel/mL) dalam LB + Km 50 µg mL-1 + Rif 50 µg mL-1 disuntikkan ke daun tanaman tembakau menggunakan syringe 1 mL (tanpa jarum). Tipe liar (Wild type [Wt]) Xanthomonas oryzae pv. oryzae digunakan sebagai kontrol positif dan akuades steril serta E.coli DH5α digunakan sebagai kontol negatif. Pengamatan munculnya respon hipersensitif dilakukan setiap hari selama 14 hari.
Uji Patogenisitas
Benih padi IR64 disterilkan menggunakan Natrium-hipokrit 2% selama 10-20 menit, dibilas dengan alkohol 70%, kemudian dibilas dengan air steril dan direndam dengan air steril selama 24 jam. Benih padi ditumbuhkan pada tanah yang sudah disteril hingga berusia dua minggu.Selanjutnya, daun padi digunting untuk dilukai dan dicelupkan ke dalam suspensi bakteri (±108 sel mL-1) selama ±10 detik. Setelah itu, tanaman disungkup dengan plastik bening. Pengamatan gejala penyakit dilakukan pada hari ke-3 dan ke-14 hari setelah inokulasi. Konfirmasi penyebab gejala HDB dilakukan dengan cara reisolasi Xoo STG 21 dan mutan dari daun yang bergejala. Hasil uji patogenisitas yang positif dianggap sah bila hasil reisolasi menunjukkan adanya koloni Xoo.
HASIL DAN PEMBAHASAN
Mutagenesis dengan transposon merupakan salah satu cara untuk mengontruksi mutan yang diharapkan bersifat nonpatogen sebagai upaya pengendalian HDB. Mutan tersebut ditujukan untuk menjadi kompetitor nutrisi pada relung yang sama dengan tipe liarnya sehingga dapat menjadi agens biokontrol yang baik.
Seleksi mutan hasil konjugasi dilakukan pada media LA + Rif (50 µg/ml) + Km (50 µg/ml ). Sifat seleksi media tersebut disebabkan oleh antibiotik rifamfisin dan kanamisin yang terkandung di dalamnya. Xoo STG 21 merupakan produk mutasi spontan pada media yang mengandung rifamfisin sehingga resisten terhadap antibiotik ini (Meliah 2010). Mutagenesis dengan transposon yang dilakukan dalam penelitian ini menghasilkan Xoo yang resisten terhadap rifamfisin dan kanamisin, yang menunjukkan bahwa gen resisten kanamisin yang terdapat pada plasmid pUTmini-Tn5Km1 yang dibawa oleh E.coli S17-1(λ pir) berhasil diinsersikan ke dalam genom Xoo melalui proses konjugasi. Berikut merupakan koloni mutan yang tumbuh pada media selektif dengan perbandingan donor : resipien 1:1, 1:10, dan 10:1 (Gambar 2).
5
Gambar 2 Koloni mutan hasil mutagenesis transposon pada media LA + Rif (50µg/ml) + Km (50 µg/ml ) setelah diinkubasi 24-120 jam pada suhu ruang
Pertumbuhan koloni mutan pada media selektif mulai teramati setelah diinkubasi selama 24 sampai 120 jam. Salah satu sifat mutan yang dihasilkan melalui mutagenesis dengan transposon adalah pertumbuhannya lambat (slow growth) jika dibandingkan tipe liarnya. Seperti pada mutan Xanthomonas campestris pv. campestris yang dihasilkan melalui mutagenesis transposon menunjukkan fenotipe yang bervariasi, diantaranya variasi fenotipe tersebut adalah pertumbuhan yang lambat dan bersifat non mukoid (Shaw et al. 1988). Selain itu, berdasar pada penelitian Ray et al. (2000) karakter lain pada mutan (BXO803) adalah aktivitas xylanase-nya rendah pada fraksi ekstraselular sedangkan pada periplasmik dan fraksi sitoplasmik aktivitasnya tinggi. Secara fenotipe koloni mutan Xoo yang dihasilkan pada penelitian ini sama dengan tipe liarnya kecuali M99 yang koloninya berwarna putih. Mutan M99 diduga mengalami mutasi pada gen penyandi xanthomonadin sehingga tidak berwarna kuning seperti tipe liarnya (Tabel 2).
Tabel 2 Karakteristik mutan Xoo hasil mutagenesis dengan transposon
Isolat mutan Warna koloni Kecepatan pertumbuhan Reaksi hipersensitivitas STG 21 (Wt) kuning ++ + M93 kuning + - M99 putih ++ - M107 kuning + - M112 kuning ++ - M120 kuning + - Keterangan:
pertumbuhan ++ : koloni bakteri teramati setelah inkubasi 24 jam pertumbuhan + : koloni bakteri teramati setelah inkubasi >24 jam
Konjugasi untuk konstruksi mutan umumnya dilakukan dengan perbandingan 1:1 antara donor dan resipien. Sedangkan pada penelitian ini konjugasi dilakukan dengan tiga perbandingan untuk mengetahui perbandingan yang optimum. Konjugasi dengan perbandingan donor dan resipien 1:1 menghasilkan jumlah koloni mutan rata-rata 442 dengan nilai frekuensi konjugasi rata-rata sebesar 2. 6× 10-6. Perbandingan 1:10 menghasilkan jumlah koloni mutan rata-rata 134 dengan nilai frekuensi konjugasi sebesar 1.1×10-6. Perbandingan 10:1 menghasilkan jumlah koloni mutan rata 175 dengan nilai frekuensi rata-rata 2. 4×10-6 (Tabel 3). Hal ini menunjukkan bahwa konjugasi yang dilakukan pada perbandingan antara sel donor dan resipien yang sama menghasilkan jumlah
M 1:1 0, 5 mm M 1:10 M 10:1
6
koloni lebih banyak dan frekuensi konjugasi yang lebih tinggi dibandingkan dengan perbandingan lainnya. Sel bakteri donor dan resipien masing-masing berjumlah kurang lebih 108 sel mL-1.
Seleksi mutan melalui uji hipersensitif pada daun tembakau dilakukan terhadap 300 mutan yang ditentukan secara acak. Kontrol positif yang digunakan adalah Xoo STG 21 yang merupakan tipe liar, sedangkan kontrol negatifnya adalah akuades steril dan E.coli DH5α. Gejala yang ditimbulkan oleh kontrol positif muncul dalam waktu 24 jam setelah inokulasi. Reaksi hipersensitif merupakan kematian sel yang cepat dan terlokalisir. Reaksi ini muncul akibat tanaman terinfeksi dan sel mati untuk menghambat pertumbuhan patogen. Induksi reaksi hipersensitif dan patogenisitas dipengaruhi oleh gen hrp yang umum ditemukan pada bakteri fitopatogen Gram negatif .
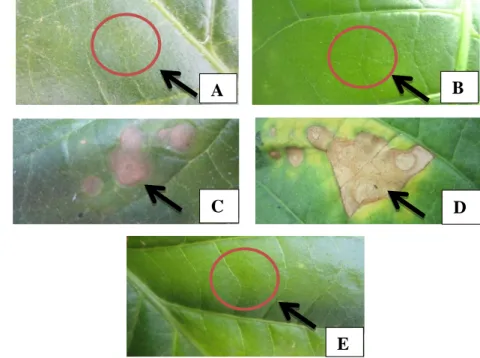
Hasil reaksi hipersensitivitas yang dilakukan pada daun tembakau menghasilkan lima mutan yang kehilangan kemampuannya untuk menimbulkan reaksi hipersensitivitas (Tabel 4). Sebanyak 295 mutan tetap menimbulkan respon hipersensitif yang ditandai dengan adanya bercak berwarna kuning kecoklatan (Gambar 3). Hilangnya kemampuan untuk menimbulkan reaksi hipersensitif juga pernah dilaporkan pada Xanthomonas axonopodis pv. glycines M715 (mutan isogenik dari Xag YR32) oleh Rukayadi et al. (2000). Selain itu, mutan Xag tersebut juga tidak menyebabkan klorosis pada kotiledon kedelai dan tidak menimbulkan gejala pustul pada tanaman kedelai. Pengamatan secara visual terhadap pertumbuhan koloni kelima mutan tersebut pada media agar cawan menunjukkan pertumbuhan yang sama dengan tipe liarnyanya (Gambar 4). Oleh karena itu, hilangnya kemampuan untuk menimbulkan reaksi hipersensitivitas (RH) pada daun tembakau oleh kelima mutan tersebut bukan disebabkan oleh terhambatnya pertumbuhan mutan.
Tabel 3 Frekuensi konjugasi pada mutagenesis dengan transposon pada berbagai perbandingan konsentrasi sel donor dan resipien
Perbandingan konsentrasi (donor:resipien)
Ulangan Frekuensi
konjugasi Jumlah koloni
1:1 I 5. 3×10-6 930 II 2. 2×10-6 261 II 3. 8× 10-7 137 Rata-rata 2. 7× 10-6 442 1:10 I 2. 4×10-6 244 II 0. 9×10-6 147 III 3. 1×10-8 11 Rata-rata 1. 9×10-6 134 10:1 I 6. 7×10-7 7 II 2. 1×10-6 359 III 4. 4×10-6 160 Rata-rata 2. 4×10-6 175
7
Gambar 3 Respon pada daun tembakau yang dinokulasikan dengan isolat E.coli
DH5α (A), akuades steril (B), STG 21(C), mutan hasil transposon hipersensitif positif (D) dan mutan hipersensitif negatif (E) . Bagianyang disuntik ditunjukan dengan anak panah
Tabel 4 Hasil reaksi hipersensitivitas yang dilakukan pada daun tembakau dari mutan hasil mutagenesis dengan transposon
Isolat Hasil uji
Ulangan 1 Ulangan 2 Ulangan 3
STG 21*) + + + E.coli DH5α **) - - - M93 - - - M899 - - - M107 - - - M112 - - - M120 - - -
Keterangan: *)Kontrol positif + menimbulkan RH
**)
Kontrol negatif - tidak menimbulkan RH
Ketiadaan reaksi hipersentif pada daun tembakau setelah diinfeksi mutan M93, M99, M107, M112, dan M120 diduga akibat mutasi yang terjadi menyebabkannya tidak dikenali sebagai fitopatogen oleh tanaman tembakau. Gen-gen hrp adalah gen-gen yang berperan penting dalam interaksi antara bakteri fitopatogen dengan tumbuhan inang. Gen-gen ini terlibat dalam mekanisme timbulnya reaksi hipersensitivitas dan patogenisitas sehingga mutasi pada gen
D A
E C
8
tersebut dapat menyebabkan hilangnya kedua kemampuan itu (Chan dan Goodwin 1999).
Gambar 4 Koloni isolat M93 dan M99 pada media LA+Kan Rif (50 µg mL-1) setelah diinkubasi 48 jam pada suhu ruang
Infeksi Xoo menyebabkan terbentuknya lesio warna kuning kecoklatan (gejala HDB) pada daun padi yang diinokulasi. Lesio tersebut memanjang pada tepian daun atau seluruh helai daun. Panjang lesio bertambah seiring dengan bertambahnya waktu. Uji patogenisitas pada daun padi menunjukkan bahwa mutan M93, M99 dan M107, M112 dan M120 masih dapat menimbulkan gejala HDB walaupun gejalanya jauh lebih ringan dibandingkan dengan gejala yang ditimbulkan oleh Xoo STG21 (Tabel 5). Panjang lesio yang ditimbulkan mutan berkurang hingga 96% dibandingkan dengan lesio yang ditimbulkan oleh tipe liarnya (Gambar 5), yang menunjukkan bahwa mutan-mutan tersebut belum sepenuhnya kehilangan sifat virulensinya. Sejalan dengan penelitian Hu et al. (2007) dari uji virulensi pada IR24 didapatkan panjang rata-rata lesio 2.5-4.0 cm (Wt=23.1 cm), artinya presentase reduksinya sekitar 79.4%. Penelitian Sun et al. (2005), tipe liar Xanthomonas oryzae pv.oryzae PXO99 menyebabkan panjang lesio sampai 20 cm setelah 10-12 hari inokulasi pada padi IR24, sedangkan lebih dari 200 mutan dapat mereduksi virulensi pada padi IR24 dengan panjang lesio kurang dari 2 cm. Persentasi reduksi pada penelitian tersebut sekitar 82%. Defisiensi virulensi pada mutan menunjukkan telah terinsersinya transposon pada tempat yang berbeda pada ORF yang sama.
Tabel 5 Perbandingan panjang gejala HDB tipe liar terhadap mutan Isolat Panjang gejala
(cm) Persentase reduksi (%) STG 21(Wt) 8. 97 - M93 4. 5 96 M99 3. 1 94 M107 3. 07 94 M112 0. 4 91 M120 0.33 91 0, 45 mm 1 mm
9
Gambar 5 Penampilan daun padi yang diinfeksi mutan M93, M99 dan M107, M112, M120, akuades steril (K-) dan STG 21 (K+)
Beberapa faktor yang mempengaruhi virulensi pada Xanthomonas spp. adalah eksopolisakarida (EPS), lipopolisakarida (LPS), dan setidaknya enam sistem sekresi protein (I-VI). EPS menghambat aliran air dalam pembuluh xilem pada tanaman inang (Chan dan Goodwin 1999). LPS berperan dalam interaksi dengan tanaman inang. Sedangkan pada Xoo faktor virulensi yang berhasil diidentifikasi adalah sistem Xps-T2S (lipase/esterase, selobiosidase, selulase, endoglukanase EglXoB) (Hu et al. 2007; Jha et al. 2007 ; Rajeshwari et al. 2005; Ray et al. 2000; Sun et al. 2005; Wang et al. 2007) T3S system (efektor AvrXa7, AvrXa10, PthXo6, PthXo7) (Sugio et al. 2007; Yang et al. 2004; Yang et al. 2000) sistem T5S (adhesin XadA, XadB, YapH).
Kluster gen hrp pada Xoo telah diidentifikasi termasuk 26 gen inklusif dari hpa2 dan hrpF yang berperan besar pada sifat virulensi dan patogenisitas (Lee et al. 2005). Mutasi pada gen hrpF dapat mengurangi virulensi, sedangkan mutasi pada hpa3, hpa4, dan hpaF tidak mempengaruhi virulensi (Sugio et al. 2005). Oleh karena itu, mutan Xoo yang diperoleh dari penelitian ini tidak sepenuhnya kehilangan patogenisitasnya diduga karena penyisipan transposon melalui proses konjugasi ini tidak menyebabkan gen hrpF bermutasi.
M112 K + K - M120 3.0 cm
[
2.5 cm[
]
3.0 cm]
1.5 cm ] 0. 33 cm 11 cm10
SIMPULAN
Mutagenesis transposon pada Xanthomonas oryzae pv oryzae STG 21 dengan perbandingan donor dan resipien 1:1, 1:10, dan 10:1, frekuensi konjugasinya secara berturut-turut, yaitu 2. 7× 10-6 ; 1. 1×10-6 ; dan 2. 4×10-6. Konjugasi donor dan resipien dengan perbandingan 1:1 lebih banyak koloni yang dihasilkan dan frekuensi konjugasi yang lebih tinggi dibandingkan perbandingan lainnya. Dari 300 isolat yang diujikan pada daun tembakau didapatkan lima mutan Xoo yang tidak menginduksi reaksi hipersensitif, yaitu M93, M99, M107, M112, dan M120. Mutan-mutan tersebut dapat mereduksi patogenisitas hingga 96% dibandingkan dengan tipe liar Xoo (STG 21).
DAFTAR PUSTAKA
[BB PADI] Balai Besar Penelitian Tanaman Padi (ID). 2012. Penyakit Hawar Daun Bakteri. http://bbpadi.litbang.deptan.go.id/index.php/in/penyakit-padi-karena-bakteri/204–penyakit-hawar-daun-bakteri-blb-(5 Desember 2012). [DITJEN TP] Direktorat Jendral Tanman Pangan (ID). 2012. Evaluasi Prakiraan
Serangan OPT Utama Tanaman Padi, Jagung, Kedelai MT. 2011/2012 Terhadap Angka Kejadian Selama Bulan Oktober 2011 – Maret 2012.
http://tanamanpangan.deptan.go.id/index.php/ (5 Desember 2012)
Chan JWYF, Goodwin PH. 1999. The molecular genetics of virulence of Xanthomonas campestris. Biotechnol Adv. 17(1):489-508.
Gnamanickam SS, V. Brindha Priyadarisini, N.N.Narayanan, Preeti Vasudevan, dan S. Kavitha. 1999. An overview of bacterial blight disease of rice and strategies for its management. Curr Sci. 77(1):1435-1443.
Herlina L, Silitonga TS. 2011. Seleksi lapang ketahanan beberapa varietas padi terhadap infeksi hawar daun bakteri strain IV dan VIII. Bul Plas Nut. 17(2) :80-87.
Hoang DD, Nghi KO, Nguyen DT, Pham VD, Le CL. 2008. Pathotype profile of Xanthomonas oryzae pv. oryzae isolates from the rice ecosystem in Cuulong River Delta. Omonrice. 16(1):34-40.
Herrero M, de Lorenzo V, Timmis KN. 1990. Transposon vectos containing non-antibiotic resistance selection markers for cloning and stable chromosomal insertion of foreign genes in Gram negative bacteria. J Bacteriol.172(11) 6557-6567.
Hu J, Qian W, He C. 2007. The Xanthomonas oryzae pv. oryzae eglXoB endoglucanase gene is required for virulence to rice. FEMS Microbiol Lett 269(1): 273–279.
Jha G, Rajeshwari R, Sonti RV.2007. Functional interplay between two Xanthomonas oryzae pv. Oryzae secretion systems in modulating virulence on rice. Mol Plant Microbe Interac 20(1):31–40.
Lee BM et al. 2005.The genome sequence of Xanthomonas oryzae pathovar oryzae KACC10331, the bacterial blight pathogen of rice. Nucleic Acids Res. 33(2): 577-586.
11 Lindow SE. 1987. Competitive exclusion of epiphytic fitness. Appl Environ
Microbiol. 53(1):2520-2627.
Meliah S. 2010. Telaah awal dan mutagenesis transposon Xanthomonas oryzae pv.oryzae penyebab hawar daun bakteri pada padi [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Muneer N, Rafi A, Akhtar MA. 2007. Isolation and characterization Xanthomonas oryzae pv. oryzae from North West Frontier Province (NWFP) Pakistan. Sarhad J Agric. 23(3): 743-751.
Rajeshwari R, Jha G, Sonti RV. 2005. Role of an in plantaexpressed xylanase of Xanthomonas oryzae pv. oryzae inpromoting virulence on rice. Mol Plant Microbe Interac. 18(8):830–837.
Ray SK, Rajeshwari R, Sonti RV. 2000. Mutants of Xanthomonas oryzae pv. Oryzae deficient in general secretory pathway are virulence deficient and unable to secrete xylanase. Mol Plant Microbe Interac. 13(4): 394–401. Rukayadi Y, Suwanto A, Tjahjono, Harling R. 2000. Survival and epiphytic
fitness of a nonpathogenic mutant of Xathomonas campestris pv. glycines. Appl Environ Microbiol. 66(3): 1183-1189.
Shaw JJ, Settles LG, Kado CI. 1988. Transposon Tn4431 mutagenesis of Xanthomonas campestris pv. campestris: characterization of a nonpathogenic mutant and cloning of a locus for pathogenicity. Mol Plant Microb Interac. 1(1): 39-45.
Sugio A, Yang B, White FF. 2005. Characterization of the hrpF pathogenicity Peninsula of Xanthomonas oryzae pv. oryzae. Mol Plant Microb Interac.18(6): 546-554
Sun QH, Hu J, Huang GX, Ge C, Fang RX, He CZ. 2005. Type-II secretion pathway structural gene xpsE, xylanase- and 11ellulose secretion and virulence in Xanthomonas oryzae pv.oryzae. Plant Pathol 54(1): 15–21. Wang L, Makino S, Subedee A, Bogdanove AJ. 2007. Novel candidate virulence
factors in rice pathogen Xanthomonas oryzae pv. Oryzicola as revealed by mutational analysis. Appl Environ Microb 73(24): 8023–8027.
Yang B, White FF. 2004. Diverse members of the AvrBs3/PthA family of type III effectors are major virulence determinants in bacterial blight disease of rice. Mol Plant Microbe Interac. 17(11): 1192–2000.
Yang B, Zhu W, Johnson LB, White FF. 2000. The virulence factor AvrXa7 of Xanthomonas oryzae pv. oryzae is a type III secretion pathway-dependent nuclear-localized doublestranded DNA-binding protein. Proc Natl Acad Sci 97(17): 9807–9812.
Zou L et al. 2006. Elucidation ot the hrp clusters of Xanthomonas oryzae pv. oryzicola that control the hypersensitive response in nonhost tobacco and pathogenicity in susceptible host rice. Appl Environ Microbiol. 72(9): 6212-6224.
12
RIWAYAT HIDUP
Penulis dilahirkan di Cianjur pada tanggal 9 November 1990 dari ayah Ajat Sudrajat dan ibu Juariah. Tahun 2009 lulus dari SMAN 1 Sukaresmi (Kabupaten Cianjur) dan pada tahun yang sama penulis lulus seleksi masuk Institut Pertanian Bogor melalui jalur USMI IPB dan diterima di Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam.
Selama mengikuti perkuliahan penulis menjadi asisten praktikum Mikrobiologi Dasar tahun ajaran 2012/2013. Penulis juga aktif mengajar mata pelajaran Sains di lembaga belajar LP3 Diagnostiq. Penulis juga pernah aktif di Kerohanian Islam Asrama sebagai Koordinator Wanita, LDK Al-Hurriyyah sebagai bendahara Divisi Syiar, Serambi Ruhiyah FMIPA (SERUM-G) sebagai bendahara umum, Forum for Scientific Studies (FORCES) sebagai sekretaris divisi HRD dan Kepala Sekolah FORCES. Penulis pernah mengikuti magang di LIPI Cibinong bagian Applied Microbiology pada tahun 2010, studi lapang di Hutan Pendidikan Gunung Walat, dan praktek lapang di LIPI Kebun Raya Cibodas pada tahun 2012.
Tahun 2012 penulis berhasil menulis buku berjudul Belajar Merawat Indonesia: Kepemimpinan Alternatif bersama penerima manfaat Beasiswa Aktivis Nusantara (BAKTI NUSA) lainnya. Sesuai minatnya bergelut dalam bidang tulis menulis, pada tahun 2011 berhasil mendapatkan juara 1 Lomba Inovasi Teknologi Lingkungan di ITS, lolos PKM-AI DIKTI dengan judul artikel “Isolasi dan identifikasi bakteri di sekitar perakaran tanaman legum di Hutan Pendidikan Gunung Walat” pada tahun 2012, Juara 1 LKTI Festival Tanaman IPB pada tahun 2011, Juara 3 Essay YASMINA se-Jawa Barat, Juara 3 Essay Neo-Eksmus pada tahun 2011, delegasi PIMNAS ke-25 di UMY dan Juara 2 Lomba Karya Tulis Ilmiah Al-Quran pada tahun 2013. Berkat beberapa capaian tersebut Republika pernah memuat profilnya di rubrik Uswah pada tahun 2011. Prestasi lain yang diraih penulis antara lain ialah Juara 1 Mahasiswa Berprestasi Departemen Biologi pada tahun 2013.