ISNAINI ET AL.: DAYA GABUNG JAGUNG HIBRIDA TOLERAN KEKERINGAN
Pendugaan Nilai Daya Gabung Jagung Hibrida
Toleran Kekeringan
Muzdalifah Isnaini1, Sriani Sujiprihati2, dan Firdaus Kasim3 1
Balai Penelitian Tanaman Serealia Jl. Ratulangi 274 Maros, Sulawesi Selatan 2Departemen Agronomi dan Hortikultura, Faperta IPB
Dramaga Bogor 3
Pusat Penelitian dan Pengembangan Tanaman Pangan Jl. Merdeka 147 Bogor
ABSTRACT. Estimation of Combining Ability of Hybrid Maize, Tolerant to Drought. The objective of this research was to estimate
General Combining Ability (GCA) and Spesific Combining Ability (SCA) effect. The F1 of 7 x 7 diallel crosses were evaluated for combining ability under normal and drought conditions in Muneng experiment farm East Java, during July to October 2007. The experiment used randomized complete block design with three replications. The mean squares due to genotypes, GCA, SCA were f ound highly significance for grain yield under both conditions, however Anthesis Silking Interval (ASI) and ear number were not significantly difference under normal condition. Based on GCA/SCA variance ratio all traits were controlled by non additive genes. Inbred MR14 was found as the best general combiner for grain yield and ear number under drought condition, while CML 165 for ASI and ear number character. The most promising specific combiners for grain yield and ear number per plant were P1/P4 and P1/P5 under drought condition, and P3/P6, P4/P7 and P6/P7 for ASI character. Hybrids P4/P2 and P7/P4 produced grain yield higher than did Pioneer 21, where drought decreased yield of about 54.5 and 64.3%, less than both of check varieties. Tolerance index of both hybrids was 0.8. Hybrid combinations of P1/P4, P7/P4, P1/P5, P6/P7 and P2/P4 were potential to be developed for high yielding hybrid varieties with drought tolerance.
Keywords: Zea mays, GCA, SCA, drought tolerance
ABSTRAK. Tujuan penelitian ini adalah untuk menduga nilai daya
gabung umum dan daya gabung khusus. Persilangan dialel 7 x 7 dievaluasi daya gabungnya pada kondisi normal dan kekeringan di Kebun Percobaan Muneng Jawa Timur pada bulan Juli-Oktober 2007. Penelitian menggunakan rancangan acak kelompok dengan tiga ulangan. Kuadrat tengah genotipe, daya gabung umum, dan daya gabung khusus berbeda nyata untuk karakter bobot biji per tanaman pada dua kondisi, namun anthesis silking interval (ASI) dan jumlah tongkol per tanaman tidak berbeda nyata pada kondisi normal. Rasio varian daya gabung umum dan daya gabung khusus memperlihatkan semua karakter dikontrol oleh gen nonaditif. Inbred MR 14 merupakan penggabung umum yang baik untuk karakter bobot biji dan jumlah tongkol pada kondisi kekeringan dan CML 165 untuk karakter ASI dan jumlah tongkol. Persilangan dengan daya gabung khusus yang baik untuk karakter bobot biji dan jumlah tongkol terdapat pada P1/P4 dan P1/P5 pada kondisi kekeringan, P3/P6, P4/P7, dan P6/P7 untuk karakter ASI. Hibrida P4/P2 dan P7/P4 mempunyai bobot biji yang lebih tinggi dibanding Pioneer 21 dengan penurunan hasil 54,5% dan 64,3%, lebih rendah dibanding kedua varietas pembanding. Indeks toleransi kedua hibrida tersebut adalah 0,8. Berdasarkan hasil yang diperoleh, kombinasi persilangan P1/P4, P7/P4, P1/P5, P6/P7, dan P2/P4 potensial dalam pembentukan hibrida yang mempunyai toleransi terhadap kekeringan.
Kata kunci: Jagung, DGU, DGK, toleran cekaman kekeringan
L
ahan pertanian di Indonesia didominasi oleh lahankering seluas 144 juta ha yang berpotensimeng-alami cekaman kekeringan, terutama di Papua, Jawa, Sulawesi, Nusa Tenggara, dan Sumatera. Hal ini akan berdampak terhadap penurunan produktivitas tanaman jagung. Semakin terbatas dan ketidakpastian pasokan air karena bersaing dengan sektor industri dan pemukiman. Iklim yang tidak menentu akibat pemanasan global merupakan faktor penyebab tanaman tercekam kekeringan. Cekaman kekeringan merupakan cekaman abiotik tertinggi, mencapai 26%, kemudian diikuti oleh cekaman mineral 20%, cekaman suhu rendah 15%, sedangkan sisanya adalah cekaman biotik (Blum 1986
dalam Kalefetoðlu dan Ekmekci 2005).
Faktor lingkungan merupakan salah satu faktor yang menentukan keberhasilan usahatani. Keragaman ling-kungan pertanaman jagung dapat menimbulkan interaksi antara genotipe dengan lingkungan. Interaksi ini dapat diperkecil dengan mengembangkan varietas potensi hasil tinggi. Sekitar 57% produksi jagung di Indonesia dihasilkan dari pertanaman pada musim hujan (MH), 24% pada musim kemarau I (MK I), dan 19% pada MK II (Kasryno 2002). Permasalahan yang dihadapi di lahan kering adalah musim hujan yang terlalu pendek.
Pemuliaan jagung hibrida umumnya dilaksanakan di lingkungan dengan kondisi optimum sehingga pengembangannya diarahkan pada lahan subur. Oleh karena itu, perlu perakitan varietas hibrida toleran kekeringan yang dapat mengatasi permasalahan budi daya jagung pada lahan kering. Banziger et al. (1997) merekomendasikan karakter yang terkait dengan toleransi terhadap cekaman kekeringan, yaitu bobot biji, jumlah tongkol per tanaman, anthesis silking Interval (ASI), leaf senescence, ukuran tassel, dan penggulungan daun.
Perakitan varietas unggul yang toleran terhadap kekeringan dapat dilakukan melalui persilangan dialel untuk melihat daya gabung, baik daya gabung umum maupun daya gabung khusus (Hallauer and Miranda
1981). Daya gabung yang diperoleh dari suatu per-silangan antarkedua tetua, dapat memberikan informasi tentang kombinasi yang dapat memberikan turunan yang berpotensi hasil tinggi (Silitonga et al. 1993).
Menurut Setiyono dan Subandi (1996), hasil hibrida F1
akan tinggi apabila kedua tetua komponen pembentuk-nya memiliki efek daya gabung umum dan daya gabung khusus yang tinggi.
Penelitian ini bertujuan untuk menduga nilai daya gabung umum dan daya gabung khusus yang diperoleh pada kondisi pengairan normal dan lingkungan ter-cekam kekeringan. Informasi yang diperoleh dapat digunakan untuk mengidentifikasi kombinasi per-silangan yang memiliki nilai daya gabung umum dan daya gabung khusus tinggi untuk karakter toleran kekeringan.
BAHAN DAN METODE
Penelitian terdiri atas dua tahap. Tahap pertama adalah
pembentukan benih F1 dengan metode persilangan
dialel lengkap. Kegiatan dilaksanakan di Kebun Percobaan Cikeumeuh, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian, Bogor. Lokasi penelitian terletak pada ketinggian 240 m dari permukaan laut, jenis tanah Inseptisol dengan 6-8 bulan basah dan tanaman sebelumnya adalah jagung. Penelitian berlangsung dari bulan Maret sampai Juni 2007. Tahap kedua adalah
evaluasi tetua, F1 dan F1 resiprokal, dilaksanakan di
Kebun Percobaan Muneng, Probolinggo, Balai Penelitian Tanaman Kacang-kacangan dan Umbi-umbian pada bulan Juli sampai Oktober 2007.
Tetua yang digunakan dalam penelitian ini adalah empat genotipe jagung toleran kekeringan dan tiga genotipe yang mempunyai potensi hasil tinggi, tetapi peka cekaman kekeringan (Tabel 1). Dua varietas pem-banding yang digunakan adalah Bisi 2 dan Pioneer 21.
Pupuk (urea, KCl, SP36), ridomil, carbofuran 30% dan Gandasil B.
Metode Percobaan
Pada tahap pembentukan benih F1, setiap genotipe
ditanam sebanyak tujuh baris pada petak sepanjang 5 m. Persilangan dilakukan antartanaman dari masing-masing kombinasi. Hasil persilangan kemudian dievaluasi pada kegiatan tahap kedua.
Pada tahap evaluasi terhadap 7 tetua, 21 F1 dan 21 F1
resiprokal digunakan rancangan acak kelompok dengan tiga ulangan. Setiap ulangan terdiri atas 49 kombinasi genotipe hasil persilangan, ditanam pada jarak 70 cm x 20 cm. Satu petak percobaan berupa barisan tunggal dengan panjang 5 m.
Perlakuan terdiri atas dua kondisi yakni kondisi normal dan cekaman kekeringan. Pemberian air pada kondisi normal dilakukan tiap dua minggu sekali, sedangkan perlakuan pada kondisi cekaman kekeringan merujuk pada prosedur CIMMYT (Banziger et al. 2000). Pemberian air dihentikan dua minggu sebelum tanaman berbunga.
Pengamatan berdasarkan prosedur CIMMYT untuk seleksi terhadap cekaman kekeringan, meliputi:
1. Umur berbunga (hari)
a. Berbunga jantan (anthesis)
b. Berbunga betina (silking)
c. Anthesis silking interval (ASI) dihitung
berdasar-kan selisih umur berbunga jantan dan betina
2. Jumlah tanaman panen/baris
3. Jumlah tongkol panen/baris
4. Bobot tongkol kupasan (kg)
10.000 100-KA
Hasil (kg/ha) = ——— x ———— x B x 0,80 LP 100-15
KA = Kadar air biji waktu panen
LP = Luas panen (m2)
B = Bobot tongkol kupasan (kg)
0,80 = Rata-rata shelling percentage/rendemen
5. Kadar air panen (%)
6. Bobot biji pipilan (g)
7. Lengas tanah (%)
Analisis Data
Analisis daya gabung lakukan jika respon genotipe berbeda nyata. Analisis daya gabung terdiri atas analisis daya gabung umum dan daya gabung khusus dengan
menggunakan metode I (tetua, F1 dan resiprokalnya)
dari Griffing (1956).
Tabel 1. Tetua, asal, dan kriteria genotipe yang digunakan dalam persilangan dialel.
Tetua genotipe Asal Kriteria
P1 CML 161 CIMMYT Peka kekeringan P2 CML 165 CIMMYT Peka kekeringan P3 MR 4 Balitsereal Peka kekeringan P4 MR 14 Balitsereal Toleran kekeringan P5 DTPY 1 CIMMYT Toleran kekeringan P6 DTPY 2 CIMMYT Toleran kekeringan P7 G18Seq CIMMYT Toleran kekeringan CML: Cimmyt Maize Line, MR: Maros, Seq: Sequia (Kering) DTPY: Drought Tolerance Population Yellow
Perbedaan pengaruh daya gabung umum diuji dengan uji-t dan nilainya dibandingkan dengan nilai beda krisis. Nilai beda krisis digunakan untuk melihat perbedaan efek pengaruh daya gabung umum dari dua galur yang dibandingkan.
Nilai beda krisis dihitung dengan formula Singh dan Chaudhary (1979) sebagai berikut:
BK = S.E. x t (tabulated)
dimana: SE =
a
a = varians beda efek DGU
Toleransi terhadap kekeringan dinilai dengan meng-ukur perbedaan hasil antara kondisi pengairan normal dengan kondisi kekeringan, perbedaan produktivitas rata-rata pada kondisi normal dan kering dengan menggunakan indeks kepekaan terhadap kekeringan (Blum 1980). Indeks toleransi untuk mengukur indeks kepekaan/sensitivitas kekeringan (S) dihitung dengan menggunakan rumus yang dikemukakan oleh Fischer dan Maurer (1978) dalam Banzinger (2000):
1
Xp
/
X
)
Y
/
Yp
1
(
S
S: Indeks toleransi kekeringan
Yp: Rata-rata suatu genotipe yang mengalami kekeringan
Y: Rata-rata suatu genotipe yang tidak mengalami kekeringan
Xp: Rata-rata dari seluruh genotipe yang mengalami kekeringan
X: Rata-rata dari seluruh genotipe yang tidak mengalami kekeringan
Kriteria untuk menentukan tingkat toleransi ke-keringan adalah jika nilai S < 0,5 maka genotipe toleran, jika 0,5 < S < 1,0 maka genotipe agak toleran, dan jika S > 1,0 maka genotipe tersebut peka.
HASIL DAN PEMBAHASAN
Hasil analisis varians (Tabel 2) menunjukkan karakter bobot biji/tanaman pada kondisi pengairan normal dan kekeringan berbeda nyata antar genotipe pada taraf 1%. Hal yang sama ditunjukkan oleh karakter anthesis silking
interval (ASI) dan jumlah tongkol/tanaman pada kondisi
kekeringan, sehingga ketiga karakter tersebut dapat dilanjutkan ke analisis daya gabung.
Hasil analisis daya gabung untuk semua karakter toleransi kekeringan menunjukkan bahwa nilai daya gabung umum berbeda nyata pada taraf 5% dan 1% pada kondisi normal maupun kekeringan. Hal ini berarti terdapat satu atau lebih genotipe atau tetua jagung penggabung yang baik berdasarkan karakter toleransi kekeringan
Demikian juga hasil analisis daya gabung khusus berbeda nyata pada taraf 1% pada semua karakter toleransi kekeringan pada kondisi normal maupun kekeringan. Nilai kuadrat tengah daya gabung khusus yang berbeda nyata mengindikasikan bahwa karakter toleransi kekeringan dikendalikan oleh gen nonaditif. Hal serupa dikemukakan oleh Mahmood et al. (2002) bahwa nilai daya gabung khusus yang tinggi menunjuk-kan adanya pengaruh aksi gen nonaditif yang tinggi pada karakter tersebut.
Pengaruh resiprokal pada semua kondisi untuk karakter bobot biji/tanaman dan ASI tidak berbeda nyata, kecuali pada karakter jumlah tongkol/tanaman pada kondisi kekeringan. Pengaruh resiprokal yang tidak berbeda nyata mengindikasikan tidak adanya pengaruh tetua betina (maternal effect).
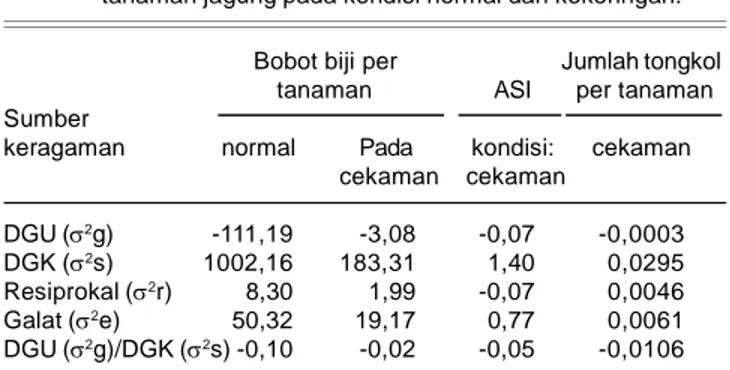
Rasio varian daya gabung umum/daya gabung khusus (Tabel 3) menunjukkan bahwa karakter bobot biji/tanaman, ASI, dan jumlah tongkol/tanaman di-kontrol oleh gen nonaditif (dominan), baik pada kondisi normal maupun kekeringan, di mana varian daya
Tabel 2. Kuadrat tengah genotipe, daya gabung umum, daya gabung khusus, dan resiprokal pada persilangan dialel (7 x 7) genotipe jagung pada kondisi normal dan kekeringan.
Bobot biji/tanaman ASI Jumlah tongkol/tanaman Sumber keragaman db
normal cekaman normal cekaman normal cekaman Genotipe 48 2541.78** 586.70** 0.51ns 5.86** 0.022ns 0.0083**
DGU 6 211.59** 290.34** - 2.16* - 0.006**
DGK 21 1809.21** 340.90** - 3.23** - 0.004**
Resiprokal 21 66.92ns 23.15ns - 0.62ns - 0.001**
Galat 96 50.33 19.18 - 0.77 - 3.10-4
ASI = anthesis silking interval ns= tidak berbeda nyata ** = berbeda nyata pada taraf 1% * = berbeda nyata pada taraf 5%
gabung khusus untuk semua karakter lebih besar dari varian daya gabung umum, sehingga rasio varian daya gabung umum/daya gabung khusus lebih kecil. Hal serupa diperoleh Tabassum et al. (2007) bahwa ratio varian daya gabung umum/daya gabung khusus terhadap bobot biji/tanaman jagung dikontrol oleh gen nonaditif pada kondisi lingkungan normal dan kekeringan.
Analisis Daya Gabung Umum
Genotipe MR 14 mempunyai nilai daya gabung umum tinggi dan positif pada karakter bobot biji/tanaman dan jumlah tongkol/tanaman pada kondisi kekeringan (Tabel 4). Hal ini menunjukkan genotipe MR 14 mempunyai kemampuan yang baik bergabung dengan semua genotipe tetua yang diuji, sehingga genotipe ini dapat digunakan sebagai tetua dalam perakitan varietas jagung berpotensi hasil tinggi.
Genotipe CML 165 dan G18 Seq mempunyai daya gabung umum negatif tinggi untuk karakter ASI pada kondisi kekeringan. Hal ini mengindikasikan bahwa ke-dua genotipe mempunyai penyerbukan dan sinkronisasi bunga jantan dan betina yang lebih baik dibanding genotipe lain, sehingga akan mendukung proses pem-bentukan biji. Nilai daya gabung umum yang tinggi dan positif ditunjukkan oleh genotipe CML 165 dan MR 14 untuk karakter jumlah tongkol/tanaman pada kondisi kekeringan. Genotipe yang memiliki nilai daya gabung umum tinggi pada karakter yang diamati, diarahkan untuk perakitan varietas bersari bebas. Menurut Sprague dan Tatum (1942) dalam Sujiprihati et al. (2001) daya gabung umum relatif lebih penting dibanding daya gabung khusus di antara galur murni yang tidak diseleksi.
Analisis Daya Gabung Khusus
Daya gabung khusus adalah penampilan kombinasi pasangan persilangan tertentu. Bila nilai pasangan persilangan tertentu lebih baik daripada nilai rata-rata
keseluruhan persilangan maka daya gabung khususnya baik (Poehlman 1990). Nilai daya gabung khusus persilangan disajikan pada Tabel 5.
Persilangan P2/P4, P1/P5, P6/P7, P1/P4 mempunyai nilai daya gabung khusus yang tinggi untuk karakter bobot biji pada kondisi kekeringan. Nilai daya gabung khusus merupakan indikator adanya aksi gen dominan dan epistasis yang mengendalikan suatu karakter. Daya
Tabel 4. Nilai daya gabung umum tujuh genotipe tetua jagung pada kondisi normal dan kekeringan.
Bobot biji per Jumlah tongkol tanaman ASI per tanaman Tetua Genotipe
normal Pada kondisi: cekaman cekaman cekaman P1 CML 161 1,12 -5,96 0,69 -0,079 P2 CML 165 4,78 0,48 -0,43 0,074 P3 MR 4 0,75 -4,52 0,19 -0,072 P4 MR 14 4,44 7,79 -0,19 0,067 P5 DTPY 1 -5,23 -1,18 -0,14 -0,012 P6 DTPY 2 -3,90 1,18 0,22 0,027 P7 G18 Seq -1,95 2,20 -0,35 -0,004 SE(gi-gj) 2,68 1,66 0,33 0,029 BK 5,33 3,29 0,66 0,058 Tabel 3. Estimasi varian komponen genetik persilangan dialel 7 x 7
tanaman jagung pada kondisi normal dan kekeringan.
Bobot biji per Jumlah tongkol tanaman ASI per tanaman Sumber
keragaman normal Pada kondisi: cekaman cekaman cekaman DGU (2g) -111,19 -3,08 -0,07 -0,0003 DGK (2s) 1002,16 183,31 1,40 0,0295 Resiprokal (2r) 8,30 1,99 -0,07 0,0046 Galat (2e) 50,32 19,17 0,77 0,0061 DGU (2g)/DGK (2s) -0,10 -0,02 -0,05 -0,0106
Tabel 5. Nilai daya gabung khusus persilangan dialel (7 x 7) genotipe jagung pada kondisi normal dan kekeringan.
Bobot biji per Jumlah tongkol tanaman ASI per tanaman Sumber
keragaman normal Pada kondisi: cekaman cekaman cekaman P1/P2 -17,05 -6,57 -0,57 -0,001 P1/P3 26,01 -0,07 -0,69 -0,035 P1/P4 11,69 11,26 -0,81 0,162 P1/P5 24,92 13,69 -1,03 0,146 P1/P6 13,58 5,20 -0,05 0,091 P1/P7 -8,06 -4,52 -0,15 -0,053 P2/P3 10,08 4,65 -0,74 0,099 P2/P4 17,92 13,95 -0,19 0,059 P2/P5 29,23 8,15 -0,07 -0,017 P2/P6 15,36 7,43 -0,93 0,160 P2/P7 -7,25 -4,33 0,97 0,005 P3/P4 -1,18 -5,48 1,35 0,050 P3/P5 13,82 8,42 -1,03 0,118 P3/P6 11,29 6,73 -1,22 0,071 P3/P7 8,54 5,44 0,02 0,082 P4/P5 13,80 3,58 -0,65 0,109 P4/P6 13,63 2,92 0,00 -0,031 P4/P7 7,15 7,33 -0,10 0,099 P5/P6 -37,30 -14,78 -0,38 -0,072 P5/P7 28,86 7,63 -0,48 0,095 P6/P7 24,72 13,64 -1,00 0,088 SE (Sij-Skl) 6,00 3,70 0,74 0,006 BK 5,34 7,37 1,48 0,132
gabung khusus yang tinggi didukung oleh nilai ragam dominan positif akan memberikan hasil yang lebih baik (Mangoendidjojo 2003).
Persilangan P2/P5 dan P5/P7, karakter bobot biji pada kondisi normal memiliki nilai daya gabung khusus tertinggi, diikuti oleh persilangan P1/P3, P1/P5, P6/P7 dan P2/P4, sehingga persilangan P2/P4, P1/P5, dan P6/P7 mempunyai daya gabung khusus yang baik pada kondisi normal maupun kekeringan. Persilangan P2/P5 dan P5/P7 memiliki kemampuan genotipe bergabung dengan genotipe tersebut hanya pada kondisi normal, sedang-kan pada kondisi kekeringan kemampuan bergabung-nya sangat kecil.
Daya gabung khusus positif berarti genotipe tersebut mempunyai nilai daya gabung khusus yang baik. Semakin tinggi nilai daya gabung khusus maka ke-mampuan genotipe untuk bergabung dengan tetuanya semakin baik juga. Daya gabung khusus negatif artinya genotipe tersebut tidak dapat bergabung dengan baik. Persilangan P1/P2, P1/P7, P2/P7, P3/P4, dan P5/P6 memiliki nilai daya gabung khusus negatif untuk karakter bobot biji per tanaman di kondisi normal maupun kekeringan. Hal ini berarti kombinasi persilangan tersebut tidak dapat bergabung dengan baik.
Pada persilangan galur peka dan toleran P1/P5, P2/P4, dan P1/P4, bobot biji mempunyai pengaruh daya gabung khusus yang tinggi pada kondisi kekeringan. Hal ini mungkin disebabkan oleh gen-gen yang meng-untungkan pada genotipe tahan dapat menutupi gen-gen yang merugikan pada gen-genotipe peka dan mampu bergabung dengan baik. Pada kondisi kekeringan, karakter bobot biji nilai daya gabung khusus tertinggi (13,95) terlihat pada persilangan P2/P4, di mana nilai daya gabung umum tetua P2 termasuk kategori sedang dan P4 tinggi.
Persilangan P1/P5 karakter bobot biji berasal dari persilangan antara dua tetua yang masing-masing memiliki nilai daya gabung umum rendah, tetapi hasil persilangan keduanya memiliki nilai daya gabung khusus tinggi. Hal ini sesuai dengan pendapat Maurya and Singh (1977) bahwa hibrida terbaik kemungkinan besar dapat diperoleh dari persilangan antara dua penggabung umum yang kurang baik. Persilangan ini mengindikasikan adanya aksi gen epistasis yang berperan menutupi sifat jelek tetuanya, sehingga meng-hasilkan bobot biji yang tinggi sebagai karakter toleran kekeringan. Menurut Pradhan et al. (2006), persilangan antara dua tetua dengan nilai daya gabung umum rendah mengindikasikan adanya aksi gen epistasis.
Persilangan antara genotipe toleran dan toleran seperti P6/P7 mempunyai nilai daya gabung khusus yang tinggi pada karakter bobot biji dan ASI. Persilangan ini berasal dari tetua P6 yang bernilai daya gabung umum
rendah dengan P7 yang mempunyai nilai daya gabung umum sedang, yang mengindikasikan adanya interaksi antara gen aditif dan dominan. Menurut Pradhan et al. (2006), kombinasi persilangan antara dua tetua dengan nilai daya gabung umum tinggi dan rendah, atau sedang dan rendah menunjukkan adanya interaksi antara gen aditif dan dominan.
Nilai daya gabung khusus tinggi mengindikasikan kemampuan suatu genotipe untuk bergabung dengan genotipe tertentu, yang dapat dilihat dari tingginya bobot biji per tanaman. Menurut Sprague dan Tatum (1942)
dalam Sujiprihati et al. (2001), nilai daya gabung khusus
lebih penting dibanding daya gabung umum di antara galur murni yang diseleksi untuk karakter hasil.
Persilangan dengan nilai daya gabung khusus tinggi diharapkan berpotensi untuk dijadikan hibrida yang mempunyai toleran kekeringan. Persilangan P6/P7 yang memiliki nilai daya gabung khusus yang tinggi pada karakter bobot biji per tanaman mempunyai nilai daya gabung khusus negatif yang tinggi pada karakter ASI. Persilangan P3/P6 memberikan nilai daya gabung khusus negatif tinggi, diikuti oleh persilangan P1/P5 dan P3/P5. daya gabung khusus negatif untuk karaker ASI, artinya genotipe mempunyai kemampuan bergabung dengan genotipe tersebut lebih baik, sedangkan daya gabung khusus positif mengindikasikan tidak dapat bergabung dengan baik. Daya gabung khusus positif ditunjukkan oleh persilangan P2/P7, P3P4, P3/P7, dan P4/P6.
Karakter jumlah tongkol per tanaman pada kondisi kekeringan memiliki nilai resiprokal nyata. Artinya, ada pengaruh tetua betina terhadap pewarisan karakter toleransi kekeringan. Persilangan P1/P4 dan P2/P6 memiliki nilai daya gabung khusus positif tertinggi, diikuti oleh P1/P5>P3P5>P4P5. Artinya, persilangan ini memiliki kemampuan genotipe untuk bergabung dengan lebih baik dibanding persilangan P1/P2, P1/P3, P1/P7, P2/P5, P4/P6, dan P5/P6 yang bernilai daya gabung khusus negatif.
Persilangan P1/P4 dan P1/P5 memperlihatkan kemampuan bergabung khusus yang baik untuk semua karakter pada kondisi kekeringan. Hal ini berarti kombinasi persilangan tersebut mempunyai toleransi yang lebih baik terhadap cekaman kekeringan.
Bobot biji per tanaman berkisar antara 51,5-156,8 g pada kondisi normal dan 3,8-60,5 g pada kondisi kekeringan dengan penurunan bobot biji 54,5-94,5%. Hibrida P4/P2 dan P7/P4 memiliki bobot biji yang berbeda nyata dengan varietas pembanding Pioneer 21 (Gambar 1).
Varietas pembanding mengalami penurunan bobot biji sebesar 84,0% (Bisi-2) dan 71,3% (Pioneer 21), yang disebabkan oleh kekeringan. Hal ini sesuai dengan yang dikemukakan oleh Poehlman dan Sleeper (1990) bahwa
kekeringan yang terjadi pada fase generatif akan memperlambat waktu panen dan kualitas biji menjadi rendah. Hal tersebut berkaitan dengan menurunnya bobot biji jagung akibat menurunnya laju fotosintesis.
Penurunan bobot biji lebih rendah dengan meng-gunakan tetua yang toleran (P4, P5, P6, dan P7) dibanding tetua peka (P1, P2, P3) sebagai tetua betina. Fenomena ini menggambarkan besarnya peranan tetua betina yang toleran kekeringan dalam pewarisan karakter toleransi. Menurut Sastrosumarjo (2006), tetua betina memberikan sumbangan yang lebih besar kepada zuriatnya dibanding tetua jantan, sehingga karakter-karakter keturunan mewarisi karakter-karakter tetua betinanya. Hasil perhitungan indeks kepekaan terhadap cekaman kekeringan diperoleh beberapa kombinasi persilangan yang menunjukkan toleransinya terhadap kekeringan dengan kriteria agak toleran (0,5<S<1,0) sedangkan kombinasi persilangan yang toleran cekaman kekeringan belum ada (S<0,5). Persilangan P7/P4 dan P4/P2 mengalami penurunan lebih kecil (54,5 dan 64,3%) dibanding kedua varietas pembanding. Indeks toleransi kedua persilangan tersebut adalah 0,8. Artinya, kedua hibrida ini termasuk ke dalam katagori agak toleran terhadap kekeringan. Varietas pembanding pioneer 21 agak toleran, sedangkan Bisi 2 tergolong peka cekaman kekeringan (S>1,0).
Lengas tanah pada penelitian ini pada kondisi cekaman kekeringan dan kondisi normal mempunyai nilai yang berbeda, terutama pada kondisi kekeringan. Air yang tersimpan telah dimanfaatkan tanaman pada
saat pengairan dihentikan menjelang fase pembungaan sampai panen, sehingga lengas tanah pada saat per-lakuan cekaman kekeringan sampai panen mengalami penurunan sampai 9,3% (Tabel 6). Selama penelitian berlangsung tidak terjadi hujan. Keadaan ini meng-indikasikan lokasi penelitian berada dalam keadaan tercekam kekeringan.
KESIMPULAN
1. Genotipe MR 14 memiliki daya gabung umum yang
baik pada karakter bobot biji pada kondisi ke-keringan. Genotipe CML 165 merupakan peng-gabung umum yang baik untuk karakter bobot biji
Tabel 6. Data pengukuran lengas tanah pada kondisi normal dan kekeringan. Muneng, Jawa Timur.
Kondisi normal Cekaman kekeringan Pengairan
Lengas Lengas Lengas Lengas tanah tanah tanah tanah awal (%) akhir (%) awal (%) akhir (%) Pengairan 1 12,5 37,6 11,5 38,7 Pengairan 2 13,1 38,0 11,9 37,5 Pengairan 3 19,3 38,6 - -Pengairan 4 20,2 40,1 - -Pengairan 5 24,1 38,3 - -Pengairan 6 26,5 39,6 - 9,3 Gambar 1. Rata-rata bobot biji per tanaman jagung pada kondisi normal dan kekeringan.
0 20 40 60 80 100 120 140 160 180 P 1 P 1 P 1 P 3 P 1 P 5 P 1 P 7 P 2 P 2 P 2 P 4 P 2 P 6 P 3 P 1 P 3 P 3 P 3 P 5 P 3 P 7 P 4 P 2 P 4 P 4 P 4 P 6 P 5 P 1 P 5 P 3 P 5 P 5 P 5 P 7 P 6 P 2 P 6 P 4 P 6 P 6 P 7 P 1 P 7 P 3 P 7 P 5 P 7 P 7 P io n e e r 2 1 Kondisi normal Kondisi kekeringan 0 20 40 60 80 100 120 140 160 180 0 20 40 60 80 100 120 140 160 180 P 1 P 1 P 1 P 3 P 1 P 5 P 1 P 7 P 2 P 2 P 2 P 4 P 2 P 6 P 3 P 1 P 3 P 3 P 3 P 5 P 3 P 7 P 4 P 2 P 4 P 4 P 4 P 6 P 5 P 1 P 5 P 3 P 5 P 5 P 5 P 7 P 6 P 2 P 6 P 4 P 6 P 6 P 7 P 1 P 7 P 3 P 7 P 5 P 7 P 7 P io n e e r 2 1 P 1 P 1 P 1 P 3 P 1 P 5 P 1 P 7 P 2 P 2 P 2 P 4 P 2 P 6 P 3 P 1 P 3 P 3 P 3 P 5 P 3 P 7 P 4 P 2 P 4 P 4 P 4 P 6 P 5 P 1 P 5 P 3 P 5 P 5 P 5 P 7 P 6 P 2 P 6 P 4 P 6 P 6 P 7 P 1 P 7 P 3 P 7 P 5 P 7 P 7 P io n e e r 2 1 Kondisi normal Kondisi kekeringan
pada kondisi normal dan karakter ASI, jumlah tongkol pada kondisi kekeringan.
2. Semua karakter pada kondisi normal dan
kekeringan dikendalikan oleh gen nonaditif.
3. Pengaruh tetua betina terdapat pada karakter
jumlah tongkol pada kondisi kekeringan.
4. Persilangan P1/P5 (CML 161/DTPY 1), P2P4 (CML
165/MR 14), dan P6/P7 (DTPY 2/G18Seq) memiliki nilai daya gabung khusus tinggi untuk karakter bobot biji pada kondisi normal dan kekeringan. Persilangan P3/P6 (MR 4/DTPY 2), P1/P5 (CML 161/DTPY 1), P6/P7 (DTPY 2/G18Seq), dan P3/P5 (MR 4/DTPY 1) memiliki nilai daya gabung khusus tinggi untuk karakter ASI. Persilangan P2/P6 (CML 165/DTPY 2) dan P1/P4 (CML 161/MR14) memiliki nilai daya gabung khusus yang baik untuk karakter jumlah tongkol/tanaman.
5. Kombinasi persilangan P7/P4 (G18Seq/MR 14), P2/P4
(CML 165/MR 14), dan P1/P4 (CML 161/MR 14) memiliki potensi hasil yang lebih baik dibanding kedua varietas pembanding dan baik digunakan sebagai pembentuk hibrida yang memiliki toleransi terhadap kekeringan.
6. Penurunan bobot biji lebih rendah dengan
meng-gunakan tetua betina yang toleran kekeringan.
DAFTAR PUSTAKA
Banziger, M., GO. Edmeades, and H.R. Lafitte. 1997. Selection for drought tolerance increase maize yields a cross a range of nitrogen levels. Crop Sci. 39:1035-1040.
Banziger, M., S. Mugo, and G.O. Edmeades. 2000. Breeding for drought tolerance in tropical maize-conventional approach and challenges to molecular approaches. In J.M. Ribaut and D. Poland (Eds). Molecular approaches for the genetic improvement of cereals for stable production in water limited enviroments. A Strategic Planning Workshop held at CIMMYT El Batan, Mexico 21-25, June 1999. Mexico p. 69-72.
Blum, A. 1980. Breeding and selection for adaptation to stress; genetic inprovement of draught adaptation. p.450-452. In Adaptation plants to water and high temperature stress. John Willey & sons, Inc. New York.
Griffing, B. 1956. Consept of general and spesific combining ability in relation to diallel crossing system. Aus. Biol. Sci. 9(4):463-493.
Hallauer, A.R. Miranda. 1981. Quantitatif genetic in maize breeding 1st
. Iowa State University Press/Ames. Photosynthesis [ulasan]. Hayati 11:164-169.
Kalefetoglu, E. 2005. The effects of drought on plants and tolerance mechanisms. Journal of Science18(4):723-740.
Kasryno, F. 2002. Perkembangan produksi dan komsumsi jagung dunia selama empat dekade yang lalu dan implikasinya bagi Indonesia. Makalah pada Diskusi Nasional Agribisnis Jagung di Bogor, 24 Juni 2002. Badan Litbang Pertanian. Jakarta. Mahmood, T. 2002. Combining ability studies in rice (Oryza sativa
L.) under salinized soil conditions. Asian J. Plant Sci. 1:88-90. Mangoendidjojo, W. 2003. Dasar-dasar pemuliaan tanaman.
Kanisius. Yogyakarta. 182 p.
Maurya, D.M. dan D.P. Singh. 1977. Combining ability in rice for yield and fitness. Indian J. Agric. Sci. 47(2):65-70.
Poehlman, J.M. and Sleeper. 1990. Breeding field crops, 4th eds. Iowa State University Press/Ames.
Pradhan, S.K., K.B. Lotan, and M. Jitendriya. 2006. Studies on gene action and combining ability analysis in Basmati Rice. Journal Central European Agiculture 7(2):267-272.
Sastrosumarjo. 2006. Sitogenetika tanaman. Bagian Genetika dan Pemuliaan Tanaman. Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor.
Setiyono, R.T. dan Subandi. 1996. Analisis heterosis dan daya gabung jagung. Penelitian Pertanian Tanaman Pangan 15(1):30-34.
Silitonga, T.S. 1993. Evaluasi daya gabung padi bulu dan cere. Penelitian Pertanian. 1:6-11.
Singh, R.K. Chaudhary. 1979. Biometrical methods in quantitatif genetic analysis. Kalyani Publishers.
Sujiprihati, S. 1996. Heterosis combining ability and yield prediction in hybrids from local maize inbred lines. Disertation on Fakulty of Agriculture, University Pertanian Malaysia.
Sujiprihati, S.S., G.B. Saleh, and E.S. Ali. 2001. Combining ability analysis of yield and related characters in single cross hybrids of tropical maize (Zea mays L.). Sabrao Journal of Breeding Genetics 33(2) 111-120.
Tabassum, M. 2007. Combining ability studies in maize under normal and water stress conditions. J. Agric. Res. 45(4).