www.elsevier.com / locate / livprodsci
Impact of biotechnology on (cross)breeding programmes in
qpigs
a ,
*
b a bPeter Visscher
, Ricardo Pong-Wong , Colin Whittemore , Chris Haley
a
University of Edinburgh, West Mains Road, Edinburgh EH9 3JG, Scotland, UK
b
Roslin Institute, Roslin, Midlothian EH25 9 PS, Scotland, UK
Received 28 May 1999; received in revised form 8 November 1999; accepted 15 November 1999
Abstract
Crossbreeding programmes in pigs exploit between breed complementarity of additive genetic effects and heterosis generated by non-additive genetic effects. Within breed, improvement programmes may focus on additive effects and hence the enhancement of complementarity, but non-additive variation is not generally used in within line selection or for mate selection at the multiplier or commercial level. In this paper, we discuss the impact of new biotechnological tools, particularly molecular markers, multiple ovulation and embryo transfer (MOET), and cloning, on structures and methods in crossbreeding. At the between line level, genetic marker information could allow better prediction of heterosis in novel crosses from information on genetic distances. Within the crossbreeding structure, the same technique might be applied at the multiplier and commercial level to exploit specific combining abilities of particular animals. Combining simple MOET and cloning protocols could radically alter the dissemination of crossbreeding benefits and their delivery to the farmer. The combination of MOET, cloning and genomic tools could result in speed genetics programmes, i.e. fast introgression and recurrent selection methods. Thus, the ultimate impact of biotechnology will be increased rates of progress, efficient use of variation, reduced genetic lag, and the removal of one or two tiers in the breeding pyramid. The costs of new technologies are discussed briefly. 2000 Elsevier Science B.V. All rights reserved.
1. Introduction breed (or line) complementarity. A typical practical
crossbreeding programme produces the slaughter Crossbreeding has been widely practised by pig generation as a four-way cross. Such a crossing breeders since the 1960s. The benefits of crossbreed- design utilises individual, maternal and paternal ing programmes in pigs were demonstrated in a heterosis in the slaughter and parental generation,
classical paper by Smith and King (1964). and has been shown to be optimum in terms of
Crossbreeding is performed to utilise heterosis for overall industry efficiency (Smith, 1964; Sellier, reproduction (fitness) and growth traits, and for 1976; Clutter and Brascamp, 1998). In practice there any many other crossing designs employed, from
q backcrosses and three-way crosses to crosses
involv-Based upon a paper presented at the 49th EAAP meeting in
ing more than four lines. In addition to the advantage
Warsaw, August 1998.
*Corresponding author. of utilising heterosis, different breeds or lines may be
kept to breed for different objectives, for example MOET, in vitro production of embryos, molecular specialised sire and dam lines selected for lean tissue genetics, and cloning on breeding programmes. The growth rate and reproduction, respectively. With a likely impact of each of these techniques is discussed suite of different lines the needs of a range of separately, as well as the effect of combining several markets or environments can be addressed by select- techniques.
ing the appropriate combination of lines and crossing structure.
A practical consequence of the need to keep 2. Reproduction techniques separate lines, together with the desire to keep the
best genetics in-house and to disseminate genetic 2.1. Artificial insemination and MOET superiority to the slaughter pig, is the traditional
pyramid structure of the industry. In such a structure Although A.I. in pigs is fairly standardly used in there are a few nucleus herds at the apex of the developed countries, we included a brief description pyramid where genetic progress is achieved and for completion. A.I. and MOET increase the re-many multiplier herds which multiply up either productive rate of the male and female, respectively. purebred or crossbred pigs to satisfy the need for a A.I. in pigs has had a much slower start than in large number of pigs in the commercial tier at the cattle, but is increasing rapidly. In some countries base of the pyramid. From the breeder’s (breeding more than 50% of breeding females are inseminated company or national scheme) perspective, the current artificially. By using A.I., genetically superior nu-structure of the pig industry is not only necessary but cleus boars can be used extensively, particularly at also potentially lucrative, because commercial pig the nucleus and multiplier level. At the nucleus level, farmers need a constant supply of replacement A.I. has made it possible to link several farms to crossbred gilts (and boars) which cannot be home create a large ‘super nucleus’, thereby increasing
bred. genetic gain at nucleus level and a decrease in
There are drawbacks, however, with the current genetic lag between the nucleus herds and the
crossbreeding structures: commercial population. In addition, the introduction
of A.I. is changing the marketing strategy of breed-1. The cost of maintaining several lines and the cost ing companies, from selling live boars to selling of making crosses between lines to produce the semen. However, compared to dairy cattle the impact
crossbred animals of superior boars on the population will be more
2. The genetic lag that is potentially introduced by limited, because of the relatively lower reproductive the need to produce crossbred product for sale to rate of A.I. boars compared to dairy bulls.
commercial producers Compared to cattle, multiple ovulation and
em-3. The difficult in actually selecting for crossbred bryo transfer (MOET) and in vitro fertilisation performance when the nucleus populations tested techniques in pigs have encountered many practical
are purebred. obstacles, including the difficulty with
cryopreserva-tion (of semen and embryos), polyspermy, and non-In future, new biotechnology may help overcome surgical methods of embryo transfer (see e.g., Prather these drawbacks by facilitating a reduction in the and Day, 1998). For these reasons such techniques number of lines that need to be kept, by the removal are not yet routinely used in practical breeding of one or two tiers from the traditional breeding programmes.
pyramid through changing the way in which genetic
superiority is disseminated to the commercial level, 2.2. In vitro production of embryos and by the effective use of crossbreeding information
slaughter house or from live females), non-surgical control over the multiplication process for the breed-embryo transfer techniques, breed-embryo storage and ing companies. Effectively, such a scheme might freezing techniques and cloning. The high female remove the need for a purebred multiplication tier, reproductive rate of the pig raises the question how and reduce the crossbred tier in the industry, thereby and why MOET or IVEP can be beneficial in reducing genetic lag. The use of Chinese pigs is not breeding programmes. MOET by itself is not likely a necessary part of this programme, but used for to be cost-effective if the aim is to increase female illustration because of their superior reproductive selection intensity, and is not a standard practice in performance. It could be argued that using the breeding programmes (except for the hygienic trans- current Chinese genotypes (e.g., the Meishan) is not fer of stock). However, as will be argued below, in desirable because of the small size of the neonates. It combination with other techniques such as marker is possible that a desirable incubator female can be assisted selection and marker assisted introgression bred either from current (white) dam lines (by programmes, these techniques may have a role to increasing litter size and keeping piglet weight play. The production of embryos in vitro is still in its constant) or from Meishan genotypes (increasing infancy stage in pigs, although progress has been litter weight and maintaining litter size).
made recently (Prather and Day, 1998). If large scale
IVEP becomes a reality, what will be the conse- 2.3. Cloning quences for breeding programmes? At the nucleus
level, superior females could have as much influence The prospect of cloning through nuclear transfer on genetic progress as males through an increase in (Wilmut et al., 1997) offers the possibility to reduce selection intensity. For example, if the female selec- the genetic lag between the nucleus, multiplier, and tion intensity could be increased from 1.27 to 1.76, commercial tiers, since genetically superior (per-corresponding to a proportion selected of 25% and formance tested) animals can be cloned and supplied 10%, respectively, and the male selection intensity is to the commercial farmer. The difference with large 2.06 (selected fraction of 5%), the annual genetic scale IVEP is that the clones are genetically identi-gain for growth traits would be increased by, approx- cal, whereas the embryos from a single donor and imately, (2.0611.76) /(2.0611.27)515%. For re- single sire vary in genetic merit. A large scale production traits, where the amount of information nuclear transfer operation requires a constant source on males and females is not equal, the proportional of ‘good’ oocytes, and protocols for in vitro
develop-increase would be larger. ment of oocytes, using for example the abundant
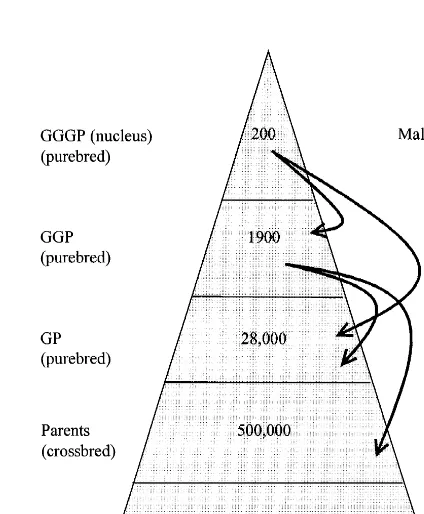
Using a cloning technique on its own is likely to For the traditional breeding scheme, we suppose result in a one-off lifting of the genetic merit at the an integrated breeding scheme with a typical commercial level. Substantial increases in sustain- pyramid structure of parents, grandparents (GP), able rates of improvement in the nucleus herds will great grandparents (GGP) and nucleus animals depend upon the application and factors such as the (GGGP). Grandparents produce on average 18 heritability, accuracy of selection, and rate of in- female parents in their lifetime, assuming a prod-breeding. Reasonable improvements in gains are uctive life of 2 years and a reproductive rate of 18 possible where improvement is currently difficult. It live piglets per year. We further assume that GGP is generally believed that for pig breeding, there is produce on average 15 female GP in their lifetime no or little effect of the application of cloning on the and GGGP produce on average 10 female GGP in rate of progress in the nucleus population. This may their lifetime. There are approximately 190 million be true, but ignores the difference between rate of pigs slaughtered in the EU annually, and for the progress at the nucleus and rate of progress at the purpose of these examples we express the numbers commercial level. At present, the rate of progress at of animals per 10 million slaughter pigs. These the commercial level for slaughter-traits is the aver- simple assumptions suggest that, per 10 million age of the rates of progress for these traits in the sire slaughter pigs, there are approximately 500 000 and dam lines. If cloning and embryo transfer could parents females in the population, whose genes could be applied (cheaply), the slaughter generation con- be ultimately supplied by a small nucleus of 200 tain no genes from the dam line so the rate of sows. See Fig. 1 for the numbers of females in all the progress at the commercial level would be the same tiers of the breeding pyramid. Hence, with the as the rate of progress in the sire line. This would be ‘traditional’ breeding pyramid to breed F females,1
of great economic advantage, because selection only 200 nucleus sows could be required to supply a pressure on reproductive genes in the dam lines are
irrelevant at the commercial level, and therefore an inefficiency of the present breeding schemes. For example, if the rate of progress in the dam lines for slaughter-traits is 90% of the corresponding rate in the sire lines, the rate of progress at the commercial level would be 95% of what could be achieved with cloning. Hence, by completely separating the genes for production and reproduction, a larger rate of progress could be achieved at the commercial level. As with the implementation of large scale IVEP, cloning could change the structure in the industry. We will argue below that the most powerful way to utilise cloning in animal breeding is in combination with molecular technologies.
2.4. Reducing genetic lag
To look at the potential impact of reproduction techniques on the structure of the breeding pyramid, we examine three simple breeding schemes: a tradi-tional breeding scheme, a modern breeding scheme, and a future scheme. These are only example of many possible breeding schemes, and for both the
traditional and the modern scheme, many variations Fig. 1. Traditional breeding pyramid based upon 10 million
market of 10 million slaughter pigs with improved genes. In reality the number of nucleus sows sup-plying such a market is much larger, because of inefficiencies in non-integrated breeding schemes, competition between breeding companies, the need to maintain several lines, and the need to have large enough lines to avoid high levels of inbreeding depression. The time between selection of the nu-cleus sows and the slaughter of commercial animals is approximately 7 years, assuming that nucleus sows have their first litter 6 months after being selected, females in all tiers have four litters with a farrowing interval of 6 months, and an age of first farrowing of 12 months. Under these assumptions, the time when the ‘average’ GGP, GP, P, and slaughter pigs are born is, approximately, 15, 36, 57, and 78 months since the selection of the nucleus sows. If males are passed down the breeding pyramid as live animals, the lag from the male side would be similar to that of the females. However, the use of A.I. to link the purebred tiers (see Fig. 1) reduces the lag on the male side, so that the overall lag can be reduced
Fig. 2. ‘Modern’ breeding pyramid based upon 10 million
slaugh-from 7 years to approximately 5–6 years.
ter pig per year.
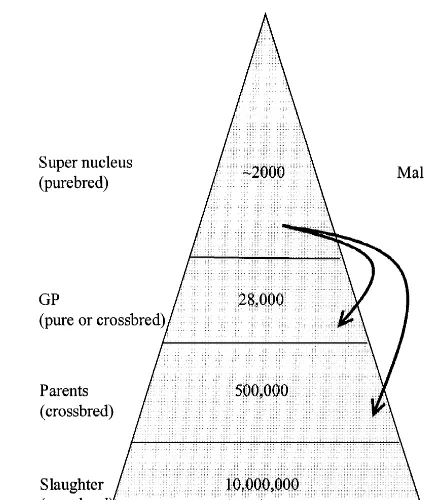
To reduce lag, the above scheme can be amended, by combining the GGGP and GGP into a super nucleus scheme by using A.I. (Fig. 2). Thus the
modern breeding scheme removes one tier of the is more difficult than selection for growth traits, we breeding pyramid. The lag from the females in the assume a larger nucleus for the recipient line breed-breeding pyramid is reduced by approximately 21 ing scheme. Finally, we assume that future recipients
months, to approximately 5 years. females produce about 40 piglets per year. These
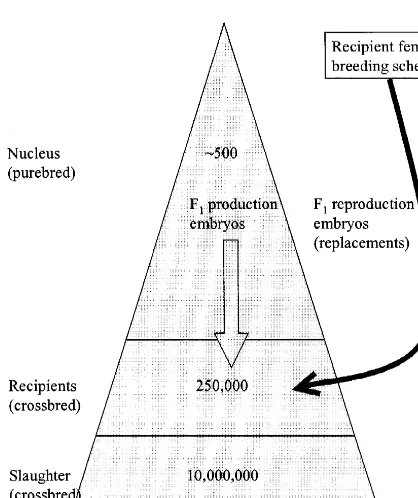
A further tier can be removed if we consider a simple assumptions suggest a large reduction in the future breeding scheme, in which embryos of the number of sows present at any point in time, and a slaughter generation genotype are implanted into large reduction in genetic lag (Fig. 3). Hence, in recipient females which are bred for lifetime re- comparison with the traditional breeding scheme the productive rate (as many live piglets born as pos- sow population has halved, the lag between nucleus sible). We further assume that there is an unlimited and commercial level is virtually zero (or negative, if supply of embryos / clones from donors, that embryos only the top nucleus animals are used to produce are produced at the nucleus level (using selected clones), and only 1000 parents of replacement recipi-nucleus genotypes), and that commercial pig produc- ents are needed. The number of tiers has now been ers buy embryos / clones which are non-surgically reduced to only three.
through generations and relationship between ani-mals can be estimated using identity-by-decent (IBD) measures. The study of associations between these IBD measurements with phenotypic perform-ance has allowed the detection of linkage between marker loci and a QTL, and thereby the identification of genome regions with major effects in quantitative traits. In the pig genome, several regions affecting quantitative traits have already been found, mainly in studies using crosses between divergent breeds (see, e.g. Rothschild, 1998; Walling et al., 1998). Given the trend in the discovery of molecular markers and advances in the statistical techniques to map QTL, the list of known single genes and genome regions affecting quantitative traits is expected to sharply increase.
What is the impact of this powerful technology on pig breeding programmes and, in particular, on the use of crossbreeding? Previously we discussed ways in which genetic markers might be used in pig breeding programmes (Visscher and Haley, 1995, 1998). Markers may be used to combine alleles
Fig. 3. Possible future breeding pyramid based upon 10 million
slaughter pig per year. (variants of a gene or QTL) from different breeds or
lines (using gene introgression programmes or by
3. Molecular techniques creating synthetic lines) or to improve the response
of selection with lines. In addition to these two main One of the aims in molecular genetics is to uses, there is a variety of ways to use genetic identify and locate the genes involved in the expres- markers. They include the control of heterozygosity sion of discrete or quantitative traits. The use of such and inbreeding, control and identification of product information in breeding programmes would provide and stock, and the prediction of dominance / heterosis a measurement of the genetic value of individuals, within and between lines. This list is by no mean without necessarily looking at their phenotype or of exhaustive, and breeding companies will surely find their relatives’ phenotypes. Since DNA can be other ways of utilising molecular marker techniques. sampled from individuals of any sex or age, it also
enables the evaluation of all individuals for traits that 3.1. Marker assisted introgression (MAI) are difficult to measure, such as for sex-specific or
carcass traits. The aim of genome introgression is to introduce
genome region of the donor population, and to speed the QTL (see, e.g. Smith, 1967; Fernando and up the genome recovery of the recipient genotype. Grossman, 1989; Lande and Thompson, 1990; Larzul Currently, there are several examples where MAI is et al., 1997; Henshall and Goddard, 1998; Pong-already possible in commercial pig breeding. For Wong and Woolliams, 1998).
example, the dominant white allele, which gives pigs What would be the impact of MAS in pig breed-the white phenotype, has been identified (Johansson ing? The results from several studies generally et al., 1992) and can be introgressed into non-white associate MAS approaches with a greater short-term lines to create homozygous white recipient lines. genetic gain than traditional methods using only There have been several reports of a QTL allele from performance records. The magnitude of this extra the Meishan on chromosome 7 which increases gain is dependent on the assumed genetic parameters
2
leanness in crosses with European pigs (Rothschild, (e.g. h of the trait, type and size of the QTL effect, 1998). Introgression of this favourable allele into frequency of the QTL, recombination rate between commercial dam lines may be desirable, in particular marker and QTL). Conversely, a reduction in the if it can be combined with introgressing additional long term cumulated gain when using genotype QTL for reproductive performance. Studies assessing information has also been observed, specially with the efficiency of MAI programmes suggest that a mass selection (i.e. the individual’s breeding value is successful introgression programme could result in a estimated using only information their own per-new line which incorporates one or more genome formance and genotypic record) or progeny test regions from a donor line in 5–7 generations of selection programmes (e.g. Gibson, 1994; Larzul et selection (e.g. Hospital et al., 1992; Visscher et al., al., 1997; Pong-Wong and Woolliams, 1998). How-1996). The genetic cost of introgressing the desirable ever, this negative effect in the long term gain is genome region would result in the new line having a limited to MAS schemes that only increase the genetic lag from the recipient population. Assuming accuracy of the estimated breeding values, and also it that MAI is carried out using the genotype of the may be controllable. For instance, under a BLUP QTL itself, this genetic cost was equivalent to one to evaluation framework (which is widely used in pig two generations of selection in the nucleus popula- breeding), the long term loss is substantially
mini-tion (Gama et al., 1992). mised (Villanueva et al., 1998), and it may be
completely eliminated by optimising the relative
3.2. Marker assisted selection (MAS) weight given to both the DNA and the phenotypic
information across generations (Dekkers and Van The purpose of marker assisted selection is to use Arendonk, 1998; Manfredi et al. 1998).
proposed by Fernando and Grossman (1989), the populations is a measure of the number of genera-increased complexity from accounting for an extra tions they have diverged from a presumed common QTL would be equivalent to add two more traits in ancestor, and is usually calculated from differences the multivariate BLUP evaluation. Obviously, some in allele frequencies between the populations at a consideration should be made of the size of the number of loci. Although the correlation between computational problem, but this is unlikely to invali- genetic distance and heterosis is not always large, date the use of MAS with multiple QTL. Perhaps the this method might target specific line combinations most important concern about the Fernando and which are more likely to produce fit, high performing Grossman (1989) approach is that the genetic model progeny. If this technique of combining lines for used to describe the QTL is based on assumptions of crossbreeding works, it should also be possible to normality so does not account for changes in allele use the same principles for mate selection. For frequency and it cannot consider dominant QTL. The example, among a pool of selected males and use of finite locus models, coupled with Monte Carlo females, mate allocation is performed based upon the Markov chain (MCMC) computer methods, may be expected mean heterozygosity of the progeny. How another viable alternative in the near future. Consid- can mate allocation be used in purebred lines? The ering that this model can handle each locus in- benefit of maximising heterozygosity to exploit non dependently, it is also straightforward to extend the additive variation is obvious at the multiplier and model to assume some loci to be linked to a given commercial level. Favourable combinations of alleles genetic marker. An additional advantage of such in the nucleus may result in a greater efficiency, for models is the fact that they easily allow considera- example by increasing the fitness of animals at this tion of QTL with dominance effects. However, level. To achieve a better commercial animal, the before such models can be widely used, further exploitation of non-additive variation in purebred studies are required to fully understand their be- lines should aim to produce the favourable allelic haviour and properties. Early results show that a combinations in the crossbred generation. For in-naive and simplistic implementation of a finite locus stance, if the cross of a pair of purebred lines is model appears sensitive to the number of loci known to show appreciable heterosis, we may be assumed in the model of analysis (Pong-Wong et al., able to associate genetic markers with this effect and 1998). As our current state of knowledge does not within line MAS designed to fix the lines for give us confidence in assumptions about the number alternative alleles would be expected to increase of loci, we require analytical models that are robust performance in the crossbred.
to such assumptions. Selection methods such as recurrent reciprocal
selection (RRS) may be implemented to select the 3.3. Prediction of crossbreeding performance and purebred lines at the nucleus level, with or without
mate selection the use of molecular markers. RSS is a traditional
(1998) concluded that using crossbred information in sequence. Driven by human medicine, these tech-a poultry breeding progrtech-amme wtech-as superior to using nologies are probably not far away, perhaps only 5 pureline information only if the ratio between domi- years or so. Surely the livestock industry will utilise nance variance and total genetic variance was ap- advances in such technologies, and should be pre-proximately 0.3 or higher. Note that the increasing pared to use the information which is generated by spread of A.I. in the pig also means that elite boars them.
from the nucleus level would also have offspring at
the commercial level, so their information would be 3.5. Genetic modification technologies available at the moment when their half sib groups in
the nucleus level are to be selected. Although In this review, we have concentrated on reproduc-technology may already be available to implement tive and molecular technologies which do not alter such a structure, it is likely to need a massive the genome other than through the standard pro-investment in the infrastructure to allow data of cesses of selective breeding. The reason for not guaranteed quality to be retrieved from all levels of discussing transgenic or genetic modification
tech-the breeding structure. nologies in great length is because it is unlikely, in
our opinion, that these technologies will be used for
3.4. Genetic marker technology pig improvement in the next 5–10 years. This
prediction is based both on technological and con-For some of the schemes suggested above, cheap sumer attitude arguments. In our opinion, genetically genomic information, for example in the form of a modified meat is not likely to be acceptable to the dense marker map, would be advantageous. Is this a consumer in the near future. The technological realistic scenario for the livestock industries? We problems and prospects were recently discussed by think it is, for a number of reasons. Firstly, the Pursel (1998), and are related to which genes to current marker technology of choice, i.e. microsatel- target, and how to control the expression of genes lite markers (i.e., markers based on genetic variation which have been introduced.
in the number of repeat sequences at a particular locus), is relative new (since the late 1980s), and new
technologies which could overtake them have been 4. Combining several techniques: speed genetics developed and are being developed. In particular, in
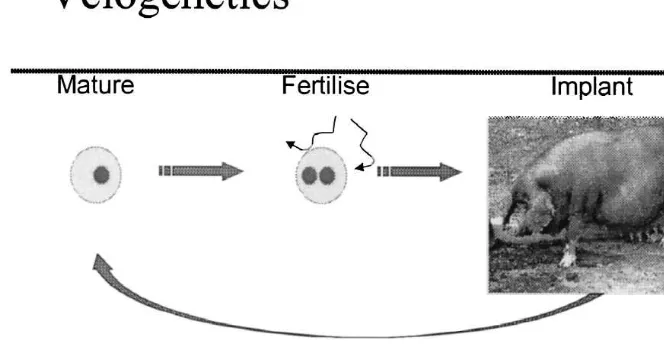
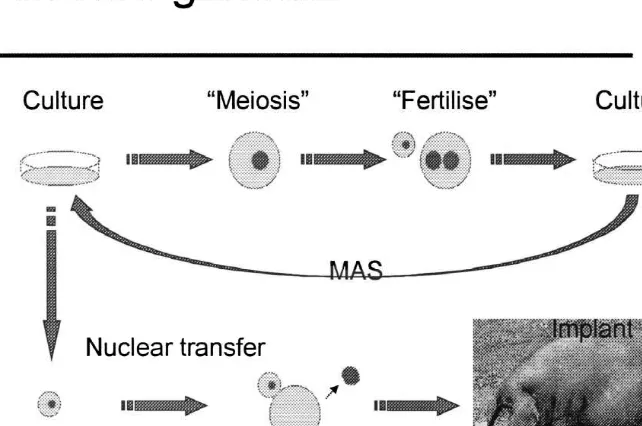
Fig. 4. Velogenetics using IVEP.
Meishan as an example, suitable recipients could be gene introgression programmes for which no addi-from other sources (e.g., improved white lines, tional phenotypic information is required.
crosses between Chinese and European pigs). After If we allow ourselves to imagine that the technolo-the piglets are born, technolo-they can be selected immedi- gy will develop to a stage where cell differentiation ately using marker assisted selection, and immature can be controlled in vitro we can imagine that in oocytes can be harvested from the selected female vitro meiosis followed by fertilisation may become piglets. The duration of one cycle of selection of possible (Fig. 6). Utilising this would allow for very such a rapid selection programme is about 4–5 rapid introgression, or with high-density marker months. The disadvantage is that the breeder has to maps and knowledge of marker–QTL associations, wait until the piglets are born before MAS is applied. more generalised selection objectives (e.g., Haley It is assumed here that the loss of embryos would be and Visscher, 1998). For example, introgression of too great if MAS is attempted on the early embryos. fertility QTLs from exotic breeds could be performed The combination of MAS and such embryo tech- entirely in vitro, thereby reducing the total time of an nologies could be further enhanced by technologies introgression programme to months rather than currently under development, such as nuclear trans- years.
fer (Wilmut et al., 1997). In comparison to ‘stan-dard’ velogenetics, we can now apply marker
as-sisted selection on a diploid cell line, after in vitro 5. Discussion and conclusions production of embryos (Fig. 5). The selected cells
Fig. 5. Nuclear velogenetics using IVEP, MAS, and nuclear transfer.
which does not depend on the sex and age of In considering crossbreeding programmes, we animals, and which usually does not depend on argued that reproductive techniques impact upon the waiting for phenotypic information. Therefore, these efficiency of selection in the nucleus, and on the breeding programmes differ fundamentally from genetic lag between the nucleus and multiplier level. traditional breeding programmes such as reciprocal Hence, the structure of the industry could change by recurrent selection because for those schemes pheno- reducing the number of tiers in the pyramid. The typic information is required at each cycle of selec- components we have discussed could be put together
tion. in a number of ways to form a new commercial
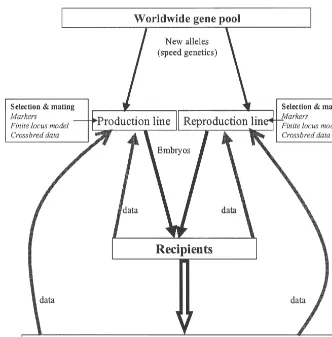
As mentioned previously, crossbreeding is prac- crossbreeding structure. Such a structure might have tised to utilise heterosis and breed / line complemen- the following components (see also Fig. 7)
tarity. In a possible future scenario, where the
parents (recipients) of the slaughter generation pigs • Separate production and reproduction lines with are unrelated to the embryos they carry, the question customised breeding objectives
arises whether there is still a need for crossbreeding. • Within line selection designed to optimise per-For example, a single large nucleus breeding popula- formance in crossbred animals. Selection under-tion could be maintained to produce slaughter gener- taken using marker information and the finite ation clones, utilising mate selection to combine locus model
genotypes. If heterosis is caused by dominance at • Data from nucleus augmented by electronic data many loci, within line selection and mating designs capture from all levels of pyramid
should be able to utilise this dominance variation, • New alleles introgressed into lines using speed
and there is no need to keep separate lines to genetics
maximise response to selection. However, if • Crossbred embryos for production animals (or heterosis is due to overdominance, i.e. heterozygotes alternatively, reproduction animals) produced by are superior to either parent, maximising hetero- A.I. from one production (or reproduction) line zygosity is important, and most easily achieved by into second production (or reproduction) line.
crossing different lines. Matings designed using finite locus model to
The disadvantage of keeping a single line is that optimise pairwise combination of genes
inbreeding will reduce heterozygosity, both at the • Embryos transported to commercial farm and nucleus level and further down the pyramid. This is non-surgical embryo transfer used on host sows. not so in the current schemes, where inbreeding
occurs at the nucleus level, but not at the commercial Thus, both production and reproduction animals level. Hence, the risk of using cloning techniques at are F hybrids and production animals do not contain1
a large scale is that negative effects due to inbreed- alleles from the reproduction line. The difference ing will be expressed at the commercial level, and between the reproduction animals in this example not just in the nucleus herds. This implies either and the reproduction animals in a present-day dam keeping the option of producing a final generation line is that the latter is likely to contain alleles from crossbred animal, or ensuring that the nucleus popu- the production lines, and that the slaughter pigs lation is sufficiently large to maintain levels of receive half of their genes from the dam line, inbreeding at an acceptable level. Genetic markers whereas in the future scheme the slaughter genotype can be used to control the realised levels of inbreed- might be completely unrelated to the reproduction ing (Visscher and Haley, 1995), and new quantita- genotype. We do not claim that this example is in tive genetic techniques have been developed to any way an optimum design — it simply serves to balance genetic gain with homozygosity (e.g., illustrate the sort of structures that may become Grundy et al., 1998). However, unless there is an possible in future. The actual structure that evolves easy way to select against deleterious recessives in will depend upon a number of factors such as the the population, the negative effects of inbreeding practical efficiency of various techniques and when depression probably dictate that a level of they become practical realities. We summarise the crossbreeding will remain to produce parents and likely implementation and impact of various
Fig. 7. Possible future biotechnology driven breeding structure.
Table 1
Predicted impact of biotechnology techniques in terms of the likelihood of effective use in the next 5 years and the estimation of the potential impact on genetic change (scores on scale 1–3, 15low, 35high)
Technique Likelihood of effective use Potential impact
Non-surgical ET 2 1
IVEP 1 2
Cloning 1 2
ET1IVEP1cloning 1 3
Molecular techniques 3 2
Speed genetics 1 3
Genetic modification 0 –
selection in the improvement of quantitative traits. Genetics
because of the pressures in the human genetics
124, 743–756.
community worldwide. Hence, for example, it is
Larzul, C., Manfredi, E., Elsen, J.M., 1997. Potential gain from
realistic to foresee the standard sequencing of in- including major gene information in breeding values estima-dividuals within a decade. It is a challenge both for tion. Genet. Sel. Evol. 29, 161–184.
Manfredi, E., Barbieri, M., Fournet, F., Elsen, J.M., 1998. A
researchers and breeders to effectively utilise such an
dynamic deterministic model to evaluate breeding strategies
enormous amount of genomic information.
under mixed inheritance. Genet. Sel. Evol. 30, 127–148. Prather, R.S., Day, B.N., 1998. Practical considerations for the in
vitro production of pig embryos. Theriogenology 49, 23–32. Pong-Wong, R., Woolliams, J.A., 1998. Response to mass
selec-Acknowledgements tion when an identified major gene is segregating. Genet. Sel.
Evol. 30, 313–337.
Pong-Wong, R., Shaw, F., Woolliams, J.A., 1998. Estimation of
We acknowledge support from BBSRC, MAFF,
dominance variation using a finite-locus model. Proc 6th World
EC, and consortia of UK pig breeding companies.
Congr. Genet Appl. Livest. Prod. (Armidale, Australia) 26, 41–44.
Pursel, V.G., 1998. Modification of production traits. In: Animal Breeding, Technology For the 21st Century, Harwood
Aca-References demic, Amsterdam, pp. 183–200.
Rothschild, M.F., 1998. Identification of quantitative trait loci and Clutter, A.C., Brascamp, E.W., 1998. Genetics of performance interesting candidate genes in the pig: progress and prospects. traits. In: Rothschild, M.F., Ruvinsky, A. (Eds.), The Genetics In: Proc. 6th World Congress on Genetics Applied to Livest of the Pig, CAB International. Prod, pp. 403–409.
Dekkers, J.C.M., Van Arendonk, J.A.M., 1998. Optimum selection Sellier, P., 1976. The basis of crossbreeding in pigs; a review. on identified quantitative trait loci. Genet. Res. 71, 257–275. Livest. Prod. Sci. 3, 203–226.
Gama, L.T., Smith, C., Gibson, J.P., 1992. Transgene effects, Smith, C., 1964. The use of specialised sire and dam lines in introgression strategies and testing schemes in pigs. Anim. selection for meat production. Anim. Prod. 6, 337–344. Prod. 54, 427–440. Smith, C., 1967. Improvement of metric traits through specific Georges, M., Massey, J.M., 1991. Velogenetics, or the synergistic genetic loci. Anim. Prod. 9, 349–358.
use of marker assisted selection and germ-line manipulation. Smith, C., King, J.W.B., 1964. Crossbreeding and litter production Theriogenology 35, 151–159. in British pigs. Anim. Prod. 6, 265.
Gibson, J.P., 1994. Short-term gain at expense of long-term Uimari, P., Gibson, J.P., 1998. The value of crossbreeding response with selection of identified loci. In: Proceedings 5th information in selection of poultry under a dominance model. World Congress on Genetics Applied to Livestock Production, Anim. Sci. 66, 519–528.
pp. 201–204. Villanueva, B., Pong-Wong, R., Grundy, B., Woolliams, J.A., Fernando, R.L., Grossman, M., 1989. Marker-assisted selection 1998. Potential benefit of using identified major gene in BLUP using best linear unbiased prediction. Genet. Sel. Evol. 21, evaluation with truncation and optimal selection. Genet. Sel.
467–477. Evol. 31, 115–133.
Grundy, B., Villanueva, B., Woolliams, J.A., 1998. Dynamic Visscher, P.M., Haley, C.S., 1995. Utilizing genetic markers in pig selection procedures for constrained inbreeding and their breeding programmes. Anim. Breedg. Abstr. 63, 1–8. consequences for pedigree development Genet. Res. 72, 159– Visscher, P.M., Haley, C.S., Thompson, R., 1996. Marker assisted
168. introgression in backcross breeding programs. Genetics 144,
Haley, C.S., Visscher, P.M., 1998. Strategies to utilise marker– 1923–1932.
QTL information. J. Dairy Sci. 81 (Suppl. 2), 85–97. Visscher, P.M., Haley, C.S., 1998. Strategies for marker-assisted Henshall, J.M., Goddard, M.E., 1998. Marker assisted selection in selection in pig breeding programmes. In: Proceedings 6th complex pedigrees using maximum likelihood. In: Proceedings World Congress on Genetics Applied to Livest Prod, pp. 6th World Congress on Genetics Applied to Livestock Pro- 503–510.
duction, pp. 345–348. Walling, G.A., Archibald, A.L., Cattermole, J.A., Downing, A.C., Hospital, F., Chevalet, C., Mulsant, P., 1992. Using markers in Finlayson, H.A., Nicholson, D., Visscher, P.M., Walker, C.A., gene introgression breeding programs. Genetics 132, 1199– Haley, C.S., 1998. Mapping of quantitative trait loci in porcine
1210. chromosome 4. Anim. Genet. 29, 415–424.
Johansson, M., Ellegren, H., Marklund, L., Gustavsson, U., Wei, M., Van der Werf, J.H.J., 1994. Maximizing genetic response Ringmar-Cederberg, E., Andersson, K., Edfors-Lilja, I., Ander- in crossbreds using both purebred and crossbred information. sson, L., 1992. The gene for dominant white color in the pig is Anim. Prod. 59, 401–413.