THERE ARE NO CORRELATION BETWEEN RETINOL BINDING PROTEIN 4 WITH APOB100 IN WOMEN WITH CENTRALOBESITY
IN NORTH SUMATERA Dina Keumala Sari 1
Nutrition Department, Faculty of Medicine, University of Sumatera Utara
ABSTRACT
Background:Prevalence of obesity increased every year especially in developing country and there were multi aspects that affecting it. Adipocyte cell is an active endocrine organ that released many active mediator, one of it was retinol binding protein 4 (RBP4). Through retinol dependent or independent mechanism, there are a relation between RBP4 and trygliceride through ApoB 100, but the mechanism is uncleared.
Objectives: This study aimed to determine was there a correlation between RBP4 and ApoB in women with central obesity.
Methods: This was a cross sectional study including 50 women with central obesity and age 20-50 years old. Examination of RBP4 and ApoB100 were conducted and the correlation was assessed.
Results: Mean aged of the subjects were 30.5±8.9 years old and mean of waist circumference was 99.4±12.3 cm.The mean of RBP4 was 30.4±8.4µg/mL and ApoB 100 was 103.6±16.2 mg/dL. There are no correlation between RBP4 and ApoB100 (p>0.05).
Conclusion: There was no correlation between RBP4 and ApoB100, there should be another parameters that correlated to ApoB100 in central obesity.
Keywords:RBP4, Apo B100, adipocyte, central obesity, women
*corresponding author
INTRODUCTION
Obesity was became an epidemic in developing countries (Sanchez, 2011), the obesity prevalence was increased. Obesity prevalence around the world still high and it predict higher number in obesity more than 250 million people, > 7% from the adult population in the world. Mortality related to obesity had higher mortality that related to metabolic syndrome (Malik, 2004).
Adipocyte was active endocrine that released active mediators for glucose and lipid metabolism. Some of the mediators also had other functions such as balancing blood pressure, inflammation control, and controlling atherosclerosis proccess. One of the active mediator was Retinol Binding Protein 4 (RBP4) (Rabe, 2008). RBP4 adalah protein yang disimpan dihati dan juga didalam jaringan adiposit sebesar kira-kira 20% dari hati dan dilepaskan ke sirkulasi. (Wolf, 2007).
Retinol binding protein 4 is a transport protein for vitamin A, is synthesized mainly by the hepatocytes and secondly by the adipose tissue.1 Plasma RBP4 levels are upregulated in insulin resistant states associated with obesity, while RBP4 also induces insulin resistance.1,2 Furthermore, elevated circulating RBP4 has been associated with the development of cardiovascular disease.3-6 Circulating RBP4 has been shown to be positively correlated with serum triglyceride and low-density lipoprotein-cholesterol (LDL-C) and negatively with high-density lipoprotein-cholesterol (HDL-C).1,7 Among these associations the strongest and the most consistently reported has been the association with TG.1,8 Moreover, serum RBP4 levels have been shown to increase TG in mice.9 These data indicate that RBP4 are possibly associated with TG metabolism.
marker, but it had an association with lipoprotein pro-atherogenic and key enzyme for lipoprotein metabolism(Eynatten (2007).
Retinol binding protein 4 had a regulation with triglyceride through apolipoprotein B (apoB) (Liu, 2015 dan Verges, 2012) and secreted protein apoB through microsomal triglycerida transfer protein(MTP) mechanism (Liu, 2015), apo B needed MTP to transfer lipid and it was a regulator to stabilize and degradation apo B (Hussain, 2003).Based on these previous study, this present study was to investigated whether there was a correlation between RBP4 and ApoB 100 in women with obesity central who lived in North Sumatera, Indonesia.
MATERIALS AND METHODS Study design and Subjects
Subjects In the present study 46 subjects were recruited from November 2015 to January 2016 who lived in Kota Medan, North Suamtera, Indonesia.. Inclusion criteria were: man or women within the range of 20-50 years old, had waist circumference more that 90 cm (man) and 80 cm (women). Exclusion criteria were: pregnancy and breast feeding, any endocrine disorder or metabolic disease other than obesity or type 2 diabetes mellitus (T2DM).
All observations were carried out in the subject’s home or workplace, subjects were included in the study after completing an interview. All subjects were also asked to complete an informed consent approved by the Health Research Ethics Committee of the Faculty of Medicine, Sumatera Utara University (No. 120/KOMET/FK USU/2015).
Anthropometric and biochemical measurements:
immunosorbent assay (ELISA) kit (ALPCO DIAGNOSTICS). Measurement of ApoB 100: serum apolipoprotein ApoB levels were measured with a Behring Holding GmbH analyzer (Liederbach, Germany).
Statistical Analysis
Numerical variables are expressed as means ± standard deviations, while categorical variables are expressed as the percentage values of proportion. Chi-squared tests were used to examine differences between the two groups (p<0.05). Unpaired t-test was used to compare numerical data (p<0.05). This study used SPSS (version 11.5; SPSS Inc, Chicago, IL) for data analysis.
RESULTS AND DISCUSSIONS
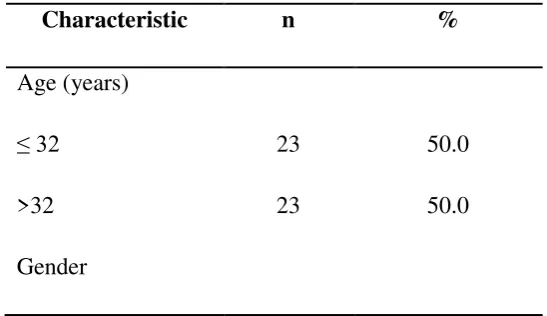
Age is related to changes in body composition. After the age of 20-30 years, Fat Free Mass (FFM) progressively decreases, where Fat Mass increases. FFM (especially muscle tissue) decreases by 40% starting from 20-70 years (Baumgartner, 1995 and Gallagher, 1997). An increase in the muscle fat mass that occurs with increasing age can increase energy intake and decrease energy expenditure, where the relationship between energy intake and energy expenditure is an important factor of body fat tissue (Halflfrisch, 1990). Increase in age is also associated with the division between body fat and FFM. With increasing age, there is a large increase in intra-abdominal fat than in subcutaneous or total body fat and there is a large decrease in peripheral FFM than in central FFM due to skeletal muscle loss (Beaufrere, 2000), (Table 1).
Table 1. Characteristic of subjects
Characteristic n %
Age (years)
≤ 32 23 50.0
>32 23 50.0
Male 20 43.5
Female 26 56.5
Table 2. Characteristic’s subject based on age Characteristic Mean Median Standard
deviation
Minimum maximum
Age (year) 30.52 32 8.96 20 50
Table 3. Mean subjects based on abdominal circumference Characteristic Mean Median Standar
d deviation
Minimum maximum
Abd
circumference (cm)
99.48 99.50 12.32 81.00 134.00
Table 4.Mean subjects based on RBP4
Characteristic Mean Median Standard deviation
Minimum maximum
RBP4 (μg/mL) 30.49 30.03 8.4 16.95 59.08
Tabel 5.Mean subjects based onapo B100
Characteristic Mean Median Standard deviation
Minimum maximum
Table 6. Mean subjects based ontriglyceride
Characteristic Mean Median Standard deviation
Minimum maximum
Triglyceride (mg/dL) 132.52 115.00 57.34 55.00 287.00
Tabel 7. Subjects distribution based on triglyceride
Triglyceride N %
< 150 mg/dL 31 67,39
≥ 150 mg/dL 15 32,6
Table 8. Analysis correlation on RBP4,triglyceride,and apo B100
Triglyceride
RBP4 R
p
n
0,478
0,001
46
Apo B100
p
n
0,322
46
Spearman Correlation Test
This study showed that there was not much difference in the frequency distribution of gender between male and female, 43.5% and 56.5% respectively (Table 1). However, women had a higher percentage of body fat compared to men. There is a difference between the fatty acid metabolism site in men and women. Firstly, catecholamines that mediate the release of free fatty acids are lower in women than in men, where free fatty acids are released from the upper body depots to be comparable. Secondly, free fatty acids released from upper body subcutaneous fats are higher in men than in women, which means there is high resistance of antilipolytic effects of the food digestive process in the upper body fat storage in men. Thirdly, there is a suggestion that basal fat oxidation (regulating Fat Free Mass) is lower in women than in men, therefore women have higher fat stores. Lastly, postprandial fat stores will be higher in the subcutaneous adipose tissue in women than in men (Blaak, 2001).
The upper limit of RBP4 level for normal people has not been established, but the reported RBP4 minimal concentration is 15-30 μg / mL in healthy individuals without vitamin A deficiency (Gamble, 2001 and De Pee, 2002). In 2007, Graham reported the mean of the highest RBP4 concentrations in people with normal glucose tolerance using several methods which are sandwich ELISA with the amount of 34 μg/mL (SB: 9), competitive Enzyme-linked immunoassays (EIAs): EIA no.1 of 25 μg / mL (SB: 4) and EIAs no.2 of 34 μg / mL (SB: 5) (Table 3).
concluded that RBP4 could be a mediator that connected visceral obesity with insulin resistance and atherosclerotic vascular changes (Table 3).
Retinol Binding Protein 4 (RBP4) is secreted by adipose tissue and RBP4 is exclusively expressed in mature adipocytes (Vitkova, 2007). Although RBP4 is expressed primarily in mature adipocytes, there is a difference in gene expression between two fat stores, this may be affected by higher absorption by macrophages in the viscera compared to subcutaneous (Cancello, 2006). As a result, it is expected that the higher down regulation of RBP4 in visceral fat is due to signals derived from macrophages such as TNF α (Sell, 2007).
Obesity is associated with the increased prevalence of dyslipidemia. Dyslipidemia is the amount of abnormal lipids such as cholesterol and triglycerides, in the blood and is considered a risk factor for cardiovascular disease. Obesity is associated with dyslipidemia mainly characterized by increase in plasma free fatty acid (FFA) and elevated triglycerides, decreased high-density lipoprotein (HDL) and increased low-density lipoprotein (LDL) (Saleh, 1999). An increase in free fatty acid (FFA) released from adipose tissue through lipolysis can increase FFA to the liver tissue. Increased FFA will increase triglycerides and Very-Low-Density Lipoprotein (VLDL) in the liver as well as inhibiting lipoprotein in adipose tissue and skeletal muscle, thereby causing hypertriglyceridemia. Triglycerides present in the VLDL are subsequently exchanged for cholesterol ester derived from LDL and HDL, and produce triglyceride-rich LDL and HDL (Jung, 2014) (Table 4).
Many significant contributing factors for obesity associated with dyslipidemia such as uncontrolled release of fatty acids from adipose tissue, especially visceral adipose tissue, after lipolysis leads to increased fatty acid delivery to the liver and for VLDL synthesis. Increased free fatty acid (FFA) may decrease the expression of mRNA or lipoprotein lipase (LPL) activity in adipose and skeletal muscle tissue as well as increased synthesis of VLDL in the liver may inhibit chylomicron lipolysis which may increase hypertriglyceridemia (Saleh, 1999).
(PI3K) in the muscle where this organ is one of the targets of insulin action and decrease in phosphorylation of Insulin Receptor Substrate-1 (IRS-1) (Wolf, 2007). Insulin resistance and insulin deficiency can decrease the activity of lipoprotein lipase (LPL), leading to decreased VLDL breakdown and increased triglyceride production in the liver (Annuzzi, 2004) (Table 5).
The second is that RBP4 activates Sterol regulatory element binding Protein 1 SREBP-1 through peroxisome proliferator-activated receptor-ϒcoactivator 1 β (PGC-1β) dependent in HepG2 cells and contributes to increase lipogenesis in the liver (Xia, 2013). RBP4 increases both PGC1β mRNA and its protein content (Xia, 2013). In which overexpression of PGC1β can lead to hypertriglyceridemia, hypercholesterolemia and abnormal VLDL, IDL/LDL. Overexpression of PGC1β induces SREBP which can increase serum triglyceride and VLDL cholesterol and lower triglycerides contained in the liver (Lin, 2005).
Sterol regulatory element binding Protein (SREB) is known as an important transcription factor in controlling the biosynthesis of cholesterol, fatty acid, and triglyceride. The mammalian genome has two SREBP genes: SREBP-1 and SREBP-2. (Horton, 2002). Hepatic overexpression of SREBP1 can stimulate lipogenesis (Horton, 2003). The expression of SREBP1 produces 2 different isoforms namely REBP1a and SREBP 1c (Horton, 2002). Although HepG2 cells more commonly contain SREBP1a than SREBP 1c, only SREBP 1c can be the main isoform of RBP4 work. Thus in vitro and in vivo RBP4 induce SREBP1c and produce triglyceride accumulation and VLDL secretion in the liver (Xia, 2013).
The third is that the variation of RBP4 is also associated with RBP4 level and triglyceride level as well as the risk of hypertriglyceridemia. Similar with the results of the study by Wu, 2009 it was found that gene polymorphisms rs 3758538 (-1265> C) and rs 10882273 (+11880 T> C) are associated with low levels of RBP4. However, gene variation rs 3758538 C-Allele showed significantly strong association with decreased risk of hypertriglyceridemia and people who carry this allele have significantly low plasma triglyceride levels. As a result, the rs 3758538 C-allele gene can protect against high triglyceride levels and the risk of hypertriglyceridemia. Munkhtulga, 2007 found that genetic variation (-803 G> A, rs 3758539) can efficiently increase the transcription within hepatocarcinoma cells as well as the efficient bonding of the hepatic 1α nuclear factor and its alleles and may be associated with circulating RBP4 level (Table 6,7,8).
reduce the level of RBP4. In addition, triglyceride level was not normally distributed because researchers did not consider the food intake factor.
CONCLUSIONS
The conclusion of this study is that there are no correlation between Retinol Binding Protein 4 with Apo B1000 in women with central obesity in North Sumatera
ACKNOWLEDGEMENT
The authors declare no conflict of interest regarding the publication of this work.
References:
[1] Sanchez, A.F., Santilan, E.M., Bautista, M., Soto, J.E., Gonzalez, A.M., Chirino, C. E., Montiel, I. D., Rivera, G. S., Vega, C. V., and Gonzalez, J.M., 2011. Inflammation, Oxidative Stress, and Obesity. Int. J. Mol. Sci.,12 : 3117-3132.
[2] Amos, A. F., McCarty, D. J., Zimmett, P., 1997. The rising global burden of diabetes and its complications: estimates and projections to the year 2010. Diabet Med., (14 suppl 5): S1-S85.
[3] Malik, S., Wong, N. D., Franklin, S.S., Kamath, T.V., L’Italien, G. J., Pio, J .R., Williams, G. R., 2004. Impact of the Metabolic Syndrome on Mortality From Coronary Heart Disease, Cardiovascular Disease, and All Causes in United States Adults. Circ.aha journals., 110 : 1245-1250
[4] Usfar, A. A., Lebenthal, E., Achadi, A., Soekirman, Hadi, H., 2010. Obesity as a poverty-related emerging nutrition problems: the case of Indonesia. International Association for the Study of Obesity. 11 : 924–928.
[5] Ferrier, D.R., 2014. Metabolisme kolesterol, lipoprotein dan sterol dalam Edisi 6. Tangerang selatan. Binarupa aksara publisher. 341-379.
[6] WHO, 2000. Obesity: Preventing and Managing the Global Epidemic. Report of a WHO consultation. Geneva,Switzerland.
[7] Rabe, K., Lehrke, M., Parhofer, K. G., Broedl, U. C., 2008. Adipokines and Insulin Resistance. Mol med., 14 : 741-751
[9] Kovacs, P., Geyer, M., Berndt, J., Kloting, N., Graham, T.E., Bottcher, Y., Enigk, B., Tonjes, A., Schleinitz, D., Schon, M.R., Kahn, B. B., Bluher, M., Stumvoll, M. 2007. Effects of genetic variation in the human retinol binding protein-4 gene (RBP4) on insulin resistance and fat depot-spesific mRNA expression. Diabetes., 56: 3095-3100.
[10] Chambon, P. A. 1996. Decade of molecular biology of retinoic acid receptors.FASEB J., 10 : 940–954.
[11] Eynatten, M.P., Lepper, P.M., Liu, D., Lang, K., Baumann, M., Nawroth, P.P., Bierhaus, A., Dugi, K.A., Heemann, U., Allolio, B., Humpert, P.M. 2007. Retinol-binding protein 4 is associated with components of the metabolic syndrome, but not with insulin resistance, in men with type 2 diabetes or coronary artery disease.Diabetologia., 50 : 1930–1937.
[12] Xia, M., Liu, Y., Guo, H., Wang, D., Wang, Y., Ling, W. 2013. Retinol binding protein 4 stimulates hepatic sterol regulatory element-binding protein 1 and increases lipogenesis through the peroxisome proliferator-actived receptor-ϒcoactivator 1 β -dependent pathway. Hepatology., 58 : 564-575.
[13] Kotnik, P., Fischer, P. P., Wabitsch, M. 2011. RBP4: a controversial adipokine. EJE. 165: 703-711.
[14] Haider, D. G., Schindler, K., Prager, G., Bohdjalian, A., Luger, A ., Wolzt, M., Ludvik,B. 2007. Serum retinol-binding protein 4 is reduced after weight loss in morbidly obese subjects. JClinEndocrinolMetab., 92 : 1168–1171.
[15] Qi, Q., Yu, Z., Ye, X., Zhao, F., Huang, P., Hu, F. B., Franco, O. H., Wang, J., Li, H., Liu, Y., Lin, X. 2007. Elevated retinol-binding protein 4 levels are associated with metabolic syndrome in chinese people.J Clin Endocrinol Metab., 92 : 4827-4834. [16] Liu, Y., Chen, H., Wang, J., Zhou, W., Sun, R., Xia, M. 2015. Elevated retinol binding
protein 4 induces apolipoprotein B production and associates with hypertriglyceridemia. J Clin Endocrinol Metab., 1- 9.
[17] Baumgartner, R. N., Stauber, P. M., McHugh, D., Koehler, K. M., Garry, P. J. 1995. Cross sectional age differences in body composition in persons 60_ years of age. J Gerontol A Biol Sci Med Sci., 50: 307–16.
[19] Hallfrisch, J., Muller, D., Drinkwater, D., Tobin, J., Andres, R. 1990. Continuing diet trends in men: the Baltimore Longitudinal Study of Aging (1961– 1987). J Gerontol., 45:186–91.
[20] Sivaprasadarao, A., Findlay, J.B., 1988. The interaction of retinol binding protein with its plasma-membrane receptor. Biochem J. 255: 561-569.
[21] Blaak, E. 2001. Gender differences in fat metabolism. Curr Opin Clin Nutr Metab Care. 4: 499-502.
[22] Gamble, M.V., Ramakrishnan, R., Palafox, N.A., Briand, K., Berglund, L., Blaner, W.S. 2001. Retinol Binding Protein as a surrogate measure for serum retinol: studies in vitamin A-deficient children from the republic of the Marshall Islands. Am J Clin Nutr., 73: 594-601.
[23] De Pee, S., Dary, O. 2002. Biochemical indicators of vitamin A deficiency: serumretinol and serum retinol binding protein. J Nutr., 132: 2895S-2901S.
[24] Lee, J. W., Im, J. A., Lee, H.R., Shim, J. Y., Youn, B. S., Lee, D. C. 2007. Visceral adiposity is associated with serum retinol binding protein-4 levels in healty women.Obesity., 15 : 2225-2232.
[25] Maeda, K., Okubo, K., Shimomura, I., Muzuno, K., Matsuzawa, Y., Matsubara, K. 1997. Analysisi of an expression profile of genes in the human adipose tissue. Gene. 190(2): 227-235.
[26] Wu, Y., Li, H., Loos, RJ. F.,Qi, Q., Hu, FB., Liu, Y., Lin, X. 2009. RBP4 variants are significantly associated with plasma RBP4 levels and hypertriglyceridemia risk in Chinese Hans. Journal of Lipid Research. 50: 1479- 1486.
[27] Chambon, P. A. 1996. Decade of molecular biology of retinoic acid receptors. FASEB J. 10 : 940–954.
[28] Cancello, R., Tordjman, J., Pitou, C., Guilhem, G., Bouilloty, J. L., Hugol, D., Coussieu, C., Basdevnaty, A., Bary, H. A., Bedossa, P., Guerre-Millo, M., Clement, K. 2006. Increased Infiltration Of Macrophages In Omental Adipose Tissue Is Associated With Marked Hepatic Lesions In Morbid Human Obesity. Diabetes. 55: 1554-1561.
[30] Saleh, J., Sniderman, A.D., Cianflone, K. 1999. Regulation of Plasma fatty acid metabolism. Clin. Chim. Acta.286: 163–180.
[31] Jung, U. J., Choi, M. S. 2014. Obesity and Its Metabolic Complications: The Role of Adipokines and the Relationship between Obesity, Inflammation, Insulin Resistance, Dyslipidemia and Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci.,15: 6184-6223.
[32] Christou, G.A., Tselepis, A.D., Kiortsis, D.N. 2012. The Metabolic Role of Retinol Binding Protein 4:An Update.Horm Metab Res., 44: 6–14.
[33] Annuzi,G., De Natale, C., Lovine, C., Patti, L., Di Marino, L., Coppola, S., Del Prato, S., Riccardi, G., Rivellese, A. A. 2004. Insulin resistance is independently associated with postprandial alterations of triglyceride-rich lipoproteins in type 2 diabetes mellitus. Arterioscler Thromb
[34] Lin, J., Yang, R., Tarr, P. T., Wu, P. H., Handschin, C., Li, S., Yang, W., Pei, L.,Uldry, M., Tontonoz, P., Newgard, C. B., Spiegelman, B. M. 2005. Hyperlipidemiceffects of dietary saturated fats mediated through PGC-1_ coactivation ofSREBP. Cell., 120:261–273.
[35] Horton, J.D., Goldstein, J.L., Brown, M.S. 2002. SREBPs: activators of the completeprogram of cholesterol and fatty acid synthesis in the liver. J Clin Invest. 109:1125-1131.
[36] Horton, J.D., Shah, N.A., Warrington, J.A., Anderson, N.N., Park, S.W., Brown, M.S., Goldstein, J.L. 2003. Combined analysis of oligonucleotide microarray data from transgenic and knockout mice identifies direct SREBP target genes. Proc Natl Acad Sci U S A.100:12027-12032.