ISSN 0012-4966, Doklady Biological Sciences, 2007, Vol. 416, pp. 341–344. © Pleiades Publishing, Ltd., 2007.
Original Russian Text © S.V. Naidenko, A.L. Antonevich, M.N. Erofeeva, 2007, published in Doklady Akademii Nauk, 2007, Vol. 416, No. 2, pp. 274–277.
341 The ACTH-challenge test is mainly used for evalua-tion of the animal physiological state (stress level) before the injection [1] or determination of metabolites of glucocorticoids in excretions during the develop-ment of noninvasive methods of hormonal monitoring [2]. Now this test is widely used in different animals. Introduction of adrenocorticotropic hormone (ACTH) results in a rapid increase in the concentration of gluco-corticoids in the blood plasma of mammals. The metab-olites of hormones released in blood are excreted from body in urea and excrements after a certain time period. Their determination in the excretions and detection of the peak corresponding to the release of the hormone into blood is the basis for validation of noninvasive methods of hormonal monitoring.
The hormonal response to introduction of ACTH has not been studied well in Felidae. The levels of test-osterone and lactogenic hormone in the blood plasma have been evaluated only in single cases along with concentration of glucocorticoids [3]. The changes in the release of other hormones of adrenal gland have never been studied. However, injection of ACTH may increase the release of other hormones of the adrenal gland, and this may considerably complicate the devel-opment of noninvasive methods due to an increase in the concentrations of the metabolites of other hormones with similar structure in the excretions. The age depen-dence of the response to ACTH injection and specific characteristics of this response during early postnatal ontogeny have never been studied before.
The lynx (Lynx lynx) is one of the most widespread wild representatives of the cat family, which recently became a model animal for hormonal studies. These species were used to study the seasonal dynamics of sex hormones and their metabolism [4, 5]. However, pre-liminary studies, including studies using the
ACTH-challenge test, did not help in the development of a non-invasive method of monitoring of the level of glucocor-ticoids in lynx (our data). Evaluation of the effects of ACTH injection on the release of different hormones into blood may help to find the solution to this problem. In addition, preliminary studies on behavior of lynx pups during ontogeny suggest that intrabrood aggres-sion in lynx pups [6] may be associated with the func-tioning of the adrenal glands [4, 7]. The intrabrood aggression was observed in lynx pups aged 36–64 days and the maximum of fights corresponds to the seventh week of development of the pups. Introduction of a syn-thetic analog of ACTH at this age induced fights between littermates within one hour (our data). In this study, we evaluated the changes in the level of hor-mones in lynx pups after the introduction of a synthetic analogue of ACTH. The data obtained are necessary for further development of noninvasive methods. In addi-tion, we tried to find differences in the response induced by ACTH in lynx pups of different ages—in the begin-ning and end of the period of intrabrood aggression.
The work was performed at the Chernogolovka research base of the Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences, in June–July 2006. The pups of the European subspecies of lynx (L. l. lynx) born in May 2006, were kept together with their mothers in open-air cages whose area was 74 m2
[6, 7].
The synthetic analog of ACTH (referred to as ACTH hereinafter) was injected to 11 pups from 4 different broods at ages of 37 (three pups), 43 (two pups), 58 (three pups), and 65 (three pups) days. Synacten (1 mg/ml, Novartis Pharma, Nurnberg, Germany) was intramuscularly injected to the pups in the morning (8.00 a.m.–10.30 a.m.). Each animal was weighed, and the dose of hormone was calculated for each individual pup with its weight taken into account (0.25 ml/10 kg, i.e., 25 IU/10 kg). A control blood sample was taken from each pup to analyze the level of hormones. The time interval from the moment of capture of the pup to the first blood sampling was as long as 10 min; hence, the level of hydrocortisone in the blood plasma could
PHYSIOLOGY
Hormonal Response to a Synthetic Analog
of Adrenocorticotropic Hormone in Lynx Pups (
Lynx lynx L.
)
S. V. Naidenko, A. L. Antonevich, and M. N. Erofeeva
Presented by Academician D.S. Pavlov March 20, 2007Received March 22, 2007
DOI: 10.1134/S0012496607050055
342
DOKLADY BIOLOGICAL SCIENCES Vol. 416 2007
NAIDENKO et al.
be considered basal [8]. Blood samples of 0.5–1.5 ml were collected from the shoulder superficial vein or femoral vein 30, 60, 90, and 120 min after the injection. The blood was centrifuged (6000 rpm, 20 min), the plasma was collected, frozen at –18°C, and kept at this temperature until measurements. The levels of test-osterone, progesterone, hydrocortisone, and dehydroe-piandrosterone sulfate (DHEA) were measured by ELISA with the use of kits produced by Immunotekh (Moscow, Russia), and the level of androstenedione was determined using a kit purchased from DRG (Ger-many). Each measurement was performed twice to evaluate the variation coefficient; the mean value was used for further analysis. Statistical analysis was per-formed with the use of Wilcoxon’s matched pair test (for evaluation of changes in the level of hormones after the injection) and Student’s test (for comparison of changes in the level of hormones in the pups of differ-ent age groups: pups younger than 45 days and older ones).
The average basal level of hydrocortisone in the lynx pups was 98 ± SE14 ng/ml (lim, 50–182 ng/ml) (Fig. 1). A threefold increase in the ACTH level was observed half an hour after the injection, and the maxi-mum level of this hormone (441 ± 46 ng/ml; lim, 257– 801 ng/ml) was achieved 120 min after the ACTH intro-duction. The level of hydrocortisone in the blood plasma of lynx pups was significantly higher 30, 60, 90, and 120 min after the injection as compared to the con-trol (in each case, T = 0, Z = 2.93; p < 0.01). We noted a considerable individual variation in the response of pups to the injection: the glucocorticoid index varied from 1.9 to 12.6 (5.6 ± 1.1). The peak of hydrocortisone in all five pups of the junior group and one of six older pups (p < 0.05 in the test of differences between propor-tions) was achieved 90 min after the injection, whereas in the rest of older pups, this peak appeared only 120 min after the injection. The increase in the level of hydrocortisone was practically the same: by a factor of 5.4 in junior pups and by a factor of 5.7 in the older pups.
The average basal level of testosterone in lynx pups was 0.31 ± 0.05 ng/ml (lim, 0.16–0.74). The level of this hormone was not significantly changed during the experiment (T = 21–32; Z = 0.18–1.07; ns), but, within 30–120 min, its average value was higher by a factor of 1.26–1.47 than before the injection. A twofold increase in this parameter was observed in older pups; however, the differences in the response of pups of different ages were insignificant (t = 1.22; d.f. = 9; ns). We also did not find any significant changes in the concentration of another androgen, DHEA. The basal level of DHEA was 0.017 ± 0.004 ng/ml, and 1 h after the injection, it was negligibly decreased.
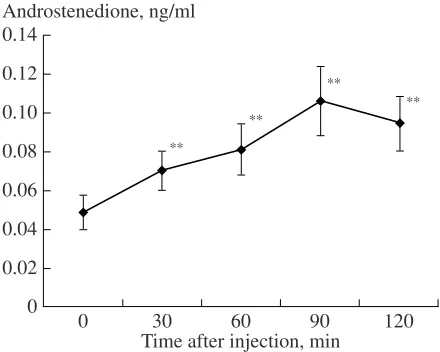
The basal level of androstenedione in the blood plasma of lynx pups was 0.049 ± 0.01 ng/ml (Fig. 2). The level of androstenedione measured during 2 h after the ACTH injection was significantly higher than the basal level (T = 0–2, Z = 2.60–2.85; n = 10–11; p < 0.01). This parameter increased by a factor of 2 (2.3 ±
0.22), achieved the maximum within 90 (in five of nine pups) or 120 min (in four pups). In one pup, we did not observe an increase in the level of androstenedione after the injection, whereas in others, it increased by a factor of 1.4–3.5. No differences in the level of andros-tenedione were found in lynx pups of different ages: it increased by a factor of 2.2 or 2.3.
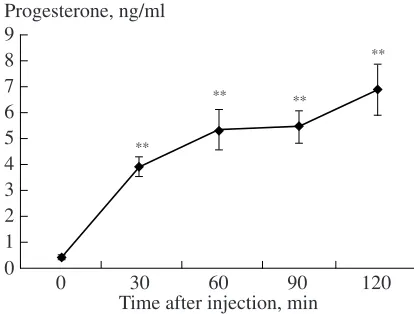
The basal concentration of progesterone in lynx pups was very low (0.42 ± 0.06 ng/ml). The introduc-tion of ACTH induced a rapid increase in the level of progesterone in the blood plasma of lynx pups. This parameter remained at a high level for two hours (in each case, T = 0; Z = 2.80–2.93; n = 10–11; p < 0.01) (Fig. 3). The level of progesterone was increased, on average, by a factor of 26.5 ± 7.5 (lim, 5.8–74.1); in eight of ten pups, it was maximum two hours after the ACTH injection (in two pups, it achieved the maximum within 90 min). The level of progesterone in lynx pups
100 0
0
** **
** **
60
30 90 120
Time after injection, min
200 300 400 500 600
Hydrocortisone, ng/ml
Fig. 1. The average level of hydrocortisone in the blood plasma of lynx pups after the injection of ACTH. Here and in Figs. 2 and 3: ** significant difference (p < 0.01) from the control (point 0).
0.02 0
0
** **
**
**
60
30 90 120
Time after injection, min
0.14
Androstenedione, ng/ml
0.04 0.06 0.08 0.10 0.12
DOKLADY BIOLOGICAL SCIENCES Vol. 416 2007
HORMONAL RESPONSE TO A SYNTHETIC ANALOG 343
of different ages also increased to the same extent, by a factor of 24.7 or 28.2.
Stimulation of the adrenal glands by ACTH intro-duction was previously performed in a number of Fel-idae. The most common model species is the cat (Felis catus) [8], however, similar experiments were also per-formed with the puma (Puma concolor), cheetah ( Aci-nonyx jubatus), tiger (Pantera tigris), and leopard (Panthera pardus) [3]. The ACTH introduction signifi-cantly increased the level of hydrocortisone in all spe-cies [3]. It achieved the maximum 45 min after the injection in leopards, 60 min in tigers, and 75 min in cheetahs, whereas in pumas, it continued to increase even 120 min after the injection. The high plateau level of hydrocortisone in cats was observed, depending on the experimental conditions, 60 [10], 60–120 [9], and 180 [11] min after the injection of ACTH. On average, the peak level of hydrocortisone in the blood plasma was maximum in pumas (715.8 ng/ml) and leopards (535.6 ng/ml), whereas in tigers and cheetahs it was significantly lower (238.6 and 252.0 ng/ml) [3]. Note that the dose introduced to all animals in the experi-ments was the same and independent of the body weight [12]. The injection of ACTH resulted in an increase in the level of hydrocortisone in the blood plasma of tigers by a factor of 1.7 and in leopards by a factor of 1.4, whereas in cheetahs and pumas, this increase was significantly larger (by a factor of 5.3 and 5.5, respectively). These specific differences may be determined by different dosage of ACTH per unit body weight (pumas and cheetahs are significantly smaller than tigers and leopards). The ACTH-challenge test did not induce any changes in the concentration of test-osterone and lactogenic hormone in any of the four spe-cies [3].
The ACTH injection to the lynx pups induced a 5.6-fold increase in the level of hydrocortisone, which is comparable to the response observed in pumas and cheetahs. No doubt, preliminary regular blood sam-pling may delay the achievement of the maximum con-centration of hydrocortisone as compared to the case when this stress was absent. However, according to our data, prolonged “stresses” (different manipulations and electroejaculation) did not significantly affect the level of hydrocortisone in adult animals 30, 60, and 90 min after the beginning of the procedures. In addition, the continuing increase in the concentration of other ste-roid hormones (androstenedione and progesterone) in lynx pups suggests that the increased secretory activity of the adrenal gland 2 h after the ACTH introduction resulted from the action of ACTH. Thus, the ACTH introduction to lynx pups resulted in the release of a considerable amount of hydrocortisone to blood as soon as 30 min after the treatment, which may have had a considerable influence on the animal behavior.
Treatment with ACTH induced a considerable increase in the blood level of androstenedione, the androgen produced by the adrenal glands [13]. These
changes in the level of androstenedione may be very important for the regulation of intrabrood aggression in lynxes [4]. In addition, introduction of ACTH resulted in a considerable increase in the blood level of proges-terone. The relative increase in the concentration of this hormone was maximum (by a factor of more than 25). A considerable increase in the concentration of terone was also found in cats [9]. It seems that proges-terone was secreted by the adrenal gland because the female cats used in these experiments were sterilized [9]. The level of progesterone in a sterilized male cat with developed progesterone-secreting substances in the adrenal glands was increased by a factor of 7.6 dur-ing the ACTH test, whereas in a normal male, it increased only by a factor of 1.5 [14]. An increase in this parameter in intact females was considerably higher (about sevenfold) [9]. Greater changes in the level of progesterone during the ACTH test in lynx pups may be determined by specific differences between these cat species. The lynx females usually achieve maturity only at an age of two years [7], hence, the ova-ries of female lynx pups did not contain yellow bodies, which are the main source of progesterone in adult lynx females. Therefore, it seems that the changes in the level of progesterone observed in lynx pups are also determined by activation of the adrenal glands.
Note that we did not observe any changes in the con-centration of testosterone or dehydroepiandrosterone sulfate during 2 h after the ACTH introduction. Their level remained low and close to the basal level.
We did not observe considerable age-dependent changes in the response of lynx pups to ACTH intro-duction. We noted only a small difference in the respon-siveness of lynx pups: the peak concentration of hydro-cortisone was achieved earlier in the younger pups (37– 43 days of age) than in older pups (57–65 days). The second month of the pups' life, when the experiment was performed, is very important for their develop-ment. This age is characterized by completion of the formation of the tooth system, transition to eating meat,
0 0
**
60
30 90 120
Time after injection, min
9
Progesterone, ng/ml
** **
**
8 7 6 5 4 3 2 1
344
DOKLADY BIOLOGICAL SCIENCES Vol. 416 2007
NAIDENKO et al.
and development of play behavior. Intrabrood aggres-sion, which results in death of some pups, also appears during this period [6]. Although the consequences of intrabrood aggression (differentiation of rate of growth and social relationships between pups) are clearly seen, the mechanisms that initiate intrabrood aggression are not clear [7]. Some increase in the level of hydrocorti-sone and androstenedione in lynx pups during this period [4] and the fact that some lynx pups begin to attack their littermates one hour after the ACTH intro-duction suggest that intrabrood aggression is associated with the functioning of the adrenal glands. Introduction of ACTH and activation of the adrenal glands results in an increase in the level of androstenedione, hydrocorti-sone, and progesterone in the blood plasma of pups as soon as half an hour after the injection. However, the problem of the trigger mechanism of intrabrood aggres-sion in lynx pups still remains unsolved and requires further studies. The complex response to ACTH sug-gests that injections of androstenedione and hydrocor-tisone at physiologically normal doses should be used in further studies.
The complex response to treatment with ACTH may also complicate the development of methods of nonin-vasive monitoring. The metabolism and release of hor-mones in the body of most Felidae have not been stud-ied well; hence, it is possible that metabolites of hydro-cortisone, androstenedione, and progesterone may have similar structures and cross-react with specific antibod-ies used during ELISA. This is crucial for progesterone and androstenedione, but, presumably, is also important for evaluation of the level of hydrocortisone metabo-lites in excrements. The mean basal level of cortisone (98 ng/ml) was more than 200 times higher than the level of progesterone (0.42 ng/ml) and more than 2000 times larger than the level of androstenedione (0.049 ng/ml). About 82% of hydrocortisone is released from the body in excrements [15]. Hence, progesterone and androstenedione add a very small error during the determination of the level of hydrocortisone metabo-lites in excrements, and this error may be neglected. However, the high level of hydrocortisone in Felidae should be kept in mind during noninvasive measure-ments of the levels of progesterone and androstenedi-one, otherwise, the results should be validated by experiments with introduction of radiolabeled hor-mones [15].
ACKNOWLEDGMENTS
We are grateful to E.V. Pavlova and A.A. Chagaeva for the help with this study.
This work was supported by the Program for Funda-mental Studies of the Presidium of the Russian Acad-emy of Sciences “Biodiversity and Gene Pool Dynam-ics” and the Russian Foundation for Basic Research, project no. 07-04-00899.
REFERENCES
1. Roedel, H.G., Monclus, R., and van Holst, D., Physiol. Behav., 2006, vol. 89, pp. 180–188.
2. Goymann, W., Mostl, E., Van’t Hof, T., et al., Gen. Comp. Endocrinol., 1999, vol. 114, pp. 340–348. 3. Wildt, D.E., Phillips, L.G., Simmons, L.G., et al., Biol.
Reprod., 1988, vol. 38, pp. 245–255.
4. Naidenko, S.V. and Erofeeva, M.N., Dokl. Biol. Sci., 2005, vol. 400, no. 3, pp. 51–53 [Dokl. Akad. Nauk, 2005, vol. 400, no. 3, pp. 423–425].
5. Jewgenow, K., Naidenko, S.V., Goeritz, F., et al., Gen. Comp. Endocrinol., 2006, vol. 149, pp. 151–158. 6. Sokolov, V.E., Naidenko, S.V., and Serbenyuk, M.A.,
Zool. Zh., 1994, vol. 73, pp. 132–138.
7. Naidenko, S.V., Osobennosti razmnozheniya i postna-tal’nogo razvitiya evraziiskoi rysi (Characteristics of the Reproduction and Postnatal Development of the Eur-asian Lynx), Moscow: KMK, 2005.
8. Carlstead, K., Brown, J.L., and Strawn, W., Appl. Anim. Behav. Sci., 1993, vol. 38, pp. 143–158.
9. Chatdarong, K., Ponglowhapan, S., Karlsson, A., and Linde-Forsberg, C., Theriogenology, 2006, vol. 66, pp.1482–1487.
10. Sparkes, A.H., Adams, D.T., Douthwaite, J.A., and Gruffydd-Jones, T.J., J. Small Anim. Pract., 1990, vol.31, pp. 1–4.
11. Schoeman, J.P., Evans, H.J., Childs, D., and Her-rtage,M.E., J. Small Anim. Pract., 2000, vol. 41, pp.552–557.
12. Graham, L.H. and Brown, J.L., Zoo Biol., 1996, vol. 15, pp. 71–82.
13. Frank, L.G., Glikman, S.E., and Light, P., Science, 1991, vol. 252, pp. 702–704.
14. Boord, M. and Griffin, C., J. Am. Vet. Med. Assoc., 1999, vol. 214, pp. 666–669.