Ž .
Animal Reproduction Science 63 2000 65–76
www.elsevier.comrlocateranireprosci
Effect of ACTH-challenge on progesterone and
cortisol levels in ovariectomised repeat
breeder heifers
R. Bage
a,), M. Forsberg
b, H. Gustafsson
c, B. Larsson
a,
˚
H. Rodrıguez-Martınez
´
´
aa ( )
Department of Obstetrics and Gynaecology, Centre for ReproductiÕe Biology Uppsala CRU , Swedish
( )
UniÕersity of Agricultural Sciences SLU , PO Box 7039, SE-750 07 Uppsala, Sweden
b
Department of Clinical Chemistry, Faculty of Veterinary Medicine, Centre for ReproductiÕe Biology Uppsala
(CRU , Swedish Uni) Õersity of Agricultural Sciences SLU , PO Box 7039, SE-750 07 Uppsala, Sweden( )
c
Swedish Dairy Association, Hallsta, SE-631 84 Eskilstuna, Sweden˚
Received 13 January 2000; received in revised form 2 May 2000; accepted 15 May 2000
Abstract
In order to investigate the potential influence of stress as a component of the repeat breeding syndrome, the adrenocortical capacity for steroid production was evaluated in ovariectomised
Ž .
dairy heifers. In repeat breeder heifers RBH , marginally elevated plasma progesterone levels during oestrus, so-called suprabasal progesterone levels, have earlier been measured and are believed to impair fertility. The aim was to distinguish if this progesterone could be of extra-gonadal or in this case, adrenal origin. Baseline levels of plasma cortisol and progesterone were determined as well as the corresponding response after induced acute stress in the form of an
Ž .
adrenocorticotropin ACTH -challenge. Comparisons were made between strictly selected RBH,
Ž .
ns5 and virgin heifers VH , ns5 of the Swedish Red and White breed. The heifers were used as their own pre-challenge controls in a 2-day trial. On the control day, saline was injected i.v. and
Ž w.
on the treatment day, a synthetic analogue of ACTH 60 mg Synachten . Via a jugular vein catheter, blood samples were collected every 30 min for 6 h each day of the experiment. Analyses
Ž .
for plasma progesterone and cortisol were made. RBH had a significantly higher P-0.01
Ž y1. Ž y1.
pretreatment baseline cortisol level 10.1"2.3 nmol l than VH 2.6"0.2 nmol l . Moreover, the cortisol response after stimuli was stronger in RBH than VH, especially concerning
Ž .
total hormone production P-0.001 , but there was also a tendency towards higher peak values
ŽPs0.06 and longer duration of significantly increased hormone concentrations. ŽPs0.08 ..
)Corresponding author. Tel.:q46-18-67-21-74; fax:q46-18-67-35-45.
Ž .
E-mail address: [email protected] R. Bage .˚
0378-4320r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved. Ž .
( )
R. Bage et al.˚ rAnimal Reproduction Science 63 2000 65–76
66
Progesterone concentrations, however, did not differ between the groups. Both baseline levels
ŽPs0.25 and posttreatment production P. Ž s0.45 were of the same magnitude in RBH and VH..
In conclusion, the study could not confirm that suprabasal progesterone concentrations during oestrus in RBH derive from the adrenal glands. Still, apparent differences were found in adrenocortical activity when ovariectomised heifers, VH and RBH, were subjected to an ACTH-challenge. It is suggested that a sustained adrenal stimulation associated with environmental or social stress could be one factor in the repeat breeding syndrome.q2000 Elsevier Science B.V.
All rights reserved.
Keywords: Cattle endocrinology; Repeat breeding; Stress; ACTH; Progesterone; Cortisol
1. Introduction
The influence of stress on reproductive performance of dairy cattle has been shown to
Ž
be of importance where intensive livestock management is applied Nanda et al., 1990;
.
Dobson and Smith, 1995; Wilson et al., 1998a,b . The peri-oestral period is very delicate with its precise schedule for the final development of a pre-ovulatory follicle towards
Ž .
ovulation Dieleman et al., 1983; Hyttel et al., 1989 and is dependent on the interplay
Ž
and regulatory actions of the hypothalamic–pituitary–gonadal axis Rivier and Rivest,
.
1991 . Another neuroendocrine connection, the hypothalamic–pituitary–adrenal axis
ŽHPA exerts profound multilevel inhibitory effects on reproduction. In order to study.
the pituitary–adrenal response to stress, evaluating stress effects on, e.g. reproduction, or
Ž .
simply to test adrenal function, the administration of adrenocorticotropin ACTH has
Ž
been used in several studies Gwazdauskas et al., 1972; Wagner et al., 1972; Alam et al.,
.
1986; Verkerk et al., 1994; Dobson and Smith, 1995 . Concerning stress and reproduc-tion, different hypotheses have been proposed regarding which mechanisms are respon-sible for the negative effects, e.g. inhibiting the pre-ovulatory LH-surge or delaying
ovulation. It may be an effect of glucocorticoids andror progesterone released from the
Ž
adrenal cortex or ACTH from the pituitary affecting the reproductive function for
.
reviews, see Moberg, 1991; Rivier and Rivest, 1991; Rivest and Rivier, 1995 . The secretion of pituitary–adrenal hormones in response to stress is controlled by
hypothala-Ž .
mic corticotropin-releasing hormone CRH and arginine vasopressin, and even CRH itself has been shown to directly affect reproduction by inhibiting
gonadotropin-releas-Ž .
ing hormone secretion Chrousos et al., 1998 .
The events that bring about ovulation are highly dependent and integrated. Slight changes may disturb the maturation of the oocyte and the timing of ovulation and render oocytes that may be able to fertilise but have impaired developmental competence, thus leading to early embryonic death. With a delayed ovulation, the chances of pursuing normal timing during AI are impaired, resulting in fertilisation failure or early embry-onic death and, consequently, in repeat breeding. Repeat breeding is regarded to be one of the most costly reproductive dysfunctions in cattle. During a number of years, strictly
Ž .
defined repeat breeder heifers RBH have been subjected to different studies at the Swedish University of Agricultural Sciences, SLU, with the objective to find possible causes for the syndrome. Findings from these various studies have identified asyn-chronous and deviating hormone patterns in connection with disturbed oestrous events
Ž
( )
R. Bage et al.˚ rAnimal Reproduction Science 63 2000 65–76 67
.
Albihn 1991; Bage et al., 1997 . Marginally elevated, so-called suprabasal, plasma
˚
Ž y1 y1 .
progesterone concentrations 0.5–1.0 nmol l vs. -0.5 nmol l as normality were
measured during oestrus and could either be a consequence of an incomplete pre-ovula-tory luteolysis or be caused by an increased progesterone release from the adrenal glands. In different ways, increased levels of progesterone are known to delay the onset of oestrus: inhibition of the pre-ovulatory LH-surge by negative feedback on the
Ž .
hypothalamus Stoebel and Moberg, 1982; Duchens et al., 1994, 1995b , inhibition of
Ž
endometrial oxytocin receptor formation McCracken et al., 1984; Lamming and Mann,
. Ž
1995 and subsequent release of uterine PGF2a Porter and Behrman, 1971; Beard et al.,
.
1994 . Prolonged exposure to progesterone leads to a variety of uterine responses all of
Ž
which may be due to an inhibitory effect on the synthesis of receptors for oxytocin for
.
reviews, see Silvia et al., 1991; McCracken et al., 1999 .
The present study was designed to measure plasma progesterone and cortisol response to ACTH-treatment in ovariectomised RBH, thereby investigating the relation-ship between adrenocortical function and reproductive physiology and, eventually, to evaluate the potential influence of stress as a component of the repeat breeding syndrome.
2. Materials and methods
2.1. Animals
Ten clinically healthy heifers of the Swedish Red and White breed were studied: five
Ž . Ž
repeat breeders RBH aged 2.5–4 years, with a mean weight of 633 kg range: 560–700
. Ž . Ž
kg and five virgin heifers VH aged 2–2.5 years, with a mean weight of 401 kg range:
.
340–500 kg . An RBH was hereby defined as a heifer that had failed to conceive after three or more inseminations in spite of normal, regular oestrous cycles and no palpatory pathological findings in the genital tract. The VH group consisted of normal, sexually mature heifers never mated or inseminated prior to the experiment. All animals were purchased from farms declared free from bovine virus diarrhea virus and bovine leucosis virus. Prior to the experiment, they were clinically and gynaecologically monitored for at least three consecutive sexual cycles. Mean plasma progesterone concentration around ovulation, determined from six blood samples collected 24 h before and after ovulation,
y1 Ž . y1
was 0.7 nmol l in RBH i.e. suprabasal concentration and 0.3 nmol l in VH.
The heifers were kept tethered in tie stalls and fed hay and water ad libitum. They had become accustomed to handling several months before the actual trial started. During the 2 days of the experimental period, efforts were made to feed and manage the heifers according to normal routines in order to minimise adrenocortical activity in response to experimental procedures or disturbed routines. One single person, familiar to the animals, carried out all sampling.
2.2. Experimental procedure
Prior to the trial, the heifers were bilaterally ovariectomised by lateral laparotomy
( )
R. Bage et al.˚ rAnimal Reproduction Science 63 2000 65–76
68
ovarian origin, represented by plasma progesterone, had reached a constant basal level
y1 Ž .
-0.2 nmol l in all animals a minimum of 7 days were required . Blood was
Ž w
collected via an indwelling jugular vein catheter central venous catheter set, Cook
.
Veterinary Products, Australia . The catheter was inserted under local anaesthesia 1 day prior to the sampling period, or alternatively without local anaesthesia at least 2 h before
Ž
the first sample on day 1 to avoid stress effects from the catheterisation Alam and
.
Dobson, 1986 . A silicone tubing connected to the catheter was stitched to the heifer’s neck to reduce stress at sampling or treatment injection.
The heifers served as their own controls during a 2-day experiment. As a
pretreat-Ž . Ž
ment control sday 1 , 5 ml saline natrium chloride 9 mgrml, Kabi Pharmacia,
. Ž .
Sweden was injected. On the day of treatment sday 2 , a synthetic analogue of
Ž w .
ACTH Synachten , Ciba, Switzerland was administered. In order to obtain a
physio-Ž . w
logical adrenocortical response, a suitable dose of ACTH, 60 mg 48 IU Synachten
Ž
made up to a volume of 5 ml in saline solution, was selected Alam and Dobson, 1986;
.
van der Kolk and Breukink, 1991; Hein and Allrich, 1992; Verkerk et al., 1994 . Blood samples were collected every 30 min during a period of 6 h. Directly after the third blood sample, NaCl or ACTH was injected i.v. via the catheter. Sampling always started at 0800 h to avoid effects from circadian and ultradian rhythms in plasma cortisol
Ž .
concentrations Lefcourt et al., 1993 . Immediately after collection, the samples were
centrifuged at 3000=g for 10 min and the plasma was separated and stored aty208C
to await hormone assay. During the experiment, the behaviour and reactions of the animals were noted in order to explain unexpected elevations of plasma cortisol.
2.3. Hormone determinations
Blood plasma concentrations of progesterone were determined by a modified
lumi-Ž
nescence immunoassay already validated for bovine plasma Forsberg et al., 1993;
.
Duchens et al., 1995a . The intra-assay coefficient of variation was 7.04% and the corresponding inter-assay variation was 10.6%. The detection limit of the assay was 0.1
nmol ly1. Cortisol was measured by a Coat-A-Count, solid phase radioimmunoassay
Ž . y1
Diagnostic Products, Los Angeles CA, USA . The detection limit was 5.5 nmol l .
According to the manufacturer, the antiserum shows low cross-reactivity with
proges-Ž .
terone 0.15% . The intra-assay variation was between 2.2% and 6.3%. The inter-assay coefficient of variation was between 3.8% and 5.2%.
2.4. Statistical analyses
The heifers served as their own controls, thereby reducing the influence of individual variation. Individual baseline levels of plasma progesterone and cortisol were calculated
Ž .
from the day 1 saline treatment analytical results plus the first three pretreatment
samples of day 2. Mean hormone concentration"SD was calculated for each animal.
Values within the limits of the mean"SD were selected and subjected to a repeated
calculation of mean"SD with an additional exclusion of values beyond the range.
( )
R. Bage et al.˚ rAnimal Reproduction Science 63 2000 65–76 69
concentrations. Baseline values are presented with range. Total production of proges-terone and cortisol after ACTH administration was calculated from the area under each
ŽŽ . .
respective curve according to the formula: RsÝ TiqTiq30 r2 300 min, where is0,
30 . . . 300 min. The duration of significantly increased plasma hormone concentrations was determined in relation to individual baseline levels. Comparison of the mean group data concerning total hormone production and duration of significantly increased plasma
Ž w
hormone concentrations was done by a Student’s t-test using Minitab Minitab Release
.
10.2, Minitab, PA, USA .
In accordance with the lower detection limit of the assays, cortisol concentrations
were fixed at 5.5 nmol ly1 and progesterone concentrations at 0.1 nmol ly1. The
concentrations of cortisol and progesterone in the baseline samples were transformed to
Ž .
logarithms in base 10 and differences between groups tested by a repeated measures analysis of variance for effect of group, time and group-time interaction using the
Ž
MIXED procedure of the statistical analysis systems package SAS Institute, Cary, NC,
.
USA, 1996 . Least-square means were obtained for combination of effects and were compared using Student’s t-test.
Correlations between cortisol and progesterone concentrations were calculated for baseline samples and after treatment, respectively.
Results are presented as mean"SEM. Differences P-0.05 were considered
signifi-cant.
3. Results
( )
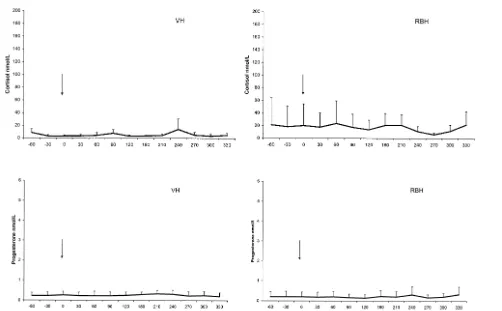
3.1. Baseline concentrations Fig. 1
In many cases, progesterone baseline concentrations were below the detection limit
Ž .
and did not differ between the groups Ps0.25 . In RBH, the mean level was
y1 Ž y1. y1Ž
0.1"0.03 nmol l , range: 0.1–0.5 nmol l , and in VH, 0.2"0.02 nmol l range:
y1. Ž .
0.1–0.4 nmol l . RBH had a significantly higher P-0.01 mean baseline
concentra-y1 Ž y1.
tion of cortisol with a mean of 10.1"2.3 nmol l range: 5.5–54 nmol l compared
y1 Ž y1.
with 2.6"0.2 nmol l in VH range: 2–6 nmol l . For neither hormone was there
any difference over time and no interaction between group of animals and time. One of the RBH had a considerably higher cortisol baseline concentration than the rest of the RBH. She showed consistently nervous and aggressive behaviour during day 1 of the trial. Without her values, the RBH cortisol mean baseline concentration would
y1 Ž y1.
have been 6.2"4.29 nmol l range: 5.5–17 nmol l , which is still significantly
Ž .
higher P-0.001 than the mean for VH. Since her baseline progesterone concentration
did not differ significantly from the rest of the group, it was included in the baseline calculation.
( )
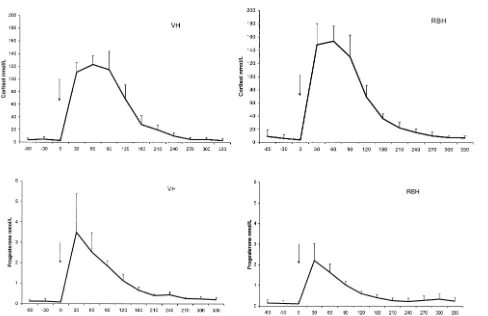
3.2. Posttreatment concentrations Fig. 2
Within 30 min after the ACTH-treatment, all animals had a significant and highly
Ž .
()
R.
Bage
et
al.
r
Animal
Reproduction
Science
63
2000
65
–
76
˚
70
Ž . Ž .
Fig. 1. Control: mean plasma concentrations and SDs of cortisol upper panel and progesterone lower panel in ovariectomised VH, ns5 and RBH, ns5 after
Ž . Ž .
()
R.
Bage
et
al.
r
Animal
Reproduction
Science
63
2000
65
–
76
˚
71
Ž . Ž .
Fig. 2. ACTH-challenge: mean plasma concentrations and SDs of cortisol upper panel and progesterone lower panel in ovariectomised VH, ns5 and RBH, ns5
Ž .
( )
R. Bage et al.˚ rAnimal Reproduction Science 63 2000 65–76
72
Ž .
had a significantly greater P-0.001 cortisol response than VH, most prominent
ŽPs0.06 in the peak values within an hour after the treatment samples no. 4 and 5; 30. Ž
.
and 60 min after the ACTH injection, respectively . Progesterone levels after treatment
Ž .
did not differ Ps0.45 between the groups, although it was complementary to the
release of cortisol. Compared with VH, RBH had a higher peak in cortisol but a smaller rise in progesterone.
The duration of significantly elevated hormone concentrations after treatment did not differ between the groups. In RBH as a group, progesterone concentration was elevated
Ž .
for 204 min range: 150–300 min and the VH group returned to pretreatment
Ž . Ž .
progesterone level after 246 min range: 180–300 min , Ps0.30 . Cortisol
concentra-Ž .
tions were elevated for a longer period, with a mean of 276 min range: 240–300 min
Ž . Ž .
in RBH and 240 min range: 210–300 min in VH Ps0.08 .
4. Discussion
4.1. Stress and repeat breeding
Oestrus is an extremely stress-sensitive period whose carefully regulated events aim to produce a fertilisable oocyte at ovulation and to convey gametes with precise timing in the oviduct. If any of these events are disrupted or delayed, fertilisation failure will occur. In RBH, it is suggested that a hormonal asynchrony during oestrus, induced by suprabasal progesterone concentrations, is the underlying cause for an unsuitable oocyte maturation, a malfunction in sperm transport and an improper timing of the encounter of spermatozoa and the delayed, newly ovulated oocyte. Experimentally induced suprabasal progesterone concentrations in normal heifers caused asynchronies similar to those observed in RBH: prolonged oestrous duration, prolonged and increased oestradiol-17b
Ž
release, delayed LH-peak, resulting in a subsequent lower fertility Duchens et al., 1994;
.
1995a,b . The suprabasal plasma progesterone concentrations could either be a conse-quence of an incomplete luteolysis or an increased progesterone release from extra-gonadal sources. Treatment of RBH with luteolytic drugs has not proven effective according to field practice experience. A second alternative to increased progesterone levels is a sustained adrenal stimulation associated with environmental or social stress. Under experimental conditions, heifers treated with ACTH in the pre-ovulatory period presented increased progesterone concentrations with a inhibition of the LH-surge and
Ž .
oestrous behaviour Stoebel and Moberg, 1982 , fairly similar to what has been detected
Ž .
in RBH Gustafsson et al., 1986; Bage et al., 1997 .
˚
4.2. Adrenal steroid secretion in RBH
( )
R. Bage et al.˚ rAnimal Reproduction Science 63 2000 65–76 73
secretion from the adrenals was, however, definitely higher in RBH. This is obvious already when looking at the hormone baseline concentrations, which were significantly higher for cortisol in RBH, and after treatment, with a significantly higher cortisol peak. Progesterone baseline concentrations were equally low in both groups, with many samples below the detection limit, making it difficult to draw any conclusions. When considering the conversion of pregnenolone to progesterone, there are two pathways
Ž .
available, depending on which of two possible enzymes acts first review: Hall, 1994 . It might be so that in RBH, the cortisol-synthesising pathway is mostly employed in the adrenal glands. Bearing in mind that cortisol and progesterone are not only products deriving from the same precursor pregnenolone, but that progesterone itself is a well-known precursor for cortisol in an additional synthesis pathway, it is suggested
Žalthough not further investigated in this study that RBH might possibly have a higher.
progesterone production capacity when cortisol is not secreted. The duration of signifi-cantly increased cortisol concentrations after treatment was notably longer in RBH than
Ž .
VH Ps0.08 . ACTH administration generally gives a peak response independent of
Ž .
dose and higher doses result in a more prolonged effect Verkerk et al., 1994 . The
significantly heavier RBH were de facto given a lower dose than VHrkg body weight.
When taking into account the dilution effect from their high body weight, large body mass and blood volume, one might speculate that their adrenal response could be even higher than the actual results shown in this study. To clarify this, ACTH should be
administered on unitsrkg body weight in a future study. In the light of the current
results, the advantages of a CRH-challenge should also be considered for an overall physiological reaction that besides adrenal steroid release also includes suppressed immune function and central nervous system-mediated behavioural stress responses.
4.3. Chronic stress and indiÕidual stress sensitiÕity
Ž .
da Rosa and Wagner 1981 reported that when adrenal-ectomised cows were
continuously infused with ACTH from day 2 of the oestrous cycle, plasma concentra-tions of progesterone exhibited a transitory increase and then remained suppressed. Similar observations have been reported in ewes treated twice daily with ACTH on days
Ž .
12–16 of the oestrous cycle Cooke and Benhaj, 1989 . These authors found that while secretion of cortisol remained relatively unchanged, progesterone concentrations de-clined progressively with an increasing number of ACTH injections. The results of these experiments, together with the present findings, suggest that one component of the repeat breeding syndrome may be related to frequent stimulation of the HPA axis. Frequent ACTH administration and chronic stress might change the microsomal environ-ment and the enzyme affinity, and therefore result in a shift in the pathways. Chronic stress may magnify the efficiency of the ACTH-secreting mechanism, expressed by a faster increase in corticosterone in stressed rats compared with controls when additional
Ž .
stress was introduced Sakellaris and Vernikos-Danellis, 1975 . The same response pattern was exhibited in cows already exposed to stressors that showed increased HPA
Ž .
( )
R. Bage et al.˚ rAnimal Reproduction Science 63 2000 65–76
74
There was a consistent ability in cows for the cortisol response to increase, as the ACTH
Ž .
dose increased Verkerk et al., 1998 . In a behavioural study, growing cattle showed a
Ž .
consistency in baseline ACTH and individual stereotypy levels over age Redbo, 1998 . Individual stress sensitivity is thought to be acquired in early postnatal life when stable
Ž
individual differences in HPA responsiveness to stressful stimuli develops Meaney et
.
al., 1996 . This was clearly illustrated in a study where 10 pairs of monozygous twin cows responded differently within pairs when they were subjected to the same ACTH treatment, suggesting that environmental and management experiences rather than
Ž
genetic similarities may modulate an individual’s adrenal response Verkerk et al.,
.
1994 .
In intensive management of dairy cows, genetic selection has increased production levels considerably. However, apart from a favourable increase in production, animals that have been selected for high production efficiency seem to be more at risk for
Ž
behavioural, physiological, and immunological problems for review, see Rauw et al.,
.
1998 . Chronic stress and stereotypies can develop under sub-optimal life conditions where normal behaviour cannot be expressed to a full extent. Many acute stress situations are inevitable in livestock management such as palpation per rectum at oestrus and AI, which causes significant increase in plasma cortisol and may be potential
Ž .
stressors for cows Nakao et al., 1994 .
5. Conclusions
In conclusion, the overall results from the present study indicate a higher capacity for steroid production in the adrenal cortex of RBH compared with control heifers both before and after ACTH-treatment. The results suggest the involvement of the adrenal glands in cases of repeat breeding, thereby including stress as a potential factor in the syndrome.
Acknowledgements
The statistical help from H. Andersson is gratefully acknowledged. Financial support was received from The Swedish Council for Forestry and Agricultural Research.
References
Alam, M.G.S., Dobson, H., 1986. Effect of various veterinary procedures on plasma concentrations of cortisol, luteinizing hormone and prostaglandin F2ametabolite in the cow. Vet. Rec. 118, 7–10.
Alam, M.G.S., Dobson, H., Fitzpatrick, R.J., 1986. Endocrine response to different doses of ACTH in cows. Br. Vet. J. 142, 239–245.
Albihn, A., 1991. Standing oestrus ovarian function and early pregnancy in virgin and repeat breeder heifers. J. Vet. Med., A 38, 212–221.
( )
R. Bage et al.˚ rAnimal Reproduction Science 63 2000 65–76 75
Bage, R., Gustafsson, H., Forsberg, M., Larsson, B., Rodrıguez-Martınez, H., 1997. Suprabasal progesterone˚ ´ ´
levels in repeat breeder heifers during the pro- and oestrous period. Theriogenology 47, 141.
Chrousos, G.P., Torpy, D.J., Gold, P.W., 1998. Interactions between the hypothalamic–pituitary–adrenal axis and the female reproductive system: clinical implications. Ann. Intern. Med. 129, 229–240.
Cooke, R.G., Benhaj, K.M., 1989. Effects of ACTH and cortisol on luteolysis in the ewe. Anim. Reprod. Sci. 20, 201–211.
da Rosa, G.O., Wagner, W.C., 1981. Adrenal–gonad interactions in cattle. Corpus luteum function in intact an adrenalectomized heifers. J. Anim. Sci. 52, 1098–1105.
Dieleman, S.J., Kruip, Th.A.M., Fontijne, P., de Jong, W.H.R., van der Weiden, G.C., 1983. Changes in estradiol, progesterone and testosterone concentrations in follicular fluid and in the micromorphology of preovulatory bovine follicles relative to the peak of luteinizing hormone. J. Endocrinol. 97, 31–42. Dobson, H., Smith, R.F., 1995. Stress and reproduction in farm animals. J. Reprod. Fertil., Suppl. 49,
451–461.
Duchens, M., Forsberg, M., Edqvist, L.-E., Gustafsson, H., Rodrıguez-Martınez, H., 1994. Effect of induced´ ´
suprabasal progesterone levels around estrus on plasma concentrations of progesterone, estradiol-17ß and LH in heifers. Theriogenology 42, 1159–1169.
Duchens, M., Maciel, M., Gustafsson, H., Forsberg, M., Rodrıguez-Martınez, H., Edqvist, L.-E., 1995a.´ ´
Influence of perioestrous suprabasal progesterone levels on cycle length, oestrous behaviour and ovulation in heifers. Anim. Reprod. Sci. 37, 95–108.
Duchens, M., Forsberg, M., Gustafsson, H., Edqvist, L.-E., Rodrıguez-Martınez, H., 1995b. Reproductive´ ´
performance of heifers induced to oestrus asynchrony by suprabasal plasma progesterone levels. Anim. Reprod. Sci. 39, 171–182.
Duchens, M., Rodrıguez-Martınez, H., Forsberg, M., Gustafsson, H., 1996. Ultrastructure of bovine ovarian´ ´
follicles induced to extended growth by perioestrous suprabasal progesterone levels. Acta Vet. Scand. 37, 239–250.
Forsberg, M., Linde-Forsberg, C., Karlsson, A., Carlsson, M.A., 1993. Progesterone and oestradiol in canine plasma monitored by enhanced luminescence immunoassays. J. Reprod. Fertil., Suppl. 47, 127–132. Gustafsson, H., Larsson, K., Kindahl, H., Madej, A., 1986. Sequential endocrine changes and behaviour during
oestrus and metoestrus in repeat breeder heifers and virgin heifers. Anim. Reprod. Sci. 10, 261–273. Gwazdauskas, F.C., Thatcher, W.W., Wilcox, C.J., 1972. Adrenocorticotropin alteration of bovine peripheral
plasma concentrations of cortisol, corticosterone and progesterone. J. Dairy Sci. 55, 1165–1169.
Ž .
Hall, P.F., 1994. Testicular steroid synthesis: organization and regulation. In: Knobil, E., Neill, J.D. Eds. , Physiology of Reproduction vol. 1. Academic Press, New York, Chap. 21.
Hein, K.G., Allrich, R.D., 1992. Influence of exogenous adrenocorticotropic hormone on estrous behaviour in cattle. J. Anim. Sci. 70, 243–247.
Hyttel, P., Greve, T., Callesen, H., 1989. Ultrastructural aspects of oocyte maturation and fertilization in cattle. J. Reprod. Fertil., Suppl. 38, 35–47.
Lamming, G.E., Mann, G.E., 1995. Control of endometrial oxytocin receptors and prostaglandin F2 alpha production in cows by progesterone and oestradiol. J. Reprod. Fertil. 103, 69–73.
Lefcourt, A.M., Bitman, J., Kahl, S., Wood, D.L., 1993. Circadian and ultradian rhythms of peripheral cortisol concentrations in lactating dairy cows. J. Dairy Sci. 76, 2607–2612.
McCracken, J.A., Schramm, W., Okulicz, W.C., 1984. Hormone receptor control of pulsatile secretion of PGF2 from the ovine uterus during luteolysis and its abrogation in early pregnancy. Anim. Reprod. Sci. 7, 31–55.
McCracken, J.A., Custer, E.E., Lamsa, J.C., 1999. Luteolysis: a neuroendocrine-mediated event. Physiol. Rev. 79, 263–323.
Meaney, M.J., Diorio, J., Francis, D., Widdowson, J., LaPlante, P., Caldji, C., Sharma, S., Seckl, J.R., Plotsky, P.M., 1996. Early environmental regulation of forebrain glucocorticoid receptor gene expression: implica-tions for adrenocortical responses to stress. Dev. Neurosci. 18, 49–72.
Moberg, G.P., 1991. How behavioral stress disrupts the endocrine control of reproduction in domestic animals. J. Dairy Sci. 74, 304–311.
( )
R. Bage et al.˚ rAnimal Reproduction Science 63 2000 65–76
76
Nakao, T., Sato, T., Moriyoshi, M., Kawata, K., 1994. Plasma cortisol response in dairy cows to vaginoscopy, genital palpation per rectum and artificial insemination. Zentralbl. Veterinaermed. 41, 16–21.
Nanda, A.S., Dobson, H., Ward, W.R., 1990. Relationship between an increase in plasma cortisol during transport-induced stress and failure of oestradiol to induce a luteinising hormone surge in dairy cows. Res. Vet. Sci. 49, 25–28.
Porter, D.G., Behrman, H.R., 1971. Prostaglandin induced myometrial activity inhibited by progesterone.
Ž .
Nature London 232, 627–628.
Rauw, W.M., Kanis, E., Noordhuizen-Stassen, E.N., Grommers, F.J., 1998. Undesirable side effects of selection for high production efficiency in farm animals: a review. Livest. Prod. Sci. 56, 15–33. Redbo, I., 1998. Relations between oral stereotypies, open-field behavior, and pituitary–adrenal system in
growing dairy cattle. Physiol. Behav. 64, 273–278.
Rivest, S., Rivier, C., 1995. The role of corticotropin-releasing factor and interleukin-1 in the regulation of neurons controlling reproductive functions. Endocr. Rev. 16, 177–199.
Rivier, C., Rivest, S., 1991. Effect of stress on the activity of the hypothalamic–pituitary–gonadal axis: peripheral and central mechanisms. Biol. Reprod. 45, 523–532.
Sakellaris, P.C., Vernikos-Danellis, J., 1975. Increased rate of response of the pituitary–adrenal system in rats adapted to chronic stress. Endocrinology 97, 597–602.
Silvia, W.J., Lewis, G.S., McCracken, J.A., Thatcher, W.W., Wilson, L., 1991. Review: hormonal regulation of uterine secretion of prostaglandin F2aduring luteolysis in ruminants. Biol. Reprod. 45, 663–665. Stoebel, D.P., Moberg, G.P., 1982. Effect of adrenocorticotropin and cortisol on luteinizing hormone surge and
estrous behaviour in cows. J. Dairy Sci. 65, 1016–1024.
van der Kolk, J.H., Breukink, H.J., 1991. Adrenocortical function testing in dairy cows and its effect on milk yield. Vet. Q. 13, 144–147.
Verkerk, G.A., Macmillan, K.L., McLeay, L.M., 1994. Adrenal cortex response to adrenocorticotropic hormone in dairy cattle. Domest. Anim. Endocrinol. 11, 115–123.
Verkerk, G.A., Phipps, A.M., Carragher, J.F., Matthews, L.R., Stelwagen, K., 1998. Characterization of milk cortisol concentrations as a measure of short-term stress responses in lactating dairy cows. Anim. Welfare 7, 77–86.
Wagner, W.C., Strobehn, R.E., Harris, P.A., 1972. ACTH corticoids and luteal function in heifers. J. Anim. Sci. 35, 789–793.
Wilson, S.J., Marion, R.S., Spain, J.N., Spiers, D.E., Keisler, D.H., Lucy, M.C., 1998a. Effects of controlled heat stress on ovarian function of dairy cattle: 1. Lactating cows. J. Dairy Sci. 81, 2124–2131.