www.elsevier.comrlocateranireprosci
In vitro culture of bovine preantral follicles
Sukumar Saha
), Manabu Shimizu, Masaya Geshi, Yoshiaki Izaike
Department of Animal Reproduction, National Institute of Animal Industry, Ministry of Agriculture, Forestry and Fisheries, Tsukuba Norindanchi, P.O. Box 5, Ibaraki 305-0901 Japan
Received 19 August 1999; received in revised form 13 December 1999; accepted 18 April 2000
Abstract
Ž .
Bovine preantral follicles 40–100mm diameter at collection were collected from ovaries of slaughtered cows and cultured in vitro with one of the four treatments: follicle stimulating
Ž . Ž .
hormone FSH; 100 ngrml alone; FSH plus epidermal growth factor EGF; 100 ngrml ; FSH
Ž . Ž .
plus insulin–transferrin–selenium ITS;q1% or FSH plus hypoxanthine 4 mM in tissue culture
Ž . Ž .
medium TCM 199 supplemented with 10% fetal calf serum FCS , 0.1 mgrml sodium pyruvate, 100 IUrml of penicillin and 100mgrml streptomycin. The control culture medium was TCM 199 with supplements without any treatments. Follicles of each size were cultured separately in groups of one to three in 24-well multidishes each containing 500ml of the appropriate culture medium.
Ž . Ž .
Culture commenced at follicle recovery day 1 and continued for 10 days harvested on day 11 . In each case, half the medium was removed and replaced by fresh medium every third day. Follicle diameters were recorded on days 1, 5 and 11 of the experiment. At the end of the 10-day culture period, half of the follicles were stained with trypan blue to assess their potential viability and half were stained with bisbenzimide plus propidium iodine to estimate various morphological features of the follicles.
Follicles of all initial sizes, on all culture treatments, increased in diameter during in vitro cultures with the greatest increases, both in absolute and proportional size, occurring between days 1 and 5 of culture. All of the culture medium supplements caused greater increases in follicle diameters than control medium at both days 5 and 11 of culture for all initial sizes of follicles
Žp-0.01 . The most effective culture supplements for follicles of 40-, 60- and 80-. mm initial diameter were FSH alone and FSHqEGF. The size of these follicles at both days 5 and 11 of
Ž .
culture on both the treatments was significantly larger p-0.01 than follicles cultured in the presence of the other two supplementary treatments. The growth of follicles of 100-mm initial diameter did not differ between culture medium supplements. None of the culture media caused follicle size to increase to the initial diameters of the next larger size category during the 10 days
)Corresponding author. Present address: Agricultural and Forestry Research Center, University of Tsukuba, Tsukuba, Ibaraki, 305-8577, Japan. Tel.:q81-298-53-2596; fax:q81-298-53-6205.
Ž .
E-mail address: [email protected] S. Saha .
0378-4320r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved. Ž .
of culture although follicles of 100-mm diameter achieved a diameter of 120mm, after 4 days of culture.
The overall follicular viability and morphology were better with treatments than the controls in
Ž .
all cases; however, there was no significant difference p)0.05 among them.
From this experiment, FSH and FSH plus EGF may be recommended for in vitro culture of
Ž .
smaller 40, 60 and 80mm follicles.q2000 Elsevier Science B.V. All rights reserved.
Keywords: Cattle ovary; Follicles; In vitro culture; In vitro growth
1. Introduction
Thousands of small oocytes are contained in bovine ovaries, as in the case in other
Ž .
mammalian ovaries, but about 99.9% of oocytes undergo atresia Erickson, 1966 . It would be of great practical benefit if these follicles, destined to become atretic, could be rescued before they degenerate. Periodically, among the pool of primordial follicles, a small population of non-growing oocytes, approximately 30 mm in diameter, begins to
Ž .
grow reaching a final size of 120 mm Fair et al., 1995 . The oocyte grows and the
Ž .
surrounding granulosa cells proliferate as the follicle grows Hulshof et al., 1992 . In cattle, a follicle that has formed an antrum needs 40 days to reach ovulatory size
ŽLussier et al., 1987 , and it is estimated that the preantral follicle requires at least.
60–80 days to reach maturity.
Ž
Growth after culture of preantral follicles could be observed in mice Carroll et al.,
. Ž . Ž
1991; Eppig, 1977; Nayudu and Osborn, 1992 , rats Daniel et al., 1989 , pigs Hirao et
. Ž .
al., 1992 , and cats Jewgenow and Pitra, 1993 .
Several endocrine and growth factors can stimulate the proliferation of granulosa cells from bovine preantral follicles. The ability of gonadotropins to stimulate the proliferation of granulosa cells in primary follicles has been demonstrated in vivo
ŽChiras and Greenwald, 1978 and in vitro Roy and Greenwald, 1989; Carroll et al.,. Ž
. Ž .
1991 . Follicle stimulating hormone FSH has been reported to promote follicle
Ž .
development in cultured bovine preantral follicles Wandji et al., 1996 . FSH has also been demonstrated to be involved in proliferation and differentiation of preantral granulosa cells in vitro and thus in growth and normal in vitro development of preantral
Ž .
follicle of many species, including cows Ralph et al., 1995 . FSH binding and
Ž .
expression of the FSH receptor gene in granulosa cells of human Zheng et al., 1996
Ž .
and bovine Wandji et al., 1992 follicles and in oocytes of primordial follicle from
Ž .
small laboratory animals Roy, 1993 support the view of a physiological action of FSH
Ž .
on preantral follicles Hurk et al., 1997 . FSH is an anti-apoptotic factor and helps to
Ž .
maintain the viability of granulosa cells Chun et al., 1996 . On the other hand, growth factors are ubiquitous peptides, acting in a paracrine andror an endocrine manner, and are involved in regulation of cell proliferation, differentiation and survival. Epidermal
Ž .
growth factor EGF may influence preantral folliculogenesis. EGF has been shown to
Ž .
be involved in proliferation of granulosa cells in pigs Morbeck et al., 1993 , hamsters
ŽRoy, 1993 and cows Wandji et al., 1996 . FSH and EGF are important for the in vitro. Ž .
hormonal and neuropeptidergic compounds as well as cytokines and nutritional
compo-Ž .
nents that initiate and maintain follicular growth Hurk et al., 1997 . There is report of
Ž .
using insulin–transferrin–selenium ITS to maintain granulosa cells within the follicu-lar microenvironment. Insulin is essential for follicle culture; lack of insulin results in follicle degeneration. However, insulin alone is insufficient to maintain healthy follicles. Moreover, selenium acts as an acceptor of free radicals generated during cell metabolism
Ž .
and it improves culture conditions Roy and Treacy, 1993 . The presence of additives such as FSH, ITS, glutamine and sodium pyruvate greatly improved development of the follicles. This was manifested by a twofold increase in the percentage of growth and
Ž
survival time of follicles cultured in the supplemented medium Katska and Rynska,
.
1998 . The number of morphologically normal oocytes increases when bovine preantral
Ž .
follicles are cultured in hypoxanthine-supplemented medium Figueiredo et al., 1994 .
Ž .
Eppig and Downs 1987 demonstrated that hypoxanthine promoted the association of oocytes and accompanying granulosa cells during the culture of mouse preantral follicles. The maintenance of oocyte–granulosa cell communication via gap junctions
Ž .
has been shown to promote oocyte growth in vitro Buccione et al., 1990 . It is probable that hypoxanthine improves the utilization of additional energy substrate by maintaining
Ž
interactions between the oocyte and the surrounding granulosa cells Figueiredo et al.,
.
1994 . Larger variations in the increase of follicular diameter were observed among
Ž .
follicle cultured in the absence of hypoxanthine Jewgenow, 1998 .
Ž
There are several reports using wide range of culture media Daniel et al., 1989;
. Ž
Eppig, 1977; Torrance et al., 1989 , pyruvate Daniel et al., 1989; Eppig, 1977; Eppig
. Ž .
and Schroeder, 1989; Torrance et al., 1989 and glutamine Nayudu and Osborn, 1992 as energy substrate. The addition of pyruvate or glutamine to the culture medium increased the percentage of intact follicle. It was shown that pyruvate was the
predomi-Ž
nant substrate used by immature and mature bovine oocytes and zygotes Khurana and
. Ž .
Niemann, 1992 . Eppig 1976 reported that isolated growing mouse oocytes utilized exogenously administered pyruvate more efficiently than glucose, lactate or succinate. It has been shown that glutamine is an efficient energy substrate for oocytes and embryos
ŽFigueiredo et al., 1994 ..
Cellular maturation and follicular development from the primary to tertiary stages and their regulation by endocrine and paracrine factors are areas of limited knowledge. The ability to culture isolated preantral follicles constitute a new tool for investigation of paracrine and autocrine factors involved in early folliculogenesis. In this experiment, we have studied the effects of some endocrine and growth factors on follicular growth by culturing preantral follicles over a period of 10 days.
2. Materials and methods
2.1. Isolation of preantral follicles
Ovaries were obtained from a local slaughterhouse and placed in isotonic sodium
Ž . Ž .
chloride solution 0.9% NaCl supplemented with penicillin 200 IUrml and
strepto-Ž .
mycin 200mgrml at 30–358C. Sections of the ovarian cortex were excised by scalpel
Ž
.
Engineering, Gomshal, Surrey, England adjusted to produce 500-mm sections. These small to minute segments were placed in Dulbecco’s phosphate-buffered solution
ŽNissui Pharmaceutical, Tokyo, Japan supplemented with 36. mgrml sodium pyruvate, 1mgrml glucose and 3 mgrml BSA. After several washings and repeated pipetting, the
Ž
samples were filtered first through a 500-mm filter Testing Sieve, Tokyo Screen,
.
Tokyo, Japan to remove all large fragments and debris, then through a 38-mm filter
ŽTesting Sieve to exclude blood cells and other minute particles. The tissue remaining. Ž
on the 38-mm filter was washed and collected in an embryo searching dish 100=20
.
mm, Falcon, Becton Dickinson Labware, Becton Dickinson, NJ, USA .
2.2. Culture of isolated preantral follicles
Freshly isolated morphologically normal follicles appearing healthy, spherical under the inverted microscope with one or more compact layers of granulosa cells around the
Ž .
central part containing oocytes within an intact basal membrane, with no apparent sign of necrosis and no antrum were used for present experiment. Preantral follicles with diameters of 40, 60, 80 and 100 mm were collected, selected on the above criteria and
Ž .
cultured in vitro using one of the four treatments culture media . The treatments were:
Ž . Ž
FSH 100 ngrml; pFSH, Denka Pharmaceuticals, Kawasaki, Japan ; EGF 100 ngrml;
. Ž . qŽ
Toyobo, Osaka, Japan qFSH 100 ngrml ; ITS 1%;
Insulin–Transferrin–Selenium-. Ž . Ž
X, Gibco, Grand Island, NY 14072, USA qFSH 100 ngrml ; and hypoxanthine 4
. Ž .
mM; Kohjin, Tokyo, Japan qFSH 100 ngrml . The basal medium for all treatments
Ž
was tissue culture medium TCM 199 with Earle’s salts, L-glutamine, 2200 mgrl
.
sodium bicarbonate, 25 mM HEPES buffer; Gibco supplemented with 10% FCS, 0.1 mgrml sodium pyruvate, 100-IUrml of penicillin and 100 mgrml streptomycin. The basal medium without any supplements for treatments was considered as control. Preantral follicles were cultured in groups of one to three in 500ml of culture medium
Ž .
in 24-well dishes Falcon at 38.58C in 5% CO2 for 10 days. Half the medium was removed and replaced by fresh medium every third day.
Follicular diameters were measured under a microscope with an ocular micrometer on the day of collection and onset of culture, on days 5 and 11 to monitor volume, morphology and any deformity or degeneration. At the end, half of the cultured preantral follicles were stained with trypan blue to categorize their viability on the basis of the degree of dye exclusion. Unstained follicles were classified as viable and fully stained follicles as dead. Follicles with medium staining were regarded as damaged. And the
Ž .
rest of the cultured preantral follicles were double stained by bisbenzimide H 33342
Ž .
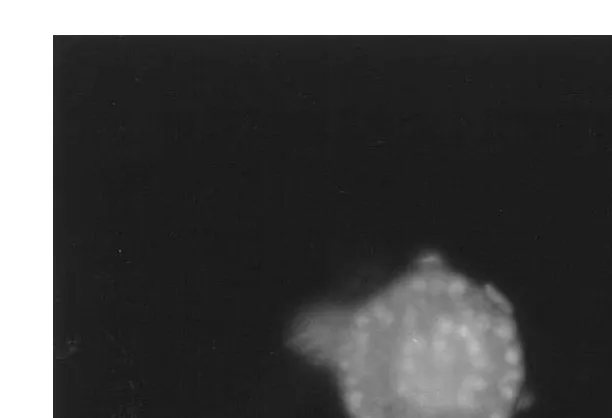
plus propidium iodine 10 mgrml each in PBS and compared with fresh controls to examine the morphological aspects as to determine the presence of an intact follicle membrane, a flattened or cuboidal granulosa cell layer, and the presence of a nucleus. Double staining helped to analyze the proportions of live and dead granulosa cells and
Ž .
also the nuclear components Fig. 1 .
2.3. Statistical analysis
Ž .
Statistical analyses were carried out by one-way analysis of variance ANOVA and
Ž
Fig. 1. A healthy, morphologically normal bovine preantral follicle with circular granulosa cell layers visible
Ž .
after 10 days of in vitro culture after staining with Hoechst 33342 =400 .
.
Concepts, Berkeley, CA, USA for the diameter gain by follicles on different days of culture. The analysis for maintenance of viability and morphology was done by Chi-square test.
3. Results
Follicles of all initial sizes, and on all control and culture treatments increased in diameters during the period of in vitro culture with the greatest increases in absolute and proportional size occurring between days 1 and 5 of culture. The culture medium supplements caused the increase in size to be greater than control at both days 5 and 11
Ž .
of culture for all initial size categorizes p-0.01; Table 1 . For follicles with an initial diameter of 40mm, the FSH alone and FSH plus EGF supplements were more effective
Ž .
at both days 5 and 11 of culture than the other two treatments p-0.01 . The same two
Ž .
treatments were most effective for 60-mm follicles Table 2 at day 5 of culture
Žp-0.01. and, of the two poorer supplements, FSH plus ITS was significantly
Žp-0.01. better than FSH plus hypoxanthine. By day 11 of culture, there were
Ž .
differences between all the treatments with FSHqEGF)FSH p-0.01 )FSHqITS
Žp-0.01.)FSH plus hypoxanthineŽp-0.05 . This pattern of treatment differences.
was established by day 5 of culture when the initial diameter of the follicles was 80mm
ŽTable 3 , FSH. qEGF)FSHŽp-0.05.)FSHqITSŽp-0.01.)FSH plus
hypox-Ž .
Table 1
q Ž
Effect of FSH, FSHqEGF, FSHqITS , FSHqhypoxanthine in culture medium on follicle diameter 40
.
mm , viability and morphology, in vitro cultured for 10 days
Ž .
Treatments Days in culture At the end of culture follicular
Žmeans"SEM; ns20. Žintactrnormal. Values with different superscripts in the same columns are significantly different p-0.05, p-0.01,
.
ANOVA; p)0.05, Chi-square test .
difference between FSH and FSHqEGF was not significant. The four media
supple-Ž .
ments did not affect the size of 100-mm follicles Table 4 by day 5, and there was little difference between them at day 11 although the difference in size at day 11 was
Ž .
significant for the comparison between the FSH and FSHqITS p-0.05 culture supplements. None of the culture media caused the follicle diameter to increase in size to that of the initial diameter of the next larger category of follicle during the 10 days of
Table 2
q Ž
Effect of FSH, FSHqEGF, FSHqITS , FSHqhypoxanthine in culture medium on follicle diameter 60
.
mm , viability and morphology, in vitro cultured for 10 days
Ž .
Treatments Days in culture At the end of culture follicular
Žmeans"SEM. Žns20. Žintactrnormal. Values with different superscripts in the same columns are significantly different p-0.05, p-0.01,
.
Table 3
q Ž
Effect of FSH, FSHqEGF, FSHqITS , FSHqhypoxanthine in culture medium on follicle diameter 80
.
mm , viability and morphology, in vitro cultured for 10 days
Ž .
Treatments Days in culture At the end of culture follicular
Žmean"SEM. Žns20. Žintactrnormal. Values with different superscripts in the same columns are significantly different p-0.05, p-0.01,
.
ANOVA; p)0.05, Chi square test .
culture although follicles, which were 100 mm in diameter at the start of culture had achieved a diameter of 120 mm, the size of an in vitro mature follicle, after 4 days in culture.
The overall follicular viability and morphology were better maintained with the
Ž
treatments than the controls in all cases but, were not significantly different p)0.05,
.
Chi-square test .
Table 4
q Ž
Effect of FSH, FSHqEGF, FSHqITS , FSHqhypoxanthine in culture medium on follicle diameter 100
.
mm , viability and morphology, in vitro cultured for 10 days
Treatments Days in culture At the end of culture
Žmean"SEM. Žns20. Žfollicular. Žintactrnormal.
Values with different superscripts in the same columns are significantly different p-0.05, p-0.01,
.
4. Discussion
We have to be careful to avoid time loss during follicle isolation as follicular viability appeared to be negatively affected by excessive manipulation of preantral follicles before culture, by the duration of time between the beginning of the isolation procedure
Ž
and the onset of culture, and by the composition of the isolation medium Figueiredo et
.
al., 1994 .
There are reports of spreading of granulosa cells from the oocytes when preantral
Ž .
follicle were cultured Eppig, 1992; Mareash et al., 1990 . These preantral follicles were obtained after enzymatic digestion with collagenase or trypsin, and it was reported that they were not surrounded by a basal lamina. In our experiment, we avoided using enzymatic isolation of follicle and replaced it with a mild mechanical isolation
proce-Ž .
dure with modifications as used by Figueiredo et al. 1993a,b . In this case, the basal lamina was preserved, and this might have prevented the spreading of granulosa cells in
Ž .
culture and consequently maintained follicle structure in vitro Figueiredo et al., 1994 . Selection of isolated follicles is also very important as follicles with disrupted basal
Ž
membranes tend to spread on the culture plate surface Jewgenow and Pitra, 1993;
.
Jewgenow and Stolte, 1996 . During in vitro culture, the preantral follicles surrounded by a natural basement membrane may have many advantages including preservation of follicular morphology and maintenance of follicular adhesion to extracellular com-pounds. In addition, it is known that basement membrane contains proteoheparansulfate
ŽWoodley and Sarret, 1993 able to bind a variety of growth factors Gospodarowicz et. Ž .
al., 1978 . Thus, presence of a basement membrane around the follicles might optimize
Ž
the effects of growth factors and hormones added to the culture medium Figueiredo et
.
al., 1995 .
Folliculogenesis from the primordial to the preovulatory stage is regulated by an
Ž
interaction among endocrine, paracrine and autocrine factors in the ovary Jewgenow,
.
1996 . Although very little is known about the regulation of the development of preantral follicles, FSH seems to be the most effective factor in maintaining follicle
Ž .
viability Ralph et al., 1996; Wandji et al., 1996 . Treatment of preantral follicles with FSH stimulates granulosa cell proliferation, antrum formation, steroidogenesis and
Ž
lactate production Boland et al., 1993; Nayudu and Osborn, 1992; Ralph et al., 1995,
.
1996; Wandji et al., 1996 . Growth factors modulate survival, proliferation and
differen-Ž
tiation of follicular cells, acting in interactions with gonadotropins Franchimont et al.,
.
1986; Gospodarowicz and Birdwell, 1977 . Peptide growth factors such as EGF play a crucial role in ovarian folliculogenesis. These are produced locally within the ovary and
Ž .
are involved in the regulation of ovarian processes Jewgenow, 1996 such as cell
Ž .
proliferation, differentiation and steroidogenesis. EGF Boland and Gosden, 1994 is effective in supporting preantral follicle growth. EGF modulate follicular cell growth
Ž .
and differentiation Jewgenow, 1996 and has been demonstrated as having an
anti-Ž .
apoptotic effect on granulosa cells in vitro Hsueh et al., 1994 .
In this experiment, EGF plus FSH and FSH alone improved follicle development. ITS in combination with FSH, in most cases, acted better than hypoxanthine plus FSH. The
Ž
addition of EGF and ITS to the culture medium had a structural supporting effect less
.
factors reduced the percentage of atretic follicle cells and increased the proliferation rate of granulosa cells over controls.
This experiment has improved the growth and development of bovine preantral
Ž
follicles over those achieved in previous experiments Wandji et al., 1996; Figueiredo et
. Ž .
al., 1994 and extended the culture Fig. 2 . The growth rate was high at the beginning of culture and a gradual reduction of these rates was noticed at the end of culture. Our
Ž .
observation is similar to the observation of Katska and Rynska 1998 . Follicles of all initial sizes, and on all control and culture treatments increased in diameters during the period of in vitro culture with the greatest increases in absolute and proportional size occurring between days 1 and 5 of culture. The culture medium supplements caused the increase in size to be greater than control at both days 5 and 11 of culture for all initial size categorizes.
It is shown that supplementation of serum is better for supporting preantral follicle
Ž .
growth Butcher and Ullmann, 1996 . In mice, FCS supports preantral follicular growth
ŽNayudu and Osborn, 1992 . Bovine small preantral follicles can be sustained in culture.
Ž .
in FCS-supplemented medium Figueiredo et al., 1994 . FCS also promotes the
attach-Ž .
ment of any cell type in vitro by providing attachment factors Klebe, 1974 . Further-more, there are reports that preantral follicle survival was influenced by addition of pyruvate to culture medium. Addition of pyruvate, glutamine andror hypoxanthine alone and in mixtures, increase the percentage of morphologically normal follicles
ŽJewgenow, 1998 . In the present experiment, glutamine was already included in TCM.
199, and we added sodium pyruvate to the basal medium.
Trypan blue staining of follicles had been used as a suitable, quick assessment of
Ž .
viability of the cultured follicles Jewgenow and Goritz, 1995 . Hoechst staining allowed classification according to form and number of granulosa cells. Staining small preantral follicles with Hoechst allowed the visualization of individual granulosa cells and thus
Ž .
the determination of their number and shape Motlik and Fulka, 1976 .
The development of an in vitro system that supports small follicle growth in domestic species is an ambitious one. The span of oocyte development is lengthy in the cow when compared with the comparatively short period in the mouse. By extrapolation of data
Ž .
calculated by Lussier et al. 1987 , it was estimated that an early follicle needs
Ž .
approximately 40 days to grow to the preovulatory stage Britt, 1991 . This means that the culture of large bovine preantral follicle will take many weeks, while the culture of
Ž
small preantral follicle will require months to reach full maturity Katska and Rynska,
.
1998 .
In this experiment, we have improved the growth and development of bovine preantral follicles and extended the culture period over some previous observations
ŽWandji et al., 1996; Figueiredo et al., 1994 . Follicles of all initial sizes in culture with.
all the treatments and follicles in controls, increased in diameters. Significantly better follicle growth than the controls was observed with all the treatments, which produced more viable and more morphologically good follicles. Follicular growth was better in
Ž
medium with FSH and FSH plus EGF than in other two media with FSH plus ITS and
. Ž .
treat-Ž . Ž .
Fig. 2. Bovine preantral follicles a on the day of collection b after 10 days of culture in TCM 199
Ž . Ž .
supplemented with FSH 100 ngrmlqEGF 100 ngrml ; an increase in follicular diameters with expansion
Ž .
ments. These limitations might be overcome regarding 100-mm follicles and as a result, no difference among treatments was noticed. This experiment may provide a new route for in vitro culture of bovine early follicle. However, culture systems for the cow are at an early stage of development and are at present being used to define the characteristics of preantral follicle growth rather than as a method of producing meiotically competent
Ž .
oocytes Figueiredo et al., 1993a,b, 1994; Hulshof et al., 1995 . More research is necessary for reaching the final goal to create an environment that will support granulosa cell differentiation and maintain granulosa–oocyte cell contact to enable oocyte development to occur as has been demonstrated in the mouse culture system
ŽEppig and Schroeder, 1989; Eppig and Telfer, 1993 ..
References
Boland, N.I., Humpherson, P.G., Leese, H.J., Gosden, R.G., 1993. Pattern of lactate production and steroidogenesis during growth and maturation of mouse ovarian follicles in vitro. Biol. Reprod. 48, 798–806.
Boland, N.I., Gosden, R.G., 1994. Effects of epidermal growth factor on the growth and differentiation of cultured mouse ovarian follicles. J. Reprod. Fertil. 101, 369–374.
Britt, J.H., 1991. Impacts of early postpartum metabolism on follicular development and fertility. Bovine Prac. 24, 39–43.
Buccione, R., Schroedeer, A.C., Eppig, J.J., 1990. Interactions between somatic cells and germ cells throughout mammalian oogenesis. Biol. Reprod. 43, 543–547.
Butcher, L., Ullmann, S.L., 1996. Culture of preantral ovarian follicles in the grey, short-tailed opossum, Monodelphis domestica. Reprod. Fertil. Dev. 8, 535–539.
Carroll, J., Whittingham, D.G., Wood, M.J., 1991. Effect of gonadotrophins environment on growth and development of isolated mouse primary ovarian follicles. J. Reprod. Fertil. 93, 71–79.
Chiras, D.D., Greenwald, G.S., 1978. Effects of steroids and gonadotrophins on follicular development in the hypophysectomized hamster. Am. J. Anat. 152, 307–320.
Chun, S.-Y., Eisenhauer, K.M., Minami, S., Billig, H., Perlas, E., Hsueh, A.J.W., 1996. Hormonal regulation of apoptosis in early antral follicles: follicle-stimulating hormones as a major survival factor. Endocrinol-ogy 137, 1447–1456.
Daniel, A.J., Armstrong, D.T., Gore-Langton, R.E., 1989. Growth and development of rat oocyte in vitro. Gamete Res. 24, 109–121.
Eppig, J.J., 1976. Analysis of mouse oogenesis in vitro. Oocyte isolation and the utilization of exogenous energy sources by growing oocytes. J. Exp. Zool. 198, 375–382.
Eppig, J.J., 1977. Mouse oocyte development in vitro with various culture systems. Dev. Biol. 60, 371–378. Eppig, J.J., Downs, S.M., 1987. The effect of hypoxanthine on mouse oocyte growth and development in vitro:
maintenance of meiotic arrest and gonadotropin-induced oocyte maturation. Dev. Biol. 119, 313–321. Eppig, J.J., Schroeder, A.C., 1989. Capacity of mouse oocytes from preantral follicles to undergo
embryogene-sis and development to live young after growth, maturation, and fertilization in vitro. Biol. Reprod. 41, 268–276.
Eppig, J.J., 1992. Growth and development of mammalian oocytes in vitro. Arch. Pathol. Lab. Med. 116, 379–382.
Eppig, J.J., Telfer, E.E., 1993. Isolation and culture of oocytes. Methods Enzymol. 225, 77–84.
Erickson, B.H., 1966. Development and senescence of the postnatal bovine ovary. J. Anim. Sci. 25, 800–805. Fair, T., Hyttel, P., Greve, T., 1995. Bovine oocyte diameter in relation to maturational competence and
transcriptional activity. Mol. Reprod. Dev. 42, 437–442.
Figueiredo, J.R., Hulshof, S.C.J., Thiry, M., Van den Hurk, R., Bevers, M.M., Nusgens, B., Beckers, J.F., 1993b. Extracellular matrix proteins and basement membrane: their identification in bovine ovaries and significance for the attachment of cultured preantral follicles. Theriogenology 43, 845–858.
Figueiredo, J.R., Hulshof, S.C.J., van den Hurk, R., Nusgens, B., Bevers, M.M., Ectors, F.J., Beckers, J.F., 1994. Preservation of oocyte and granulosa cell morphology in bovine preantral follicles cultured in vitro. Theriogenology 41, 1333–1346.
Figueiredo, J.R., Hulshof, S.C.J., Thiry, M., van den Hurk, R., Bevers, M.M., Nusgens, B., Beckers, J.F., 1995. Extracellular matrix proteins and basement membrane: their identification in bovine ovaries and significance for the attachment of cultured preantral follicles. Theriogenology 43, 845–858.
Franchimont, P., Hazee-Hagelstein, M.T., Charlet-Renard, C., Jaspar, J.M., 1986. Effect of mouse epidermal growth factor on DNA and protein synthesis, progesterone and inhibin production by bovine granulosa cells in culture. Acta Endocrinol. Copenhagen 111, 122–127.
Gospodarowicz, D., Birdwell, C.R., 1977. Effects of fibroblast and epidermal growth factors on ovarian cell proliferation in vitro: I. Characterization of the response of granulosa cells to FGF and EGF. Endocrinol-ogy 100, 1108–1120.
Gospodarowicz, D., Greenburg, G., Birdwell, C.R., 1978. Determination of cellular shape by the extracellular matrix and its correlation with the control of cellular growth. Cancer Res. 38, 4155–4171.
Hirao, Y., Miyano, T., Kato, S., 1992. In vitro growth of porcine oocytes. 12th Int. Congr. Anim. Reprod. 1, 333–335.
Hsueh, A.J.W., Billig, H., Tsafriri, A., 1994. Ovarian follicle atresia: a hormonally controlled apoptotic process. Endocr. Rev. 15, 707–724.
Hulshof, S.C.J., Bevers, M.M., van der Donk, H.A., van den Hurk, R., 1992. The isolation and characteriza-tion of preantral follicles from foetal bovine oocytes. 12th Int. Congr. Anim. Reprod., Hague 1, 336–338. Hulshof, S.C.J., Figueiredo, J.R., Beckers, J.F., Bevers, M.M., van der Donk, J.A., van den Hurk, R., 1995. Effects of fetal bovine serum, FSH and 17 b-estradiol on the culture of bovine preantral follicles. Theriogenology 44, 217–226.
van den Hurk, R., Bevers, M.M., Beckers, J.F., 1997. In vivo and in vitro development of preantral follicles. Theriogenology 47, 73–82.
Jewgenow, K., Pitra, C., 1993. Hormone-controlled culture of secondary follicle of domestic cats. Theri-ogenology 39, 527–535.
Jewgenow, K., Goritz, F., 1995. The recovery of preantral follicles from ovaries of domestic cats and their characterization before and after culture. Anim. Reprod. Sci. 39, 285–297.
Jewgenow, K., Stolte, M., 1996. Isolation of preantral follicles from non-domestic cats — viability and ultrastructural investigations. Anim. Reprod. Sci. 44, 183–193.
Jewgenow, K., 1996. Impact of peptide growth factors on the culture of small preantral follicles of domestic cats. Theriogenology 45, 889–895.
Jewgenow, K., 1998. Role of media, protein and energy supplements on maintenance of morphology and DNA-synthesis of small preantral domestic cat follicles during short-term culture. Theriogenology 49, 1567–1577.
Katska, L., Rynska, B., 1998. The isolation and in vitro culture of bovine preantral and early antral follicles of different size classes. Theriogenology 50, 213–222.
Khurana, N., Niemann, H., 1992. Energy metabolism of bovine oocytes matured and fertilized in vitro. 12th Int. Congr. Anim. Reprod. 1, 348–350.
Klebe, R.J., 1974. Isolation of a collagen-dependent cell attachment factor. Nature 250, 248–251.
Lussier, J.G., Matton, P., Dufour, J.J., 1987. Growth rates of follicles in the ovary of the cow. J. Reprod. Fertil. 81, 301–307.
Mareash, G.A., Timmons, T.T., Dunbar, B.D., 1990. Effect of matrix on the expression of specific ovarian proteins. Biol. Reprod. 43, 965–976.
Morbeck, D.E., Flowers, W.L., Britt, J.H., 1993. Response of porcine granulosa cells isolated from primary and secondary follicles of FSH, 8-bromo-cAMP and EGF in vitro. J. Reprod. Fertil. 99, 577–584. Motlik, J., Fulka, J., 1976. Breakdown of the germinal vesicle in pig oocytes in vivo and in vitro. J. Exp. Zool.
198, 155–162.
Nayudu, P.L., Osborn, S.M., 1992. Factors influencing the rate of preantral and antral growth of mouse
Ž .
Ralph, J.H., Wilmut, I., Telfer, E.E., 1995. In vitro growth of bovine preantral follicles and the influence of
Ž .
FSH on follicular and oocyte diameters. J. Reprod. Fertil. 15, Abstr. Series , 6 abstr.
Ralph, J.H., Wilmut, I., Telfer, E.E., 1996. The effect of FSH on bovine preantral to early antral ovarian
Ž .
follicle growth in vitro. Biol. Reprod. 54 Suppl 1 , 5 abstr.
Roy, S.K., Greenwald, G.S., 1989. Hormonal requirements for the growth and differentiation of hamster preantral follicles in long-term culture. J. Reprod. Fertil. 87, 103–114.
Roy, S.K., 1993. TGF-bpotentiation of FSH-induced DNA synthesis in hamster preantral follicles is mediated by a latent induction of EGF. Biol. Reprod. 48, 558–563.
Roy, S.K., Treacy, B.J., 1993. Isolation and long-term culture of human preantral follicles. Fertil. Steril. 59, 783–790.
Torrance, C., Telfer, E., Gosden, R.G., 1989. Quantitative study of the development of isolated mouse pre-antral follicles in collagen gel culture. J. Reprod. Fertil. 87, 367–374.
Wandji, S.A., Pelletier, G., Sirard, M.A., 1992. Ontogeny and cellular localization of125I-labeled insulin-like growth factor-I,125I-labeled follicle-stimulating hormone, and125I-labeled human chorionic gonadotropin binding sites in ovaries from bovine fetuses and neonatal calves. Biol. Reprod. 47, 814–822.
Wandji, S.A., Eppig, J.J., Fortune, J.E., 1996. FSH and growth factors affect the growth and endocrine function in vitro of granulosa cells of bovine preantral follicles. Theriogenology 45, 817–832.
Ž .
Woodley, D.T., Sarret, Y., 1993. Connective tissue diseases of the skin. In: Lapiere, C.M., Kriegf, T. Eds. , Marcel Dekker, New York, pp. 41–57.