Marine-mollusc fossils (Mollusca: Bivalvia and Gastropoda) and their
paleoenvironmental interpretation from the Situs manusia purba Sangiran area
Donan Satria Yudha and Rusyad Adi SuriyantoCitation: AIP Conference Proceedings 1744, 020012 (2016); doi: 10.1063/1.4953486 View online: http://dx.doi.org/10.1063/1.4953486
View Table of Contents: http://scitation.aip.org/content/aip/proceeding/aipcp/1744?ver=pdfcov Published by the AIP Publishing
Articles you may be interested in
Accumulation of radionuclides in selected marine biota from Manjung coastal area AIP Conf. Proc. 1659, 050009 (2015); 10.1063/1.4916879
Analysis of Northern bottlenose whale pulses and associated reflections recorded from the Gully Marine Protected Area
J. Acoust. Soc. Am. 133, 3443 (2013); 10.1121/1.4806089
Preliminary analysis of C and H in a “Sangiran” fossil using laser-induced plasma at reduced pressure J. Appl. Phys. 98, 093307 (2005); 10.1063/1.2121930
Some Sounds from Marine Life in the Hawaiian Area J. Acoust. Soc. Am. 24, 116 (1952); 10.1121/1.1917427
Marine-mollusc Fossils
(Mollusca: Bivalvia and Gastropoda)
and Their Paleoenvironmental Interpretation
from the
Situs Manusia Purba Sangiran
Area
Donan Satria Yudha
1, a)and Rusyad Adi Suriyanto
21
Animal Systematics Laboratory, Faculty of Biology, Universitas Gadjah Mada, Jl. Teknika Selatan Sekip Utara, Yogyakarta 55281, Indonesia
2
Laboratory of Bioanthropology and Paleoanthropology, Faculty of Medecine Universitas Gadjah Mada, Jl. Medika, Sekip, Yogyakarta 55281, Indonesia
a)Corresponding author: donan_satria@ugm.ac.id
Abstract. Research on mollusc fossils on the Sangiran area is not widely known. Sangiran area is known as one of the world prehistoric sites because the area has uncovered many fossil organisms. Sangiran stratigraphic chart extends from the oldest geological time to recent one, i.e.: Late Pliocene to Holocene. The stratigraphic chart is divided into four formations, i.e., Kalibeng, Pucangan, Kabuh and Notopuro. Each formation is characterized by different geological sediments and distinctive fossil organisms. The Laboratory of Bioanthropology and Paleoanthropology, Faculty of Medicine, UGM has frequently been conducting excavation in the Sangiran area. Fossil of marine molluscs that had been found so far is not yet identified. The aims of this research are: to identify fossil collection of marine-molluscs to study its diversity during the Pleistocene, and to interpret the paleo-environment of where those fossils were found. The results obtained: from 35 samples of bivalve there are six orders with 12 families and 3 identified genera. From 29 samples of gastropod, there are four orders with 14 families and 13 identified genera. All identified genera of bivalves and gastropods are fauna which habitat in the shallow sea of the intertidal zone. These marine molluscs habitat is matched with the geological studies of Sangiran area in which during the Lower Pleistocene, some part of Sangiran area were shallow sea and estuaries
Keywords: Fossils, identification, marine-mollusc, paleoenvironment, Sangiran.
INTRODUCTION
Research on mollusc fossils on the Sangiran area is not widely known. Some researches on fossil molluscs of Sangiran are Di Geronimo and Sartono [1] and Fae [2, 3]. On the Fae research [2, 3], the mollusks that have been identified is a collection belonging to Ing. Premonowati, from the University of Veteran (UPN) Yogyakarta. The mollusc fossil of the Di Geronimo and Sartono [1] research were the collection of Sartono himself that have been kept in Bandung.
and hominids). The fossils found in the Kalibeng and Pucangan Formations are mostly marine molluscs or other marine fauna such as foraminifers and sharks [1, 2, 3, 4, 5, 6, 7].
The study of paleontology is strongly connected to the study of geology, taxonomy, anatomy, taphonomy, paleo-climatology and paleo-ecology. Taphonomy is the study of the transition of animal remains from the biosphere into the lithosphere [8]. The taphonomical study aimed to reconstruct past environment and biotas in some specific area. In the taphonomical study, it is known that the fossilization occurs on the hard parts of the organism remains such as bones, shells and teeth. The postmortem processes, especially physical process, are highly influenced by the morphological forms of the organism remains to cause incomplete morphological form (undergone fragmentation). The incomplete morphological characters of some fossils create a bias in the taxonomic identification, especially for one who are not experienced about taxonomy, anatomy and taphonomy [8, 9, 10].
Laboratory of Bioanthropology and Paleoanthropology, Faculty of Medicine, UGM has frequently been conducting excavation in the Sangiran area. Vertebrate fossils especially hominids and other animals have been published or used as scientific research (undergraduate thesis) by students of UGM. The invertebrate fossils
especially molluscs’ collection of Laboratory of Bioanthropology and Paleoanthropology have not been identified
and studied until recently. In 2015, we make taxonomical identification of these fossils based on shells
characteristics. This identification is to know molluscs’ diversity in the Sangiran area during Pleistocene epoch and
to interpret its environment.
The purpose of this research is to know and understand the diversity of molluscs; classis Bivalvia dan Gastropoda found in the Pucangan Formation area of the Association of Conservation Sangiran’s Ancient Man
Archaeological Site , Sragen, Jawa Tengah. Samples of molluscs’ fossil obtained from the collection of the
Laboratory of Bioanthropology and Paleoanthropology, Faculty of Medicine, UGM. The identified fossils are important to complete the existing database of the fossils of the phylum Mollusca in Java, Indonesia. These identified fossils could be used to determine paleo-ecology and paleo-climatology [11], and it’s also used as a
reference for future research especially for students how interested in Palaeontology.
MATERIAL AND METHODS
Materials examined are 35 fossil shells of bivalve fossil and 29 fossil shells of gastropod. For gastropods fossil, in each plastic bag, there are 5 to 7 fossil shells which considered same species according to people who did the excavation in the 1970s. There are two types of fossil shells in this collection examined, i.e. mold and cast. Mold and cast are types of fossilization in which physical characters of an organism imprinted on rocks, usually coarse porous rocks such as sandstones. Physical characters imprinted on rocks are hard parts of an organism such as
molluscs’ shells, reptile’s scales, cnidarian exoskeleton structure, vertebrates’ bones and teeth.
On the molluscs’ fossil, “mold” is an imprint of shell’s external characters and “cast” is an imprint of shell’s
internal part. The fossil cast is formed when a mold fossil filled with sediment, and sometimes after cast is formed, the mold then was broken down and leave only its internal part (a cast). A mold type of shell fossil is usually easier
to identified due to the preservation of shell’s external or morphological characteristics.
All molluscs fossil (bivalves and gastropods) analyzed are a collection of the Laboratory of Bioanthropology and Paleoanthropology, Faculty of Medicine, UGM. All samples were results of Sangiran Excavation Project in 1970–
1980s which have not been taxonomically identified. All samples were found in the sediment of the Pucangan Formation.
This research is to analyze shells’ morphological structure and ornamentation of bivalves and gastropods.
Morphological and taxonomical identification based on Rogers [12]; Kira [13]; Habe [14]. Paleo-environmental interpretation is based on two things, first is based on geological references of the Sangiran area during Pleistocene, and second based on the paleo-habitat of identified fossil shells which is identical compare to modern shells habitat of the same genus. The Sangiran’s geological references used are Sartono [5], Di Geronimo & Sartono [1], Huffman
[15], Sémah, et al., [16] and Sémah, et al., [17].
RESULT AND DISCUSSION
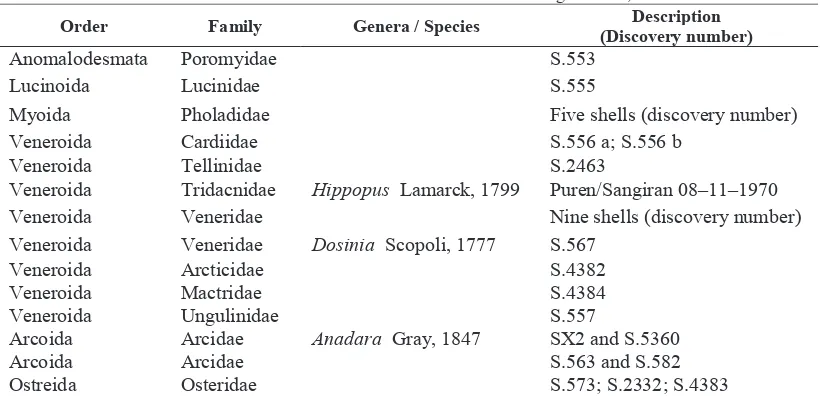
shells identified until genus are belong to 3 different genera, i.e. Hippopus from family Tridacnidae, Dosinia from family Veneridae and Anadara of family Arcidae (Table 1).
Four fossil shells which could not identify until taxon level family, due to its incomplete physical characters caused by taphonomical processes. Shell A (number S.552) the shell remains is a small fragment. Shell B (number S.2402). The shell remains incomplete, of which two lateral sides of shells are not well preserved or fragmented.
TABLE 1. The list of Bivalvia fossil shells discovered in the Sangiran area, Central Java.
Order Family Genera / Species (Discovery number) Description
Anomalodesmata Poromyidae S.553 Lucinoida Lucinidae S.555
Myoida Pholadidae Five shells (discovery number) Veneroida Cardiidae S.556 a; S.556 b
Veneroida Tellinidae S.2463
Veneroida Tridacnidae Hippopus Lamarck, 1799 Puren/Sangiran 08–11–1970
Veneroida Veneridae Nine shells (discovery number) Veneroida Veneridae Dosinia Scopoli, 1777 S.567
Veneroida Arcticidae S.4382 Veneroida Mactridae S.4384 Veneroida Ungulinidae S.557
Arcoida Arcidae Anadara Gray, 1847 SX2 and S.5360 Arcoida Arcidae S.563 and S.582 Ostreida Osteridae S.573; S.2332; S.4383
Systematic Descriptions and Habitat
Classification and morphological description of the marine bivalves and gastropods are mainly based on [13] and [14], with new systematic revision based on “World Register of Marine Species (WoRMS): www.marinespecies.org”. Additional morphological and additional habitat descriptions are based on Carpenter &
Niem [18]. morphology. Discovery number: Puren/Sangiran 08–11–1970.
Diagnostic characters: shown on the fossil shell are shell very convex; generally shell’s profile is fan-shape;
equivalve; inequilateral; shell strongly inflated and longer than high; umbo position is in midline; Hinge with 1 ridge-like cardinal tooth, 2 lateral teeth on right valve, 1 lateral tooth on left valve; outer surface sculptured with 9 to 14 large radial fold with 2 to 3 small rib-like at each interstices. The coloration on outer surface whitish, irregular reddish blotches scattered in irregular concentric bands; inner surface porcelaneous white (Fig. 1).
Habitat: recent Hippopus habitat is in the shallow sea or littoral zone. They found on the sandy areas in clear water among coral reefs, sometimes among sea grass beds adjoining reefs. Their habitat alongside intertidal zone because their diet is small drifting plants on sea current or from very small plant cells (zooxanthellae) that live in the flesh of the clam. Zooxanthellae that lived with the flesh of giant clams require sunlight, so giant clams can only live and grow in the clear and shallow sea.
(a) (b)
FIGURE 1. Fossil of bivalve shell genera Hippopus discovered in Sangiran: (a) is the dorsal view of fossil shell; (b) is an internal and umbilical view of fossil shell (Photo: Yudha, 2015).
Description: A. large radial fold; B. small rib-like; C. Umbo; D. cardinal teeth; E. lateral teeth.
Order Veneroida Superfamily Veneroidea
Family Veneridae Genus Dosinia Scopoli, 1977.
Syn. Dosinorbis Dall, 1902.
Material examined: is an external shell fossil remain which its diagnostic characters are shown on its shell morphology. Discovery number: S.567.
Diagnostic characters: shown on the fossil shell are; shell thick, strong, solid, flattened, equivalve and circular. Umbo position is in the midline. Lunule is small and located in front of the beak, wide and heart-shaped. Escutcheon narrow, lens-shaped. Outer surface with obvious regular concentric ridges and become lamellate and irregular toward antero-dorsal and postero-dorsal margin. Hinge plate wide with three cardinal teeth on each valve and a small bulge on the anterior lateral left valve. Shell color is ivory white with brown radial streaks (Fig. 2).
Habitat: recent Dosinia habitat is in the intertidal zone. They found in the shallow sea with soft bottoms such as sandy, muddy or gravel flats. This animal commonly found in low intertidal to shallow subtidal depth to 80 m in depth. Some species are active burrowers in various soft bottom sediment.
Distribution: recent Dosinia distribution is wide, nearly found in coastal area worldwide such as New Zealand, Atlantic and West Hindies beaches, Japanese, Korean, African and Chinese beaches.
(a) (b)
FIGURE 2. Fossil of bivalve shell genera Dosinia discovered in Sangiran: (a) is dorsal view of fossil shell; (b) is an umbilical view of fossil shell (Photo: Yudha, 2015).
Description: A. umbo; B. concentric lamellate ridges; C. lunule; D. escutcheon.
Subclass Pteriomorphia Order Arcoida Superfamily Arcoidea
Family Arcidae Genus Anadara Gray, 1847
Material examined: there is two fossils of external shell remains which their diagnostic characters can be seen
on shell’s morphology. Discovery number: SX2; and S.5360. Number S.5360 is a fossil of the external shell which
embedded in the hard sediment, but its shell external characters are clearly shown.
Diagnostic characters: on the fossil shell are: shell thick (0.35 to 0.45) mm, whitish and convex; shell covered
with thick periostracum; shell’s posterior part is not twisted towards the left; Shell equivalve, valves exactly meets
on margins; Ventral margin without a byssal gape; inner, ventral margin with strong crenulations corresponding with the external ribs; protruding umbones; strong radial ribs with median grooves (Fig. 3).
Habitat: recent Anadara habitat is in the intertidal zone and estuary, only small number species lived near or on the margin of subtidal zone. This animal lives sedentarily, living attached to the substrate, sometimes nestling in rock crevices or buried in soft bottoms. Mostly found in the sandy and muddy flats, or in mangrove area and seagrass area. This animal prefers to choose habitat which it substrate is rich in organic materials.
Distribution: recent Anadara distribution is coastal areas of Australia, Tropical Indo-West Pacific, Red Sea, South China Sea, Vietnam, China, Hongkong, Thailand, Philippines, New Caledonia, Japan and Indonesia.
(a) (b)
FIGURE 3. Fossil of bivalve shell genera Anadara (S.5360) discovered in Sangiran: (a) is an umbilical view of fossil shell; (b) is a dorsal view of fossil shell (Photo: Yudha, 2015).
Description: A. strong and thick radial ribs; B. thick and wrinkled ribs.
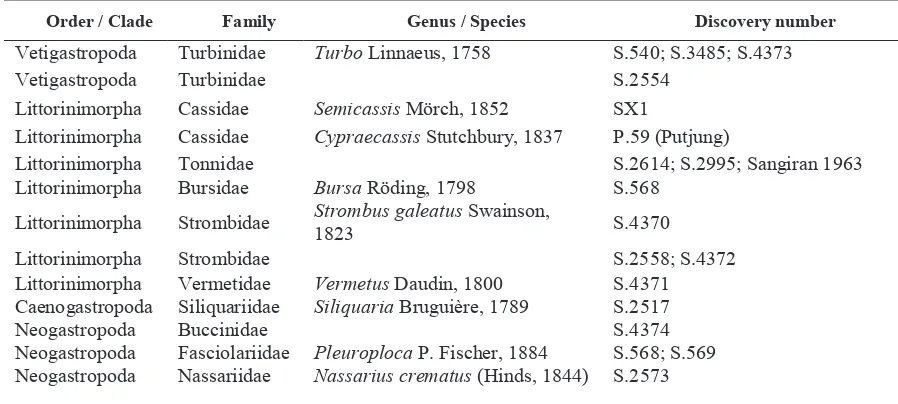
Of twenty-nine gastropods examine fossil shells of gastropods examined, all shells can be determined until taxa level family, 18 shells can determine until taxa level genus and 1 shell is determined by species level. Total taxa of gastropods fossil are four orders with 13 families. Of eighteen fossil shells identified to genus are belong to 13 different genera, i.e. Turbo (Turbinidae), Cypraecassis and Semicassis (Cassidae), Bursa (Bursidae), Strombus (Strombidae), Vermetus (Vermetidae), Siliquaria (Siliquariidae), Pleuroploca (Fasciolariidae), Marginella and Persicula (Marginellidae), Murex (Muricidae), Zeuxis (Nassariidae), and Neptuneopsis (Volutidae) (Table 2).
TABLE 2. The list of Gastropod Fossil shells discovered in the Sangiran area, Central Java.
Order / Clade Family Genus / Species Discovery number
Vetigastropoda Turbinidae Turbo Linnaeus, 1758 S.540; S.3485; S.4373 Vetigastropoda Turbinidae S.2554
Littorinimorpha Cassidae Semicassis Mörch, 1852 SX1
Littorinimorpha Cassidae Cypraecassis Stutchbury, 1837 P.59 (Putjung)
Littorinimorpha Tonnidae S.2614; S.2995; Sangiran 1963 Littorinimorpha Bursidae Bursa Röding, 1798 S.568
Littorinimorpha Strombidae Strombus galeatus Swainson, 1823 S.4370 Littorinimorpha Strombidae S.2558; S.4372 Littorinimorpha Vermetidae Vermetus Daudin, 1800 S.4371 Caenogastropoda Siliquariidae Siliquaria Bruguière, 1789 S.2517 Neogastropoda Buccinidae S.4374 Neogastropoda Fasciolariidae Pleuroploca P. Fischer, 1884 S.568; S.569 Neogastropoda Nassariidae Nassarius crematus (Hinds, 1844) S.2573
Table 2. Continued
Order / Clade Family Genus / Species Discovery number
Neogastropoda Nassariidae 4 individuals Neogastropoda Marginellidae Marginella Lamarck, 1799 SX3; S.791 Neogastropoda Marginellidae Persicula Schumacher, 1817 SX4
Neogastropoda Muricidae Murex trapa Röding, 1798 S.4393; Tgl 10–12–70
Neogastropoda Volutidae Neptuneopsis Sowerby III, 1898 S.564
Class Gastropoda Subclass Vetigastropoda Superfamily Trochoidea
Family Turbinidae Genus Turbo Linnaeus, 1758
Material examined: there are three fossils cast of internal shell remains which its diagnostic characters only a little. Discovery number: S.540; S.3485; and S.4373.
Diagnostic characters: shown on the fossil shell are Shell heavy (11 to 15) gr, thick (0.3 to 0.45) mm, turbinate. Aperture rounded, anterior canal lacking.
Habitat: recent Turbo habitat mainly on rocky and coral reef in shallow waters of warm temperate and tropical sea [18].
Distribution: Widespread in the Indo-West Pacific; and all of the Indonesian seas[18].
Subclass Caenogastropoda Order Littorinimorpha Superfamily Tonnoidea
Family Cassidae
There is two identified genus of Cassidae’s fossil from this collection, i.e.; Semicassis and Cypraecassis. The
Semicassis fossil type is an external mold, which means the outer shell details is preserved/imprinted, while the Cypraecassis fossil type is a cast, which means sediment filled in the external shell or the mold fossil, and after that, the external shell or mold fossil is broken leave only the three-dimensional form of internal shell.
Genus Semicassis Mörch, 1852
Syn. Xenogalea Iredale, 1927
Material examined: is external shell fossil remains which its diagnostic characters on its shell morphology. Discovery number: S X 1.
Diagnostic characters: shown on the fossil shell are: shell is egg-shaped, transversely obscurely sulcate (grooved). The shell has a large aperture, with outer lip thick and has a series of strong lirae, giving it a toothed appearance. The body whorl is sculptured by spiral grooves crossed by mostly rather faint growth lines. The tail or siphonal canal is recurved (Fig. 4).
Habitat: recent Semicassis habitat mainly on the sandy substrate in moderately shallow water, and also on rocky tide pools close to the shore at depths approximately 15 m to 46 m.
Genus Cypraecassis Stutchbury, 1837
Syn. Cassis Scopoli, 1771
Material examined: is a cast of internal shell remains which its diagnostic characters only a little. Discovery number: P. 59.
Habitat: Recent Cypraecassis habitat is on the sandy substrates close to coral reefs in the shallow waters at depths of about 0 m to 60 m.
Distribution: Indo-Pacific.
(a) (b)
FIGURE 4. Image number (1) is Semicassis external shell, and number (2) is a cast of internal shell of Cypraecassis; (a) is an image of whorl part of fossil; (b) is an image of aperture (Photo: Yudha, 2015). Description: A. internal part of the apex; B.
internal part of anterior siphonal canal; C. apex; D. anterior siphonal canal; E. internal part of outer lip; F. outer lip.
Order Littorinimorpha Superfamily Tonnoidae
Family Bursidae Genus Bursa Röding, 1798
Material examined: is external shell fossil remains which its diagnostic characters on its shell morphology. Discovery number: S.568.
Diagnostic characters: Shell ovate to slightly elongate and dorsoventrally compressed; two strong axial varices every 180°, then aligned up sides of spire. Shell thick, with coarse knobbed sculpture; nucleus of the operculum near the anterior end. Aperture with a short anterior siphonal canal. Outer lip thick and denticulate inside, inner lip more or less calloused, with transverse ridges or granules
Habitat: recent Bursa mainly occurs on rocks or coral reefs in shallow waters, low tide levels and shallow sublittoral zone to about 20 m, only a few are found on sandy bottoms of deeper waters of the continental shelf.
Distribution: Tropical oceans, including the Indo-Pacific, the Caribbean Sea and other marginal warm seas, they are also found in the Atlantic Ocean and the Mediterranean Sea.
Order Littorinimorpha Superfamily Stromboidea
Family Strombidae
Species original name: Strombus galeatus Swainson, 1823
Species accepted name: Lobatus galeatus (Swainson, 1823)
Material examined: a fossil cast of internal shell remains which its diagnostic characters only showed a little. Discovery number: S.4370.
Diagnostic characters: Shell thick and solid, with a low spire and a very large body whorl. Aperture elongated, with a well-marked siphonal canal. The inner part of thickened and flaring outer lip is shown.
Habitat: recent Strombus galeatus mainly living in shallow water, on sandy, muddy or rubble bottoms or marine grassflats[18].
Distribution: Tropical and subtropical waters, widespread in the Indo-West Pacific, also occur in the Eastern Pacific and Western Atlantic.
Order Littorinimorpha Superfamily Vermetoidea
Material examined: is external shell fossil remains which its diagnostic characters are shown on its shell morphology. Discovery number: S.4371.
Diagnostic characters: Shell long, irregularly coiled resembling a worm tube. On the shell remains there is a trace of attachment to a hard substrate. The sculpture is weak, longitudinal and transverse. Trace of rounded aperture, sharp-edged, without a siphonal canal.
Habitat: recent Vermetus are abundant in the intertidal zone of warm-temperate to tropical seas. Adults are living attached to hard substrates of shallow reefs (rocks, corals and other shells).
Distribution: recent Vermetus distribution had not been reviewed yet.
Subclass Caenogastropoda
Material examined: a fossil cast of internal shell remains which its diagnostic characters only showed a little. Discovery number: S.2517.
Diagnostic characters: early whorls of the shell are spirally coiled with four spirals, and protoconch is missing.
Habitat: recent Siliquaria is fixed to the substrate, embedded in sponges between intertidal and subtidal zones.
Distribution: recent Siliquaria commonly found in Indo-West Pacific zones.
Subclass Caenogastropoda Order Neogastropoda Superfamily Buccinoidea
Family Fasciolariidae Genus Pleuroploca P. Fischer, 1884
Material examined: two fossils of external shell fossil remains which its diagnostic characters are shown on their shell morphology. Discovery number: S.568 and S.569.
Diagnostic characters: Shell is thick and heavy. Shell shape is broadly fusiform with a high conical spire and stout. Siphonal canal is moderately straight long and broadly open. Whorls angulate at shoulders, with a row of thick and prominent axial nodules, most pronounced on the last two whorls. The body whorl is swollen, subangulate at the
base. Aperture’s shape is roughly ovate, inside the outer lip, there are finely marking lirate which has sharp paired
denticles at the margin. Columella with few (about 3) weak oblique folds anteriorly.
Habitat: recent Pleuroploca mainly found on sand and rubble bottoms, on inner reef flats in shallow water near the rocky area. Low tide levels to a depth of about 40 m.
Distribution: recent Pleuroploca distributed Widespread in the Indo-West Pacific, from East
Africa, including the Red Sea and the Persian Gulf, to Melanesia; north to Japan, and south to northern Queensland and New Caledonia. Apparently rare in Australia [18].
Superfamily Buccinoidea Family Nasariidae
Species Nassarius crematus (Hinds, 1844)
Original Name: Nassa cremata Hinds, 1844; Syn. Nassarius (Zeuxis) crematus (Hinds, 1844)
Material examined: is external shell fossil remains which its diagnostic characters are shown on its shell morphology. Discovery number: S.2573.
Habitat: recent Nassarius cremata is on fine sandy or muddy bottoms, in the intertidal, sublittoral and shelf zones to a depth of about 200 m[18].
Distribution: recent Nassarius cremata is widespread in the Indo-West Pacific, from East Africa to Hawaii; north to Japan and south to central Queensland[18].
(a) (b)
FIGURE 5. Fossil of gastropod shell Nassarius crematus (S.2573) discovered in Sangiran:
(a) is whorl and body whorl part; (b) is an aperture part (Photo: Yudha, 2015).
Order Neogastropoda Superfamily Muricoidea
Family Marginellidae
There is two identified genus of Marginellidae’s fossil from this collection, i.e.; Marginella and Persicula. Both
specimens fossil type are an external mold, which means the outer shell details is preserved.
Genus Marginella Lamarck, 1799
Syn. Marginellarius Duméril, 1805; Porcellana Gray, 1847
Material examined: two fossils of external shell fossil remains which its diagnostic characters are shown on its shell morphology. Discovery number: S.791 and S X 3.
Diagnostic characters: Shell minute, elongate-ovate, broader posteriorly, shell color is white, polished without sculpture. Spire is very low, outer lip nearly straight, on the last whorl thickened by callus along its outer edge, most strongly thickened on posterior portion.
Habitat: recent Marginella is in gravel or mixed sediments around rocky areas, at 10 m to 100 m [18].
Distribution: recent Marginella is commonly found in tropical and temperate seas [18].
Genus Persicula Schumacher, 1817
Material examined: is external shell fossil remains which its diagnostic characters are shown on its shell morphology. Discovery number: S X 4.
Diagnostic characters: shell white and small, outer lip are thickened, the flat spire immersed in a pad of enamel and there is pattern of callousing on the parietal wall.
Habitat: recent Persicula live from the intertidal zone to 370 m in depth.
Distribution: recent Persicula is widespread in the tropical West Pacific.
Superfamily Muricoidea Family Muricidae Species Murex trapa Röding, 1798
Material examined: two fossils of external shell fossil remains which its diagnostic characters are shown on its
shell morphology. Discovery number: S.4393 and “Tgl 10–12–70”.
margin of the outer lip with a prominent tooth-like process; Three to 4 short spines, restricted to the basal half of siphonal canal.
Habitat: recent Murex trapa habitat is commonly on muddy-sand bottom, low tide and shallow sublittoral depths.
Distribution: recent Murex is found Widespread in the Indo-West Pacific, from Madagascar, India, Sri Lanka and the Andaman Sea, to the Philippines; north to southern Japan and south to southern Indonesia.
(a) (b)
FIGURE 6. Fossil of gastropod shell Murex trapa(S.4393 and “Tgl 10–12–70”) discovered in Sangiran:
(a) is whorl and body whorl part; (b) is an aperture part (Photo: Yudha, 2015).
Superfamily Muricoidea Family Volutidae
Genus Neptuneopsis Sowerby III, 1898
Material examined: is external shell fossil remains which its diagnostic characters are shown on its shell morphology. Discovery number: S.564.
Diagnostic characters: Shell shape fusiform with a sunken spire; protoconch large, bulbous, with a pronounced calcarella; the surface of the shell is smooth, glossy and brightly coloured. Aperture of fossil is fragmented, but in living shell, aperture long, with a short and wide siphonal canal anteriorly. Inner lip with strong, oblique folds, the weaker ones situated posteriorly.
Habitat: recent Neptuneopsis mainly found on sandy or muddy bottoms, from intertidal flats to the continental shelf and slope.
Distribution: recent Neptuneopsis is not yet reviewed.
Based on shell’s morphological characters, therefore, bivalves’ fossils that can be identified until taxa that level
genus are Hippopus, Dosinia and Anadara. Those three genera still have its member whose live nowadays (Holocene time) and all of their inhabit shallow seas on the intertidal/littoral zones. Also based on the shell’s morphological characters, gastropods’ fossils that can be identified by taxa level genus are Turbo, Cypraecassis,
Semicassis, Bursa, Strombus, Vermetus, Siliquaria, Pleuroploca, Marginella, Persicula, Murex, Nassarius and Neptuneopsis. Those thirteen genuses also have the same members who live in a recent time (Holocene). All of those gastropods inhabit shallow seas on the intertidal/littoral zones.
The time-range from Pleistocene to Holocene is a short period in term of geological time scale, i.e., 2.6 × 106 yr.
A short geological period is also a short time for organisms experiencing macroevolution. Therefore, the Pleistocene-Holocene time-range has not affected the change of faunal morphology, faunal compositions and its habitat in term of biological evolution. Invertebrates fauna evolutive are not undergone morphological changes in a short period, they underwent morphological changes in a long period, for example about 100 × 106 yr, faunal
example: trilobites [20, 21, 22]. It can be concluded that all bivalves and gastropods which lived in Pleistocene of Sangiran have the same habitat with recent (Holocene) bivalves of the same genus.
seacoasts. Sémah, et al., [23] said that Java was emerged from the sea as early as the Gauss-Matuyama boundary, around 2.5 × 106 yr ago and shallow marine zone were in Sangiran area during the Late Pliocene to Lower
Pleistocene. Sémah, et al., [24]; Sémah & Sémah [7] studies mention that Java Island underwent several sea-level changes linked to the glacial-interglacial cycles, tectonic uplifts and volcanic activities and at that time, around Sangiran area were shallow lagoon and coastal swamps with immense mangrove forest near the shoreline.
CONCLUSION
The marine mollusc fossils collection of the Laboratory of Bioanthropology and Paleoanthropology, Faculty of Medicine, UGM is composed of two classes, i.e., Bivalvia and Gastropoda. There are six orders with 12 families and three genuses from the class Bivalvia. There are four orders with 14 families and 13 genuses from the class Gastropoda. All members of the bivalves and gastropods that have been identified by level genus are molluscs which inhabit shallow sea around intertidal zones. Those are identical with references about the geological study of Sangiran area during Pleistocene time. In the Lower Pleistocene time, some areas of Sangiran were shallow sea and estuary.
ACKNOWLEDGEMENTS
We are extremely grateful to the Head of the Laboratory of Bioanthropology and Paleoanthropology, Faculty of Medicine, UGM for permission to analyze samples of mollusc fossil and to work comfortably in the laboratory. We also thanks to Sugiyo, a laboratory staff who help us prepare the samples and other need for analysis.
REFERENCES
1. Di Geronimo and S. Sartono. Sangiran (Java, Indonesia): Upper Pliocene – Pleistocene molluscan
environments (Bul. Jurusan Geologi, Bandung, 1990), pp. 33–35.
2. M. Fae, “La malacofauna quaternaria dei depositi con resti umani dell’area di Sangiran in Giava central [Quartenary malacofauna in the human remains deposit area of Sangiran],” Unpublished graduation work,
Padova University, 1994. [Italiano]
3. M. Fae, Memorie di Scienze Geologiche 48, 143–153 (1996).
4. T. Simanjuntak, “Sangiran site: problems and the balance of research,” in Sangiran: Man, Culture, and Environment in Pleistocene Times, edited by T. Simanjuntak et al. (Yayasan Obor Indonesia, Jakarta. 2001), pp. 1–15.
5. S. Sartono, “On the Stratigraphic Position of Pithecanthropus Mandible” in C. Proceedings vol. 4, No. 4,
(Institut Teknologi Bandung, Bandung, 1970), pp. 91–102.
6. F. Sémah, H. Saleki, C. Falguères, G. Féraud and T. Djubiantono, Journal of Archaeological Science27, 763–
769 (2000).
7. A-M. Sémah and F. Sémah, Quaternary International249, 120–128 (2012).
8. L. R. Lyman, Journal of Taphonomy 8(1), pp.2–4(2010).
9. A. K. Behrensmeyer, S. M. Kidwell and R.A. Gastaldo, Paleobiology26(4), 103–147 (2000).
10. P. A. Allison and D. J. Bottjer, Taphonomy: Process and Bias Through Time. 1 Topics in Geobiology 32
(Springer Science+Business Media B.V, USA, 2011), pp. 2–4. DOI 10.1007/978-90-481-8643-3_1
11. O. Macsotay and R.C. Hernandez, Caribbean Journal of Earth Science 39, 93–104 (2005).
12. J. E. Rogers, The Shell Book: A Popular Guide To A Knowledge of The Families of Living Mollusks, and An Aid To The Identification of Shells Native and Foreign (Charles T. Branford co., Publishers, USA, 1936), pp. 27, 79, 111, 117, 305.
13. T. Kira, Shells Of The Western Pacific In Color, Vol 1 (Hoikusha Publishing, Co., Ltd., Japan. 1962), pp. 16–
19, 24–27, 34–39, 59–61, 63–65, 72–78, 80–82, 82–87, 92–93.
14. T. Habe, Shells Of The Western Pacific In Color, Vol 2 (Hoikusha Publishing, Co., Ltd., Japan. 1964), pp. 19–
21, 35–38, 57–61, 67–70, 75–77.
15. F. O. Huffman, “Plio-Pleistocene environmental variety in eastern Java and early Homo erectus paleoecology –
a geological perspective” in Sangiran: Man, Culture, and Environment in Pleistocene Times edited by T.
16. F. Sémah, A-M Sémah and T. Djubiantono, “From the shoreline to the slopes of the volcanoes: the long
Pithecanthropus trek,” in Sangiran: Man, Culture, and Environment in Pleistocene Times, edited by T.
Simanjuntak et al. (Yayasan Obor Indonesia, Jakarta, 2001), pp. 195–218.
17. A-M. Sémah, F. Sémah, T. Djubiantono and B. Brasseur, Quaternary International223–224, 451–454 (2010).
18. K. E. Carpenter and V. H. Niem, FAO species identification guide for fishery purposes.The living marine resources of the Western Central Pacific. Volume 1. Seaweeds, corals, bivalves and gastropods. (FAO, Rome, 1998), pp. 1–686.
19. U. E. Hernawan, Biodiversitas13(3), 118–123 (2012).
20. S. J. Gould, “Tempo and mode in the macroevolutionary reconstruction of Darwinism,” in Proc. Nadl. Acad.
Sci. USA Vol. 91, edited by I. M. Verma et al. (Colloquium Paper, USA, 1994), pp. 6764–6771.
21. J. Pojeta Jr. and D. A. Springer, Evolution and the Fossil Record (American Geological Institute The Paleontological Society, American Geological Institute Alexandria, Virginia. 2001), pp. 1–26.
22. U. Kutschera and K. J. Niklas, Naturwissenschaften 91, 255–276 (2004).
23. F. Sémah, A-M Sémah and T. Djubiantono, “From the shoreline to the slopes of the volcanoes: the long
Pithecanthropus trek,” in Sangiran: Man, Culture, and Environment in Pleistocene Times, edited by T.
Simanjuntak et al. (Yayasan Obor Indonesia, Jakarta, 2001), pp. 195–218.