GENETIKA IKAN
INBREEDING
(PERKAWINAN SEDARAH) Disusun oleh : Kelompok 9 Kelas B Ruli Aisyah 230110140091 Ristiana Dewi 230110140099Darajat Prasetya Wintantra 230110140098 Adinda Kinasih Jacinda 230110140108 Ahmad Abdul Gofur 230110140120
Egi Ramadhan 230110140125
JURUSAN PERIKANAN
FAKULTAS PERIKANAN DAN ILMU KELAUTAN UNIVERSITAS PADJADJARAN
JATINANGOR
KATA PENGANTAR
Puji dan syukur kami panjatkan kehadirat Allah Swt yang telah begitu banyak melimpahkan karunia dan rahmat-Nya sehingga kami dapat menyelesaikan makalah ini tepat pada waktunya tanpa tantangan yang berarti, Shalawat teriring salam semoga tetap terlimpah curah kepada panutan alam yakni Nabi besar Muhammad SAW, kepada para keluarganya, para sahabatnya sampai kepada kita semua selaku umatnya hingga akhir zaman.
Alhamdulillah dengan segala keterbatasan makalah Inbreeding yang merupakan salah satu penunjang mata kuliah Genetika Ikan dapat kami selesaikan, semoga dengan segal keterbatasan ini mampu menjadi acuan atau panduan untuk lebih mendalami mata kuliah Genetika Ikan khususnya dalam materi Inbreeding.
Dalam makalah ini masih terdapat begitu banyak kekurangan karena pengetahuan kami mengenai materinya pun masih belum terlalu jauh serta keterbatasan sumber. Oleh karena itu segala bentuk kritik dan saran yang sifatnya membangun untuk kesempurnaan makalah ini sangat kami harapkan.
Jatinangor, 15 Oktober 2015 Penyusun
DAFTAR ISI KATA PENGANTAR ... i DAFTAR ISI ... ii BAB I PENDAHULUAN ... 1 1.1 Latar Belakang ... 1 1.2 Tujuan ... 1 BAB II PEMBAHASAN ... 2 2.1 Pengertian Inbreeding ... 2 2.2 Kegunaan Inbreeding ... 6 2.3 Perhitungan Inbreeding ... 8
2.4 Efek ukuran populasi dalam perkawinan sedarah dan Genetik Drift ... 13
2.5 Mencegah Penurunan Dalam Angka Efektif Breeding ... 20
BAB III PENUTUP ... 39
3.1 Kesimpulan ... 39
3.2 Saran ... 39
BAB I PENDAHULUAN
1.1 Latar Belakang
Di dalam akuakultur inbreeding di dikenal dengan istilah perkawinan individu yang memiliki kekerabatan dekat. Dalam genetika, Inbreeding bertujuan untuk mendapatkan hewan yang homozygot (yang unggul dalam salah satu sifat, misalnya mempunyai kemampuan yang tinggi dalam kenaikan berat badan dan sifat ini menurun). Dalam Inbreeding frekuensi gen, seleksi, genetif drift tidak mengalami perubahan. Sehingga untuk menghasilkan benih yang baik dan unggul dilakukan seleksi atau perkawinan silang.
Inbreeding memiliki dampak positif dan negative dalam akuakultur. Dampak positif dari inbreeding adalah diperoleh benih yang unggul, ketahanan terhadap penyakit, dan efisiensi pakan yang baik. sedangkan dampak negatif dari inbreeding yaitu jika terjadi secara tidak terkendali dan tanpa dilandasi pengetahuan yang baik adalah munculnya sifat – sifat merugikan yang sebelumnya tertutup sifat dominan (resesif).permasalahan dari inbreeding yang langsung dirasakan adalah menurunnya kualitas benih. Dalam makalah ini akan diuraikan lebih jauh lagi mengenai inbreeding dan segala aspek yang terkait didalamnya.
1.2 Tujuan
Adapun tujuan dari pembuatan makalah ini antara lain : 1. Mengetahui apa itu inbreeding.
2. Mengetahui kegunaan Inbreeding. 3. Mengetahui perhitungan inbreeding.
BAB II PEMBAHASAN
2.1 Pengertian Inbreeding
Perkawinan sekerabat (inbreeding) adalah program pemulian utama ketiga yang dapat berdampak pada produktivitas. Inbreeding merupakan salah satu konsep yang diketahui oleh setiap orang tapi hanya beberapa orang saja yang benar-benar memahaminya, istilah ini biasanya memunculkan kesan individu yang cacat dan buruk, dan istilah ini digunakan sebagai gurauan untuk menjelaskan hakikat sebenarnya dari semua perilaku yang cacat, meskipun perkawinan sedarah biasanya tidak ada hubungannya dengan masalah ini. kebanyakan orang sadar tentang perkawinan sedarah karena hukum dan moral hukum yang menentang perkawinan antar kerabat. 200 tahun yang lalu, ada hukum yang melarang perkawinan antar kerabat bahkan pada ternak karena dianggap tidak bermoral dan melawan hukum tuhan dan alam. Tetapi peternak dengan cepat menemukan bahwa inbreeding adalah salah satu teknik pemuliaan yang paling penting, tanpa penggunaannya produktivitas pertanian akan menurun drastis.
Perkawinan sekerabat merupakan perkawinan antara individu yang masih memiliki keterkaitan (kurang lebih), Inbreeding tidak terlihat secara jelas, dan tidak dapat dijelaskan. Apapun mengenai kelayakan, pertumbuhan, atau produktivitas. Inbreeding bukan mengenai baik dan buruk, itu bisa, namun seperti program pemuliaan lainnya, dapat juga digunakan secara bijaksana atau serakah.
Secara genetika, semua pemuliaan menciptakan homozigositas. Alel yang terkait dalam individu tersebut melalui satu atau lebih nenek moyang yang sama. Ketika individu yang bersangkutan melakukan perkawinan, alel yang dihasilkan dari nenek moyang mereka umumnya dapat dipasangkan. Ini menghasilkan keturunan yang homozigot pada satu lokus lagi, dan keturunan tersebut merupakan bawaan. Perkawinan pada individu yang tidak ada keterkaitan juga bisa menghasilkan keturunan yang homozigot pada satu atau lebih lokus. Bagaimana kita bisa membedakan homozigot yang dihasilkan oleh inbreeding dan
perkawinan yang bukan inbreeding? Pada umumnya, bagaimana bentuk dari dua homozigot ini berbeda? Jawabannya adalah 1. Kita tidak bisa membedakan keduanya. 2. Umumnya, tidak ada perbedaan mereka sama, satu-satunya perbedaan yaitu binatang yang bersifat homozigot karena memiliki alel yang sama dengan keturunan sebelumnya. Sedangkan non inbred individu homozigot karena mereka telah alel yang sama dalam bentuk. Tidak ada cara kimiawi atau fisika untuk membedakan antara keduanya. Tidak ada cara lain dalam alel .
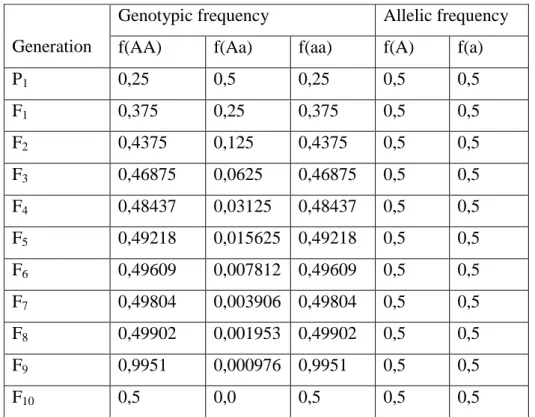
Tabel 4.8 efek perkawinan sekerabat pada Frekuensi Genotip dan Frekuensi Alel pada Lokus.
Berikut terjadinya perkawinan pada setiap generasi: AA x AA ; Aa x Aa ; aa x aa.
Karena perbedaan genotipe meningkat, perbedaan fenotip juga bertambah. Perbedaan genotipe dan fenotipe meningkat karena populasinya terbagi menjadi garis homozigot terpisah dan itu merubah distribusi normal menjadi distribusibimodal (gambar 4.16). Jika garis homozigot terpisah menjadi populasi yang berbeda, perbedaan genotipe dan fenotipe akan berkurang. Jika hal tersebut terjadi frekuensi gen dan perbedaan genetik akan mengalami perubahan juga.
Generation
Genotypic frequency Allelic frequency
f(AA) f(Aa) f(aa) f(A) f(a)
P1 0,25 0,5 0,25 0,5 0,5 F1 0,375 0,25 0,375 0,5 0,5 F2 0,4375 0,125 0,4375 0,5 0,5 F3 0,46875 0,0625 0,46875 0,5 0,5 F4 0,48437 0,03125 0,48437 0,5 0,5 F5 0,49218 0,015625 0,49218 0,5 0,5 F6 0,49609 0,007812 0,49609 0,5 0,5 F7 0,49804 0,003906 0,49804 0,5 0,5 F8 0,49902 0,001953 0,49902 0,5 0,5 F9 0,9951 0,000976 0,9951 0,5 0,5 F10 0,5 0,0 0,5 0,5 0,5
Mean (B) Mean
Mean Mean (C)
Gambar. 4.16 inbreeding bisa dirubah menjadi dristribusi yang normal (A) kedalam kurva bimodal (B) presentasi inbreeding meningkatkan homozigot setiap individu. Peningkatan perbedaan phenotip ini jika inbreeding berlangsung terus-menerus dan populasinya terpisah menjadi dua garis (C), perbedaan dalam setiap garis tiba-tiba berkurang.
Jadi apa yang telah diakibatkan oleh inbreeding sehingga mendapat stigma negatif? Hampir setiap organisme membawa alel-alel resesif merugikan yang tersembunyi dalam bentuk heterozigot. Jika alel-alel ini muncul, mereka akan menghasilkan abnormal atau letal fenotip. Individu-individu yang berhubungan kemungkinan memiliki alel-alel resesif merugikan yang sama. Karena perkawinan sekerabat menghasilkan ke-homozigot-an dari alel-alel berpasangan yang sama dari keturunan, sebagian alel-alel resesif yang merusak memiliki kemiripan yang lebih dan muncul pada individu yang berhubungan daripada individu-individu yang tidak berhubungan. Kemungkinan meningkatnya alel resesif yang merugikan disebabkan hubungan antara induk yang meningkat. Hal inilah yang menyebabkan reputasi buruk pada proses inbreeding.
Tidak ada kepastian bahwa keturunan perkawinan sekerabat akan memiliki kelainan. Namun, kemungkinan hasil keturunan yang abnormal meningkat ketika induk memiliki hubungan, dan semakin dekat hubungannya maka semakin besar kemungkinannya.
Pasangan alel resesif yang merugikan menghasilkan kecenderungan umum terhadap penurunan kelangsungan hidup, pertahanan hidup, pertumbuhan, produksi telur dan secara bersamaan meningkatkan persentase kelainan. Umumnya, semakin besar perkawinan sekerabat, depresi pada produktifitas semakin berat.
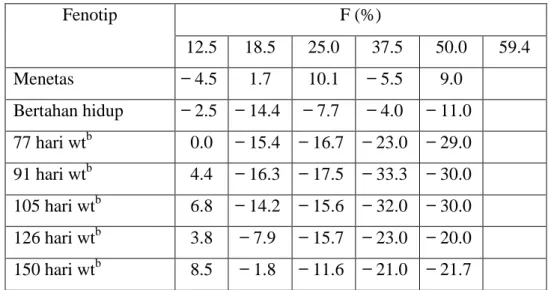
Sudah ada penelitian tentang inbreeding ikan yang relatif sedikit. Beberapa yang telah dilakukan diantaranya di Rainbow Trout (Calaprice 1969, Aulstad dan Kittelsen 1971; Bridges 1973, Kincaid 1976A, 1976B, 1983B, Davis 1976;. Gjerde et al 1983), Salmon Atlantik (Ryman 1970), brook trout (Cooper 1961; Davis 1976), brown trout (Davis 1976), common carp (Moav dan wohlfarth 1968), T.mossambica (Ch'ang 1971a), channel catfish (Bondari 1984a), zebra danio (Piron 1978, Mracovčić dan Haley 1979), dan convict cichlids (Winemiller dan Taylor 1982). Dengan beberapa pengecualian, studi menunjukkan kecenderungan yang sama, seperti produksi pertumbuhan fenotipe perkawinan sekerabat tertekan, kelangsungan hidup, dan meningkatkan jumlah kelainan. Kincaid telah melakukan studi yang paling rinci, dan beberapa hasil nya ditunjukkan pada Tabel 4.9. Rayman (1970) menemukan bahwa perkawinan sekerabat salmon Atlantik memiliki tingkat pengembalian secara signifikan lebih rendah ketika ditebar di perairan alami. Studinya menunjukkan bahwa angka kematian post hatchery, sebagai produk sampingan dari perkawinan silang, mungkin menjadi salah satu alasan mengapa sulit untuk memperoleh kembali sumber daya benih ikan diproduksi.
Tabel 4.9 Depresi Inbreeding pada Rainbow Trout
Fenotip F (%) 12.5 18.5 25.0 37.5 50.0 59.4 Menetas 4.5 1.7 10.1 5.5 9.0 Bertahan hidup 2.5 14.4 7.7 4.0 11.0 77 hari wtb 0.0 15.4 16.7 23.0 29.0 91 hari wtb 4.4 16.3 17.5 33.3 30.0 105 hari wtb 6.8 14.2 15.6 32.0 30.0 126 hari wtb 3.8 7.9 15.7 23.0 20.0 150 hari wtb 8.5 1.8 11.6 21.0 21.7
1 tahun wt, ♂ 23.3 16.4 12.8 34.8 1 tahun wt, ♀ 20.9 27.2 21.9 41.8 2 tahun wt, ♂ 26.2 32.2 33.7 41.8 2 tahun wt, ♀ 18.0 38.1 28.8 51.2 Jangka 2 tahun, ♂ 9.2 13.0 15.2 19.0 Jangka 2 tahun, ♀ 9.1 16.8 13.9 21.8 Massa telur 18.1 33.9 40.3 57.0 Persentase yang cacat 2.9 9.2
Depresi Inbreeding dinyatakan sebagai persen depresi bila dibandingkan dengan populasi kontrol (F = 0,0). Sebuah nilai positif berarti bahwa kelompok inbrida adalah lebih baik daripada control.
Kincaid (1976A) Kincaid (1983B) Kincaid (1976B)
Penelitian umumnya didasarkan pada tingkat inbreeding di kisaran 25-60, meskipun Kincaid (1976A) memandang F = 12,5%. Studi masa depan harus memeriksa tingkat yang lebih rendah dari inbreeding, karena tingkat yang lebih rendah benar-benar dapat mempertinggi produktivitas. Beberapa hasil Kinchaid (1976A) ini (Tabel 4.9) menunjukkan bahwa hal ini mungkin benar di rainbow trout sekitar 18%, di bawah 18%, perkawinan sedarah menghasilkan beberapa masalah, tetapi di atas 18%, produktivitas tertekan secara signifikan.
2.2 Kegunaan Inbreeding
Perkawinan sekerabat yang dipelajari di ikan secara umum menunjukkan bahwa perkawinan sekerabat mengurangi produktivitas, jadi bagaimana bisa ini pernah digunakan untuk meningkatkan populasi? Salah satu penggunaan utama perkawinan sekerabat adalah dalam sebuah program perkembangbiakan yang disebut linebreeding. Linebreeding terjadi ketika individu yang menonjol bagus (biasanya laki-laki) dibawa kembali ke garis untuk kawin dengan keturunan. Ini dilakukan karena hewan yang luar biasa ini ingin Anda tingkatkan kontribusinya
untuk masing-masing keturunan dan meningkatkan kontribusinya terhadap gen yang jelek. Dua jenis linebreeding yang ditampilkan pada gambar 4.17.
4.17. diagram skema dari dua tipe linebreeding. Tujuan dari keduanya adalah untuk meningkatkan kontribusi dari individu A untuk genotif keturunan. Sebagai contoh Mild breeding individu A berkontribusi 53,12% kepada gen K. Dan contoh intens linebreeding, individu A berkontribusi 93,75% kepada gen G.
Penggunaan utama yang kedua dari perkawinan sekerabat adalah untuk menciptakan inbreadlines yang akan meng-hibridisasi untuk menghasilkan hibrida F1 untuk tumbuh. Di sini, dipilih dua atau lebih garis inbread untuk memperbaiki alel tertentu. Ketika jalur inbread kawin, hibrida akan sama dan akan seragam, dimana itu merupakan salah satu tujuan dalam cross breeding program. Perkawinan sekerabat dalam dua atau lebih baris yang diikuti oleh hibridisasi adalah cara klasik dalam menghasilkan keturunan yang sama untuk tumbuh.
Perkawinan sekerabat juga digunakan oleh banyak peneliti untuk menghasilkan hewan yang akan digunakan dalam berbagai eksperimen. Dalam beberapa kasus, para ilmuwan tidak menyadari bahwa mereka menimbulkan perkawinan sekerabat, tidak mempertimbangkan efek perkawinan sekerabat terhadap ternaknya, dan tidak menyadari bahwa perkawinan sekerabat mungkin mengacaukan variabel yang mereka periksa dalam percobaan mereka. Konsekuensi alami dari perkawinan sekerabat kadang-kadang memberi kejutan kepada peneliti, lalu mereka menarik kesimpulan yang salah. Salah satu contohnya adalah sebuah studi yang dilakukan oleh Piron (1978). Ia menghasilkan
perkawinan sekerabat tingkat tinggi kepada zebra danio yang digunakan dalam toksisitas tes. Perkawinan sekerabat menghasilkan beberapa ikan dengan kelainan bentuk kerangka, dan hal ini menyebabkan Piron (1978) menyimpulkan bahwa spesies ini tidak cocok untuk digunakan dalam toksisitas tes. Dihasilkannya kelainan sebagai hasil dari perkawinan sekerabat adalah bukan alasan yang sah untuk mencapai kesimpulan akhir ini.
Sampai semua alel resesif yang merugikan diambil, hampir setiap populasi akan menghasilkan beberapa kelainan sebagai hasil dari perkawinan sekerabat. Sebagai soal fakta, banyak spesies yang digunakan untuk penelitian biomedis melakukan perkawinan sekerabat tingkat tinggi untuk menghasilkan populasi homozigot sehingga semua binatang akan bereaksi dengan cara yang sama untuk variabel eksperimental. Produksi populasi perkawinan sekerabat untuk penelitian tersebut meminimalkan variasi individu, dimana merupakan bagian penting dari total varians dalam percobaan.
Bahkan ketika digunakan secara tidak benar dan ketika perkawinan sekerabat mengalami depresi, perkawinan sekerabat masih dapat menghasilkan keturunan yang bagus. Depresi yang terlihat untuk berbagai fenotipe berarti populasi. Individu yang unggul dapat diproduksi meskipun populasi rata-rata mungkin menurun. Hewan inbrida yang beredar dapat dinilai sebagai induk karena mereka berkembang biak untuk banyak fenotipe dan tidak akan menunjang fenotipe tidak diinginkan.
Genetika inbeeding hampir sama dengan crooss breeding. keduanya bergantung pada interaksi alel. Perkawinan sekerabat menekan produksi oleh pasangan determental alel resesif. Dengan demikian, perkawinan sekerabat adalah pada dasarnya fungsi VD.
2.3 Perhitungan Inbreeding
Nilai-nilai individu Inbreeding dapat dihitung dengan menggunakan teknik yang disebut analisis jalur. Dalam analisis jalur, anda mengubah sebuah silsilah ke sebuah diagram alur dan menentukan Inbreeding dari seorang individu dengan
menambahkan perbedaan alur yang memungkinan untuk satu atau lebih nenek moyang.
Contoh :
Setiap panah dalam diagram alur mewakili gamet dan 50% dari seorang individu genom. Perhatikan bahwa F tidak digunakan untuk menunjuk seorang individu; F tidak pernah digunakan karena simbol untuk Inbreeding.
Nilai-nilai individu Inbreeding ditentukan dengan menggunakan rumus berikut:
Fx = ∑ [(0.5)N (1+FA)]
Dimana Fx adalah inbreeding individu, ∑ merupakan simbol untuk
“jumlah" atau "menambahkan," N adalah jumlah individu yang mengikuti jalur, dan Fa adalah inbreeding dari nenek moyang bersama. Jika F = 0, menjadi
Fx = ∑ [(0.5)N]
Individu G di silsilah sebelumnya bawaan karena salah satu nenek moyangnya muncul di kedua sisi ibu dan ayah dari silsilah (definisi dari nenek moyang yang sama). Individu A adalah nenek moyang dari G. Inbreeding dari G ditentukan dengan menelusuri jalur dari G ke A. Ketika Anda menelusuri jalur, apa yang akan Anda lakukan adalah menentukan bagaimana gen A berakhir di G. Untuk melakukannya, Anda mulai dengan salah satu orang tua G, menelusuri jalur ke A, dan kemudian menelusuri dari A ke orangtua G lain.
Untuk menghitung FG, jalur dari D ke E , melalui indukan umum:
Indukan umum dari G: A Jalur dari G ke A: D-A-E
Disana terdapat 3 individu pada jalur ini, jadi N= 3. Individu A tidak inbred, jadi Eq. (4.9) dapat dipakai untuk mengkalkulasikan Fg
Fg = = 0.125
Jika terdapat lebih dari satu indukan umum yang muncul kamu dapat menambahkan hasilnya pada masing-masing bagian.
Silsilah Path diagram
Individu G adalah inbred karenaa keduanya memiliki induk umum. Untuk mengkalkulasikan FG jejak dari keturunan C ke D, seluruh nenek moyang G.
Jalur diagram G ke A: C-A-D
Jalur dari G ke B: C-B-D
Disana terdapat 2 bgian dan semuanya individu , pada bagian ini N= 3 di keduanya. Individu A dan B tidak inbred, jadi Eq dapat di gunakan untuk menghitung FG untuk menghitung FG tambahkan dari bagian yang terpisah:
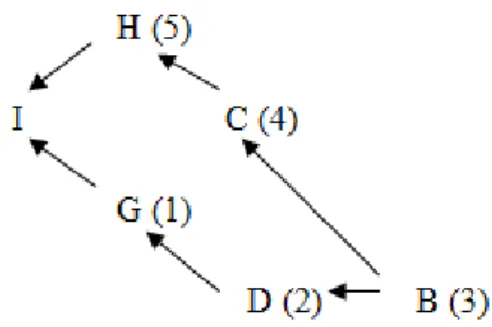
Fg = + Fg= 0,25
Untuk menghitung F1, melacak jalur dari G ke H, melalui induk I Secara umum induk I : C,A, dan B
Jalur dari I ke C: C-G-H
Jalur dari I ke B: G-D-B-C-H
Ada tiga jalur. Ada tiga individu di jalurnya dari I ke C dan lima individu di jalurnya dari I ke A dan I ke B. Akibatnya, N adalah 3, 5, dan 5, untuk jalur masing-masing. Seperti sebelumnya, tidak ada induk yang sama yang , sehingga persamaan. (4.9) dapat digunakan untuk menghitung F1. Untuk menghitung F1, produk-produk dari tiga jalur terpisah ditambahkan:
F1 = (0,5)3 + (0,5)5 + (0,5)5
F1 = 0,1875
Bahkan mungkin ada lebih dari satu jalur antara individu dan induk yang sama. Jika ada jalur kedua, Anda hanya menghitung dan menambahkannya ke total.
Ada satu aturan penting tentang menentukan jalur antara individu dan induk : Anda tidak dapat menelusuri jalur, yaitu, Anda tidak bisa pergi melalui individu dua kali dalam jalur yang diberikan. Dengan demikian, Anda tidak dapat membuat jalur C-D-A-C-G untuk jalur dari I ke A dalam contoh sebelumnya, karena Anda akan pergi melalui individu G dua kali.
Diagram jalur kedua menggambarkan konsep penting. Perkawinan sekerabat dapat dikurangi menjadi nol jika dua individu terkait kawin. Individu K tidak melakukan kawin sekerabat karena K tidak memiliki induk yang sama. Dua induk K adalah bawaan, tapi karena orang tua K terkait, Fk = 0. Akibatnya, jika Anda dapat mengidentifikasi individu ikan dan silsilahnya, Anda dapat mencegah perkawinan sekerabat hanya dengan mengawinkan individu yang tidak berhubungan. Cara klasik untuk menghilangkan perkawinan sedarah pada hewan
yang akan digunakan untuk pertumbuhan adalah untuk menghasilkan hibrida. Jika strain atau garis keturunan disimpan murni, hibrida F1 akan selalu memiliki F = 0. Apakah F itu memiliki nilai rata-rata? F adalah ukuran dari pertambahan di homozigosit seperti hasil dari perkawinan sekerabat. Dengan demikian, seekor ikan dengan F=25% dimana 25% lebih banyak homozigot dari pada rata-rata ikan didalam populasi. F tidak menyatakan tentang tingkat yang tetap dari homozigosit atau rata-rata populasi. F adalah nilai relatif untuk rata-rata populasi.
Perhitungan individu nilai F memiliki 1 kekurangan yang sangat besar di budidaya perikanan. Untuk itu, anda harus mengetahui silsilah individu. Informasi ini memiliki kekurangan, karena lebih banyak tempat penetasan tidak melengkapi atau tidak dapat untuk memberikan nilai-nilai identifikasi pada setiap individu ikan. Oleh karena itu, ini tidak mungkin untuk memperhitungkan tingkat individu dari perkawinan sekerabat untuk ikan yang lebih banyak.
Apakah rata-rata perkawinan sekerabat itu tidak dapat diukur dan dapat diabaikan? Jawabanya pasti tidak. Anda tidak boleh menentukan silsilah individu dan perhitungan nilai perkawinan sekerabat untuk individu ikan, tetapi karena terjadi di beberapa populasi, sangat penting sekali untuk perhitungan rata-rata perkawinan sekerabat untuk individu di dalam populasi jika kamu dapat mengaturnya dengan pantas.
2.4 Efek ukuran populasi dalam perkawinan sedarah dan Genetik Drift
Perkawinan sedarah yang tidak disengaja dan hanyutnya genetik tejadi di populasi hatchery karena ukurannya kecil dan tetutup. Kombinasi ini dengan cepat mengguncang variasi populasi dan peningkatan perkawinan sedarah, yang akan menurunkan produktivitas dan menambah biaya produksi. Masalah ini mungin lebih penting untuk populasi ikan. Ikan yang dibesarkan untuk pakan atau umpan hidupnya relatif dimanjakan. Pada manajemen hatchery dilakukan segala hal untuk menjaga agar ikan tetap hidup. Ikan yang ditebar pada air alami pergi dari lingkungan penetasan yang relatif ringan ke lingkunngan yang sangat keras dimana kelangsungan hidup lemah. Akibatnya hilanglah varians genetik dan perkawinan sedarah mungkin dapatmempengaruhi populasi yang ditebar dialam
jauh lebih banyak daripada populasi yang tidak pernah meninggalkan pembenihan (hatchery).
Secara genetik, populasi ideal adalah populasi yang besar. Sayangnya (atau untungnya tergantung pada pandangan anda), manajemen hatchery tidak bisa bekerja dengan populasi yang banyak/besar. Manajemen hatchery harus bekerja dengan populasi yang sedikit/kecil yang terbatas. Ketika populasi terbatas, cara yang terbaik untuk menggambarkan hal itu adalah bukan oleh jumlah populasi tetapi dengan nomor efektif pemuliaan. Nomor efektif pemuliaan tergantung pada beberapa faktor. Yang sangat penting adalah totall dari individu hasil pemuliaan, rasio sex, sistem perkawinan, dan varians dari ukuran keluarga. Ketika tidak ada pilihan yang terjadi , ada dua sistem perkawinan yang bisa digunakan dalam manajemen hatchery. Perkawinan acak atau perkawinan baik, perkawinan acak sering digunakan secara eksklusif dalam akuakultur. Angka efektif perkawinan populasi ketika dikawinkan secara acak dihitung dengan menggunakan rumus berikut
♀ ♂ ♀ ♂ (4.10)
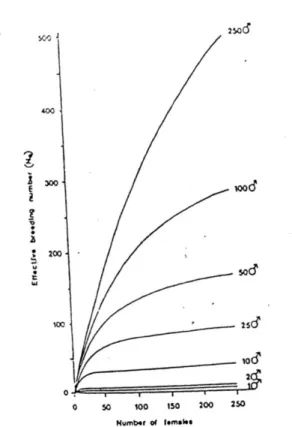
Dimana Ne adalah angka efektif pembiakan, ♀ adalah jumlah betina yang menghasilkan keturunan yang layak, dan ♂ adalah jumlah jantan yang menghasilkan keturunn yang layak. Pemeriksaan rumus sebelumnya menunjukan bahwa Ne dapat meningkat dalam dua cara : meningkatkan jumlah pembiakan individu atau membawa populasi lebih dekat untuk rasio jenis kelamin 50:50 (gambar 4.18)
Angka efektif pembiakan adalah salah satu konsep penting dalam pengelolaan populasi,yang memberikan indikasi tentang stabilitas genetik populasi karena Ne berbanding terbalik dengan perkawinan sedarah dan hanyutan genetik. Ketika Ne menurun, perkawinan sedarah dan varians dari perubahan dalam frekuensi gen akibat hanyutan genetik meningkat. Perkawinan sekerabat yang dihasilkan oleh satu generasi kawin dalam populasi tertutup adalah
Perubahan dalam frekuensi gen karena sampling error disebut hanyutan genetik. Hanyutan genetik mengekspresikan varians dari perubahan frekuensi gen. Dalam hal ini berbanding terbalik dengan Ne.
Dan itu berbanding terbalik dengan Ne : =
Gambar 4.18 angka pembibitan yang efektif yang dihasilkan dengan perkawinan dari berbagai kombinasi jantan dan betina. angka pembibitan yang efektif dihitung dengan menggunakan dua asumsi: (1) perkawinan acak yang telah berlaku: (2) semua induk memberikan kontribusi sama untuk generasi berikutnya.
Dimana adalah variasi dari perubahan frekuensi gen, dan p dan q adalah frekuensi alel p dan q pada lokus tertentu.
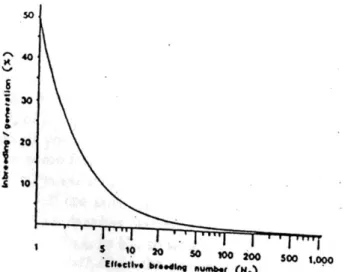
Gambar 4.19 Hubungan antara angka pembibitan yang efektif dan persilangan Sumber. Tare (1986b)
Melayang dan artinya bahwa ketika menurun, genetik melayang meningkat. Efek utama dari pergeseran genetik adalah hilangnya beberapa alel dan fiksasi lain. Alel jarang mudah hilang, tetapi alel lebih umum juga bisa hilang melalui pergeseran genetik.
Efek keseluruhan dari kecil adalah homozigot diproduksi oleh persilangan dan juga kehilangan alel sebagai akibat dari pergeseran genetik. Dengan demikian, reduksi ireversibel merusak gen dengan menghilangkan alel dan menciptakan homozigot. Setelah homozigot meningkat melalui persilangan, alel mungkin akan hilang lebih cepat karena sampel kecil dapat mengumpulkan jumlah yang tidak proporsional dari ikan yang homozigot pada lokus tertentu. Akibatnya, ada penurunan kebugaran secara keseluruhan, kelangsungan hidup, dan produktivitas, ketika populasi menjadi tidak dapat menyesuaikan diri dengan perubahan lingkungan, karena beberapa potensi genetik yang telah hilang.
Garis pewarisan sifat populasi pembenihan seragam mungkin menjadi salah satu alasan mengapa sulit untuk membuat populasi mereproduksi dirinya sendiri di badan air alamiah oleh stocking pembenihan memproduksi ikan. garis pewarisan sifat seragam mungkin memadai dalam lingkungan terkendali seperti pembenihan, tetapi mereka merugikan di alam liar di mana lingkungan adalah apapun tetapi seragam. Ini adalah alasan bahwa tujuan utama dalam peternakan
laut dari salmon adalah untuk mempertahankan lebih banyak keragaman genetik yang memungkinkan.
Sekali penurunan dalam telah menghasilkan persilangan, persilangan dalam menurunkan masa dalam siklus umpan balik positif. Setelah persilangan telah terjadi, menjadi
=
Dimana adalah pada populasi inbrida. Dengan demikian, pembatasan dalam dan F cenderung untuk memberi makan satu sama lain, dan situasi dapat terus memburuk .
Ketika sebuah populasi memiliki yang telah direduksi, kenaikan berikutnya tidak memperbaiki kerusakan yang telah terjadi. Pembatasan dalam sering terjadi selama transfer persediaan dari satu pembenihan ke yang lainnya. Itu mahal dan sulit untuk membuka populasi besar, begitu banyak pembenihan dimulai dengan beberapa ikan, terutama jika ikan sangat produktif. Ketika ini terjadi, populasi dikatakan telah melalui hambatan. penurunanDramatis dari persediaan dan produktivitas. melalui serangkaian generasi adalah sebuah harmoni artinya dari di setiap generasi. Selama t generasi, kelesuruhan dapat ditentukan dari rumus berikut:
= )
Dimana adalah efektif keseluruhan pembibitan, dan , , dan adalah S di generasi 1, 2, dan t masing-masing. Rumus sebelumnya
menunjukkan bahwa generasi dengan terkecil memiliki dampak terbesar pada keseluruhan
Sebuah hambatan telah berefek tahan lama pada genetika sebuah populasi genetika . Nei et al (1975) menunjukkan bahwa hambatan mengurangi varians genetik dan rata-rata heterozigositas, dan tergantung pada dari hambatan dan
peningkatan ukuran populasi setelahnya, sebuah populasi mungkin tidak kembali pulih ini adalah keanekaragaman genetik untuk ratusan generasi mungkin tidak kembali pulih ini adalah keanekaragaman genetik untuk ratusan generasi, selain itu juga melalui pengenalan persediaan baru.
Ini memiliki implikasi praktis yang penting untuk pengelolaan persediaan pembenihan. Jika populasi pembenihan dimulai dengan hanya beberapa individu, dengan keturunan dari hanya sedikit menumbuhkan, atau populasi hancur oleh penyakit, peningkatan berikutnya dalam hanya menjaga persilangan dan pergeseran genetik dari semakin buruk; mereka tidak memperbaiki masalah yang terjadi. Jadi, sangat penting untuk mengetahui untuk setiap generasi, baik di pembenihan dan sebelum persediaan tiba.
suatu populasi juga bisa melewati hambatan selama seleksi. Jika Anda
membangun nilai cutoff yang ekstrim, Anda mungkin dapat menghemat lebih sedikit ikan , dan akan berkurang drastis pada populasi yang dipilih. Cara lain bahwa seleksi dapat membuat hambatan adalah jika ikan yang dipilih berasal dari hanya sedikit famili. Ikan dari beberapa keluarga bisa menjadi lebih unggul kepada yang lainnya tersebut baik dikarenakan VA, VD, V1, VG-E, atau VE,, jadi
ketika Anda memilih ikan, Anda mungkin mengurangi pada generasi sebelumnya jika hanya sedikit induk yang dihasilkan populasi yang dipilih. Sementara kedua jenis hambatan mungkin sama-sama merusak, yang terakhir adalah salah satu yang tak terlihat karena Anda mungkin tidak akan menyadari bahwa Anda secara drastis mengurangi .
Faktor terakhir yang dapat mengurangi adalah ukuran famili. Jika semua penetas menghasilkan jumlah yang sama dari keturunan, dihitung seperti yang dijelaskan sebelumnya. Namun, produksi gamet dan keturunan viabilitas sangat bervariasi. Ini memiliki dampak negatif pada karena kontribusi yang tidak sama untuk generasi berikutnya akan menurunkan dari
apa yang akan telah memiliki semua penetas membuat kontribusi yang sama. Ketika ada produksi yang keturunannya tidak sama, dalam suatu populasi dengan perkawinan acak adalah
=
♂ ♀
Dimana adalah dengan produksi dengan keturunan yang tidak sama dan ♂ dan ♀ adalah varians produksi keturunan jantan dan betina (ukuran famili), masing-masing. Ukuran keluarga sering berasumsi apa yang disebut distribusi Poisson. Dalam distribusi Poisson, mean dan varians adalah
sama, sehingga ketika hal ini terjadi, berarti ukuran famili dapat digantikan untuk varians (Latter 1959).
Hilangnya sengaja varians genetik melalui pengurangan dapat sangat merusak. Varians genetik adalah bahan baku dengan kedua alam dan genetika bekerja, dan kehilangan adalah merusak karena biasanya ireversibel. Hilangnya varian genetik dapat menurunkan produktivitas, meningkatkan anomali perkembangan, dan melakukan perbaikan di masa depan melalui seleksi yang sulit.
Sebuah penelitian angka telah menyarankan bahwa penurunan memiliki efek buruk pada kandungan gen dari persediaan tempat penetasan (Allendrof dan Ulter 1979; Allendrof dan Phelps 1980; Ryman dan Stahl 1980; Tave dan Smitherman 1980; Salib dan Raja 1983; Stahl 1983; Taniguchi et al. 1983). Sebagai contoh, Teichert-Coddington (1983) tidak dapat meningkatkan laju pertumbuhan di Auburn University (Pantai Gading) strain T. Nilotica dengan seleksi. Ini mungkin karena penurunan selama beberapa alur transfer ini yang
mungkin telah menghilangkan sebagian besar untuk meningkatkan pertumbuhan (Tave dan Smitherman 1980). Leary et al. (1985a) menemukan frekuensi tinggi dari dua cacat morfologi dan jumlah individu yang sangat besar yang asimetris pada fenotipe meristic bilateral pada populasi pembenihan ikan cutthroat trout. Mereka di keadaan jajal ini mengembangkan anomali untuk pengurangan heterozigositas, sebagai akibat dari pergeseran genetik. Tingkat penetasan telah menurun di galur Donaldson dari rainbow trout (Hershberger 1985). Hal ini mungkin karena penurunan ; berkurang menjadi sekitar 20 /
generasi selama 40 tahun terakhir. Ini telah menghasilkan perkawinan sedarah antara 40 dan 60% (Hershberger 1983), dan data elektroforesis dari Allendrof dan Utter (1979) menunjukkan bahwa pergeseran genetik telah merenggut populasi banyak alel yang berharga.
Seburuk ini untuk persediaan tempat penetasan yang digunakan dalam budidaya ikan, dapat punah ketika terjadi pada populasi yang digunakan dalam budidaya ikan, dapat punah ketika terjadi pada populasi yang digunakan untuk persediaan media air alamiah. US Fish dan Wildlife Service (1982) memperingatkan bahwa hilangnya keragaman genetik di persediaan pembiakkan
danau trout yang digunakan untuk persediaan Danau Michigan dan Ontario dapat menghalangi pemulihan spesies itu. Rasmuson (1981), Ryman (1981), dan Johansonn (1981) semua khawatir dan menyatakan bahwa praktek manajemen induk yang mengurangi variasi genetik di persediaan pembiakkan Scandanavian salmon yang digunakan untuk melengkapi produksi alami. Ryman (1981) memperingatkan bahwa banyak program manajemen benar-benar dapat menghancurkan populasi kandungan gen bukannya menyelamatkan mereka.
2.5 Mencegah Penurunan Dalam Angka Efektif Breeding
Bagaimana anda mencegah batasan di dari menjalankan potensi genetik populasi pembenihan? Pertama dan terpenting, menjaga sebesar mungkin setiap generasi. Ketika memperoleh induk, Anda harus menentukan, diawal, silsilah ikan yang anda beli. Anda perlu tahu berapa banyak induk yang digunakan untuk menghasilkan ikan yang Anda beli. Ini merupakan perhatian penting dalam akuakultur, karena hal ini menentukan kesuburan ikan. Saya pernah melihat seseorang memenuhi permintaan untuk yayasan ikan lele dengan pengiriman 2.000 benih yang berasal dari bibit tunggal. yang menghasilkan 2000 bibit ikan lel hanya 2 (kurang jika induk adalah terkait dan bawaan). Bahwa perkawinan silang pada populasi dasar ini ini akan menghasilkan pada generasi pertama adalah 25%, tetapi kehilangan banyak alel melalui pergeseran genetik bahkan lebih merusak.
Berapakah besar Ne seharusnya , untuk mencegah perkawinan sedarah dan
pengendalian genetika ?. Kincaid merekomendasikan bahwa Ne minimal 200
(1976a) dan 500 (1979); Ryman dan Stahl (1989) merekomendasikan bahwa Ne
minimal 60; Organisasi Makanan dan Pertanian dari U.N. (FAO) merekomendasikan bahwa Ne minimal 50 untuk jangka pendek dan jangka
panjang masa kerja (FAO/UNEP 1981); U.S. Ikan dan Layanan satwa liar 1984 merekomendasikan bahwa Ne minimal 100; dan Tave (1986C) merekomendasikan
Nes antara 263 dan 344 untuk makanan ikan dan populasi ikan yang dijadikan
umpan dan Nes antara 424 dan 685 untuk populasi yang akan ditebar di badan air
Manakah rekomendasi yang benar ? Sayangnya tidak ada satu nomor dari manajer perusahaan penetasan dapat digunakan untuk mencegah pergeseran genetika atau perkawinan sedarah dari masalah di persediannya. Tave(1986B,1986C) diuraikannya prosedur untuk mencegah minimum Nes yang
dapat digunakan untuk mencegah perkawinan sedarah dan pergeseran genetika terkait masalahnya.
Untuk mengitung Ne yang diperlukan untuk mencegah perkawinan sedarah
untuk mencapai tingkat yang menekan produktivitas, anda membutuhkan 2 potongan informasi. Informasi pertama adalah level perkawinan sedarah dimana perkawinan sedarah terjadi depresi. Sayangnya , informasi ini tidak ada untuk ikan; nilai tunggal yang universal mungkin tidak akan pernah ada, karena itu akan berbeda untuk perbedaan fenotip dan perbedaan populasi. Kincaid’s(1976A.1976B,1983B) mempelajari tentang rainbow trouth dengan lengkap, tapi tingkat terkecil perkawinan sedarah itu dia memproduksi hanya 12,5%. . Kincaid(1977) memperkirakan F=18% adalah tingkat di mana perkawinan depresi menjadi signifikan dalam rainbow trouth. Bagaimanapun, beberapa dari data Kincaid(1976A) memperlihatkan depresi pada perkawinan sedarah pada F=12,5%. Data dari hewan yang lain memperlihatkan bahwa depresi pada perkawinan sedarah bisa terjadi pada perkawinan sedarah (falconer1981). Karena tidak ada nilai-nilai kritis yang ada pada ikan. Nilai hipotetis harus di gunakan. Tave(1986) menyarankan menggunakan 5% sebagai nilai konservatif dan 10% sebagai perkiraan liberal.
Informasi kedua yang diperlukan adalah jumlah generasi yang ingin anda masukkan ke dalam program perkawinan sedarah sebelum F mencapai nilai kritis. Setelah itu ditentukan, Anda hanya menghitung nilai konstan minimum Ne yang
akan menghasilkan tingkat kritis perkawinan sedarah dalam jumlah yang ditentukan dari generasi. Sebagai contoh, jika kamu memilih F=5% sebagai tingkat kritis Anda dari perkawinan sedarah dan Anda tidak ingin mencapai tingkat itu sampai generasi 15, Ne dihitung sebagai berikut :
Langkah pertama, hitung F/generasi yang dibutuhkan untuk memproduksi F=0.05 dari generasi 15
F/generasi = 0.0033333/generasi
Langkah kedua, hitung Ne yang di butuhkan untuk memproduksi
F=0.0033333/generasi menggunakan Eq.(4.11)
F = 0.0033333 = (0.0033333)(2) = Ne = Ne = 150
Minimum konstan Nes yang memperoduksi F=5% dan F=10% setelah
jumlah generasi di tetapkan yang tercantum dalam tabel 4.10.
Harus seberapa besar Ne untuk mencegah pergeseran genetik? itu jauh
lebih sulit untuk mencegah pergeseran genetik daripada perkawinan sedarah yang mencapai tingkatan yang menekan produktivitas, karena setiap perubahan frekuensi gen sebagai akibat dari kesalahan sampling adalah pergeseran genetik. Jika frekuensi akibat dari perubahan alel 0,500-0,499 sebagai kesalahan hasil sampling, pergeseran genetik telah terjadi. tapi itu tidak penting, sebagai hilangnya alel sehingga pertanyaan penting menjadi seberapa besar yang N harus mencegah hilangnya alel langka (alel dengan frekuensi rendah) akan hilang dibandingkan dengan frequecies tinggi. jawaban untuk quetion ini tergantung pada dua keputusan yang Anda harus membuat (1) bagaimana berharga adalah alel langka
Percakapan Obrolan Berakhir yaitu, bagaimana langka alel akan Anda mencoba untuk menyimpan (misalnya, f = 0,1 atau 0,01 atau 0,000001); (2) apa tingkat probabilitas yang diinginkan (misalnya, p = 0,05 berarti bahwa Anda memiliki probabilitas 95% dari tabungan alel; p = 0,01 berarti bahwa Anda memiliki probabilitas 99% dari tabungan alel).
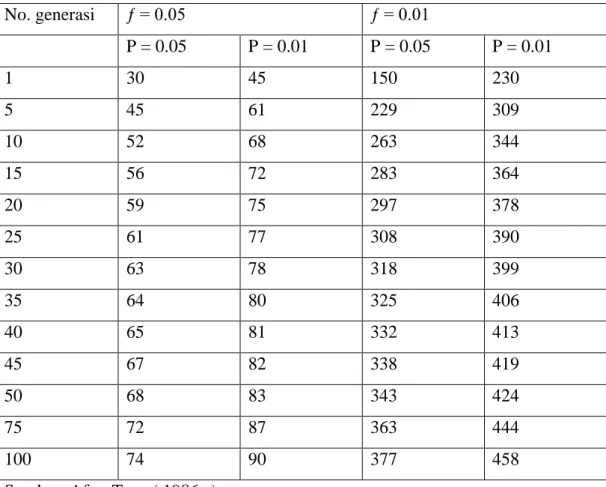
Tabel 4.10 nomor pemuliaan yang efektif (N,) yang dibutuhkan untuk menghasilkan F = 5% dan F = 10% setelah nomor-nomor tertentu generasi
F= 5% F = 10% 1 10 5 2 20 10 3 30 15 4 40 20 5 50 25 6 60 30 7 70 35 8 80 40 9 90 45 10 100 50 20 200 100 30 300 150 40 400 200 50 500 250 60 600 300 70 700 350 80 800 400 90 900 450 100 1000 500
Jika Anda ingin menyimpan salah satu alel paling langka (f = 0,000001) dan Anda ingin jaminan 100%, Anda akan memerlukan sebuah N, begitu besar sehingga tidak akan pernah cocok di hatchery a. akibatnya, kompromi harus dibuat antara apa yang ideal dan apa yang akan menimbulkan masalah. N yang Anda butuhkan dapat ditentukan dengan menghitung jumlah ikan yang diperlukan untuk memastikan bahwa alel q, pada frekuensi tertentu, hadir pada probabilitas yang diberikan. yang probabillity (p) dari tidak menjaga sebuah alel dalam sampel acak adalah
p = (1,0-q)2Ne
Di mana p adalah probabilitas kehilangan q alel dalam satu sampel (dari generaion atau pada salah satu transfer induk dari satu stasiun ke yang lain) dan q
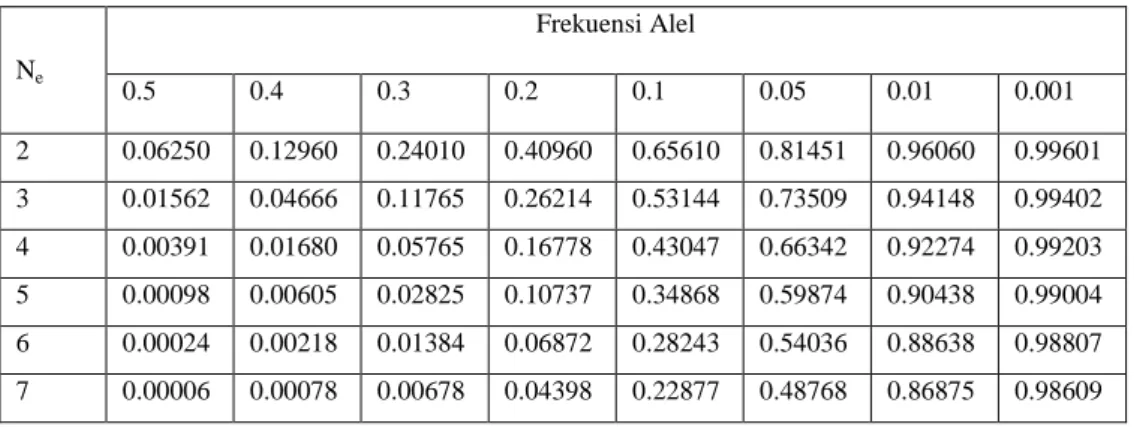
adalah frekuensi alel. eksponen adalah 2N, karena ikan yang diploid dan memiliki 2 alel per lokus. Tabel 4.11 menunjukkan probalilites kehilangan alel untuk berbagai N s pada berbagai frekuensi gen. misalnya, bahwa informasi dalam tabel 4.11 menunjukkan bahwa Anda memerlukan N, 150 untuk menghasilkan ap = 0,04904 untuk alel yang f = 0,01 (probabilitas 4,9% dari kehilangan alel). probabilitas yang tercantum dalam tabel 4.11 adalah probabilitas kehilangan alel melalui pergeseran genetik hanya satu generasi atau generasi tunggal atau transfer ke penetasan lain produk dari probabilitas untuk setiap generasi atau transfer. misalnya jika Anda mempertahankan N konstan, dari 150 untuk 10 generasi, probabilitas kehilangan sebuah alel yang f = 0,01 setelah 10 generasi dihitung sebagai berikut
Langkah 1. menghitung probabilitas kehilangan alel dalam satu generasi menggunakan Persamaan. (4.16)
p = (1,0-4) p = (1,0-0,01)
p = 0,04904
Langkah 2. menghitung probabilitas 0f menjaga allela dalam satu generasi. untuk menghitung probabilitas menjaga allela itu, kurangi kemungkinan kehilangan allela (p) from1.0;
probabilitas menjaga allela = 1,0-0,04904 probabilitas menjaga allela = 0,95096
Langkah 3. menghitung probabilitas menjaga allela setelah 10 generasi itu adalah produk dari probabilitas menjaga
Tabel 4.11 probabilitas kehilangan alel melalui pergeseran genetik selama delapan frekuensi allelia di berbagai Nomor pemuliaan yang efektif (Ne)a
Ne Frekuensi Alel 0.5 0.4 0.3 0.2 0.1 0.05 0.01 0.001 2 0.06250 0.12960 0.24010 0.40960 0.65610 0.81451 0.96060 0.99601 3 0.01562 0.04666 0.11765 0.26214 0.53144 0.73509 0.94148 0.99402 4 0.00391 0.01680 0.05765 0.16778 0.43047 0.66342 0.92274 0.99203 5 0.00098 0.00605 0.02825 0.10737 0.34868 0.59874 0.90438 0.99004 6 0.00024 0.00218 0.01384 0.06872 0.28243 0.54036 0.88638 0.98807 7 0.00006 0.00078 0.00678 0.04398 0.22877 0.48768 0.86875 0.98609
8 0.00002 0.00028 0.00332 0.02815 0.18530 0.44013 0.85146 0.98412 9 4 x 10-6 0.00010 0.00163 0.01801 0.15009 0.39721 0.83451 0.98215 10 1 x 10-6 0.00004 0.00080 0.01153 0.12158 0.35849 0.81791 0.98019 14 6 x 10-7 0.00005 0.00193 0.05233 0.23783 0.75472 0.97237 15 0.00002 0.00124 0.04239 0.21464 0.73970 0.97943 20 6 x 10-7 0.00013 0.01478 0.12851 0.66897 0.96077 25 0.00001 0.00515 0.07694 0.60501 0.95121 30 2 x 10-6 0.00180 0.04607 0.54716 0.94174 31 1 x 10-6 0.00146 0.04158 0.53627 0.93985 35 0.00063 0.02758 0.49484 0.93236 40 0.00022 0.01652 0.44752 0.92308 45 0.00008 0.00989 0.40473 0.91389 50 0.00003 0.00592 0.36603 0.90479 55 9 x 10-6 0.00354 0.33103 0.89578 60 3 x 10-6 0.00212 0.29938 0.88687 66 9 x 10-7 0.00115 0.26537 0.87628 70 0.00076 0.24487 0.86930 73 0.00046 0.22145 0.86064 80 0.00027 0.22028 0.85208 85 0.00016 0.18113 0.84359 90 0.00010 0.16381 0.83520 95 0.00006 0.14814 0.82688 100 0.00004 0.13398 0.81865 125 3 x 10-6 0.08106 0.77870 135 1 x 10-6 0.06630 0.76328 150 0.04904 0.74071 175 0.02967 0.70456 200 0.01795 0.67019 225 0.01086 0.63748 230 0.00982 0.63114 250 0.00657 0.60638 275 0.00398 0.57679 300 0.00240 0.54865 325 0.00146 0.52188 350 0.00088 0.49641 375 0.00053 0.47219 400 0.00032 0.44925 425 0.00019 0.42723 450 0.00012 0.40639 475 0.00007 0.38656 500 0.00004 0.36770
685 1 x 10-6 0.25393
1498 0.04991
2302 0.00999
6880 1 x 10-6
probabilitas ini adalah probabilitas kehilangan sebuah alel untuk satu generasi. alel untuk setiap generasi: probabilitas menjaga alel = (0,95096)
probabilitas menjaga alel = 0,60481
Langkah 4. menghitung probabilitas kehilangan allela (p) setelah 10 generasi. caculate probabilitas menjaga allela dari 1,0:
p = 1,0-,60481 p = 0,39519
Sehingga jika Anda mempertahankan N 150 / generasi selama 10 generasi Anda memiliki probabilitas 60,5% dari menjaga alel yang f = 0,01 setelah 10 generasi, walaupun memiliki probabilitas 95% dari menjaga alel dalam setiap generasi.
Jika Anda memiliki populasi berfluktuasi, probabilitas keseluruhan menjaga alel adalah, sekali lagi, produk dari probabilitas untuk setiap contoh generation.for, apa pobability untuk setiap generasi. misalnya, apa adalah probabilitas menjaga alel yang f = 0,01 setelah 10 generasi dengan Ns / generasi berikut ?
`N,s : 230, 100, 200, 50, 30, 200,10, 20,25,230
Langkah 1. menghitung probabilitas kehilangan alel (P) dan probabilitas menjaga alel yang f = 0,01 untuk setiap generasi:
Ne Kemungkinan kehilangan alel Kemungkinan
mempertahankan alel Menggunakan Eq. (4,16) 2Ne 1,0 230 2(230) 0,00982 0,99018 100 2(100) 0,13398 0,86602 200 2(200) 0,01795 0,98205 50 2(50) 0,36603 0,63397 30 2(30) 0,54716 0,45284
200 2(200) 0,01795 0,98205 10 2(10) 0,81791 0,18209 20 2(20) 0,66897 0,33103 25 2(25) 0,60501 0,39499 230 2(230) 0,00982 0,99018
Langkah 2. Menghitung kemungkinan mempertahankan alel setelah generasi ke 10. Hasil dari kemungkinan mempertahankan alel untuk setiap generasi :
Kemungkinan mempertahankan alel = (0,99018)(0,86602)(0,98205)(0,63379) (0,45284)(0,98205)(0,18209)(0,33103)
(0,39499)(0,99018)
Kemungkinan mempertahankan alel = 0,00560
Langkah 3. Menghitung kemungkinan kehilangan alel setelah generasi ke 10. Untuk menghitung kemungkinan kehilangan alel ( ), yaitu dengan cara 1,0 dikurangi hasil kemungkinan mempertahankan alel :
= 1,0 – 0,00560 = 0,99440
Contoh ini dengan jelas menunjukkan dampak yang merugikan bahwa penurunan Ne dapat menyebabkan genetic drift (pergeseran genetik). Bahkan jika Ne dikurangi hanya untuk satu generasi, maka bottleneck yang dapat bertahan lama populasi genetika tersebut.
Misalnya, jika Ne untuk generasi pertama adalah 10, lalu meningkat dan dipertahankan pada 230 untuk kepentingan 9 generasi, kemungkinan kehilangan alel ƒ = 0,01 setelah generasi ke-10 adalah
Langkah 1: Menghitung kemungkinan kehilangan alel ( ), dan kemungkinan mempertahankan alel dengan ƒ = 0,01 untuk setiap generasi :
Ne Kemungkinan kehilangan alel Kemungkinan
mempertahankan alel Menggunakan Eq. (4,16)
10 2(10) = 0,81791 0,18209 230 2(230) = 0,00982 0,99018
Langkah 2. Menghitung kemungkinan mempertahankan alel setelah generasi ke 10. Hasil dari kemungkinan mempertahankan alel untuk setiap generasi :
Kemungkinan mempertahankan alel = (0,18209)(0,99018) Kemungkinan mempertahankan alel = 0,16661
Langkah 3. Menghitung kemungkinan kehilangan alel setelah generasi ke 10. Untuk menghitung kemungkinan kehilangan alel ( ), yaitu dengan cara 1,0 dikurangi hasil kemungkinan mempertahankan alel :
= 1,0 – 0,16661 = 0,83339
Kemungkinan keseluruhan kehilangan alel memberikan kemungkinan kehilangan alel melalui pergeseran genetik setelah beberapa generasi. Jika alel yang benar-benar kehilangan, maka Ne bisa menjadi salah satu dari alel tersebut dan frekuensi alel masih akan menjadi nol. Sekali kehilangan, sebuah alel hanya dapat diperkenalkan kembali oleh mutasi atau dengan impor induk baru. Hal ini menjelaskan mengapa kemacetan, secara permanen dapat melumpuhkan perbedaan genetik suatu populasi.
Apakah manajer pembudidaya harus menggunakan beberapa pedoman NeS untuk mencegah pergeseran genetik dengan menghilangkan alel langka? Untuk menghitung ini, Anda harus menentukan tiga hal. Yang pertama adalah frekuensi alel langka yang ingin Anda simpan. Tave (1986) merekomendasikan bahwa tujuannya harus menjaga alel yang ƒ = 0,01 karena biologi populasi dan genetika populasi umumnya menganggap bahwa alel yang ƒ ≥ 0,01 berkontribusi kepada polimorfisme, dan tujuannya adalah untuk menjaga alel lokus polimorfik di negara polimorfik. Meffe (1986) merekomendasikan bahwa tujuannya harus untuk menjaga alel yang ƒ = 0,05 karena alel yang langka kemudian akan berkontribusi sedikit untuk perbedaan genetik secara keseluruhan.
Rekomendasi Meffe (1986) diterima untuk makanan ikan dan ikan umpan peternakan karena alel langka mungkin tidak mengatur produktivitas peternakan
ikan. di samping itu, alel yang paling langka mungkin hilang selama Proses domestikasi, dalam hal frekuensi akan meningkat secara dramatis. tapi rekomendasi Tave (1986) mungkin lebih tepat untuk program permainan ikan dan laut peternakan, serta bagi mereka yang mempertahankan garis acuan standar, karena tujuan utama dari program ini adalah genetika konservasi, untuk berkomunikasi dengan perbedaan genetik Anda harus menyimpan banyak alel secara praktis. Dalam program tersebut lebih baik untuk berbuat salah pada sisi konservatif dan aman.
Bagian kedua dari informasi yang diperlukan adalah kemungkinan ( ) yang ingin Anda gunakan (kemungkinan kehilangan alel). Dalam biologi, dua kemungkinan umumnya digunakan dan diterima: = 0,05 dan 0,01; = 0,05 dan 0,01 berarti bahwa ada kemungkinan 5 dan 1% dari kehilangan alel dan kemungkinan 95 dan 99% dari mempertahankan alel masing-masing.
Bagian ketiga dari informasi yang diperlukan adalah jumlah generasi yang akan dimasukkan ke dalam program pemuliaan sebelum mencapai tingkat yang diinginkan. Setelah Anda menentukan ini, Anda cukup menghitung kembali Ne / generasi yang diperlukan untuk menghasilkan yang diinginkan serta memberikan alel dalam jumlah yang telah ditetapkan generasi.
Misalnya, Anda menginginkan = 0,01 (kemungkinan 1% dari kehilangan alel dan kemungkinan 99% dari mempertahankan alel) setelah generasi ke-10 untuk alel ƒ = 0,01. Apakah Ne / generasi harus dipertahankan untuk mencapai tujuan ini? jawabannya dihitung sebagai berikut:
q= 0,01; = 0,01 setelah generasi ke-10
Langkah 1. Menghitung kemungkinan/generasi mempertahankan alel yang akan menghasilkan kemungkinan 0,99 dari mempertahankan alel tersebut ( = 0,01) setelah generasi ke 10.
0,99 = (kemungkinan/generasi)10 Kemungkinan/generasi = (0,99)1/10 Kemungkinan/generasi = 0,9989955
Langkah 2 . Menghitung generasi P ( P ) dengan mengurangi yang kemungkinan dari menjaga alel dari 1.0 :
P = 0.0010045
Langkah 3 . Menghitung tingkat Ne yang diperlukan untuk memproduksi P = 0.0010045 dengan menggunakan Eq. ( 4.16 ):
P =
0.0010045 =
Formula harus diubah menjadi logaritma untuk memecahkan untuk Ne : log o.0010045 = log
log 0.0010045 = (2N˛) (log 0.99) 2Ne = 686.867 Ne = Ne = 343.43 Ne dibulatkan menjadi 344
Dalam contoh sebelumnya, anda perlu sebuah konstanta Ne dari 344/generasi untuk menghasilkan P = 0.01 (kemungkinan dari 1 % kehilangan alel dan beberapa 99 % menjaga alel) setelah 10 generasi untuk sebuah alel yang ƒ = 0.01. Konstan Nes dibutuhkan untuk menghasilkan P = 0.05 dan 0.01 untuk alel yang ƒ = 0.05 atau 0.01 setelah berbagai jumlah generasi yang tercantum dalam tabel 4.12 .
Seperti halnya perkawinan sedarah , tidak ada universal Ne yang dapat direkomendasikan untuk setiap ikan program pengembangbiakan. Nes yang tercantum dalam tabel 4.12 harus digunakan hanya sebagai pedoman. Mereka bukan nilai mutlak. Kebanyakan makanan ikan pembudidaya harus menjaga konstan Nes diantara 68 dan 90. Sebuah konstanta Ne= 68 sudah cukup untuk pekerjaan jangka pendek ( 10 generasi), karena Ne akan menghasilkan P= 0.01 untuk sebuah alel yang ƒ = 0.05 setelah 10 generasi. Konstan Ne = 90 harus cukup untuk long-term bekerja ( 10 generasi , karena itu Ne akan menghasilkan P = 0.01 untuk sebuah alel yang ƒ = 0.05 setelah 100 generasi.
Tabel 4.12 efektif berkembang biak per (Ne) yang diperlukan untuk memproduksi
P = 0.01 dan 0.05 setelah berbagai nomor generasi untuk alel ƒ = 0.05 atau 0.01.
No. generasi ƒ = 0.05 ƒ = 0.01 P = 0.05 P = 0.01 P = 0.05 P = 0.01 1 30 45 150 230 5 45 61 229 309 10 52 68 263 344 15 56 72 283 364 20 59 75 297 378 25 61 77 308 390 30 63 78 318 399 35 64 80 325 406 40 65 81 332 413 45 67 82 338 419 50 68 83 343 424 75 72 87 363 444 100 74 90 377 458
Sumber: After Tave ( 1986c )
Efektif berkembang biak nomor itu dibulatkan ke berikutnya yang lebih tinggi jumlah keseluruhan.
Standar aquaculturists yang selalu memelihara referensi baris ikan atau yang ingin melestarikan lebih banyak alel harus mencoba untuk mempertahankan Nes ini di antara 263 dan 344/generasi. Sebuah konstanta Ne dari 263 sudah cukup memadai untuk jangka pendek bekerja ( 10 setelahnya, karena ini Ne akan menghasilkan P = 0.005/generasi, dan setelah 10 generasi P akan 0,05. Ne dari 344 adalah lebih tepat untuk jangka panjang bekerja ( 10 generasi), karena itu akan menghasilkan P = 0.001/generasi, dan setelah 10 dan 51 generasi, P = 0,01 dan 0,05 secara berturut-turut, untuk sebuah alel yang ƒ= 0,01.
Efektif berkembang biak nomor seharusnya minimal 424/generasi sebagai ikan budaya program di mana penduduk akan digunakan untuk program pengelolaan perikanan laut atau untuk peternakan. Seperti yang disarankan Ne lebih besar bagi keberhasilan program karena dua alasan: pertama, salah satu tujuan utama, jika
tidak tujuan utama, seharusnya mempertahankan seperti banyak genetik yang berbeda mungkin. Ikan ini akan dibakar di alam liar, dan tidak ada yang mengetahui yang alel preadapt ikan untuk berubah lingkungan. Hilangnya genetik yang berbeda melalui genetik melayang mungkin seorang perdana alasan mengapa sulit untuk memulihkan sebuah sumber daya alam dengan populasi produksi penetasan. Kedua, ketika bekerja dengan adanya program program tersebut, seorang manajer harus memasukkan pembudidayaan suhu tinggi perencanaan induk ke manajemen, dan 50 generasi yang baik minimum karena itu akan mencangkup di mana saja dari 50 untuk 200 + tahun. Sebuah konstanta Ne dari 424/generasi akan menghasilkan P = 0.01 dan 0.05 untuk alel yang ƒ = 0.01 setelah 50 dan 257 generasi, masing masing.
Minimum Ne ideal bagi keberhasilan program adalah 685/generasi karena itu Ne hampir meyakinkan alel yang ƒ = 0.01 tidak semuanya akan hilang. Sebuah konstanta Ne dari 685 akan menghasilkan /generasi, dan setelah 100 generasi P akan 0.0001 untuk alel yang ƒ = 0.01.
Meskipun begitu akan sangat ideal untuk menjaga alel yang rarer dari 0,01 untuk jenis program ini, Ne yang dibutuhkan untuk tujuan ini adalah hampir tidak dapat diatur. Misalnya, Ne diperlukan untuk mempertahankan sebuah alel yang ƒ = 0.001 di P = 0,05 untuk satu generasi adalah 1498. Jelas, hal ini tidak praktis untuk mencoba dan mencegah hilangnya alel banyak rarer dari 0,01.
Tidak mungkin bahwa setiap manajer pembudidayaan dapat menjaga nya diinginkan minimum Ne dari generasi ke generasi. Fluktuasi dalam Ne karena penyakit , pemijahan masalah , dll. Akan mengurangi Ne di bawah tingkat yang diinginkan. Jika tujuan adalah untuk menjaga alel yang ƒ = 0.05, Ne = 30 harus minimum hambatan, karena itu Ne akan menghasilkan P = 0.05 untuk 1 generasi. Yang lebih baik tujuannya adalah untuk menjaga hambatan minimum untuk Ne = 45, karena itu Ne akan menghasilkan P = 0.01 untuk 1 generasi. Jika tujuan adalah untuk menyelamatkan alel yang ƒ = 0.01, Ne = 150 harus menjadi pertimbangan minimal hambatan, karena itu Ne akan menghasilkan P = 0.05/generasi sebagai alel yang ƒ = 0.01 . Jika Ne tetes di bawah 150, P akan pergi 0.05/generasi dan kemungkinan menjaga alel akan turun di bawah 95%/generasi.
Nes yang telah dianjurkan untuk mencegah perkawinan sedarah mencapai tingkat produktivitas yang menekan dan untuk mencegah genetik melayang dari kehilangan dapat dikatakan pedoman; mereka tidak akan dianggap sebagai injil. Nes yang disampaikan didasarkan atas asumsi yang dijelaskan di bagian ini. Kunci aspek bagian ini adalah prosedur yang digunakan untuk menghasilkan Nes. Jika anda tahu bagaimana menggunakan prosedur, anda dapat menghasilkan Nes atas tujuan anda dan anda asumsi tentang apa yang diinginkan.
Apa yang harus dilakukan jika Ne dari populasi tetes pembudidayaan (atau catatan menunjukkan bahwa itu berkurang sebelumnya) dan mulai produktivitas lebih rendah? Satu satunya cara untuk memperbaiki seperti masalahnya adalah untuk memperoleh induk baru. Jika ini dilakukan, memastikan bahwa kamu mempunyai induk yang tidak melewati hambatan. Namun sesederhana seperti ini adalah resep, hal ini sering sulit obat untuk menelan. Banyak pengelola pembudidayaan benci ide membawa induk baru karena mereka takut impor yang penyakit baru lebih dari mereka takut kepada masalah genetik. Penyakit yang terlihat dan mudah dipahami, tetapi genetik melayang tidak terlihat. Selain itu, tambak-tambak yang menyebarkan terancam atau spesies langka sering tak bisa mengimpor induk baru karena mereka tidak tersedia.
Ketika impor induk baru tidak menunjang, beberapa opsi dapat digunakan untuk mencegah perkawinan sedarah dan genetik melayang semakin parah. Kalau opsi pertama adalah untuk meningkatkan Ne sebanyak mungkin; semakin besar pula Ne lebih baik. Yang biasa hambatan ini adalah fisik keterbatasan ruang pada pemerintah tambak-tambak dan anggaran. Kebanyakan tambak-tambak cenderung bertelur seperti banyak ikan saat dimungkinkan, sehingga kadangkala sulit untuk meningkatkan Ne cara ini.
Kedua adalah pendekatan untuk bertelur aqual yang lebih sex rasio, memberikan kisaran 50: 50. Efek miring sex rasio di perkawinan sedarah dapat ditunjukkan oleh hasil tersebut :
F = ♀ + ♂
Di mana (♀ ) adalah jumlah betina yang menghasilkan keturunan dan (♂ ) adalah jumlah jantan yang menghasilkan keturunan.
Ketika berkembang biak populasinya sedang kecil, seks rasio miring bisa lebih rendah Ne dan meningkatkan perkawinan sedarah secara dramatis (gambar .4.18). Contoh berikut menunjukkan kenyataan ini:
Populasi 1: 25 betina dan 25 jantan
F = F = 1% generasi
Populasi 2: 250 betina dan 10 jantan F =
F = 1,3 % generasi
Populasi 2 memiliki lebih dari 5 kali banyak peternakan individu, tetapi Ne populasi 1 adalah 50 sementara Ne populasi 2 adalah dlm hati 38,5. sebagai hasilnya, perkawinan sedarah yang dihasilkan oleh penduduk 2 adalah 30% lebih besar karena rasio jenis kelamin miring. Tabel 4.13 memberikan F diproduksi dalam satu generasi dengan berbagai kombinasi dari laki-laki anf perempuan.
Sering kali ada godaan besar untuk menggunakan rasio seks miring karena produksi fingerling dioptimalkan dalam hal broodfish paling sedikit diperlukan untuk mencapai kuota produksi fingerling. Bondari (1983b) menunjukkan bahwa petani lele yang menggunakan teknik pemijahan kolam terbuka dapat condong rasio jenis kelamin ikan lele broodfish hingga 4 betina: 1 jantan dan tidak mempengaruhi menggoreng produksi. Produksi ini, namun masalah genetik yang ada hanya akan bertambah buruk.
Sebagai contoh, jika seorang petani lele membutuhkan 50 telur massa, perkawinan sedarah yang dihasilkan oleh rasio dua seks
1:1 sex ratio 50 ♀ : 50 ♂ F = F = 0.5 % / generation 4 : 1 sex ratio 50 ♀ : 12.5 or 13 ♂ F = F = 1.21 % / generation
Pendekatan ketiga untuk memaksimalkan Ne adalah untuk beralih dari
kawin acak untuk keturunan baik kawin. Kawin keturunan baik berbeda dari perkawinan acak di setiap perempuan meninggalkan satu anak perempuan dan laki-laki masing-masing daun satu anak untuk digunakan sebagai induk pada generasi berikut. laki-laki yang menumbuhkan dengan sepuluh perempuan daun sebanyak anak sebagai laki-laki memunculkan dengan satu perempuan-satu. Putra dan putri yang dipilih secara acak dari dalam setiap keluarga. Sistem pembibitan ini dapat melipatgandakan Ne tanpa meningkatkan ukuran populasi Ne dengan
kawin keturunan baik adalah Ne =
Tabel 4.13
Jika ada lebih banyak perempuan, penyebut adalah 3 () + () jika ada lebih laki-laki, penyebut adalah () + 3 (). Jumlah peternakan efektif peningkatan bila Anda menggunakan kawin keturunan baik, karena Anda artifisial meningkatkan variasi genetik dengan memastikan bahwa setiap brooder diwakili pada generasi berikutnya.
Satu satunya kelemahan untuk keturunan perkawinan adalah bahwa anda harus mampu mengidentifikasi individu ikan .Meskipun beberapa sistem menandai telah dirancang (Anon. 1956; Clemens and Sneed 1959; Moav et al.
1960A, 1960B; Groves and Novotny 1965; Monan 1966; Volz and Wheeler 1966; Everest and Edmundson 1967; Fujihara and 1975; Rinne 1976; Joyce and El-Ibiary 1977; Welch and Mills 1981), kebanyakan tambak tambak tidak bisa untuk menandai ikan atau untuk memisahkan famiies sampai mereka dapat ditandai .
Ketika pengembangbiakan kependudukan tidak dapat diganti atau meningkat dalam ukuran , satu-satunya cara untuk meningkatkan produktivitas adalah baik untuk mengubah sex rasio atau untuk beralih ke perkawinan keturunan .Manfaat diproduksi oleh perubahan sex rasio atau dalam program pengembangbiakan dapat diukur sebagai efektif efisiensi penduduk berkembang biak. :
Nb =
di mana Nb adalah efektif berkembang biak efisiensi dan n adalah ukuran
populasi.
Dalam kendala tetap ukuran populasi n, nb memungkinkan anda untuk menentukan effeciency bahwa sesungguhnya sex rasio atau program pengembangbiakan memiliki memaksimalkan ne relatif terhadap kelamin lain rasio program atau berkembang biak.Tokoh 4.20 menunjukkan nb untuk semua kemungkinan seks rasio untuk kedua acak dan keturunan perkawinan.Misalnya, jika trout petani menumbuhkan 90 perempuan dan 10 laki laki ikan trout pelangi oleh perkawinan acak, dia memiliki sebuah nb = 36 %.Dengan menyesuaikan seks nya rasio 70 untuk perempuan dan 30 laki laki, dia akan menambahkan nb 84 untuk %.Yang 70: 30 sex rasio 2.3 kali lebih efisien dalam memaksimalkan ne, dan perkawinan sedarah akan hanya % sebagai besar 42.Indeks ini menunjukkan petani yang ia masih dapat menggunakan sebuah miring sex rasio fot mengoptimalkan fingerling produksi, tetapi jika ia moderat rasio dia bisa meningkatkan nya ne, dan itu quantifies peningkatan.
Gambar 4.20
Efisiensi pemuliaan yang efektif juga dapat menginformasikan manajer pembenihan bahwa ia melakukan pekerjaan yang baik dalam keterbatasan pembenihan nya. Sebagai contoh, jika kawin acak digunakan, rasio jenis kelamin 55:45 memiliki Nb dari 99%, sehingga beralih ke rasio 50:50 seks untuk mencapai Nb dari 100% mungkin tidak berharga. The kawin parabola acak digambarkan dalam Gambar. 4.20 menunjukkan bahwa perubahan dalam rasio jenis kelamin yang lebih miring dari 60:40 akan menghasilkan perbaikan yang lebih kecil.
Pemuliaan yang efektif juga memberikan sepotong informasi tambahan yang sangat penting dalam proses pengambilan keputusan. Efisiensi pemuliaan yang efektif mengkuantifikasi efek yang diusulkan perubahan rasio jenis kelamin akan memiliki produksi fingerling. Indeks mengkuantifikasi berapa banyak perempuan yang lebih sedikit akan melahirkan, sehingga sangat posible untuk
menghitung berapa banyak bibit yang lebih sedikit akan diproduksi. Jika penurunan ini diterima, maka chage diusulkan dalam rasio jenis kelamin akan menurunkan produksi fingerling ke tingkat yang tidak dapat diterima, baik perubahan rasio jenis kelamin dapat moderat rata berat badan perempuan dapat ditingkatkan sehingga jumlah yang sama telur akan diproduksi. Dengan demikian, Anda berdua bisa memuaskan kuota produksi fingerling dan juga meningkatkan genetika populasi.
Gambar 4.20 juga menunjukkan keuntungan dari pengeluaran usaha ekstra yang terlibat dalam beralih ke silsilah kawin. Rasio 79:21 seks dengan kawin keturunan baik memiliki Nb besar daripada rasio 50:50 seks dengan kawin acak, dalam kendala dari ukuran populasi tetap: 102% vs 100%. Dengan demikian, dalam kendala dari ukuran populasi tetap, penggunaan silsilah kawin dapat menghasilkan Nb yang lebih besar yang mungkin dapat diproduksi oleh perkawinan acak. Anda bahkan dapat menghasilkan Nb dari 200% dengan kawin keturunan baik (50:50 rasio jenis kelamin).
Ingatlah, semua yang penyesuaian rasio jenis kelamin dan program pemuliaan melakukan i mencegah perkawinan sedarah dan pergeseran genetik dari semakin buruk. Mereka tidak merekonstruksi gen yang rusak. Jika Anda memastikan bahwa youre Ne tidak melalui constinctions atau kemacetan, penyesuaian ini dalam manajemen induk akan membantu menjaga masalah genetik di cek. Satu-satunya obat untuk masalah yang disebabkan oleh penyempitan atau hambatan adalah untuk memperoleh induk baru dan baik menggantikan populasi atau menyeberang dengan indukan baru.
BAB III PENUTUP
3.1 Kesimpulan
Inbreeding digunakan untuk menggambarkan berbagai fenomena yang terkait dengan persilangan antar kerabat dekat yang dapat meningkatkan homozigositas genotipe. Sehingga seringkali hasil dari Inbreeding adalah memperlihakan berkurangnya keragaman genetik sehingga dapat menyebabkan penurunan pertumbuhan dan peningkatan kepunahan suatu organisme. Namun, meski demikian, Inbreeding juga ternyata digunakan untuk meningkatkan populasi. Salah satu penggunaan utama perkawinan sekerabat adalah dalam sebuah program perkembangbiakan yang disebut linebreeding. Linebreeding terjadi ketika individu yang menonjol bagus (biasanya laki-laki) dibawa kembali ke garis untuk kawin dengan keturunan. . Ini dilakukan karena hewan yang luar biasa ini ingin Anda tingkatkan kontribusinya untuk masing-masing keturunan dan meningkatkan kontribusinya terhadap gen yang jelek. Adapun untuk mengetahui hasilnya, nilai-nilai individu Inbreeding dapat dihitung dengan menggunakan teknik yang disebut analisis jalur.
3.2 Saran
Inbreeding tidak selamanya membawa dampak buruk, Inbreeding
dilakukan untuk mendapatkan kemurnian dari strain yang sudah stabil atau mendapatkan fenotip dari gen resesif yang tidak muncul pada indukannya.
Inbreeding yang dimaksud adalah pemijahan sesame anakan dalam satu lubuk
atau antara anak dan indukannya. Dalam proses pengembangan dengan teknik
Inbreeding ini sebaiknya disiapkan selalu 2 lini, jika terjadi hal-hal yang tidak
DAFTAR PUSTAKA
Tave, Douglas, 1986. Genetics for Fish Hatchery Managers, ACI Publishing Company, Inc. West Port. Conecticut.