7
TINJAUAN PUSTAKA
Ekosistem KarstKarstifikasi
Kata karst berasal dari bahasa Yugoslavia dan diperkenalkan oleh Cvijic seorang geolog asal Jerman pada tahun 1850. Kata karst tersebut mengacu pada kawasan batu gamping di Kota Trieste, Slovenia, Yugoslavia (Wirawan 2005). Sampai saat ini, kata karst telah digunakan secara internasional dan telah diserap secara utuh sebagai kata bahasa Indonesia. Salah satu definisi karst yang dikemukakan oleh ahli geologi adalah bentang alam (landscape) pada lempeng batuan gamping yang dibentuk oleh pelarutan batuan gamping. Pelarutan batu gamping tersebut menghasilkan bentukan karst dengan ciri celah sinkhole (lubang lari air), sungai bawah tanah, dan gua (Hamilton & Smith 2006; Samodra 2006).
Proses terbentuknya karst (karstifikasi) berlangsung selama jutaan tahun melalui peristiwa yang melibatkan faktor-faktor geologi, fisika, kimia, dan biologi. Karstifikasi diawali dengan pergerakan lempeng bumi yang bersifat dinamis. Pergerakan lempeng bumi tersebut menyebabkan lempeng saling bertabrakan dan menghasilkan gaya tektonik yang mendorong sebagian lempeng ke atas. Peristiwa ini menyebabkan sedimentasi sisa-sisa tumbuhan dan hewan yang mengandung kapur (kalsium karbonat) terangkat dari dasar laut ke permukaan (Gimes 2001). Menurut Yunqiu et al. (2006) biota laut tersebut antara lain, koral (Pontes, Neandrina, Acropora, Siderastrea, Ginoid), Briozoa, ganggang (Halimeda, Lithothamniam, Penicillus, Acialaria, Neomen), Foraminifera, dan Moluska. Peristiwa yang disebabkan oleh gaya tektonik ini menghasilkan deretan bukit kapur/gamping di permukaan laut. Gaya-gaya tektonik tersebut dapat menyebabkan terjadinya patahan dan retakan yang saling berasosiasi. Lempeng batuan yang terdeformasi oleh gaya-gaya tektonik ini merupakan area yang sangat potensial untuk masuknya aliran air dan terbentuknya perangkap-perangkap air (Eberhard 2006). Formasi awal terbentuknya karst tersaji pada Gambar 2.
8
Gambar 2 Formasi awal terbentuknya karst (Sumber: Subterra 2004)
Setelah proses yang disebabkan oleh gaya tektonik, peristiwa selanjutnya adalah pelarutan batuan karbonat oleh asam lemah. Reaksi karbon dioksida (CO2) di udara dengan air hujan (H20) menghasilkan H2CO3 yang bersifat asam
lemah. Larutan tersebut mengalir melalui aliran air permukaan (run off) dan akan melarutkan batu gamping sehingga terbentuk celah. Lebih rinci Samodra (2006) menjelaskan reaksi kimia pelarutan batu gamping oleh asam lemah adalah sebagai berikut :
H2O + CO2 H2CO3
H2CO3 HCO3 + H+
HCO3 + CaO CaCO3 + H2O
CaCO3 + H2O + CO2 CaH2C2O6
Celah yang dihasilkan oleh pelarutan tersebut semakin besar dari waktu ke waktu sampai membentuk patahan dan rongga yang disebut karen (patahan), sinkhole (lubang lari air), collapse sink/doline (rongga), dan gua (Gimes 2001). Gaya tektonik yang terjadi pada masa berikutnya menyebabkan rongga dan gua saling berasosiasi satu sama lain membentuk sistem perguaan dengan lorong yang panjang (Samodra 2006). Persyaratan yang harus dipenuhi supaya lempeng
9
batu gamping dapat membentuk morfologi karst menurut Hamilton & Smith (2006) adalah : 1) lempeng batuan gamping mempunyai ketebalan yang cukup, 2) berada di wilayah dengan curah hujan tinggi, 3) batuan gamping banyak mengandung celah atau rongga, 4) berada pada posisi lebih tinggi dibandingkan lingkungan di sekitarnya.
Geomorfologi karst
Ahli geologi membagi geomorfologi karst menjadi karst luar atau exokarst dan karst dalam atau endokarst (gua). Exokarst/epikarst dicirikan dengan: 1) adanya bukit-bukit kapur berbentuk kerucut atau kubah, 2) permukaan kasar berlubang-lubang membentuk dolina (cekungan), dan 3) adanya endapan sedimen lempung berwarna merah hasil pelapukan batu gamping (Samodra 2006). Selain itu menurut Roemantyo & Noerdjito (2006), exokarst biasanya tertutup oleh lapisan tanah yang tipis yang umumnya berasal dari batuan kapur yang hancur dan terdekomposisi secara mekanik dan kimiawi. Lapisan tanah tipis tersebut sebagian terkumpul pada cekungan. Proses pengayaan nutrisi pada lapisan tanah exokarst dapat terjadi oleh adanya debu vulkanis, ataupun aliran air hujan yang membawa humus dari tempat lain. Akibatnya exokarst dapat ditumbuhi oleh jenis-jenis tumbuhan tertentu.
Endokarst (gua) menurut Hamilton & Smith (2006) merupakan ruang
dengan sirkulasi udara terbatas dan sangat sedikit atau bahkan sama sekali tidak ada cahaya. Selain itu, menurut Wirawan (2005), ruang dalam gua biasanya dilengkapi dengan ornamen-ornamen gua. Ornamen tersebut merupakan hasil pengendapan kalsium karbonat (CaCO3) yang sebelumnya terlarutkan oleh
peristiwa karstifikasi. FINSPAC (1993) membagi ornamen-ornamen dalam gua menjadi: 1) stalaktit, yaitu endapan kapur yang menggelantung pada langit-langit gua, 2) stalakmit, yaitu endapan kapur yang terdapat pada lantai gua, 3) tiang (column), yaitu pertemuan antara stalaktit dan stalakmit yang membentuk tiang, 4) tirai (drip curtain/drapery), yaitu endapan yang berbentuk lembaran tipis vertikal, pada atap gua yang miring, dan 5) teras (travertin), yaitu endapan kalsium karbonat pada lantai gua yang bertingkat sehingga membentuk
10
Gambar 3 Geomorfologi karst (Sumber: FINSPAC 1993)
Hamilton & Smith (2006) membagi lingkungan dalam gua berdasarkan pengaruh sinar matahari menjadi 3 mintakat, yaitu 1) mintakat I adalah mintakat di dalam gua yang sinar matahari dapat masuk dan iklim dalam gua masih dipengaruhi oleh iklim luar gua, 2) mintakat II adalah mintakat di dalam gua yang tidak ada sinar matahari yang masuk, tetapi iklim di dalam mintakat tersebut masih dipengaruhi oleh iklim di luar gua, dan 3) mintakat III adalah mintakat yang tidak ada sinar matahari dan iklim di dalam mintakat ini stabil, tidak dipengaruhi oleh fluktuasi iklim di luar gua. Contoh denah pembagian mintakat pada Gua Jatijajar dapat dilihat pada Gambar 4.
Menurut Russo et al. (2003) dinding dan atap gua merupakan penyangga efektif yang memisahkan lingkungan dalam gua dengan lingkungan luar gua. Oleh karenanya, lingkungan dalam gua memiliki mikroklimat yang berbeda dari luar gua. Menurut Samodra (2006) mikroklimat dalam gua cenderung lebih dingin dan lebih lembap. Hal ini karena 1) adanya aliran sungai di lantai gua; 2) adanya air rembesan di atap gua; 3) tidak ada sinar matahari, dan 4) sirkulasi udara terbatas. Selain itu menurut Russo et al. (2003) mikroklimat tersebut dapat berbeda antara satu zona (mintakat) dan zona lainnya. Hal ini dipengaruhi oleh pengaruh sinar matahari, formasi gua, dan keberadaan mahluk hidup di dalamnya. Penelitian Baudinette et al. (1994) di Gua Kelelawar dan Gua
11
Robertson Afrika Selatan membuktikan gua yang dihuni kelelawar dengan jumlah besar dapat menaikkan suhu dalam gua hingga 3oC.
Gambar 4 Denah pembagian mintakat pada Gua Jatijajar (Sumber : DISPARBUD Kabupaten Kebumen 2004 yang telah dimodifikasi)
Tipe-tipe gua
Aliran air merupakan faktor utama dalam pembentukan gua karst. Karenanya menurut Hamilton & Smith (2006), berdasarkan penetrasi air pada dinding dan atap gua, dapat dibedakan tiga-tipe gua karst, yaitu 1) gua fosil, adalah gua karst yang pada dinding dan atapnya tidak ada lagi penetrasi air. Hal ini menyebabkan pertumbuhan ornamen-ornamen gua terhenti dan mikroklimat gua cenderung lebih kering dibandingkan tipe gua karst lainnya; 2) gua vedosa: gua karst yang berada pada sarang air (water table) yang datar. Tipe gua ini ditandai dengan sedikitnya penetrasi air pada atap gua sehingga tidak banyak ditemukan ornamen gua. Oleh karena berada pada sarang air datar, maka banyak terbentuk mata air di dinding gua. Banyaknya mata air tersebut menyebabkan mikroklimat di dalam gua cenderung dingin dan lembap; 3) gua pheartic adalah gua karst yang berada pada bidang miring, yang penetrasi air pada atap gua berlangsung sangat efektif. Tipe gua ini ditandai dengan banyaknya tetesan air
12
pada atap gua dan biasanya lantai gua dilalui sungai bawah tanah. Ornamen gua membentuk formasi yang kompleks dan masih aktif berkembang. Keberadaan sungai bawah tanah dan banyaknya tetesan air pada atap gua menyebabkan gua
pheartic dingin dan lembap (Samodra 2006).
Pada gua fosil, vedosa maupun pheartic terbentuk zonasi atau mintakat-mintakat. Mintakat tersebut sangat dipengaruhi oleh besarnya mulut gua, banyaknya ventilasi gua maupun formasi gua. Gua tipe pheartic memiliki formasi gua yang lebih kompleks dibandingkan tipe gua lainnya (Samodra 2006). Hal ini disebabkan lorong gua yang berliku-liku, adanya aliran sungai, dan banyaknya ornamen-ornamen gua. Kondisi ini menghasilkan mintakat III yang lebih luas, yaitu mintakat gelap dengan suhu dan kelembapan stabil tidak dipengaruhi suhu luar gua.
Gua dapat diartikan sebagai ruang /lorong yang berada di bawah permukaan tanah. Selain tipe-tipe gua karst yang telah dijelaskan di atas, menurut Ko (2004) terdapat tiga tipe gua lain, yaitu 1) gua lava: gua yang terbentuk karena aktivitas vulkanik atau gunung berapi, 2) gua litoral /gua laut: gua yang terbentuk kerena gelombang laut yang memecah karang di pantai, dan 3) gua sandstone: gua yang terbentuk karena erosi air dan angin pada batuan pasir. Ketiga tipe gua tersebut memiliki lorong pendek, formasi gua sederhana, dan cahaya matahari dapat masuk hampir keseluruh bagian lorong gua. Karena itu tidak terdapat mintakat-mintakat seperti halnya pada gua karst dan tidak membentuk ekosistem yang kompleks.
Komunitas fauna gua karst
Dinding dan atap gua merupakan pembatas yang memisahkan lingkungan dalam gua dengan luar gua. Dinding dan atap tersebut tidak tembus sinar matahari. Akibatnya, kondisi dalam gua menjadi gelap dan tumbuhan hijau (autotrof) tidak ditemukan. Meskipun demikian, menurut Ko (2004), ruang dalam gua dapat ditempati oleh mahluk hidup. Hal ini karena sumber energi didatangkan dari luar gua melalui unsur hara yang terlarut dalam aliran air, debu zat-zat organik yang terbawa oleh udara serta bahan nutrisi yang berasal dari
13
hewan yang bersarang di dalam gua tetapi mencari makan di luar gua (hewan
Troglozene).
Menurut Ko (2004), di kawasan karst penghubung utama antara ekosistem luar gua dan ekosistem dalam gua adalah burung dan Mamalia. Jenis-jenis burung di antaranya adalah walet (Aerodramus fuciphagus) dan sriti (Hirundo
tahitica), sedangkan kelompok Mamalia adalah ordo Chiroptera (kelelawar).
Menurut Whitten et al. (1999) dan Sinaga et al. (2006) fauna troglozene utama di gua-gua karst di Pulau Jawa adalah kelelawar. Bahkan jumlah populasi kelelawar tersebut dapat mencapai jutaan individu dalam satu gua. Secara sederhana suplai energi ke dalam gua disajikan dalam Gambar 5.
Gambar 5 Suplai energi ke dalam gua (Sumber: Subterra 2004)
Berdasarkan sumber energinya, jenis-jenis fauna yang hidup di gua menurut Ko (2004) dibedakan menjadi: 1) necrophagus, yaitu fauna pemakan bangkai 2)
cocroaphagus, yaitu fauna pemakan kotoran/feses 3) parasit, yaitu fauna yang
hidup pada fauna lain dan 4) predator, yaitu fauna pemakan fauna lain. Penelitian McCure (1985) di Gua Batu Malaysia mendapatkan necrophagus terdiri atas : lalat (Muscoidae:Insekta) dan semut (Formicidae: Insekta); cocroaphagus terdiri atas ekor pegas (Collembola: Insekta/Hexapoda), kumbang (Stratiomyiidae: Insekta), kecoa (Blattidae: Insekta), kumbang (Tineidae: Insekta), jangkerik (Gryllothalpidae: Insekta) dan jangkerik (Gryllidae: Insekta); parasit terdiri atas :
14
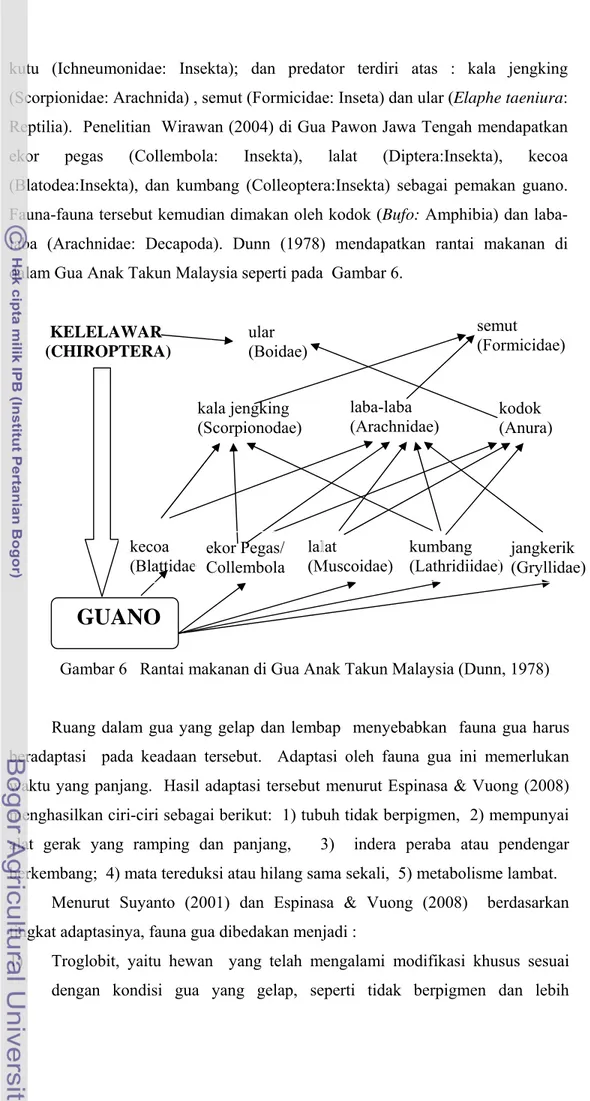
kutu (Ichneumonidae: Insekta); dan predator terdiri atas : kala jengking (Scorpionidae: Arachnida) , semut (Formicidae: Inseta) dan ular (Elaphe taeniura: Reptilia). Penelitian Wirawan (2004) di Gua Pawon Jawa Tengah mendapatkan ekor pegas (Collembola: Insekta), lalat (Diptera:Insekta), kecoa (Blatodea:Insekta), dan kumbang (Colleoptera:Insekta) sebagai pemakan guano. Fauna-fauna tersebut kemudian dimakan oleh kodok (Bufo: Amphibia) dan laba- laba (Arachnidae: Decapoda). Dunn (1978) mendapatkan rantai makanan di dalam Gua Anak Takun Malaysia seperti pada Gambar 6.
GUANO
Gambar 6 Rantai makanan di Gua Anak Takun Malaysia (Dunn, 1978)
Ruang dalam gua yang gelap dan lembap menyebabkan fauna gua harus beradaptasi pada keadaan tersebut. Adaptasi oleh fauna gua ini memerlukan waktu yang panjang. Hasil adaptasi tersebut menurut Espinasa & Vuong (2008) menghasilkan ciri-ciri sebagai berikut: 1) tubuh tidak berpigmen, 2) mempunyai alat gerak yang ramping dan panjang, 3) indera peraba atau pendengar berkembang; 4) mata tereduksi atau hilang sama sekali, 5) metabolisme lambat.
Menurut Suyanto (2001) dan Espinasa & Vuong (2008) berdasarkan tingkat adaptasinya, fauna gua dibedakan menjadi :
1) Troglobit, yaitu hewan yang telah mengalami modifikasi khusus sesuai dengan kondisi gua yang gelap, seperti tidak berpigmen dan lebih
KELELAWAR
(CHIROPTERA)
lalat
(Muscoidae) kumbang (Lathridiidae) jangkerik (Gryllidae) kecoa (Blattidae kala jengking (Scorpionodae) laba-laba (Arachnidae) semut (Formicidae) ular (Boidae) kodok (Anura) ekor Pegas/ Collembola
15
berfungsinya indera peraba, penciuman, dan pendengaran. Troglobit merupakan penghuni tetap gua yang tidak dapat hidup di habitat lain. Oleh karena itu, hewan troglobit merupakan kelompok yang paling fragil di antara kelompok lainnya. Espinasa & Vuong (2008) mendapatkan serangga troglobit: Nicoletiid (Zygentoma: Insecta) di Gua Oaxaca, Mexico. Menurut Whitten et al. (1999) fauna troglobit yang sering ditemukan di gua-gua karst di Pulau Jawa adalah kepiting (Sesarmoides jacobsoni: Crustacea ), Udang putih (Macrobrachium poeti: Crustacea) dan ikan buta (Puntius binotatus: Osteicthyes). Penelitian Wijayanti (2001) di Gua Petruk dan Gua Jatijajar Jawa Tengah mendapatkan fauna troglobit: ikan buta (Amblyopsis spelaeus: Osteicthyes), udang gua (Macrobrachium pilimanus: Crustacea), laba-laba gua (Stigophrynus darmamani: Arachnidea), dan kumbang gua (Eustra saripaensis: Insekta). Hasil penelitian Rachmadi (2003) di gua Karst Ngerong, Tuban, Jawa Timur, mendapatkan fauna troglobit: kalajengking gua (Chaerilus sabinae: Scorpionidae), kepiting gua (Cancrocaeca xenomorpha: Cruatacea), kepiting mata kecil (Sesarmoides
emdi: Crustacea), isopoda gua (Cirolana marosina: Isopoda), kumbang gua
(Eustra saripaensis:Insecta), dan ekor pegas gua (Pseudosinella maros: Insecta).
2) Troglozene, yaitu fauna yang secara teratur masuk ke dalam gua untuk berlindung, beristirahat, dan berkembang biak, tetapi mencari makan di luar gua. Meskipun hanya sebagian hidupnya berada di dalam gua, hewan troglozene telah beradaptasi dengan kondisi gua yang gelap. Menurut Vermeulen & Whitten (1999), fauna troglozene mempunyai kemampuan
echolokasi, yaitu kemampuan menangkap gelombang pantul (gema)
berfrekuensi ultrasonik (>20 KHz). Echolokasi ini berguna untuk mendeteksi mangsa dan orientasi ruang tanpa mengunakan mata. Kelompok fauna troglozene merupakan spesies kunci dalam ekosistem gua, karena fauna troglozene memindahkan energi dari luar gua ke dalam gua. Fauna troglozene yang sering ditemukan di gua karst di Indonesia adalah burung walet (Collocalia fuciphaga/Aerodramus fuciphagus), burung sriti (Hirundo tahitica), dan kelelawar (ordo Chiroptera) (Whitten et al. 1999).
16
3) Troglophil, yaitu fauna yang hidup di dalam gua, tetapi belum mengalami modifikasi khusus. Fauna ini selama hidupnya berada dalam gua, tetapi jenis yang sama juga ditemukan di luar gua. Bila terjadi gangguan di dalam gua, fauna troglophil dapat pindah ke habitat luar gua. Penelitian Castillo et
al. (2009) di Los Ricos Cave, Queretaro, Mexico mendapatkan kodok
(Eleutherodactylus longipes: Anura) sebagai fauna troglophil yang secara musiman memasuki gua. Menurut Whitten et al. (1999) jangkerik (Rhapidophora dammarmani: Insekta), kumbang (Collasoma scrutater: Insekta), laba-laba (Liphistius sp: Arachnidae), dan keong (Thiara scabra: Gastropoda) merupakan troglophil yang sering dijumpai di gua-gua karst di Pulau Jawa.
Kelelawar sebagai Kelompok Fauna Troglozene
Kelelawar merupakan fauna troglozene utama di gua-gua karst di Indonesia (Whitten et al.1999; dan Suyanto 2001). Kelelawar adalah Mamalia yang termasuk dalam ordo Chiroptera. Ciri khas ordo ini adalah tulang telapak tangan (metacarpal) dan tulang jari (digiti) mengalami pemanjangan sehingga berfungsi sebagai kerangka sayap. Sayap tersebut terbentuk dari selaput tipis (petagium) yang membentang antara tulang-tulang telapak dan jari tangan sampai sepanjang sisi tubuh (Nowak 1994; Altringham 1996). Nowak (1994) menggambarkan struktur rangka kelelawar seperti terlihat pada Gambar 7.
Gambar 7 Struktur rangka kelelawar (Sumber : Nowak 1994) digiti 1 digiti 2 digiti 3 digiti 4 digiti 5 petagium
17
Ordo Chiroptera terdiri atas 2 subordo, yaitu Megachiroptera dan Microchiroptera. Kedua subordo ini diduga tidak mempunyai hubungan kekerabatan dan merupakan hasil evolusi konvergen, yaitu evolusi yang terjadi pada dua spesies yang berbeda tetapi beradaptasi dengan cara yang sama sehingga menghasilkan morfologi yang mirip (Altringham 1996). Salah satu alasan yang mendukung adalah : saraf superior colliculus (s.c) kanan pada otak tengah Microchiroptera mengatur retina mata kiri dan sebaliknya s.c kiri mengatur retina mata kanan. Hal ini ditemukan pada semua Mamalia, kecuali primata. Pada Megachiroptera, saraf superior colliculus kanan otak tengah mengatur retina mata kiri dan mata kanan sekaligus. Keadaan ini hanya ditemukan pada Primata, Dermoptera, dan Megachiroptera (Corbet & Hill 1992; Altringham 1996). Karena alasan tersebut maka diduga Megachiroptera berasal dari nenek moyang Primata, sedangkan Microchiroptera diduga berasal dari nenek moyang bukan Primata. Penelitian HanGuan et al. (2006) tentang philogenetika kelelawar juga mendapatkan bahwa kelelawar Megachiroptera memiliki kekerabatan lebih dekat dengan primata dibandingkan dengan Microchiroptera. Saat ini diketahui terdapat 18 famili, 192 genus dan sekitar 1111 jenis kelelawar yang ada di dunia (Safi & Kerth 2004). Menurut Suyanto et al. (1998) terdapat 10 famili, 49 genus, dan sekitar 151 jenis terdapat di Indonesia.
Anggota subordo Megachiroptera makanan utamanya adalah buah (frugivora), selain itu juga memakan serbuk sari (polen) dan nektar. Subordo ini terdiri atas 1 famili, yaitu Pteropodidae dengan 42 genus dan 166 spesies (Nowak 1994). Menurut Altringham (1996) anggota subordo Megachiroptera memiliki ukuran yang relatif besar (bobot minimum 10 gram maksimum 1500 gram dengan bentangan sayap maksimum 1700 mm); memiliki mata besar; telinga tidak memiliki tragus; moncong sederhana dan ekor tidak berkembang; jari kedua dan jari ketiga terpisah relatif jauh dan memiliki cakar pada jari kedua, kecuali pada
Eonycteris, Dobsonia, dan Neopterix.
Anggota subordo Microchiroptera kebanyakan pemakan serangga (insectivora). Selain itu, ada juga yang penghisap darah (sanguivora), misalnya
Desmodus vampirus; dan penghisap madu misalnya (Leptonycteris curasoae).
18
Microchiroptera adalah berukuran kecil (bobot minimum 2 gram, maksimum 196 gram dengan bentangan sayap maksimum 70 mm); memiliki mata kecil; telinga memiliki tragus (tonjolan dari dalam daun telinga) atau anti tragus (tonjolan dari luar daun telinga); jari sayap tidak bercakar dan moncong sangat bervariasi, terutama famili Rhinolophidae dan Hipposideridae memiliki daun hidung (noselea) yang kompleks.
Klasifikasi kelelawar menurut Corbet & Hill (1992) adalah sebagai berikut : Kingdom : Animalia
Filum : Chordata Sub Filum : Vertebrata Kelas : Mamalia Ordo : Chiroptera Subordo : Megachiroptera
Famili : Pteropodidae Subordo : Microchiroptera
Famili : Rhinolophidae, Hipposideridae, Megadermatidae, Craseonycteridae,
Rhinopomatidae, Nycteridae, Emballonuridae, Phyllostomidae, Mormoopidae, Noctilionidae, Furipteridae, Thyropteridae, Mystacinidae, Myzopodidae, Vespertilionidae, Molosidae dan Natalidae
Menurut Nowak ( 1994), kelelawar ditemukan di seluruh permukaan bumi, kecuali di daerah kutub dan pulau-pulau terpencil. Kemampuan terbang kelelawar merupakan faktor penting dalam persebaran hewan ini. Selain itu, jenis pakannya sangat bervariasi sehingga memungkinkan hidup di berbagai tipe habitat. Menurut Altringham (1996), sekitar 200 spesies kelelawar ditemukan di Madagaskar dan Afrika; 300 spesies ditemukan di Amerika Selatan dan Amerika Tengah; 240 jenis ditemukan di Asia dan Australia; dan sekitar 40 spesies ditemukan di Amerika Utara dan Eropa. Menurut Suyanto et al. (1998), di Indonesia terdapat 151 jenis kelelawar. Jenis-jenis tersebut menyebar di seluruh kepulauan
19
Indonesia. Lebih lanjut Kunz & Pierson (1994) menjelaskan bahwa kelelawar merupakan Mamalia paling berhasil, karena dapat ditemukan di berbagai tipe habitat dengan ketinggian mulai 10 m dpl sampai 3000 m dpl. Winkelmann et al. (2000) meneliti penggunaan habitat oleh kelelawar Synconycteris australis di Papua New Guinea. Menurut Winkelmann et al. (2000) faktor-faktor yang dapat mempengaruhi keberadaan dan kelimpahan kelelawar pada suatu habitat ialah 1) struktur fisik habitat, 2) iklim mikro habitat, 3) ketersediaan pakan dan sumber air, 4) keamanan dari predator, 5) kompetisi, dan 6) ketersediaan sarang.
Perilaku bersarang
Sarang merupakan salah satu komponen penting dalam hidup kelelawar. Kebanyakan jenis kelelawar hidup berkoloni dalam bersarang dan pencarian makan. Menurut Zukal et al. (2005) beberapa keuntungan hidup dalam koloni adalah adanya transfer informasi, keamanan pada predator, keberhasilan reproduksi, dan thermoregulasi. Altringham (1996) menjelaskan tiga perilaku produk transfer informasi yang dilakukan dalam koloni kelelawar, yaitu 1) mengikuti (following behaviour), yaitu perilaku yang menyebabkan anggota dalam koloni bersama-sama menuju suatu lokasi tempat pencarian makan atau tempat bersarang; 2) penanda hubungan sosial (sosial signal), yaitu pemahaman signal-signal intensional, termasuk signal tanda bahaya; 3) belajar (learning
behaviour), yaitu proses pembelajaran dari induk ke anak yang menyebabkan
kelelawar muda mampu mengembangkan teknik pencarian makan, menghindar dari predator, serta hal-hal yang menguntungkan bagi kehidupannya. Willis & Brigham (2004) meneliti pembagian sarang (roost sharing) dan kebersamaan sosial (sosial cohesion) kelelawar Eptesicus fuscus (Microchiroptera) di Cypres Hill Canada. Hasil penelitian membuktikan bahwa interaksi sosial dan kerja sama intraspesifik dalam koloni dapat menghasilkan ketahanan terhadap gangguan predator dan cuaca buruk. Penelitian Baudinette et al. (1994) di Gua Kelelawar dan Gua Robertson Australia membuktikan gua yang dihuni kelelawar dengan jumlah besar dapat menaikkan suhu dalam gua hingga 3oC. Pada musim dingin, keadaan ini menguntungkan kelelawar karena mengurangi energi yang diperlukan untuk menghangatkan tubuh.
20
Setiap jenis kelelawar mempunyai beberapa alternatif dalam memilih lokasi sarang, di antaranya adalah pohon yang tinggi, di balik batu, di atap rumah, dan di dalam gua. Menurut Altringham (1996), pemilihan sarang mempengaruhi distribusi lokal dan global, kepadatan, strategi pencarian makan, strategi kawin, struktur sosial, dan pergerakan musiman. Menurut Zahn & Hager (2005) proses yang terlibat dalam memilih tempat bersarang cukup kompleks. Ketersediaan tempat bersarang yang cocok misalnya, akan mempengaruhi perilaku pencarian makan, tetapi perilaku bersarang sendiri juga dipengaruhi oleh kelimpahan dan penyebaran makanan.
Menurut Baudinette et al. (1994), Russso et al. (2003), dan Willis & Brigham (2004), dengan memilih sarang, kelelawar dapat memperoleh beberapa keuntungan, yaitu perlindungan dari cuaca buruk, perlindungan dari predator, memperkecil energi termoregulasi, keberhasilan reproduksi, serta transfer informasi tempat mencari makan dan tempat bersarang. Penelitian Willis & Brigham (2004) dan juga penelitian Seckerdieck et al. (2005) membuktikan bahwa kelelawar mempunyai home instink yang kuat, sarang yang dipilih kelelawar dipertahankan sampai beberapa generasi. Namun demikian apabila sarang mendapat ganggguan dan kelelawar tidak nyaman dan aman, sarang ini akan ditinggalkan (Willis & Brigham 2004).
Menurut Altringham (1996), kebanyakan kelelawar pemakan buah (Megachiroptera) bersarang di pohon dengan jumlah koloni besar. Pohon sarang Megachiroptera biasanya tinggi dan besar, tetapi tidak berdaun rimbun. Menurut Campbell et al. (1996), pohon tempat bersarang kelelawar biasanya menyediakan akses yang mudah menuju tempat pencarian makan (central place foraging) dan mempunyai pencahayaan yang cukup bagi perkembangan anakan. Penelitian
Storz et al. (2000) pada sarang kelelawar Cyanopterus sphinx (Megachiroptera) di
India Barat mendapatkan tanaman palem (Caryota urens : Palmaea) ditempati oleh 1 individu jantan dewasa, 37 individu betina dewasa, dan 33 individu anakan. Penelitian Soegiharto & Kartono (2009) mendapatkan kelelawar Megachiroptera:
Pteropus vampirus menempati tanaman kelapa (Cocos nucifera: Palmaea), kepuh
21
Kebun Raya Bogor. Tanaman yang dipilih memiliki ketinggian yang cukup untuk menghindar dari gangguan predator serta bertajuk relatif lebar dan mendatar.
Jenis Megachiroptera yang bersarang di gua biasanya dalam koloni kecil atau bahkan hanya satu individu saja. Jenis-jenis tersebut adalah Rousettus
amplexicaudatus, Megaderma lyra dan Eonysteris spelaea (Suyanto 2001).
Penelitian Doyle (1979) di Gua Pondok Malaysia mendapatkan Eonycteris
spelaea dengan jumlah dua puluh individu dan Megaderma lyra hanya lima
individu bersarang dalam gua.
Sebaliknya, ordo Microchiroptera bersarang di pohon dalam jumlah sedikit. Microchiroptera lebih menyukai bersarang di bangunan buatan manusia, di celah batuan atau di gua dibandingkan pada dahan pohon. Penelitian Campbell et al. (1996) di hutan Pasific Nortwest Amerika Serikat mendapatkan kelelawar
Lasionycteri noctivagans (Vespertilionidae: Microchiroptera) bersarang pada
pohon pinus (Pinus ponderosa: Pinaceae) dan pinus putih (Pinus monticola: Pinaceae). Law & Chidel (2002) meneliti sarang dan ekologi pencarian makan kelelawar Kerivoula papuensis (Vespertilionidae : Microchiroptera) di hutan hujan New South Wales Australia. Sebanyak 11 individu kelelawar ditangkap di sekitar hutan dan dipasangi radiotracking. Lima puluh empat persen (54%) di antaranya bersarang di pohon yang jaraknya 5.2 km dari sungai, dan dua puluh tiga persen (23%) bersarang di pohon yang jaraknya 2.7 km dari sungai, dua puluh tiga persen (23%) bersarang dipohon yang jaraknya 2 km. Jumlah individu dalam koloni sarang ternyata tidak lebih dari 10 individu. Tanaman yang digunakan sebagai sarang adalah pohon jeruk (Flindersia australis: Rutaceae). Russo et al. (2003) meneliti seleksi sarang oleh kelelawar jenis Barbastella barbastellus (Vespertilionidae: Microchiroptera) di hutan Italia. Tanaman pada hutan yang tidak ditebang lebih banyak dihuni kelelawar Barbastella barbastellus daripada di hutan yang telah mengalami penebangan. Hal ini karena di hutan yang belum ditebang lebih banyak terdapat tanaman tua (hampir mati) dengan kulit kayu mengelupas, tinggi, dan sedikit daun.
Kebanyakan jenis Microchiroptera bersarang di gua dalam jumlah besar. Menurut Altringham (1996) dan Zahn & Hager (2005) beberapa jenis kelelawar memilih gua sebagai tempat bersarang karena kondisi gua yang lembap, suhu
22
stabil, dan jauh dari kebisingan. Dengan kondisi demikian, kelelawar kelompok Microchiroptera dapat meminimalkan kekurangan air akibat evaporasi, dapat memilih suhu yang tepat untuk tubuhnya, dan dapat menghindari kebisingan yang dapat mengganggu bahkan dapat menyebabkan kematian. Menurut Ruczynsi et al. (2007) kelelawar Microchiroptera memiliki alat pendengaran yang sangat sensitif pada gelombang suara, terutama gelombang pantul (echolokasi) berfrekuensi ultrasonik ( > 20 KHerzt).
Hasil penelitian Seckerdieck et al. (2005) membuktikan bahwa koloni kelelawar betina Rhinolopus hipposideros (Rhinolophidae : Microchiroptera) pada masa produktif cenderung memilih ruang bawah tanah dengan suhu rata rata 2oC lebih dingin dan lebih stabil dibandingkan lokasi lainnya. Pada masa
hamil dan menyusui, sebenarnya kelelawar lebih membutuhkan suhu hangat. Meskipun lebih dingin, ruang bawah tanah lebih dipilih sebagai tempat bersarang karena mempunyai suhu stabil. Penelitian Zahn & Hager (2005) juga menunjukkan bahwa kelelawar jenis Myotis daubentonii (Vespertilionidae: Microchiroptera) ditemukan bereproduksi di gua-gua di Eropa Tengah yang juga dijadikan tempat bersarang bagi kelelawar muda dan jantan dewasa. M.
daubentonii jantan biasanya menempati lokasi yang lebih dingin dibandingkan M. daubentonii betina.
Apriandi et al. (2008) melakukan penelitian pada kelelawar penghuni gua di Karst Cibinong. Hasil penelitan menunjukkan dalam satu gua ditemukan dua koloni kelelawar Hipposideros larvatus (Hipposideridae: Microchiroptera) bersarang di satu gua yang sama. Tiap jenis kelelawar memilih sarang dalam gua dengan jarak dari mulut gua berbeda. Hasil penelitian Maryanto & Maharadatunkamsi (1991) pada gua-gua di Pulau Sumbawa mendapatkan jenis
Rhinolophus luctus (Rhinolopodidae: Microchiroptera) menyukai tempat
bersarang di ujung gua. Dunn (1978) mendapatkan Hipposideros diadema (Hipposideridae: Microchiroptera) dan H.armiger di atap gua pada jarak 200 kaki dari mulut Gua Anak Takun Malaysia. Dengan memilih sarang jauh dari mulut gua, kelelawar dapat terhindar dari gangguan manusia dan predator serta dapat memilih mikroklimat yang stabil dan sesuai bagi tubuhnya. Tetapi, pemilihan sarang dengan jarak jauh dari mulut gua harus didukung oleh kemampuan
23
orientasi ruang dalam keadaan gelap dan kemampuan terbang dalam ruang dengan banyak rintangan. Penelitian Safi & Kerth (2004) pada 35 jenis kelelawar Microchiroptera di zona temperate Eropa dan Amerika Utara menunjukkan bahwa kelelawar yang mempunyai tulang-tulang jari (phalanges) sayap panjang hanya mampu mengeksploitasi habitat dengan kanopi terbuka. Sebaliknya kelelawar yang memiliki tulang jari sayap pendek, lebih mampu mengeksploitasi habitat berkelok-kelok dan banyak rintangan. Oleh karena itu, kelelawar yang mampu bersarang pada lokasi jauh dari mulut gua kemungkinan adalah kelelawar dengan tulang jari sayap pendek.
Adaptasi struktural dan fisiologi pernapasan
Kondisi ruang gua yang sempit, sirkulasi udara terbatas, dan banyak dihuni kelelawar menyebabkan udara dalam gua menjadi rendah oksigen (hypoxic), tinggi karbon dioksida (hypercapnic), dan tinggi gas amonia (Baudinette et al. 1994). Keadaan ini kurang menguntungkan bagi hewan gua karena: 1) kurangnya oksigen dapat menyebabkan respirasi terhambat (Guyton 1995); 2) tingginya karbon dioksida dapat menyebabkan afinitas hemoglobin pada oksigen menurun (Guyton 1995); dan 3) tingginya konsentrasi gas amonia (NH3) dapat
menyebabkan gangguan metabolisme, iritasi epitel organ pernapasan serta gangguan fisiologi saraf (Hutabarat et al. 2000). Oleh karena itu, agar dapat bertahan hidup dalam gua, kelelawar harus beradaptasi pada keadaan tersebut. Hal ini sesuai dengan pendapat Alikodra (2010) bahwa untuk bertahan hidup dan berkembang dalam suatu habitat, hewan harus mengembangkan strategi, di antaranya strategi adaptasi pada habitat.
Penelitian Baudinette et al. (1994) di Afrika Selatan membuktikan bahwa kelelawar Miniopterus schreibersii (Vespertilionidae: Microchiroptera) dapat hidup pada gua dengan kondisi udara rendah oksigen (hypoxic) dan tinggi karbon dioksida (hypercapnic). Jumlah populasi kelelawar yang besar pada gua tersebut menyebabkan oksigen yang digunakan untuk respirasi lebih besar dibandingkan oksigen yang masuk ke dalam gua. Sebaliknya karbon diokasida yang dihasilkan dari proses respirasi menambah jumlah karbon dioksida yang terperangkap di dalam gua (Baudinette et al. 1994).
24
Menurut Suyanto (2001), kelelawar juga dapat bertahan hidup pada gua dengan kandungan amonia tinggi. Penelitian Sridhar et al. (2006) mendapatkan urin dan feses (guano) kelelawar Hipposideros speoris (Hipposideridae: Microchiroptera) tersusun atas 5.7 ± 1.5% nitrogen (N) berbentuk amonia (NH3).
Amonia tersebut merupakan hasil katabolisme protein. Amonia dalam guano dapat menguap menjadi gas bercampur dengan komponen udara lainnya. Hal ini menyebabkan kandungan amonia udara meningkat tajam (Shidar et al. 2006). Hutabarat (2000) melakukan penelitian pada karyawan pabrik lateks yang terkena paparan amonia sebesar 500 ppm sampai 600 ppm selama 60 hari. Hasil penelitian menunjukkan karyawan yang terkena paparan amonia mengalami gejala sebagai berikut : tenggorokan kering (80%); jalan pernapasan kering (73.3%); mata perih (66.67%); batuk (53.3%); dan pingsan (6.67%). Menurut Suyanto (2001) kelelawar gua dapat bertahan pada kandungan amonia udara mencapai 5000 ppm, sedangkan manusia hanya mampu bertahan pada kandungan amonia udara maksimum sebesar 100 ppm.
Penelitian tentang strategi adaptasi kelelawar yang bersarang di gua dengan kondisi dingin dan lembap pernah dilakukan oleh Baudinette et al. (2000) di Australia. Hasil penelitian menunjukkan laju respirasi kelelawar Macroderma
gigas (Megadermatidae: Microchiroptera) dan Rhinonycteris aurantias
(Hipposideridae: Microchiroptera) menyesuaikan dengan suhu dan kelembapan udara dalam gua. Pada saat kondisi udara kering dan dingin (kelembapan <60% dan suhu < 5.6oC) laju respirasi sama dengan pada saat kondisi udara lembap dan hangat ( kelembapan > 80% ;suhu > 9.8oC). Tetapi bila kondisi udara lembap dan dingin (kelembapan < 60% dan suhu < 9o) laju respirasinya meningkat tajam. Meningkatnya laju respirasi tersebut merupakan strategi agar tubuh tetap hangat. Namun demikian, sejauh ini belum ada penelitian mengenai strategi adaptasi fisiologi dan anatomi pernapasan kelelawar untuk bertahan hidup pada kondisi hypoxic, hypercapnic, dan tinggi amonia.
Menurut Frandson (1992) organ respirasi Mamalia terdiri atas: 1) lubang hidung (nasale), berfungsi menyaring, menghangatkan, dan melembapkan udara yang masuk ke dalam organ pernapasan; 2) laring, berfungsi melindungi jalan napas bawah dari obstruksi (masuknya) benda asing; 3) trakhea, berfungsi
25
menghantarkan udara ke paru-paru; 4) bronchus, merupakan percabangan trakhea; dan 5) alveolus, merupakan membran tempat pertukaran oksigen dan karbon dioksida. Organ pernapasan hewan Mamalia dapat dilihat pada Gambar 8.
Gambar 8 Organ pernapasan Mamalia (Sumber: Campbell et al. 2002) Bagian terpenting dari organ pernapasan adalah alveolus, yaitu tempat oksigen berdifusi ke dalam sel dan karbon dioksida berdifusi keluar sel. Menurut Guyton (1995), dinding alveolus sangat tipis dan di dalamnya banyak terdapat kapiler darah yang saling berhubungan. Kondisi demikian memudahkan pertukaran gas antara membran alveolus dan membran kapiler darah. Difusi oksigen dari alveolus ke dalam kapiler darah dan difusi karbon dioksida dengan arah berlawanan dapat dilihat pada Gambar 9.
Gambar 9 Alveolus dan kapiler darah tempat difusi oksigen dan karbon dioksida (Sumber: Campbell et al. 2002)
26
Menurut Guyton (1995) pertukaran gas respirasi melewati membran alveolus berlangsung sangat efektif. Hal ini karena permukaan membran alveolus sangat tipis. Selain itu, dengan bentuk seperti gelembung-gelembung udara, luas permukaan membran menjadi sangat besar. Hasil penelitian Setiadi (2000) menunjukkan kelelawar Scotophilus kuhli (Vespertilionidae: Microchiroptera) memiliki alveolus lebih kecil dan rapat dibandingkan alveolus tikus (Mus
musculus). Menurut Plopper & Adams (1993), alveolus beberapa Mamalia
berukuran kecil dan rapat sehingga permukaan respirasinya lebih luas. Permukaan respirasi yang luas menyebabkan paru-paru mampu mengikat oksigen lebih banyak (Plopper & Adams 1993). Menurut Guyton (1995), luas permukaan membran alveolus ini dapat berkurang karena rusaknya dinding alveolus. Kerusakan tersebut di antaranya adalah karena emfisema, yaitu keadaan alveolus bersatu disebabkan terpapar gas racun atau karena infeksi kuman.
Setelah oksigen berdifusi dari alveolus ke kapiler darah dan karbon dioksida berdifusi dari kapiler darah ke alveolus, selanjutnya oksigen akan diangkut oleh darah ke seluruh jaringan tubuh oleh sel darah merah (eritrosit). Karena adanya unsur hemoglobin dalam eritrosit, maka eritrosit dapat mengikat oksigen dan karbon dioksida. Menurut Ganong (2001), eritrosit Mamalia berbentuk cakram bikonkaf. Cakram bikonkaf tersebut memiliki permukaan yang relatif luas untuk pertukaran oksigen.
Menurut Ganong (2001) jumlah eritrosit juga berbeda antara satu individu dengan individu lainnya. Faktor yang mempengaruhi jumlah eritrosit hewan adalah kondisi nutrisi, aktivitas fisik, umur, dan lingkungan. Selain itu, menurut Ganong (2001), penurunan kadar oksigen (hipoksia) akan merangsang ginjal untuk melepaskan enzim eritrogenin (erythrogenin). Selanjutnya, enzim tersebut akan mengaktifkan eritropoietinogen yang merupakan prekusor dalam pembentukan eritopoietin. Di dalam sumsum tulang, eritropoietin akan meningkatkan jumlah sel bakal (stem cell). Sel bakal ini akan menjadi prekusor darah merah dan selanjutnya menjadi sel darah merah (Ganong 2001). Menurut Guyton (1995), binatang yang terpapar oksigen rendah selama bertahun tahun menunjukkan peningkatan jaringan hipoksik dan peningkatan jumlah sel darah merah sampai 20%.
27
Kemampuan sel darah merah mengikat oksigen disebabkan adanya
hemoglobin. Hemoglobin merupakan molekul protein yang berikatan dengan porphyrin. Di bagian tengah molekul porphyrin tersebut terdapat satu atom besi
(Fe). Menurut Ganong (2001) hemoglobin Mamalia tersusun atas empat subunit protein berbentuk globul (bola). Satu subunit dapat membawa satu molekul oksigen, dengan demikian setiap molekul hemoglobin dapat membawa empat molekul oksigen. Struktur hemoglobin dapat dilihat pada Gambar 10.
Gambar 10 Struktur kimia hemoglobin (Sumber: Campbell et al. 2002) Menurut Ganong (2001) hemoglobin yang mengikat oksigen disebut
oksihemoglobin. Afinitas (daya ikat) hemoglobin pada oksigen dipengaruhi oleh
suhu tubuh, keasaman (pH) darah, dan konsentrasi bahan-bahan kimia dalam darah. Pada keadaan suhu udara rendah, tekanan oksigen tinggi dan keasaman darah tinggi, afinitas hemoglobin pada oksigen meningkat. Sebaliknya pada kondisi tekanan oksigen rendah, suhu tinggi, dan keasaman rendah afinitas hemoglobin pada oksigen menurun (Ganong 2001). Menurut Guyton (1995), dalam keadaan normal, 100 ml darah manusia mengandung 15 gram hemoglobin yang mampu mengangkut 0.03 gram oksigen. Penelitian Fatmawati (2007), mendapatkan kandungan hemoglobin anjing (Canis familiaris) 11.8 ± 1.13 g/mm3.
Menurut Frandson (1992) jumlah hemoglobin hewan Mamalia berada pada kisaran 12 – 18 g/mm3.
28
Kesamaan relung (niche overlap) pakan
Kelelawar dari jenis (species) berbeda dapat memanfaatkan gua yang sama sebagai sarang. Hal ini dibuktikan oleh: penelitian Dunn (1978) yang mendapatkan jenis Hipposideros armiger, H. cineraceus, Rhinolophus affinis dan
R. malayanus di Gua Anak Takun Malaysia; penelitian Zukal et al. (2005) yang
mendapatkan jenis Myotis myotis dan Rhinolophus hipposideros di Gua Katerinska Cekoslvakia; dan penelitian Apriandi et al. (2008) yang mendapatkan jenis Miniopterus australis, Myotis adversus dan Rhinolophus affinis di Gua Gudawang Bogor. Menurut Kunz (1982) dan Willis & Brigmann (2004) sarang yang dipilih kelelawar memiliki akses yang mudah pada sumber pakan. Oleh karena itu, apabila jenis-jenis kelelawar yang bersarang dalam satu gua tersebut bergantung pada sumber pakan yang sama, akan terjadi kompetisi, terutama bila ketersediaan sumber pakan terbatas. Sebaliknya, bila sumber pakan berbeda, kompetisi tidak terjadi.
Menurut Cox (2002), penggunaan relung yang sama (niche overlap) menyebabkan interaksi kompetitif, yaitu tiap populasi yang berkompetisi memberikan pengaruh yang merugikan bagi pesaingnya (competitor). Menurut Reynold & Ludwig (1984) nilai niche overlap berkisar antara nol (0) sampai dengan satu (1). Apabila nilai niche overlap pakan mendekati satu berarti kedua jenis hewan tersebut memiliki pakan yang sama dan berpotensi untuk berkompetisi.
Kelelawar anggota subordo Megachiroptera adalah pemakan buah (frugivora) ataupun serbuk sari (polinator), sedangkan anggota subordo Microchiroptera kebanyakan pemakan serangga (insektivora). Menurut Altringham (1996), berdasarkan strategi pencarian makannya, kelelawar dibedakan menjadi tipe spesialis (selektif) dan opportunis (generalis). Kelelawar tipe spesialis hanya memakan jenis tertentu. Tipe ini bisa menghabiskan banyak waktu dan energi dalam pencarian makan, tetapi makanan yang didapatkan memiliki profit (nilai gizi) tinggi. Tipe opportunis menghabiskan lebih sedikit waktu dan energi dalam pencarian makannya, tetapi makanan yang didapatkan mungkin lebih sedikit nilai gizinya dibandingkan kelelawar tipe spesialis. Beberapa Megachiroptera, misalnya Rousettus amplexicaudatus (Pteropodidae:
29
Makrochiroptera) adalah pemakan buah tipe oportunis, sedangkan Macroglosus
sabrinus (Pteropodidae: Makrochiroptera) adalah tipe spesialis. Menurut Nowak
(1994) R. amplexicaudatus memakan jambu biji (Psidium guajava), pisang (Musa
paradisiaca); sawo (Manilkara kauki), dan buah masak lainnya. Sedangkan Macroglosus sabrinus terspesialissasi untuk memakan nectar bunga durian
(Durio zibethinus) dan bunga patai (Parkis speciosa) (Nowak 1994). Microchiroptera adalah pemakan serangga dengan tipe spesialis ataupun oportunis. Hal ini dibuktikan dengan hasil penelitian Razakarivony et al. (2005) yang meneliti makanan kelelawar jenis Myzopoda aurita (Myzopodidae: Microchiroptera) di Madagaskar. Pada saat ngengat (Lepidoptera) melimpah, prosentase ngengat dalam feses juga meningkat. Hasil penelitian ini membuktikan bahwa kelelawar ini memanfaatkan kesempatan (oportunis) dalam memilih jenis makanannya.